Oviduct Transcriptomic Reveals the Regulation of mRNAs and lncRNAs Related to Goat Prolificacy in the Luteal Phase

 ,
,  ,
,  , and
, and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Experimental Animals and Sampling
2.3. Total RNA Extraction and Detection
2.4. Library Preparation, RNA-seq, and Data Quality Control
2.5. LncRNA Identification and Differential Expression Analysis
2.6. Quantitative Real-Time PCR
2.7. Target Gene Prediction of DELs and Functional Bioinformatics Analyses
2.8. Statistical Analysis
3. Results
3.1. Summary of Sequencing Data in YunShang Black Goat Oviducts in the Luteal Phase
3.2. Identification of mRNA and lncRNA in YunShang Black Goat Oviduct Tissue
3.3. Profiling of Differentially Expressed Transcripts and qPCR Verification
3.4. PPI Network Related to Prolificacy Traits
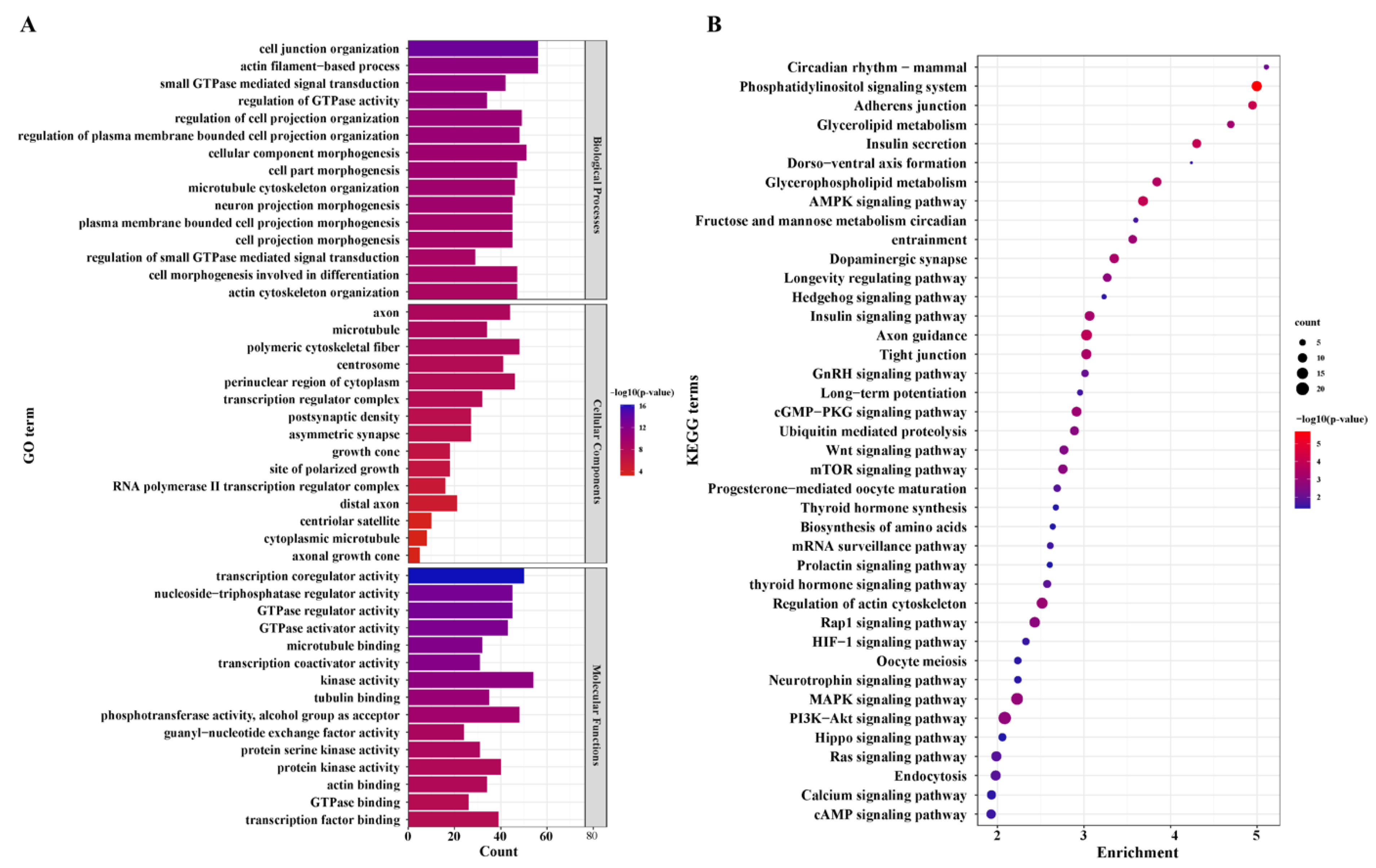
3.5. Functional Enrichment Analysis of the DEGs
3.6. Posttranscriptional Regulatory Network of DELs and the Target Genes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Liu, Y.F.; Zhou, Z.Y.; He, X.Y.; Tao, L.; Jiang, Y.T.; Lan, R.; Hong, Q.H.; Chu, M.X. Integrated analyses of miRNA-mRNA expression profiles of ovaries reveal the crucial interaction networks that regulate the prolificacy of goats in the follicular phase. BMC Genomics. 2021, 22, 812. [Google Scholar] [CrossRef] [PubMed]
- Buhi, W.C.; Alvarez, I.M.; Kouba, A.J. Secreted proteins of the oviduct. Cells Tissues Organs 2000, 166, 165–179. [Google Scholar] [CrossRef] [PubMed]
- Shirley, B.; Reeder, R.L. Cyclic changes in the ampulla of the rat oviduct. J. Exp. Zool. 1996, 276, 164–173. [Google Scholar] [CrossRef]
- Avilés, M.; Gutierrez-Adan, A.; Coy, P. Oviductal secretions: Will they be key factors for the future ARTs? Mol. Hum. Reprod. 2010, 16, 896–906. [Google Scholar] [CrossRef] [PubMed]
- Hunter, R. Components of oviduct physiology in eutherian mammals. Biol. Rev. 2011, 87, 244–255. [Google Scholar] [CrossRef]
- Coy, P.; Yanagimachi, R. The common and species-specific roles of oviductal proteins in mammalian fertilization and embryo development. BioScience 2015, 65, 973–984. [Google Scholar] [CrossRef] [Green Version]
- Chang, H.M.; Qiao, J.; Leung, P.C.K. Oocyte-somatic cell interactions in the human ovary-novel role of bone morphogenetic proteins and growth differentiation factors. Hum. Reprod. Update 2016, 23, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Miller, W.L.; Auchus, R.J. The molecular biology, biochemistry, and physiology of human steroidogenesis and its disorders. Endocr. Rev. 2011, 32, 81–151. [Google Scholar] [CrossRef] [Green Version]
- Cui, H.; Zhao, S.; Cheng, M.; Guo, L.; Ye, R.; Liu, W.; Gao, S. Cloning and Expression Levels of Genes Relating to the Ovulation Rate of the Yunling Black Goat1. Biol. Reprod. 2009, 80, 219–226. [Google Scholar] [CrossRef] [Green Version]
- Davis, G.H. Major genes affecting ovulation rate in sheep. Genet. Sel. Evol. 2005, 37, S11–S14. [Google Scholar] [CrossRef]
- Hu, W.; Tang, J.; Zhang, Z.; Tang, Q.; Yan, Y.; Wang, P.; Wang, X.; Liu, Q.; Guo, X.; Jin, M.; et al. Polymorphisms in the ASMT and ADAMTS1 gene may increase litter size in goats. Veter-Med. Sci. 2020, 6, 775–787. [Google Scholar] [CrossRef] [PubMed]
- Chu, M.X.; Jiao, C.L.; He, Y.Q.; Wang, J.Y.; Liu, Z.H.; Chen, G.H. Association between PCR-SSCP of bone morphogenetic protein 15 gene and prolificacy in Jining Grey goats. Anim. Biotechnol. 2007, 18, 263–274. [Google Scholar] [CrossRef] [PubMed]
- Pan, Z.; Zhang, J.; Lin, F.; Ma, X.; Wang, X.; Liu, H. Expression profiles of key candidate genes involved in steroidogenesis during follicular atresia in the pig ovary. Mol. Biol. Rep. 2012, 39, 10823–10832. [Google Scholar] [CrossRef] [PubMed]
- Pramod, R.; Sharma, S.; Singhi, A.; Pan, S.; Mitra, A. Differential Ovarian Morphometry and Follicular Expression of BMP15, GDF9 and BMPR1B Influence the Prolificacy in Goat. Reprod. Domest. Anim. 2013, 48, 803–809. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.; Hong, Q.; Liu, Y.; He, X.; Di, R.; Wang, X.; Ren, C.; Zhang, Z.; Chu, M. Characterization of circular RNA profiles of oviduct reveal the potential mechanism in prolificacy trait of goat in the estrus cycle. Front. Physiol. 2022, 13, 990691. [Google Scholar] [CrossRef]
- Liu, L.; Fang, F. Long noncoding RNA mediated regulation in human embryogenesis, pluripotency, and reproduction. Stem Cells Int. 2022, 2022, 8051717. [Google Scholar] [CrossRef]
- Wang, J.J.; Niu, M.H.; Zhang, T.; Shen, W.; Cao, H.G. Genome-wide network of lncRNA–mRNA during ovine oocyte development from germinal vesicle to metaphase II in vitro. Front. Physiol. 2020, 11, 1019. [Google Scholar] [CrossRef]
- Taylor, D.; Chu, E.T.-J.; Spektor, R.; Soloway, P.D. Long non-coding RNA regulation of reproduction and development. Mol. Reprod. Dev. 2015, 82, 932–956. [Google Scholar] [CrossRef] [Green Version]
- Lai, F.; Orom, U.A.; Cesaroni, M.; Beringer, M.; Taatjes, D.J.; Blobel, G.A.; Shiekhattar, R. Activating RNAs associate with Mediator to enhance chromatin architecture and transcription. Nature 2013, 494, 497–501. [Google Scholar] [CrossRef] [Green Version]
- Beltran, M.; Puig, I.; Peña, C.; García, J.M.; Álvarez, A.B.; Peña, R.; Bonilla, F.; de Herreros, A.G. A natural antisense transcript regulates Zeb2/Sip1 gene expression during Snail1-induced epithelial–mesenchymal transition. Genes Dev. 2008, 22, 756–769. [Google Scholar] [CrossRef]
- Caretti, G.; Schiltz, R.L.; Dilworth, F.J.; Di Padova, M.; Zhao, P.; Ogryzko, V.; Fuller-Pace, F.V.; Hoffman, E.P.; Tapscott, S.J.; Sartorelli, V. The RNA helicases p68/p72 and the noncoding RNA SRA are coregulators of MyoD and skeletal muscle differentiation. Dev. Cell 2006, 11, 547–560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohammad, F.; Mondal, T.; Guseva, N.; Pandey, G.K.; Kanduri, C. Kcnq1ot1 noncoding RNA mediates transcriptional gene silencing by interacting with Dnmt1. Development 2010, 137, 2493–2499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, G.L.; Liu, S.X.; Xing, G.L.; Wang, F. LncRNA PVT1/microRNA-17-5p/PTEN axis regulates secretion of E2 and P4, proliferation, and apoptosis of ovarian granulosa cells in PCOS. Mol. Ther. Nucleic. Acids 2020, 20, 205–216. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Guo, Y.X.; Duan, C.H.; Yang, R.C.; Zhang, L.C.; Liu, Y.Q.; Zhang, Y.J. Long non-coding RNA GDAR regulates ovine granulosa cells apoptosis by affecting the expression of apoptosis-related genes. Int. J. Mol. Sci. 2022, 23, 5183. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Hu, S.G.; Yao, G.X.; Zhu, Q.L.; He, Y.Q.; Lu, Y.; Qi, J.; Xu, R.; Ding, Y.; Li, J.X.; et al. A novel molecule in human cyclic endometrium: Lncrna tunar is involved in embryo implantation. Front. Physiol. 2020, 11, 587448. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Cao, Y.X.; Xu, X.F.; Xiang, H.F.; Zhang, Z.G.; Chen, B.L.; Hao, Y.; Wei, Z.L.; Zhou, P.; Chen, D.W. Increased new lncRNA-mRNA gene pair levels in human cumulus cells correlate with oocyte maturation and embryo development. Reprod. Sci. 2015, 22, 1008–1014. [Google Scholar] [CrossRef]
- Zhao, Z.F.; Zou, X.; Lu, T.T.; Deng, M.; Li, Y.K.; Guo, Y.Q.; Sun, B.L.; Liu, G.B.; Liu, D.W. Identification of mRNAs and lncRNAs involved in the regulation of follicle development in goat. Front. Genet. 2020, 11, 589076. [Google Scholar] [CrossRef]
- Liu, Y.; Qi, B.; Xie, J.; Wu, X.Q.; Ling, Y.H.; Cao, X.Y.; Kong, F.; Xin, J.; Jiang, X.; Wu, Q.Q.; et al. Filtered reproductive long non-coding RNAs by genome-wide analyses of goat ovary at different estrus periods. BMC Genome. 2018, 19, 866. [Google Scholar] [CrossRef]
- Hong, L.J.; Hu, Q.; Zang, X.P.; Xie, Y.S.; Zhou, C.; Zou, X.; Li, Y.K.; Deng, M.; Guo, Y.Q.; Liu, G.B.; et al. Analysis and screening of reproductive long non-coding RNAs through genome-wide analyses of goat endometrium during the pre-attachment phase. Front. Genet. 2020, 11, 568017. [Google Scholar] [CrossRef]
- Li, S.; Winuthayanon, W. Oviduct: Roles in fertilization and early embryo development. J. Endocrinol. 2017, 232, R1–R26. [Google Scholar] [CrossRef]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kovaka, S.; Zimin, A.V.; Pertea, G.M.; Razaghi, R.; Salzberg, S.L.; Pertea, M. Transcriptome assembly from long-read RNA-seq alignments with StringTie2. Genome Biol. 2019, 20, 278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trapnell, C.; Williams, B.A.; Pertea, G.; Mortazavi, A.; Kwan, G.; Van Baren, M.J.; Salzberg, S.L.; Wold, B.J.; Pachter, L. Transcript assembly and quantification by RNA-Seq reveals unannotated transcripts and isoform switching during cell differentiation. Nat. Biotechnol. 2010, 28, 511–515. [Google Scholar] [CrossRef] [Green Version]
- Sun, L.; Luo, H.T.; Bu, D.C.; Zhao, G.G.; Yu, K.T.; Zhang, C.H.; Liu, Y.N.; Chen, R.S.; Zhao, Y. Utilizing sequence intrinsic composition to classify protein-coding and long non-coding transcripts. Nucleic Acids Res. 2013, 41, e166. [Google Scholar] [CrossRef] [PubMed]
- Kang, Y.J.; Yang, D.C.; Kong, L.; Hou, M.; Meng, Y.Q.; Wei, L.; Gao, G. CPC2: A fast and accurate coding potential calculator based on sequence intrinsic features. Nucleic Acids Res. 2017, 45, W12–W16. [Google Scholar] [CrossRef] [Green Version]
- Li, A.M.; Zhang, J.Y.; Zhou, Z.Y. PLEK: A tool for predicting long non-coding RNAs and messenger RNAs based on an improved k-mer scheme. BMC Bioinform. 2014, 15, 311. [Google Scholar] [CrossRef] [Green Version]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [Green Version]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative C(T) method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef]
- Akison, L.K.; Robker, R.L. The critical roles of progesterone receptor (PGR) in ovulation, oocyte developmental competence and oviductal transport in mammalian reproduction. Reprod. Domest. Anim. 2012, 47, 288–296. [Google Scholar] [CrossRef]
- Cerny, K.L.; Garrett, E.; Walton, A.J.; Anderson, L.H.; Bridges, P.J. A transcriptomal analysis of bovine oviductal epithelial cells collected during the follicular phase versus the luteal phase of the estrous cycle. Reprod. Biol. Endocrinol. 2015, 13, 84. [Google Scholar] [CrossRef]
- Kornienko, A.E.; Dotter, C.P.; Guenzl, P.M.; Gisslinger, H.; Gisslinger, B.; Cleary, C.; Kralovics, R.; Pauler, F.M.; Barlow, D.P. Long non-coding RNAs display higher natural expression variation than protein-coding genes in healthy humans. Genome Biol. 2016, 17, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goodman, R.L.; Maltby, M.J.; Millar, R.P.; Hileman, S.M.; Nestor, C.C.; Whited, B.; Tseng, A.S.; Coolen, L.M.; Lehman, M.N. Evidence that dopamine acts via kisspeptin to hold GnRH pulse frequency in check in anestrous ewes. Endocrinology 2012, 153, 5918–5927. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nestor, C.C.; Bedenbaugh, M.N.; Hileman, S.M.; Coolen, L.M.; Lehman, M.N.; Goodman, R.L. Regulation of GnRH pulsatility in ewes. Reproduction 2018, 156, R83–R99. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.B.; Tang, J.S.; Di, R.; Liu, Q.Y.; Wang, X.Y.; Gan, S.Q.; Zhang, X.S.; Zhang, J.L.; Hu, W.P.; Chu, M.X. Comparative transcriptomics reveal key sheep (ovis aries) hypothalamus lncRNAs that affect reproduction. Animals 2019, 9, 152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herrera, G.G.B.; Lierz, S.L.; Harris, E.A.; Donoghue, L.J.; Hewitt, S.C.; Rodriguez, K.F.; Jefferson, W.N.; Lydon, J.P.; DeMayo, F.J.; Williams, C.J.; et al. Oviductal retention of embryos in female mice lacking estrogen receptor α in the isthmus and the uterus. Endocrinology 2020, 161, bqz033. [Google Scholar] [CrossRef] [PubMed]
- Cao, W.; Chang, T.; Li, X.-Q.; Wang, R.; Wu, L. Dual effects of fructose on ChREBP and FoxO1/3α are responsible for AldoB up-regulation and vascular remodelling. Clin. Sci. 2017, 131, 309–325. [Google Scholar] [CrossRef]
- Trenti, A.; Tedesco, S.; Boscaro, C.; Ferri, N.; Cignarella, A.; Trevisi, L.; Bolego, C. The Glycolytic Enzyme PFKFB3 Is Involved in Estrogen-Mediated Angiogenesis via GPER1. J. Pharmacol. Exp. Ther. 2017, 361, 398–407. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Zhong, Y.Q.; Liu, W.F.; Zhang, F.H.; Zhao, Y.; Zou, L.; Liu, X.X. PFKFB3-mediated glycometabolism reprogramming modulates endothelial differentiation and angiogenic capacity of placenta-derived mesenchymal stem cells. Stem Cell Res. Ther. 2022, 13, 391. [Google Scholar] [CrossRef]
- Liu, J.J.; Ran, X.Q.; Li, S.; Feng, Y.; Wang, J.F. Polymorphism in the first intron of follicle stimulating hormone beta gene in three Chinese pig breeds and two European pig breeds. Anim. Reprod. Sci. 2009, 111, 369–375. [Google Scholar] [CrossRef]
- Tríbulo, P.; Jumatayeva, G.; Lehloenya, K.; Moss, J.I.; Negrón-Pérez, V.M.; Hansen, P.J. Effects of sex on response of the bovine preimplantation embryo to insulin-like growth factor 1, activin A, and WNT7A. BMC Dev. Biol. 2018, 18, 16. [Google Scholar] [CrossRef]
- Trueb, B. Biology of FGFRL1, the fifth fibroblast growth factor receptor. Cell Mol. Life Sci. 2011, 68, 951–964. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshimura, T.; Yasuo, S.; Watanabe, M.; Iigo, M.; Yamamura, T.; Hirunagi, K.; Ebihara, S. Light-induced hormone conversion of T4 to T3 regulates photoperiodic response of gonads in birds. Nature 2003, 426, 178–181. [Google Scholar] [CrossRef]
- Anastasaki, C.; Longman, D.; Capper, A.; Patton, E.E.; Cáceres, J.F. Dhx34 and Nbas function in the NMD pathway and are required for embryonic development in zebrafish. Nucleic Acids Res. 2011, 39, 3686–3694. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, S.; Tan, Q.; Liang, J.; Cao, D.; Wang, S.; Liang, J.; Chen, K.; Wang, Z. Placental trophoblast cell-derived exosomal microRNA-1290 promotes the interaction between endometrium and embryo by targeting LHX6. Mol. Ther.—Nucleic Acids 2021, 26, 760–772. [Google Scholar] [CrossRef]
- Thompson, I.R.; Ciccone, N.A.; Xu, S.; Zaytseva, S.; Carroll, R.S.; Kaiser, U.B. GnRH pulse frequency-dependent stimulation of FSHβ transcription is mediated via activation of PKA and CREB. Mol. Endocrinol. 2013, 27, 606–618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Chen, Y.; Chen, Z.; Xiang, Z.; Ding, J.; Han, X. Microcystin-leucine arginine inhibits gonadotropin-releasing hormone synthesis in mice hypothalamus. Ecotoxicol. Environ. Saf. 2018, 163, 391–399. [Google Scholar] [CrossRef]
- Balschun, D.; Wolfer, D.P.; Gass, P.; Mantamadiotis, T.; Welzl, H.; Schütz, G.; Frey, J.U.; Lipp, H.P. Does cAMP response element-binding protein have a pivotal role in hippocampal synaptic plasticity and hippocampus-dependent memory? J. Neurosci. 2003, 23, 6304–6314. [Google Scholar] [CrossRef] [Green Version]
- Zhou, L.; Li, J.; Liu, J.; Wang, A.; Liu, Y.; Yu, H.; Ouyang, H.; Pang, D. Investigation of the lncRNA THOR in Mice Highlights the Importance of Noncoding RNAs in Mammalian Male Reproduction. Biomedicines 2021, 9, 859. [Google Scholar] [CrossRef]
- Wan, Z.; Yang, H.; Chen, P.; Wang, Z.; Cai, Y.; Yao, X.; Wang, F.; Zhang, Y. The Novel Competing Endogenous Long Noncoding RNA SM2 Regulates Gonadotropin Secretion in the Hu Sheep Anterior Pituitary by Targeting the Oar-miR-16b/TGF-β/SMAD2 Signaling Pathway. Cells 2022, 11, 985. [Google Scholar] [CrossRef]
- Hu, H.; Fu, Y.; Zhou, B.; Li, Z.; Liu, Z.; Jia, Q. Long non-coding RNA TCONS_00814106 regulates porcine granulosa cell proliferation and apoptosis by sponging miR-1343. Mol. Cell. Endocrinol. 2020, 520, 111064. [Google Scholar] [CrossRef]
- Savaris, R.F.; Da Silva, L.C.; Moraes, G.D.S.; Edelweiss, M.I.A. Expression of MUC1 in tubal pregnancy. Fertil. Steril. 2008, 89, 1015–1017. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Wang, N.; Zhang, C.; Li, M.; He, X.; Yin, C.; Tu, Q.; Shen, X.; Zhang, L.; Lv, J.; et al. Fructose-1,6-Bisphosphate Aldolase B Depletion Promotes Hepatocellular Carcinogenesis Through Activating Insulin Receptor Signaling and Lipogenesis. Hepatology 2021, 74, 3037–3055. [Google Scholar] [CrossRef] [PubMed]
- Caspi, M.; Perry, G.; Skalka, N.; Meisel, S.; Firsow, A.; Amit, M.; Rosin-Arbesfeld, R. Aldolase positively regulates of the canonical Wnt signaling pathway. Mol. Cancer 2014, 13, 164. [Google Scholar] [CrossRef] [Green Version]
- Nakabayashi, K.; Makino, S.; Minagawa, S.; Smith, A.C.; Bamforth, J.S.; Stanier, P.; Preece, M.; Parker-Katiraee, L.; Paton, T.; Oshimura, M.; et al. Genomic imprinting of PPP1R9A encoding neurabin I in skeletal muscle and extra-embryonic tissues. J. Med. Genet. 2004, 41, 601–608. [Google Scholar] [CrossRef] [Green Version]
- Zaitoun, I.; Khatib, H. Comparative genomic imprinting and expression analysis of six cattle genes1. J. Anim. Sci. 2008, 86, 25–32. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.; Millard, R.W.; Ashraf, M. Role of GATA-4 in differentiation and survival of bone marrow mesenchymal stem cells-sciencedirect. Prog. Mol. Biol. Transl. Sci. 2012, 111, 217–241. [Google Scholar] [CrossRef]
- Shelton, G.D.; Minor, K.M.; Thomovsky, S.; Guo, L.T.; Friedenberg, S.G.; Cullen, J.N.; Mickelson, J.R. Congenital muscular dystrophy in a dog with a LAMA2 gene deletion. J. Veter-Intern. Med. 2021, 36, 279–284. [Google Scholar] [CrossRef]
- Tao, L.; He, X.Y.; Jiang, Y.T.; Lan, R.; Li, M.; Li, Z.M.; Yang, W.F.; Hong, Q.H.; Chu, M.X. Combined approaches to reveal genes associated with litter size in Yunshang black goats. Anim. Genet. 2020, 51, 924–934. [Google Scholar] [CrossRef]
- Choi, K.-C.; Leung, P.C.; Jeung, E.-B. Biology and physiology of Calbindin-D9k in female reproductive tissues: Involvement of steroids and endocrine disruptors. Reprod. Biol. Endocrinol. 2005, 3, 66. [Google Scholar] [CrossRef] [Green Version]
- Luu, K.C.; Nie, G.Y.; Salamonsen, L.A. Endometrial calbindins are critical for embryo implantation: Evidence from in vivo use of morpholino antisense oligonucleotides. Proc. Natl. Acad. Sci. USA 2004, 101, 8028–8033. [Google Scholar] [CrossRef]
- Li, S.; O’Neill, S.R.S.; Zhang, Y.; Holtzman, M.J.; Takemaru, K.; Korach, K.S.; Winuthayanon, W. Estrogen receptor α is required for oviductal transport of embryos. FASEB J. 2016, 31, 1595–1607. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, R.R.; Ou, J.J.; Li, L.; Yang, Y.; Zhao, J.P.; Wu, R.R. The Wnt signaling pathway effector TCF7L2 mediates olanzapine-induced weight gain and insulin resistance. Front. Pharmacol. 2018, 9, 379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burcelin, R.; Thorens, B.; Glauser, M.; Gaillard, R.C.; Pralong, F.P. Gonadotropin-releasing hormone secretion from hypothalamic neurons: Stimulation by insulin and potentiation by leptin. Endocrinology 2003, 144, 4484–4491. [Google Scholar] [CrossRef]
- Downing, J.A.; Joss, J.; Scaramuzzi, R.J. A mixture of the branched chain amino acids leucine, isoleucine and valine increases ovulation rate in ewes when infused during the late luteal phase of the oestrous cycle: An effect that may be mediated by insulin. J. Endocrinol. 1995, 145, 315–323. [Google Scholar] [CrossRef] [PubMed]
- Salilew-Wondim, D.; Hölker, M.; Rings, F.; Phatsara, C.; Mohammadi-Sangcheshmeh, A.; Tholen, E.; Schellander, K.; Tesfaye, D. Depletion of BIRC6 leads to retarded bovine early embryonic development and blastocyst formation in vitro. Reprod. Fertil. Dev. 2010, 22, 564–579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ren, J.; Shi, M.; Liu, R.; Yang, Q.H.; Johnson, T.; Skarnes, W.C.; Du, C. The Birc6 (Bruce) gene regulates p53 and the mitochondrial pathway of apoptosis and is essential for mouse embryonic development. Proc. Natl. Acad. Sci. USA 2005, 102, 565–570. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Clean Reads | Clean Base (bp) | Length | Q20 (%) | Q30 (%) | GC (%) | Total Mapped (%) | Uniquely Mapped (%) |
|---|---|---|---|---|---|---|---|---|
| LH-1 | 53,564,145 | 16,069,243,500 | 150 | 98.10 | 94.45 | 46.70 | 97.11 | 93.06 |
| LH-2 | 53,967,718 | 16,190,315,400 | 150 | 96.85 | 91.90 | 41.85 | 95.55 | 93.61 |
| LH-3 | 53,642,740 | 16,092,822,000 | 150 | 98.55 | 95.40 | 43.10 | 97.20 | 94.62 |
| LH-4 | 70,077,091 | 21,023,127,300 | 150 | 97.65 | 93.40 | 48.95 | 96.46 | 91.29 |
| LH-5 | 53,146,812 | 15,944,043,600 | 150 | 98.10 | 94.35 | 46.05 | 97.12 | 93.33 |
| LL-1 | 57,893,047 | 17,367,914,100 | 150 | 98.40 | 94.95 | 48.30 | 97.24 | 92.03 |
| LL-2 | 64,129,075 | 19,238,722,500 | 150 | 98.40 | 95.10 | 46.70 | 97.30 | 93.89 |
| LL-3 | 66,815,604 | 20,044,681,200 | 150 | 98.30 | 94.75 | 46.70 | 97.20 | 93.20 |
| LL-4 | 53,717,853 | 16,115,355,900 | 150 | 98.20 | 94.60 | 47.10 | 97.23 | 92.67 |
| LL-5 | 60,845,824 | 18,253,747,200 | 150 | 98.40 | 95.10 | 43.90 | 97.35 | 94.48 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, Z.; Hong, Q.; Liu, Y.; Ren, C.; He, X.; Jiang, Y.; Ouyang, Y.; Chu, M.; Zhang, Z. Oviduct Transcriptomic Reveals the Regulation of mRNAs and lncRNAs Related to Goat Prolificacy in the Luteal Phase. Animals 2022, 12, 2823. https://doi.org/10.3390/ani12202823
Sun Z, Hong Q, Liu Y, Ren C, He X, Jiang Y, Ouyang Y, Chu M, Zhang Z. Oviduct Transcriptomic Reveals the Regulation of mRNAs and lncRNAs Related to Goat Prolificacy in the Luteal Phase. Animals. 2022; 12(20):2823. https://doi.org/10.3390/ani12202823
Chicago/Turabian StyleSun, Zhipeng, Qionghua Hong, Yufang Liu, Chunhuan Ren, Xiaoyun He, Yanting Jiang, Yina Ouyang, Mingxing Chu, and Zijun Zhang. 2022. "Oviduct Transcriptomic Reveals the Regulation of mRNAs and lncRNAs Related to Goat Prolificacy in the Luteal Phase" Animals 12, no. 20: 2823. https://doi.org/10.3390/ani12202823