Hopping Down the Main Street: Eastern Grey Kangaroos at Home in an Urban Matrix

Abstract
:Simple Summary
Abstract

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Methods
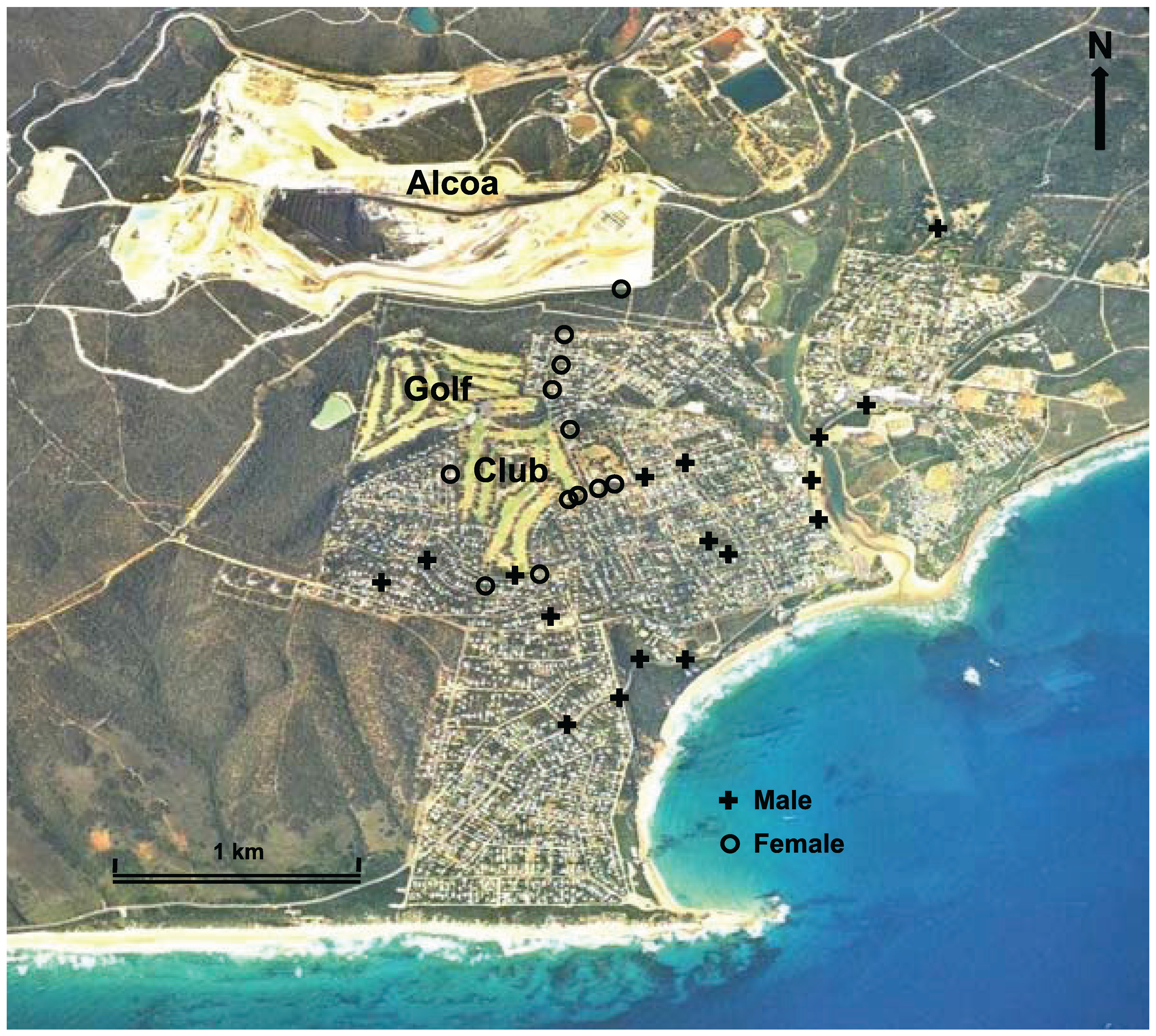
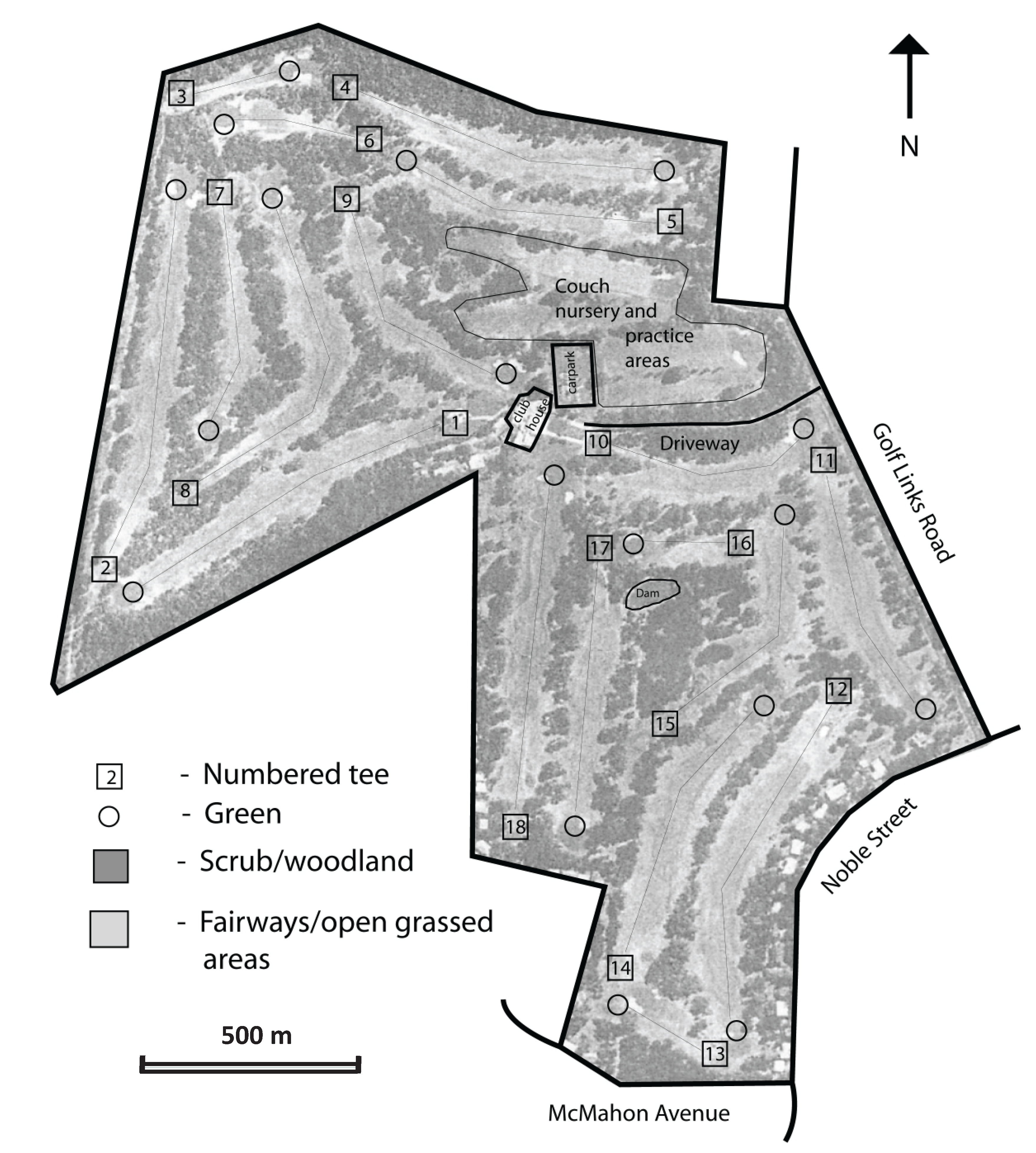
2.1. Study Sites


2.2. Capture and Marking
2.3. Population Surveys
2.4. Movements
2.5. Mortality
3. Results
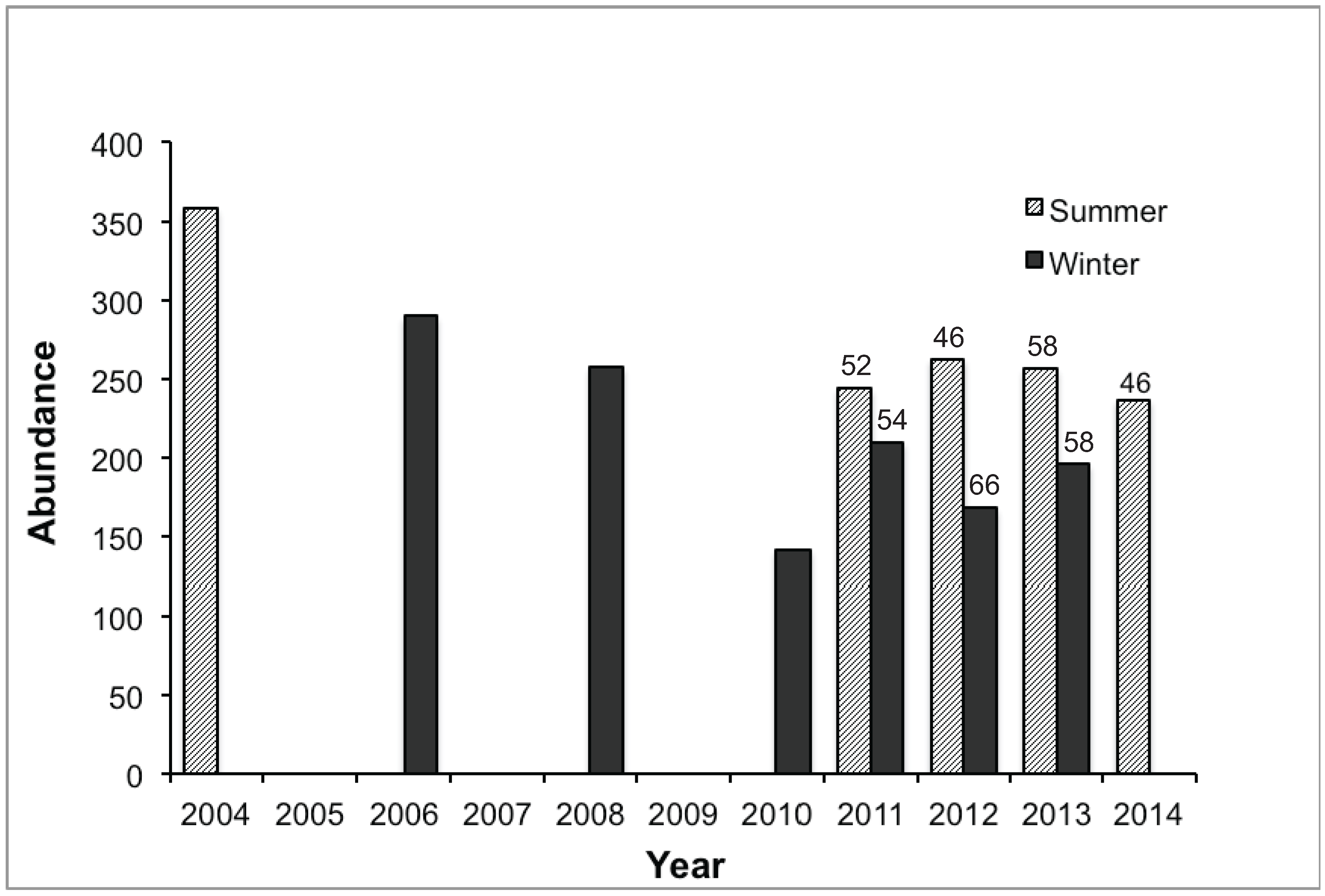
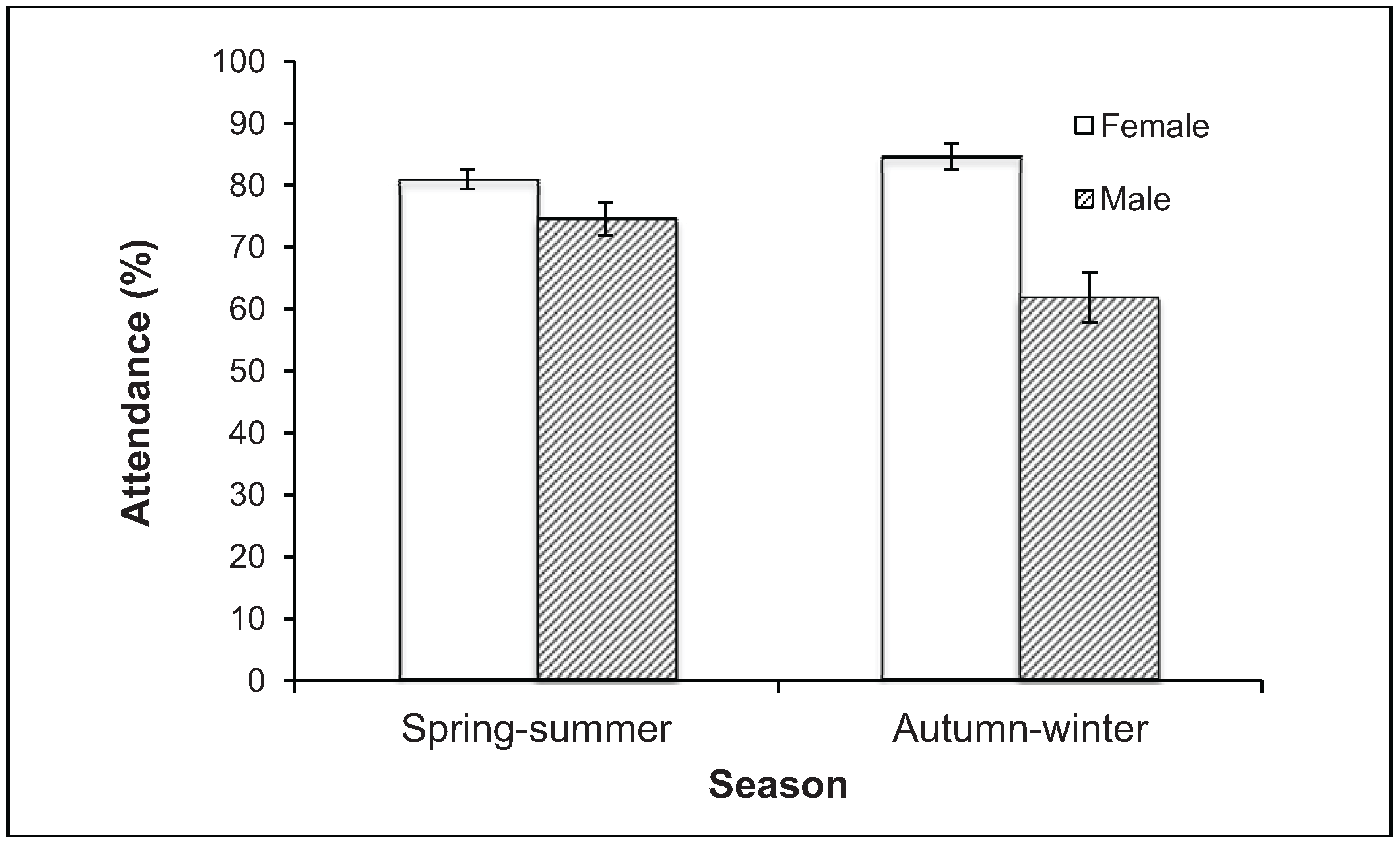
3.1. Abundance and Attendance


3.2. Fecundity and Mortality
3.3. Movement and Habitats

4. Discussion
4.1. Population Dynamics
4.2. Movement
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References and Notes
- Adams, C.E.; Lindsey, K.J.; Ash, S.J. Urban Wildlife Management; Taylor & Francis: Boca Raton, FL, USA, 2006. [Google Scholar]
- McDonnell, M.J.; Pickett, S.T. A. Ecosystem structure and function along urban-rural gradients: An unexploited opportunity for ecology. Ecology 1990, 71, 1232–1327. [Google Scholar] [CrossRef]
- Collins, J.P.; Kinzing, A.; Grimm, N.B.; Fagan, W.F.; Hope, D.; Wu, J.; Borer, E.T. A new urban ecology. Am. Sci. 2000, 88, 416–426. [Google Scholar] [CrossRef]
- Garden, J.; McAlpine, C.; Peterson, A.; Jones, D.; Possingham, H. Review of the ecology of Australian urban fauna: A focus on spatially explicit processes. Aust. Ecol. 2006, 31, 126–148. [Google Scholar] [CrossRef]
- Berry, R.J. Town Mouse, Country Mouse: adaptation and adaptability in Mus domesticus (M. musculus domesticus). Mamm. Rev. 1981, 11, 91–136. [Google Scholar] [CrossRef]
- Luniak, M. Synurbization—Adaptation of animal wildlife to urban development. In Proceedings of the 4th International Symposium on Urban Wildlife Conservation; Shaw, W.W., Harris, L.K., VanDruff, L., Eds.; University of Arizona: Tucson, AZ, USA,, 2004; pp. 267–269. [Google Scholar]
- Withey, J.C.; Marzluff, J.W. Multi-scale use of lands providing anthropogenic resources by American Crows in an urbanizing landscape. Landscape Ecol. 2009, 24, 281–293. [Google Scholar] [CrossRef]
- Dowding, C.V.; Harris, S.; Poulton, S.; Baker, P.J. Nocturnal ranging behavior of urban hedgehogs, Erinaceus europaeus, in relation to risk and reward. Anim. Behav. 2010, 80, 13–21. [Google Scholar] [CrossRef]
- Cavia, R.; Cueto, G.R.; Olga Virginia Suárez, O.V. Changes in rodent communities according to the landscape structure in an urban ecosystem. Landscape Urban Plan. 2009, 90, 11–19. [Google Scholar] [CrossRef]
- Heard, G.W.; McCarthy, M.A.; Scroggie, M.P.; Baumgartner, J.B.; Parris, K.M. A Bayesian model of metapopulation viability, with application to an endangered amphibian. Divers. Distrib. 2013, 19, 555–566. [Google Scholar] [CrossRef]
- Adams, L.W. Urban wildlife ecology and conservation: A brief history of the discipline. Urban Ecosyst. 2005, 8, 139–156. [Google Scholar] [CrossRef]
- Baker, P.J.; Harris, S. Urban mammals: What does the future hold? An analysis of the factors affecting patterns of use of residential gardens in Great Britain. Mammal Rev. 2007, 37, 297–315. [Google Scholar]
- Etter, D.R.; Hollis, K.M.; Van Deelen, T.R.; Ludwig, D.R.; Chelsvig, J.E.; Anchor, C.L.; Warner, R.E. Survival and movements of white-tailed deer in suburban Chicago, Illinois. J. Wildl. Manage. 2002, 66, 500–510. [Google Scholar] [CrossRef]
- Porter, W.F.; Underwood, B.; Woodard, J.L. Movement behavior, dispersal, and the potential for localized management of deer in a suburban environment. J. Wildl. Manage. 2004, 68, 247–256. [Google Scholar] [CrossRef]
- Harveson, P.M.; Lopez, R.R.; Collier, B.A.; Silvy, N.J. Impacts of urbanization on Florida Key deer behavior and population dynamics. Biol. Cons. 2007, 134, 321–331. [Google Scholar] [CrossRef]
- DeNicola, A.J.; Williams, S.C. Sharpshooting suburban white-tailed deer reduces deer–vehicle collisions. Human-Wildl. Confl. 2008, 2, 28–33. [Google Scholar]
- Walter, W.D.; Beringer, J.; Hansen, L.P.; Fischer, J.W.; Millspaugh, J.P.; Vercauteren, K.C. Factors affecting space use overlap by white-tailed deer in an urban landscape. Int. J. Geogr. Inf. Sci. 2011, 25, 379–392. [Google Scholar] [CrossRef]
- Ng, J.W.; Nielsen, C.; St.Clair, C.C. Landscape and traffic factors influencing deer–ehicle collisions in an urban environment. Human-Wildl. Confl. 2008, 2, 34–47. [Google Scholar]
- McCullough, D.R.; Jennings, K.W.; Gates, N.B.; Elliott, B.G.; DiDonato, J.E. Overabundant deer populations in California. Wildl. Soc. Bull. 1997, 25, 478–483. [Google Scholar]
- Wolfe, L.L.; Miller, M.W.; Williams, E.S. Feasibility of “test-and-cull” for managing chronic wasting disease in urban mule deer. Wildl. Soc. Bull. 2004, 32, 500–505. [Google Scholar] [CrossRef]
- Nielsen, C.K.; Anderson, R.G.; Grund, M.D. Landscape influences on deer-vehicle accident areas in an urban environment. J. Wildl. Manage. 2003, 67, 46–51. [Google Scholar] [CrossRef]
- Magnarelli, L.A.; Denicola, A.; Stafford, K.C.; Anderson, J.F., III. Borrelia burgdorferi in an urban environment: White-tailed deer with infected ticks and antibodies. J. Clin. Microbiol. 1995, 33, 541–544. [Google Scholar]
- Jarman, P.J. Social behavior and organization in the Macropodoidea. Adv. Stud. Behav. 1991, 20, 1–50. [Google Scholar] [CrossRef]
- Exley, B. Kangaroos on the main street? Using resources to break down stereotypes. Jigsaw: Mag. Fam. Day Care Aust. 2005, 35, 9–10. [Google Scholar]
- Merchant, J.C. Swamp wallaby Wallabia bicolor. In The Mammals of Australia, 3rd ed.; Van Dyck, S., Strahan, R., Eds.; Reed New Holland: Sydney, Australia, 2008; pp. 323–324. [Google Scholar]
- Ramp, D.; Ben-Ami, D. The effect of road-based fatalities on the viability of a peri-urban swamp wallaby population. J. Wildl. Manage. 2006, 70, 1615–1624. [Google Scholar] [CrossRef]
- Garvey, N.; Ben-Ami, D.; Ramp, D.; Croft, D B. Survival behavior of swamp wallabies during prescribed burning and wildfire. Wildl. Res. 2010, 37, 1–12. [Google Scholar] [CrossRef]
- Merchant, J.C. Agile wallaby Macropus agilis. In The Mammals of Australia, 3rd ed.; Van Dyck, S., Strahan, R., Eds.; Reed New Holland: Sydney, Australia, 2008; pp. 404–406. [Google Scholar]
- Stirrat, S.C. Seasonal changes in home-range area and habitat use by the agile wallaby (Macropusagilis). Wildl. Res. 2003, 30, 593–600. [Google Scholar] [CrossRef]
- Coulson, G. Western grey kangaroo Macropus fuliginosus. In The Mammals of Australia, 3rd ed.; Van Dyck, S., Strahan, R., Eds.; Reed New Holland: Sydney, Australia, 2008; pp. 333–334. [Google Scholar]
- Mayberry, C.; Maloney, S.K.; Mitchell, J.; Mawson, P.; Bencini, R. Reproductive implications of exposure to Toxoplasma gondii and Neospora caninum in western grey kangaroos (Macropus fuliginosus ocydromus). J. Wildl. Dis. 2014, 50, 364–368. [Google Scholar] [CrossRef]
- Coulson, G. Eastern grey kangaroo Macropus giganteus. In The Mammals of Australia, 3rd ed.; Van Dyck, S., Strahan, R., Eds.; Reed New Holland: Sydney, Australia, 2008; pp. 335–338. [Google Scholar]
- Australian Capital Territory. In ACT Kangaroo Management Plan; Territory and Municipal Services: Canberra, Australia, 2010.
- Ballard, G. Peri-urban kangaroos. Wanted? Dead or alive? In Too Close for Comfort: Contentious Issues in Human-Wildlife Encounters; Lunney, D., Munn, A., Meikle, W., Eds.; Royal Zoological Society of New South Wales: Mosman, Australia, 2008; pp. 49–51. [Google Scholar]
- Inwood, D.; Catanchin, H.; Coulson, G. Roo town slow down: A community-based kangaroo management plan for Anglesea, Victoria. In Too Close for Comfort: Contentious Issues in Human-Wildlife Encounters; Lunney, D., Munn, A., Meikle, W., Eds.; Royal Zoological Society of New South Wales: Mosman, Australia, 2008; pp. 1–8. [Google Scholar]
- King, W.J.; Wilson, M.E.; Allen, T.; Festa-Bianchet, M.; Coulson, G. A capture technique for free-ranging eastern grey kangaroos (Macropus giganteus) habituated to humans. Aust. Mammal. 2011, 33, 47–51. [Google Scholar] [CrossRef]
- Clutton-Brock, T.H.; Sheldon, B.C. Individuals and populations: The role of long term, individual-based studies of animals in ecology and evolutionary biology. Trends Ecol. Evol. 2010, 25, 562–573. [Google Scholar] [CrossRef]
- Mulder, R.A.; Guay, J.-P.; Wilson, M.; Coulson, G. Citizen science: Recruiting residents for studies of tagged urban wildlife. Wildl. Res. 2010, 37, 440–446. [Google Scholar] [CrossRef]
- Cripps, J.; Beveridge, I.; Coulson, G. The efficacy of anthelmintic drugs against nematodes infecting free-ranging eastern grey kangaroos, Macropus giganteus. J. Wildl. Dis. 2013, 49, 535–544. [Google Scholar] [CrossRef]
- Wilson, M.E.; Coulson, G.; Shaw, G.; Renfree, M.B. Deslorelin implants in free-ranging female eastern grey kangaroos (Macropus giganteus): Mechanism of action and contraceptive efficacy. Wildl. Res. 2013, 40, 403–412. [Google Scholar]
- Gélin, U.; Wilson, M.E.; Coulson, G.M.; Festa-Bianchet, M. Offspring sex, current and previous reproduction affect feeding behavior in wild eastern grey kangaroos. Anim. Behav. 2013, 86, 885–891. [Google Scholar]
- Census of Population and Housing. Economic Development, City of Greater Geelong; Australian Bureau of Statistics: Belconnen, Australia, 2001.
- Holdgate, G.R.; Smith, T.A.G.; Gallagher, S.J.; Wallace, M.W. Geology of coal-bearing Palaeogene sediments, onshore Torquay Basin, Victoria. Aust. J. Earth Sci. 2001, 48, 657–679. [Google Scholar] [CrossRef]
- Kirkpatrick, T.H. Studies of the Macropodidae in Queensland. 2. Age estimation in the grey kangaroo, the red kangaroo, the eastern wallaroo and the red-necked wallaby, with notes on dental abnormalities. Qld. J. Agric. Anim. Sci. 1965, 22, 301–317. [Google Scholar]
- Ditchkoff, S.S.; Saalfeld, S.T.; Gibson, C.J. Animal behavior in urban ecosystems: Modifications due to human-induced stress. Urban. Ecosyst. 2006, 9, 5–12. [Google Scholar] [CrossRef]
- Adderton Herbert, C. Long-acting contraceptives: A new tool to manage overabundant kangaroo populations in nature reserves and urban areas. Aust. Mammal. 2004, 26, 67–74. [Google Scholar]
- Wilson, M.E.; Coulson, G. Unpublished data. 2014.
- Porter, W.F.; Underwood, H.B. Contraception & Deer: The Irondequoit Report; Roosevelt Wild Life Station: Syracuse, NY, USA, 2001. [Google Scholar]
- van Dijk, A.I. J.M.; Beck, H.E.; Crosbie, R.S.; de Jeu, R.A. M.; Liu, Y.Y.; Podger, G.M.; Timbal, B.; Viney, N.R. The Millennium Drought in southeast Australia (2001–2009): Natural and human causes and implications for water resources, ecosystems, economy, and society. Water Resour. Res. 2013, 49, 1040–1057. [Google Scholar] [CrossRef]
- Kirkpatrick, T.H.; McEvoy, J.S. Studies of Macropodidae in Queensland. 5. Effects of drought on reproduction in the grey kangaroo (Macropus giganteus). Qld. J. Agric. Anim. Sci. 1966, 23, 339–442. [Google Scholar]
- Banks, P.B.; Newsome, A.E.; Dickman, C.R. Predation by red foxes limits recruitment in populations of eastern grey kangaroos. Aust. Ecol. 2000, 25, 283–291. [Google Scholar] [CrossRef]
- Jarman, P.J; Taylor, R.J. Ranging of eastern grey kangaroos and wallaroos on a New England pastoral property. Aust. Wildl. Res. 1983, 10, 33–38. [Google Scholar] [CrossRef]
- Jaremovic, R.V.; Croft, D.B. Comparison of techniques to determine eastern grey kangaroo home range. J. Wildl. Manage. 1987, 51, 921–930. [Google Scholar] [CrossRef]
- Moore, B.D.; Coulson, G.; Way, S. Habitat selection by adult female eastern grey kangaroos. Wildl. Res. 2002, 29, 439–445. [Google Scholar] [CrossRef]
- Viggers, K.L.; Hearn, J.P. The kangaroo conundrum: Home range studies and implications for management. J. Appl. Ecol. 2005, 42, 99–107. [Google Scholar] [CrossRef]
- Grund, M.D.; McAninch, J.B.; Wiggers, E.B. Seasonal movements and habitat use of female white-tailed deer associated with an urban park. J. Wildl. Manage. 2002, 66, 123–130. [Google Scholar] [CrossRef]
- Storm, D.J.; Nielsen, C.K.; Schauber, E.M.; Woolf, A. Space use and survival of white-tailed deer in an exurban landscape. J. Wildl. Manage. 2007, 71, 1170–1176. [Google Scholar] [CrossRef]
- MacFarlane, A.M.; Coulson, G. Sexual segregation in Australian marsupials. In Sexual Segregation in Vertebrates; Ruckstuhl, K.E., Neuhaus, H., Eds.; Cambridge University Press: Cambridge, UK, 2005; pp. 254–279. [Google Scholar]
- ACT Kangaroo Advisory Committee. Living With Eastern Grey Kangaroos in the ACT—Public Land; Third Report to the Minister for the Environment, Land And Planning; Publications and Public Communication for Environment ACT: Canberra, Australia, 1997. [Google Scholar]
- Coulson, G.M. Road-kills of macropods on a section of highway in central Victoria. Aust. Wildl. Res. 1982, 9, 21–26. [Google Scholar] [CrossRef]
- Coulson, G. Male bias in road-kills of macropods. Wildl. Res. 1997, 23, 21–25. [Google Scholar] [CrossRef]
- McAlpine, C.A.; Grigg, G.C.; Mott, J.J.; Sharma, P. Influence of landscape structure on kangaroo abundance in a disturbed semi-arid woodland of Queensland. Rangel. J. 1999, 21, 104–134. [Google Scholar] [CrossRef]
- Hennessy, C.A.; Dubach, J.; Gehrt, S.D. Long-term pair bonding and genetic evidence for monogamy among urban coyotes (Canis latrans). J. Mammal. 2012, 93, 732–742. [Google Scholar] [CrossRef]
- Cripps, J.; Beveridge, I.; Ploeg, R.; Coulson, G. Experimental manipulation reveals few subclinical impacts of a parasite community in juvenile kangaroos. IJP-PAW 2014, 3, 88–94. [Google Scholar]
- Morishita, E.; Itao, J.; Sasaki, K.; Higuchi, H. Movements of crows in urban areas, based on PHS tracking. Global Env. Res. 2003, 7, 181–192. [Google Scholar]
- Grinder, M.I.; Krausman, P.R. Home range, habitat use, and nocturnal activity of coyotes in an urban environment. J. Wildl. Manage. 2001, 65, 887–898. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Coulson, G.; Cripps, J.K.; Wilson, M.E. Hopping Down the Main Street: Eastern Grey Kangaroos at Home in an Urban Matrix. Animals 2014, 4, 272-291. https://doi.org/10.3390/ani4020272
Coulson G, Cripps JK, Wilson ME. Hopping Down the Main Street: Eastern Grey Kangaroos at Home in an Urban Matrix. Animals. 2014; 4(2):272-291. https://doi.org/10.3390/ani4020272
Chicago/Turabian StyleCoulson, Graeme, Jemma K. Cripps, and Michelle E. Wilson. 2014. "Hopping Down the Main Street: Eastern Grey Kangaroos at Home in an Urban Matrix" Animals 4, no. 2: 272-291. https://doi.org/10.3390/ani4020272