
Physiological Effects of Ergot Alkaloid and Indole-Diterpene Consumption on Sheep under Hot and Thermoneutral Ambient Temperature Conditions


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animal Management
2.2. Experimental Protocol
2.3. Statistical Analysis
3. Results
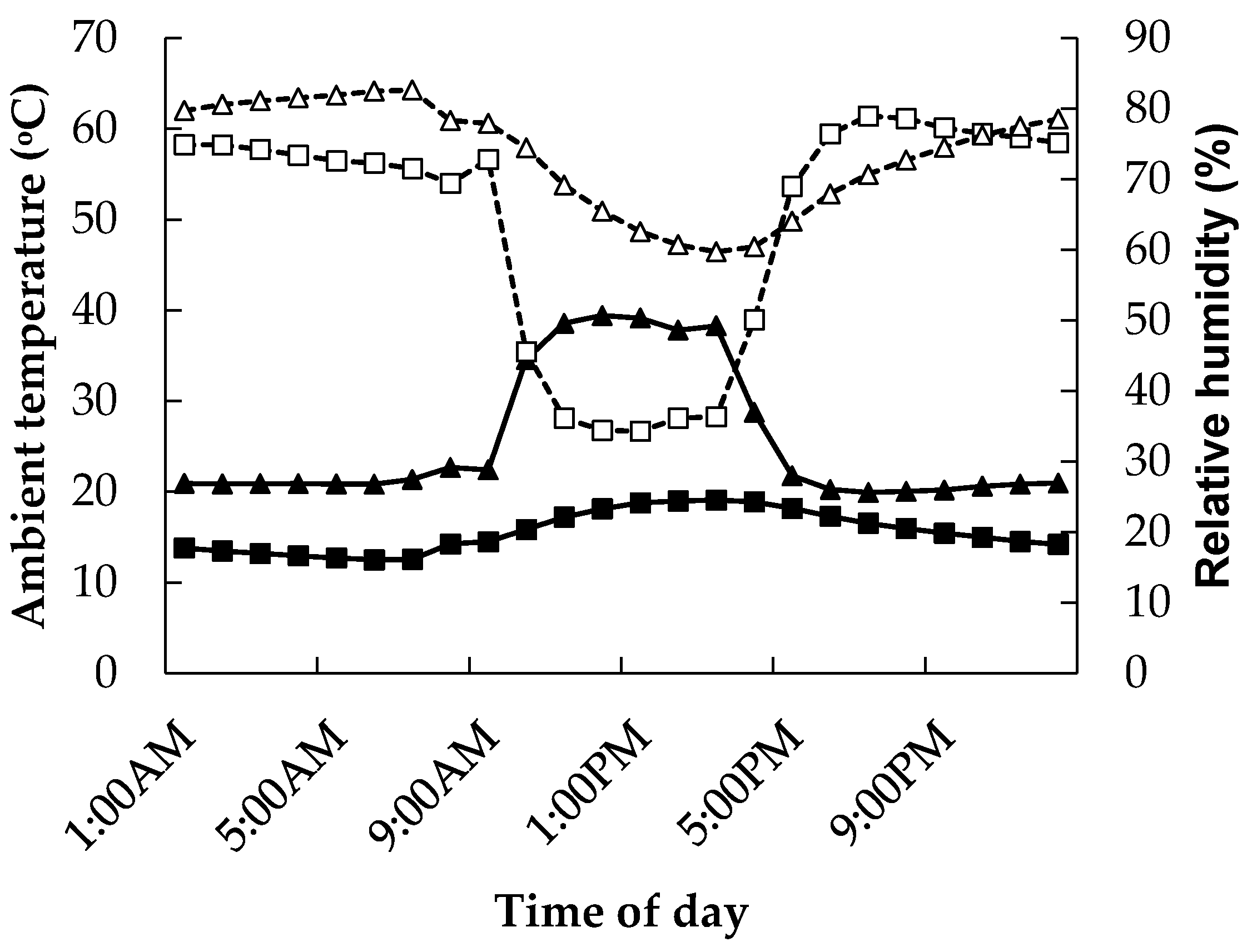
3.1. Environmental Conditions
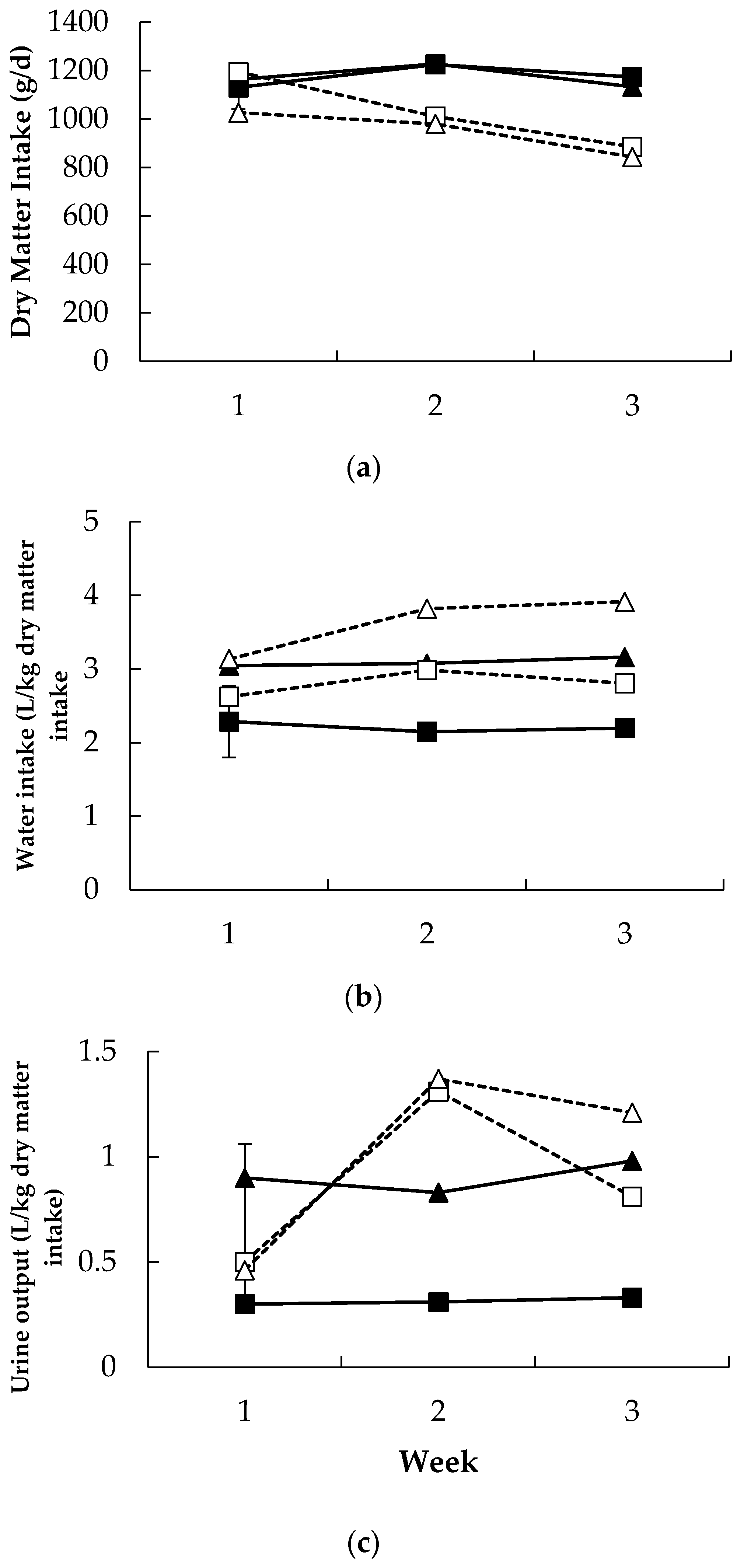
3.2. Production Response
3.3. Physiological Response
3.3.1. Overall Experimental Period
3.3.2. Temporal Patterns in Physiological Response
4. Discussion
4.1. Production Response
4.2. Physiological Response
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| PRG | Perennial ryegrass |
| PRGT | Perennial ryegrass toxicosis |
| DM | Dry matter |
| TN | Thermoneutral |
| ADG | Average daily gain |
| DMD | Dry matter digestibility |
References
- Reed, K. Perennial ryegrass toxins in australian pasture. In Perennial Ryegrass Toxicosis in Australia; Reed, K.F.M., Page, S.W., Lean, I.J., Eds.; Meat and Livestock Australia: Attwood, Australia, 2005; pp. 11–17. [Google Scholar]
- Popay, A.; Bonos, S. Biotic responses in endophytic grasses. In Neotyphodium in Cool-Season Grasses; Roberts, C.A., West, C.P., Spiers, D.E., Eds.; Blackwell Publishing: Oxford, UK, 2005; pp. 163–185. [Google Scholar]
- Osborn, T.; Schmidt, S.; Marple, D.; Rahe, C.; Steenstra, J. Effect of consuming fungus-infected and fungus-free tall fescue and ergotamine tartrate on selected physiological variables of cattle in environmentally controlled conditions. J. Anim. Sci. 1992, 70, 2501–2509. [Google Scholar] [PubMed]
- Fletcher, L.; Sutherland, B.; Fletcher, C. The impact of endophyte on the health and productivity of sheep grazing ryegrass-based pastures. In Ryegrass Endophyte: An Essential New Zealand Symbiosis; Woodfield, D.R., Matthew, C., Eds.; New Zealand Grassland Association: Napier, New Zealand, 1999; pp. 11–18. [Google Scholar]
- Di Menna, M.; Finch, S.; Popay, A.; Smith, B. A review of the neotyphodium lolii/lolium perenne symbiosis and its associated effects on animal and plant health, with particular emphasis on ryegrass staggers. N. Z. Vet. J. 2012, 60, 315–328. [Google Scholar] [CrossRef] [PubMed]
- Fiorito, I.; Bunting, L.; Davenport, G.; Boling, J. Metabolic and endocrine responses of lambs fed Acremonium coenophialum-infected or noninfected tall fescue hay at equivalent nutrient intake. J. Anim. Sci. 1991, 69, 2108–2114. [Google Scholar] [PubMed]
- Browning, R.; Leite-Browning, M. Effect of ergotamine and ergonovine on thermal regulation and cardiovascular function in cattle. J. Anim. Sci. 1997, 75, 176–181. [Google Scholar] [PubMed]
- Klotz, J. Activities and effects of ergot alkaloids on livestock physiology and production. Toxins 2015, 7, 2801–2821. [Google Scholar] [CrossRef] [PubMed]
- Fletcher, L.; Harvey, I. An association of a lolium endophyte with ryegrass staggers. N. Z. Vet. J. 1981, 32, 185–186. [Google Scholar] [CrossRef] [PubMed]
- Fletcher, L.; Harvey, I. Some observations of neurotoxin production in perennial ryegrass. N. Z. Vet. J. 1981, 30, 203–204. [Google Scholar]
- Finch, S.; Imlach, W.; Dunlop, J.; Meredith, A.; Aldrich, R.; Dalziel, J. Mode of action of tremorgens. In Proceedings of the 6th International Symposium on Fungal Endophytes of Grasses; New Zealand Grassland Association Inc.: Dunedin and Christchurch, New Zealand, 2007; pp. 363–364. [Google Scholar]
- Reed, K.; Cummins, J.; Moore, D.; Clark, A. Performance of coopworth ewe lambs exposed to low levels of ryegrass endophyte (Neotyphodium lolii) alkaloids and allowed access to a mycotoxin deactivator. Anim. Prod. Sci. 2011, 51, 225–232. [Google Scholar] [CrossRef]
- Leury, B.; Henry, M.; Kemp, S.; WebbWare, J. Developing Increased Understanding, Awareness and Potential Mitigation Strategies for Perennial Ryegrass Toxicosis in Sheep Production Systems; Meat and Livestock Australia: North Sydney, Australia, 2014. [Google Scholar]
- Henry, M.; Kemp, S.; Clarke, I.; Dunshea, F.; Leury, B. Production and physiological effects of perennial ryegrass alkaloids in merinos under mild heat conditions. Small Rumi. Res. 2016, submitted. [Google Scholar]
- Harrington, K.; Beskow, W.; Hodgson, J. Recovery and viability of seeds ingested by goats. Pasture Pests Weeds 2011, 64, 75–80. [Google Scholar]
- Henry, M.; Kemp, S.; Clarke, I.; Dunshea, F.; Leury, B. Production and physiological effects of perennial ryegrass alkaloids under thermoneutral conditions in merinos. Anim. Prod. Sci. 2015. [Google Scholar] [CrossRef]
- Hovermale, J.; Morrie Craig, A. Correlation of ergovaline and lolitrem B levels in endophyte-infected perennial ryegrass (Lolium perenne). J. Vet. Diagn. Investig. 2001, 13, 323–327. [Google Scholar] [CrossRef]
- Reed, K.; Nie, Z.; Walker, L.; Kearney, G. Fluctuations in the concentration of ergovaline and lolitrem B produced by the wild-type endophyte (Neotyphodium lolii) in perennial ryegrass (Lolium perenne) pasture. Anim. Prod. Sci. 2011, 51, 1098–1108. [Google Scholar] [CrossRef]
- Fletcher, L.; Barrell, G. Reduced liveweight gains and serum prolactin levels in hoggets grazing ryegrasses containing lolium endophyte. N. Z. Vet. J. 1984, 32, 139–140. [Google Scholar] [CrossRef] [PubMed]
- Fletcher, L.; Sutherland, B.; Fletcher, C.; Easton, H. The impact of endophyte toxins on the health of grazing sheep—An evolving story. In Proceedings of the Second Pan Pacific Veterinary Conference, Massey University, Christchurch, New Zealand, 23–28 June 1996; pp. 31–42.
- Fletcher, L.; Easton, H. The evaluation and use of endophytes for pasture improvement. In Neotyphodium/Grass Interactions; Bacon, C.W., Hill, N.S., Eds.; Plenum Press: New York, NY, USA, 1997; pp. 209–227. [Google Scholar]
- Cunningham, P.; Foot, J.; Reed, K. Perennial ryegrass (Lolium perenne) endophyte (Acremonium lolli) relationships: The australian experience. Agric. Ecosyst. Environ. 1993, 44, 157–168. [Google Scholar] [CrossRef]
- McLeay, L.; Smith, B. Effects of the mycotoxins lolitrem B and paxilline on gastrointestinal smooth muscel, and cardiovascular and respiratory systems, and temperature in sheep. In Ryegrass Endophyte: An Essential New Zealand Symbiosis; Woodfield, D.R., Matthew, C., Eds.; New Zealand Grassland Association: Napier, New Zealand, 1999; pp. 69–76. [Google Scholar]
- Sewell, J.; Hill, R.; Hume, D. Preliminary data on the effect of ryegrass endophyte on water intake and flystrike in sheep. In Proceedings of the 50th Annual Conference of the Grassland Society of Southern Australia Inc., Geelong, Australia, 6–7 August 2009; Warn, L., Ed.; Grassland Society of Southern Australia Inc.: Geelong, Australia; pp. 170–174.
- Dixon, R.; Thomas, R.; Holmes, J. Interactions between heat stress and nutrition in sheep fed roughage diets. J. Agric. Sci. Camb. 1999, 132, 351–359. [Google Scholar] [CrossRef]
- Aldrich, C.; Paterson, J.; Tate, J.; Kerley, M. The effects of endophyte-infected tall fescue consumption on diet utilization and thermal regulation in cattle. J. Anim. Sci. 1993, 71, 164–170. [Google Scholar] [PubMed]
- Hannah, S.; Paterson, J.; Williams, J.; Kerley, M.; Miner, J. Effects of increasing dietary levels of endophyte-infected tall fescue seed on diet digestibility and ruminal kinetics in sheep. J. Anim. Sci. 1990, 68, 1693–1701. [Google Scholar] [PubMed]
- Barger, J.; Tannenbaum, M. Consumption of endophyte-infected fescue seeds and osmoregulation in white-footed mice. J. Mammal. 1998, 79, 464–474. [Google Scholar] [CrossRef]
- Turner, L.; Loewer, O.; Taul, K.; Gay, N. Stimulation of summer fescue toxicosis in beef cattle by the mechanism of vasoconstriction. Am. Soc. Agric. Eng. Winter Meet. 1985, 85–4509, 17–20. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Treatment/Temperature | NilAlk | Alk | Sed 1 | p Value | ||||
|---|---|---|---|---|---|---|---|---|
| TN | Heat | TN | Heat | Alk 2 | Temp 3 | AlkxTemp | ||
| ADG (grams/day) | 83 | 83 | 34 | −34 | 61 | 0.07 | 0.44 | 0.44 |
| Faecal water (%) | 59 | 58 | 62 | 57 | 2.9 | 0.64 | 0.15 | 0.31 |
| DMD (%) | 57 | 55 | 58 | 58 | 2.2 | 0.17 | 0.58 | 0.46 |
| Treatment/Temperature | NilAlk | Alk | Sed 1 | p Value | ||||
|---|---|---|---|---|---|---|---|---|
| TN | Heat | TN | Heat | Alk 2 | Temp 3 | Week | ||
| Rectal temperature (°C) | ||||||||
| W1 | 39.55 | 39.95 | 40.12 | 40.33 | 0.11 | <0.001 | 0.002 | <0.001 |
| W2 | 39.56 | 39.88 | 39.95 | 40.11 | ||||
| Respiration rate (breaths/min) | ||||||||
| W1 | 77 | 132 | 135 | 174 | 11.5 | <0.001 | <0.001 | 0.77 |
| W2 | 77 | 134 | 119 | 184 | ||||
| Back skin temperature (°C) | ||||||||
| W1 | 38.40 | 39.05 | 38.82 | 39.31 | 0.13 | 0.004 | <0.001 | 0.002 |
| W2 | 38.32 | 39.01 | 38.63 | 39.17 | ||||
| Leg skin temperature (°C) | ||||||||
| W1 | 38.19 | 38.98 | 38.66 | 39.18 | 0.12 | 0.008 | <0.001 | <0.001 |
| W2 | 38.09 | 38.87 | 38.27 | 38.95 | ||||
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Henry, M.L.E.; Kemp, S.; Dunshea, F.R.; Leury, B.J. Physiological Effects of Ergot Alkaloid and Indole-Diterpene Consumption on Sheep under Hot and Thermoneutral Ambient Temperature Conditions. Animals 2016, 6, 37. https://doi.org/10.3390/ani6060037
Henry MLE, Kemp S, Dunshea FR, Leury BJ. Physiological Effects of Ergot Alkaloid and Indole-Diterpene Consumption on Sheep under Hot and Thermoneutral Ambient Temperature Conditions. Animals. 2016; 6(6):37. https://doi.org/10.3390/ani6060037
Chicago/Turabian StyleHenry, Michelle L. E., Stuart Kemp, Frank R. Dunshea, and Brian J. Leury. 2016. "Physiological Effects of Ergot Alkaloid and Indole-Diterpene Consumption on Sheep under Hot and Thermoneutral Ambient Temperature Conditions" Animals 6, no. 6: 37. https://doi.org/10.3390/ani6060037