
Dietary Betaine Impacts the Physiological Responses to Moderate Heat Conditions in a Dose Dependent Manner in Sheep


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Treatments
2.2. Physiological Measures
2.3. Plasma Collection and Analysis
2.4. Statistical Analysis
3. Results
3.1. Ambient Temperature, Intake and Weight
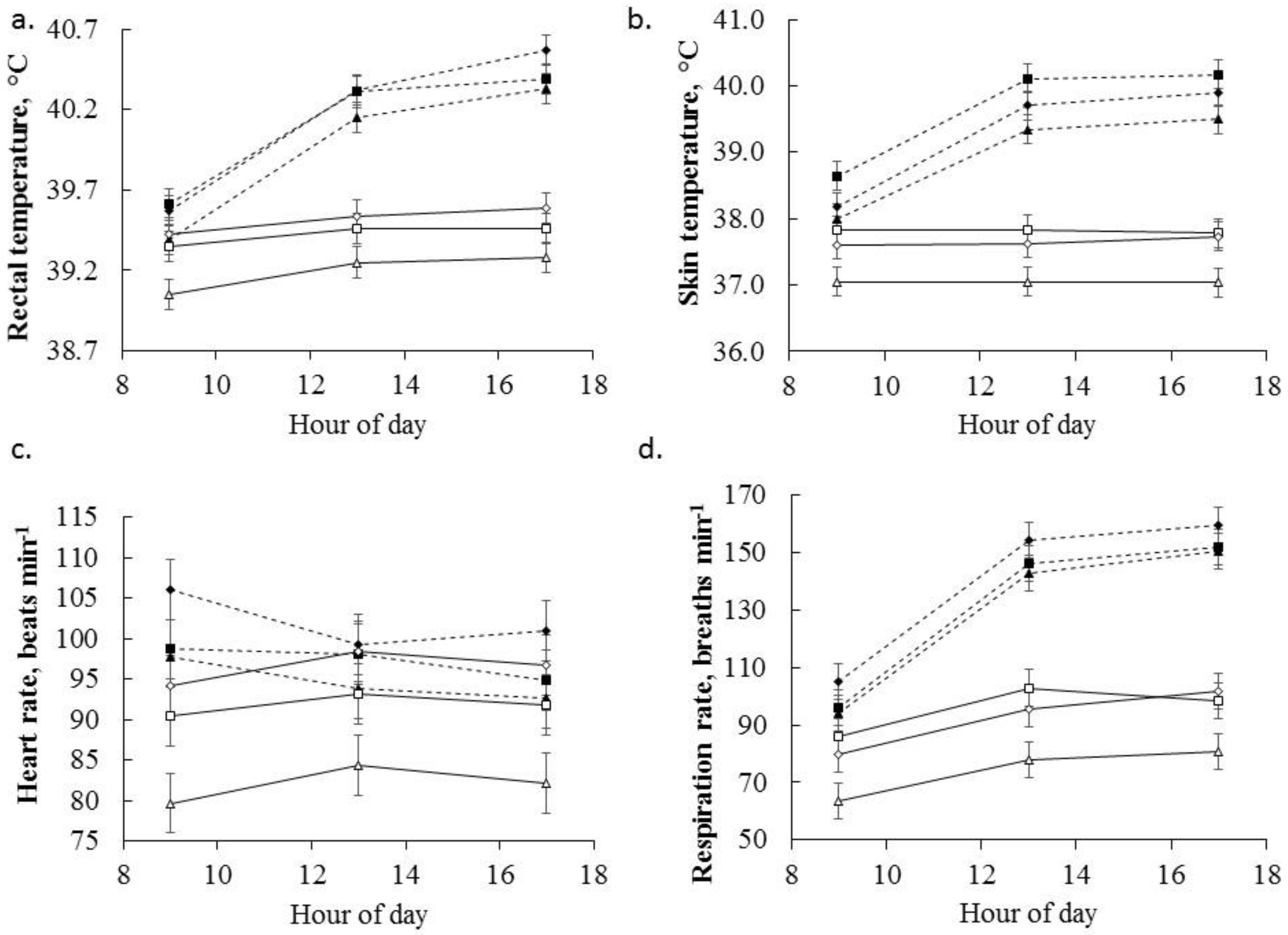
3.2. Physiological Responses to Temperature
3.3. Physiological Responses to Betaine Supplementation
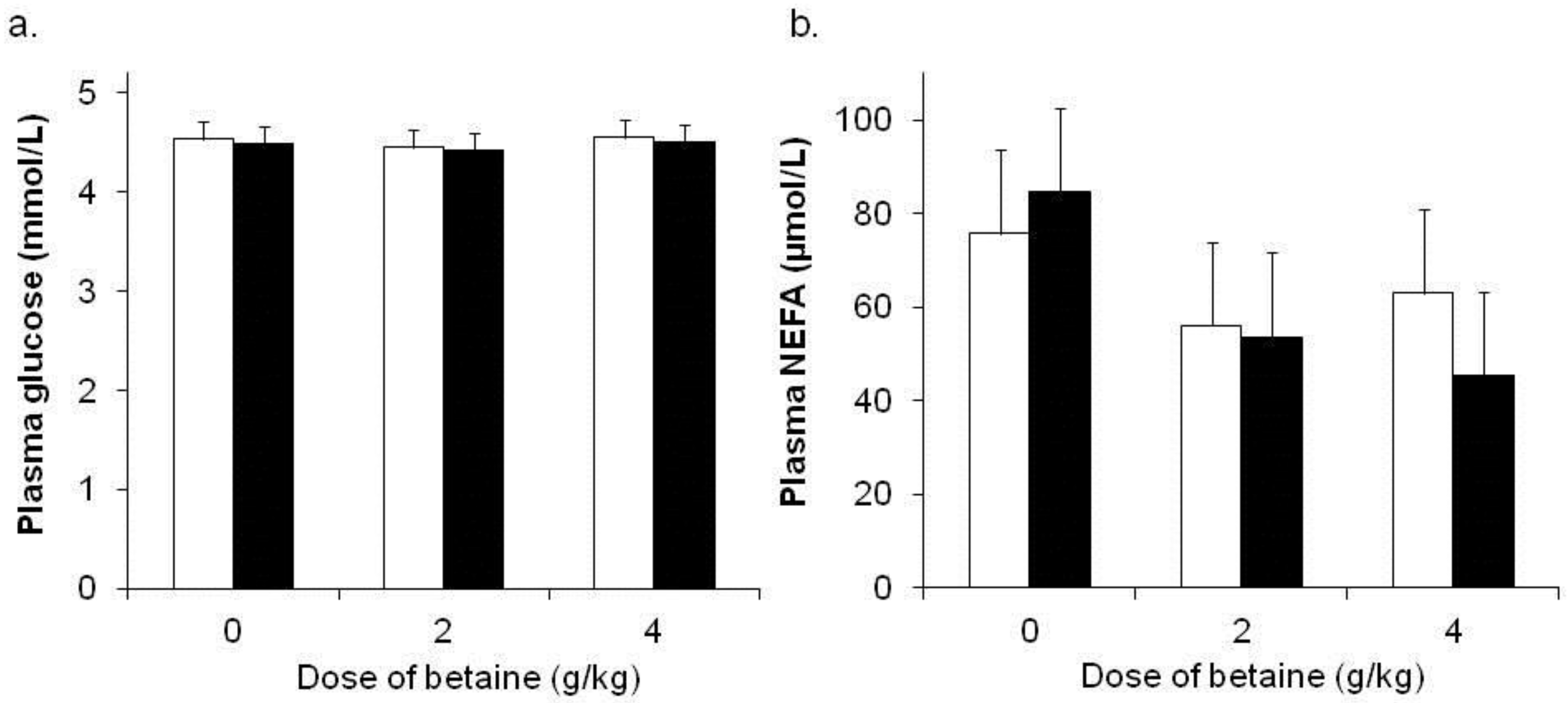
3.4. Metabolite Responses
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Horowitz, M. From molecular and cellular to integrative heat defense during exposure to chronic heat. Comp. Biochem. Physiol. A Comp. Physiol. 2002, 131, 475–483. [Google Scholar] [CrossRef]
- St-Pierre, N.R.; Cobanov, B.; Schnitkey, G. Economic losses from heat stress by US livestock industries. J. Dairy Sci. 2003, 86, E52–E77. [Google Scholar] [CrossRef]
- Vitali, A.; Segnalini, M.; Bertocchi, L.; Bernabucci, U.; Nardone, A.; Lacetera, N. Seasonal pattern of mortality and relationships between mortality and temperature-humidity index in dairy cows. J. Dairy Sci. 2009, 92, 3781–3790. [Google Scholar] [CrossRef] [PubMed]
- Alexander, L.V.; Arblaster, J.M. Assessing trends in observed and modelled climate extremes over Australia in relation to future projections. Int. J. Climatol. 2009, 29, 417–435. [Google Scholar] [CrossRef]
- Collier, R.J.; Zimbelman, R.B. Heat stress effects on cattle: What we know and what we don‘t know. In Proceedings of the 22nd Annual Southwest Nutrition & Management Conference, Tempe, AZ, USA, 22–23 February 2007; Department of Animal Sciences, The University of Arizona: Tempe, AZ, USA, 2007. [Google Scholar]
- Hahn, G.L. Dynamic responses of cattle to thermal heat loads. J. Anim. Sci. 1999, 77, 10–20. [Google Scholar] [CrossRef] [PubMed]
- Mader, T.L. Environmental stress in confined beef cattle. J. Anim. Sci. 2003, 81, E110–E119. [Google Scholar]
- Marai, I.F.M.; El-Darawany, A.A.; Fadiel, A.; Abdel-Hafez, M.A.M. Physiological traits as affected by heat stress in sheep—A review. Small Rumin. Res. 2007, 71, 1–12. [Google Scholar] [CrossRef]
- Hales, J.R.S.; Brown, G.D. Net energetic and thermoregulatory efficiency during panting in the sheep. Comp. Biochem. Physiol. A Comp. Physiol. 1974, 49, 413–422. [Google Scholar] [CrossRef]
- Macfarlane, W.V.; Morris, R.J.H.; Howard, B. Heat and water in tropical Merino sheep. Aust. J. Agric. Res. 1958, 9, 217–228. [Google Scholar] [CrossRef]
- Johnson, K.G. Body temperatures and respiratory rates of free-ranging Merino sheep in and out of shade during summer. Aust. J. Agric. Res. 1991, 42, 1347–1357. [Google Scholar] [CrossRef]
- Dunshea, F.R.; Cadogan, D.J.; Partridge, G.G. Dietary betaine and ractopamine combine to increase lean tissue deposition in finisher pigs, particularly gilts. Anim. Prod. Sci. 2009, 49, 65–70. [Google Scholar] [CrossRef]
- Fernandez-Figares, I.; Wray-Cahen, D.; Steele, N.C.; Campbell, R.G.; Hall, D.D.; Virtanen, E.; Caperna, T.J. Effect of dietary betaine on nutrient utilization and partitioning in the young growing feed restricted pig. J. Anim. Sci. 2002, 80, 421–428. [Google Scholar] [CrossRef] [PubMed]
- Lawrence, B.V.; Schinckel, A.P.; Adeola, O.; Cera, K. Impact of betaine on pig finishing performance and carcass composition. J. Anim. Sci. 2002, 80, 475–482. [Google Scholar] [CrossRef] [PubMed]
- Matthews, J.O.; Southern, L.L.; Higbie, A.D.; Persica, M.A.; Bidner, T.D. Effects of betaine on growth, carcass characteristics, pork quality and plasma metabolites of finishing pigs. J. Anim. Sci. 2001, 79, 722–728. [Google Scholar] [CrossRef] [PubMed]
- Suster, D.; Leury, B.J.; King, R.H.; Mottram, M.; Dunshea, F.R. Interrelationships between porcine somatotropin (pST), betaine, and energy level on body composition and tissue distribution of finisher boars. Aust. J. Agric. Res. 2004, 55, 983–990. [Google Scholar] [CrossRef]
- Wang, Y.Z.; Xu, Z.R.; Feng, J. The effect of betaine and DL-methionine on growth performance and carcass characteristics in meat ducks. Anim. Feed Sci. Technol. 2004, 116, 151–159. [Google Scholar] [CrossRef]
- Zhan, X.A.; Li, J.X.; Xu, Z.R.; Zhao, R.Q. Effects of methionine and betaine supplementation on growth performance, carcase composition and metabolism of lipids in male broilers. Br. Poult. Sci. 2006, 47, 576–580. [Google Scholar] [CrossRef] [PubMed]
- Bock, B.J.; Brethour, J.R.; Harmoney, K.R.; Goodall, S.R. Influence of betaine on pasture, finishing, and carcass performance in steers. Prof. Anim. Sci. 2004, 20, 53–57. [Google Scholar] [CrossRef]
- DiGiacomo, K.; Warner, R.D.; Leury, B.J.; Gaughan, J.B.; Dunshea, F.R. Dietary betaine supplementation has energy-sparing effects in feedlot cattle during summer, particularly in those without access to shade. Anim. Prod. Sci. 2014, 54, 450–458. [Google Scholar] [CrossRef]
- Fernandez, C.; Gallego, L.; Lopez-Bote, C.J. Effect of betaine on fat content in growing lambs. Anim. Feed Sci. Technol. 1998, 73, 329–338. [Google Scholar] [CrossRef]
- Moeckel, G.W.; Shadman, R.; Fogel, J.M.; Sadrzadeh, S.M.H. Organic osmolytes betaine, sorbitol and inositol are potent inhibitors of erythrocyte membrane ATPases. Life Sci. 2002, 71, 2413–2424. [Google Scholar] [CrossRef]
- Wray-Cahen, D.; Fernández-Fígares, I.; Virtanen, E.; Steele, N.C.; Caperna, T.J. Betaine improves growth, but does not induce whole body or hepatic palmitate oxidation in swine (Sus scrofa domestica). Comp. Biochem. Physiol. A Comp. Physiol. 2004, 137, 131–140. [Google Scholar] [CrossRef]
- Loest, C.A.; Titgemeyer, E.C.; Drouillard, J.S.; Blasi, D.A.; Bindel, D.J. Soybean hulls as a primary ingredient in forage-free diets for limit-fed growing cattle. J. Anim. Sci. 2001, 79, 766–774. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, A.D.; Chappell, A.; Knox, K.L. Metabolism of betaine in the ruminant. J. Anim. Sci. 1979, 49, 764–774. [Google Scholar] [CrossRef] [PubMed]
- Fernández, C.; López-Saez, A.; Gallego, L.; de la Fuente, J.M. Effect of source of betaine on growth performance and carcass traits in lambs. Anim. Feed Sci. Technol. 2000, 86, 71–82. [Google Scholar] [CrossRef]
- Baumgard, L.H.; Wheelock, J.B.; O‘Brien, M.; Schwartz, G.; Zimbelman, R.B.; Sanders, S.R.; VanBaale, M.J.; Collier, R.J.; Rhoads, M.L.; Rhoads, R.P. The differential effects of heat stress vs. underfeeding on production and post-absorptive nutrient partitioning. In Proceedings of the 22nd Annual Southwest Nutrition & Management Conference, Tempe, AZ, USA, 22–23 February 2007; pp. 116–124.
- Rhoads, M.L.; Kim, J.W.; Collier, R.J.; Crooker, B.A.; Boisclair, Y.R.; Baumgard, L.H.; Rhoads, R.P. Effects of heat stress and nutrition on lactating Holstein cows: II. Aspects of hepatic growth hormone responsiveness. J. Dairy Sci. 2009, 93, 170–179. [Google Scholar] [CrossRef] [PubMed]
- Rhoads, M.L.; Rhoads, R.P.; VanBaale, M.J.; Collier, R.J.; Sanders, S.R.; Weber, W.J.; Crooker, B.A.; Baumgard, L.H. Effects of heat stress and plane of nutrition on lactating Holstein cows: I. Production, metabolism, and aspects of circulating somatotropin. J. Dairy Sci. 2009, 92, 1986–1997. [Google Scholar] [CrossRef] [PubMed]
- Johnson, M.M.; Peters, J.P. Technical note: An improved method to quantify nonesterified fatty acids in bovine plasma. J. Anim. Sci. 1993, 71, 753–756. [Google Scholar] [PubMed]
- Payne, R.W.; Harding, S.A.; Murray, D.A.; Soutar, D.M.; Baird, D.B.; Welham, S.J.; Kane, A.F.; Gilmour, R.; Thompson, R.; Webster, R.; et al. GenStat Release 11 Reference Manual; VSN International: Hemel Hempstead, UK, 2008. [Google Scholar]
- Dunshea, F.R.; Leury, B.J.; Fahri, F.; DiGiacomo, K.; Hung, A.; Chauhan, S.; Clarke, I.J.; Collier, R.; Little, S.; Baumgard, L.H.; et al. Amelioration of thermal stress impacts in dairy cows. Anim. Prod. Sci. 2013, 53, 965–975. [Google Scholar] [CrossRef]
- Rhoads, R.P.; Baumgard, L.H.; Suagee, J.K.; Sanders, S.R. Nutritional interventions to alleviate the negative consequences of heat stress. Adv. Nutr. 2013, 4, 267–276. [Google Scholar] [CrossRef] [PubMed]
- Collier, R.J.; Eley, R.M.; Sharma, A.K.; Pereira, R.M.; Buffington, D.E. Shade management in subtropical environment for milk yield and composition in Holstein and Jersey cows. J. Dairy Sci. 1981, 64, 844–849. [Google Scholar] [CrossRef]
- Alhidary, I.A.; Shini, S.; Al Jassim, R.A.M.; Gaughan, J.B. Physiological responses of Australian Merino wethers exposed to high heat load. J. Anim. Sci. 2012, 90, 212–220. [Google Scholar] [CrossRef] [PubMed]
- Hopkins, P.; Knights, G.; Feuvre, A.L. Studies of the environmental physiology of tropical Merinos. Aust. J. Agric. Res. 1978, 29, 161–171. [Google Scholar] [CrossRef]
- Kahn, C.M.; Line, S. The Merck Veterinary Manual, 10th ed.; Merck: Whitehouse Station, NJ, USA, 2010. [Google Scholar]
- Chauhan, S.S.; Celi, P.; Leury, B.J.; Clarke, I.J.; Dunshea, F.R. Dietary antioxidants at supranutritional doses improve oxidative status and reduce the negative effects of heat stress in sheep. J. Anim. Sci. 2014, 92, 3364–3374. [Google Scholar] [CrossRef] [PubMed]
- Srikandakumar, A.; Johnson, E.H.; Mahgoub, O. Effect of heat stress on respiratory rate, rectal temperature and blood chemistry in Omani and Australian Merino sheep. Small Rumin. Res. 2003, 49, 193–198. [Google Scholar] [CrossRef]
- Stockman, C.A.; Barnes, A.L.; Maloney, S.K.; Taylor, E.; McCarthy, M.; Pethick, D. Effect of prolonged exposure to continuous heat and humidity similar to long haul live export voyages in Merino wethers. Anim. Prod. Sci. 2011, 51, 135–143. [Google Scholar] [CrossRef]
- Horowitz, M. Heat acclimation: Phenotypic plasticity and cues to the underlying molecular mechanisms. J. Therm. Biol. 2001, 26, 357–363. [Google Scholar] [CrossRef]
- DiGiacomo, K.; Dunshea, F.R.; Leury, B.J. Mild heat load alters some aspects of nutrient partitioning and gene expression in sheep. In Proceedings of the 63rd Annual Meeting of the European Federation of Animal Science, Bratislava, Slovakia, 27–31 August 2012; Wageningen Academic Publishers: Bratislava, Slovakia, 2012; p. 369. [Google Scholar]
- Caldas, T.; Demont-Caulet, N.; Ghazi, A.; Richarme, G. Thermoprotection by glycine betaine and choline. Microbiology 1999, 145, 2543–2548. [Google Scholar] [CrossRef] [PubMed]
- Craig, S.A.S. Betaine in human nutrition. Am. J. Clin. Nutr. 2004, 80, 539–549. [Google Scholar] [PubMed]
- Cronje, P. Heat stress in livestock—The role of the gut in its aetiology and a potential role for betaine in its alleviation. Recent Adv. Anim. Nutr. Aust. 2005, 15, 107–122. [Google Scholar]
- Wang, C.; Liu, Q.; Yang, W.Z.; Wu, J.; Zhang, W.W.; Zhang, P.; Dong, K.H.; Huang, Y.X. Effects of betaine supplementation on rumen fermentation, lactation performance, feed digestibilities and plasma characteristics in dairy cows. J. Agric. Sci. 2010, 148, 487–495. [Google Scholar] [CrossRef]
- Shwartz, G.; Rhoads, M.L.; VanBaale, M.J.; Rhoads, R.P.; Baumgard, L.H. Effects of a supplemental yeast culture on heat-stressed lactating Holstein cows. J. Dairy Sci. 2009, 92, 935–942. [Google Scholar] [CrossRef] [PubMed]
- Huang, Q.C.; Xu, Z.R.; Han, X.Y.; Li, W.F. Effect of dietary betaine supplementation on lipogenic enzyme activities and fatty acid synthase mRNA expression in finishing pigs. Anim. Feed Sci. Technol. 2008, 140, 365–375. [Google Scholar] [CrossRef]
- Abdelmalek, M.F.; Sanderson, S.O.; Angulo, P.; Soldevila-Pico, C.; Liu, C.; Peter, J.; Keach, J.; Cave, M.; Chen, T.; McClain, C.J.; et al. Betaine for nonalcoholic fatty liver disease: Results of a randomized placebo-controlled trial. Hepatology 2009, 50, 1818–1826. [Google Scholar] [CrossRef] [PubMed]
- Borgschulte, G.; Kathirvel, E.; Herrera, M.; French, S.W.; Morgan, T.R.; Morgan, K.; Bottiglieri, T. M1769 Betaine treatment reverses insulin resistance and fatty liver disease without reducing oxidative stress or endoplasmic reticulum stress in an animal model of NAFLD. Gastroenterology 2008, 134, A-414–A-415. [Google Scholar] [CrossRef]
- Ji, C.; Kaplowitz, N. Betaine decreases hyperhomocysteinemia, endoplasmic reticulum stress, and liver injury in alcohol-fed mice. Gastroenterology 2003, 124, 1488–1499. [Google Scholar] [CrossRef]
- Wang, Z.; Yao, T.; Pini, M.; Zhou, Z.; Fantuzzi, G.; Song, Z. Betaine improved adipose tissue function in mice fed a high fat diet: A mechanism for hepatoprotective effect of betaine in nonalcholic fatty liver disease. Am. J. Physiol. Gastrointest. Liver Physiol. 2010, 298, G634–G642. [Google Scholar] [CrossRef] [PubMed]
- Banskalieva, V.; Puchala, R.; Goetsch, A.L.; Luo, J.; Sahlu, T. Effects of ruminally protected betaine and choline on net flux of nutrients across the portal-drained viscera and liver of meat goat wethers consuming diets differing in protein concentration. Small Rumin. Res. 2005, 57, 193–202. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Temperature (T) | Thermoneutral (TN) | Heat (HE) | p-Value 1 | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Betaine (B), g/d | 0 | 2 | 4 | 0 | 2 | 4 | Sed a | T | B | 0 vs. 2 + 4 | 2 vs. 4 |
| Feed intake, g/d | 1171 | 1237 | 1317 | 1371 | 1329 | 1342 | 94.1 | 0.060 | 0.66 | 0.56 | 0.51 |
| Water intake, L/d | 3.21 | 3.38 | 3.97 | 4.84 | 4.61 | 5.16 | 0.504 | <0.001 | 0.20 | 0.40 | 0.11 |
| Daily gain, g/d | 194 | 271 | 261 | 279 | 331 | 305 | 43.7 | 0.016 | 0.11 | 0.04 | 0.55 |
| Feed conversion efficiency | 0.17 | 0.22 | 0.20 | 0.20 | 0.25 | 0.20 | 0.038 | 0.15 | 0.18 | 0.10 | 0.38 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
DiGiacomo, K.; Simpson, S.; Leury, B.J.; Dunshea, F.R. Dietary Betaine Impacts the Physiological Responses to Moderate Heat Conditions in a Dose Dependent Manner in Sheep. Animals 2016, 6, 51. https://doi.org/10.3390/ani6090051
DiGiacomo K, Simpson S, Leury BJ, Dunshea FR. Dietary Betaine Impacts the Physiological Responses to Moderate Heat Conditions in a Dose Dependent Manner in Sheep. Animals. 2016; 6(9):51. https://doi.org/10.3390/ani6090051
Chicago/Turabian StyleDiGiacomo, Kristy, Sarah Simpson, Brian J. Leury, and Frank R. Dunshea. 2016. "Dietary Betaine Impacts the Physiological Responses to Moderate Heat Conditions in a Dose Dependent Manner in Sheep" Animals 6, no. 9: 51. https://doi.org/10.3390/ani6090051