Primiparous and Multiparous Sows Have Largely Similar Colostrum and Milk Composition Profiles Throughout Lactation

 , ,
, ,  ,
,
Abstract
:Simple summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Experimental Animals
2.3. Colostrum and Milk Collection
2.4. Assay for Total Fat
2.5. Total IgG, Protein, Lactose, and Energy Calculation
2.6. Statistical Analysis
3. Results
3.1. Farrowing Performance
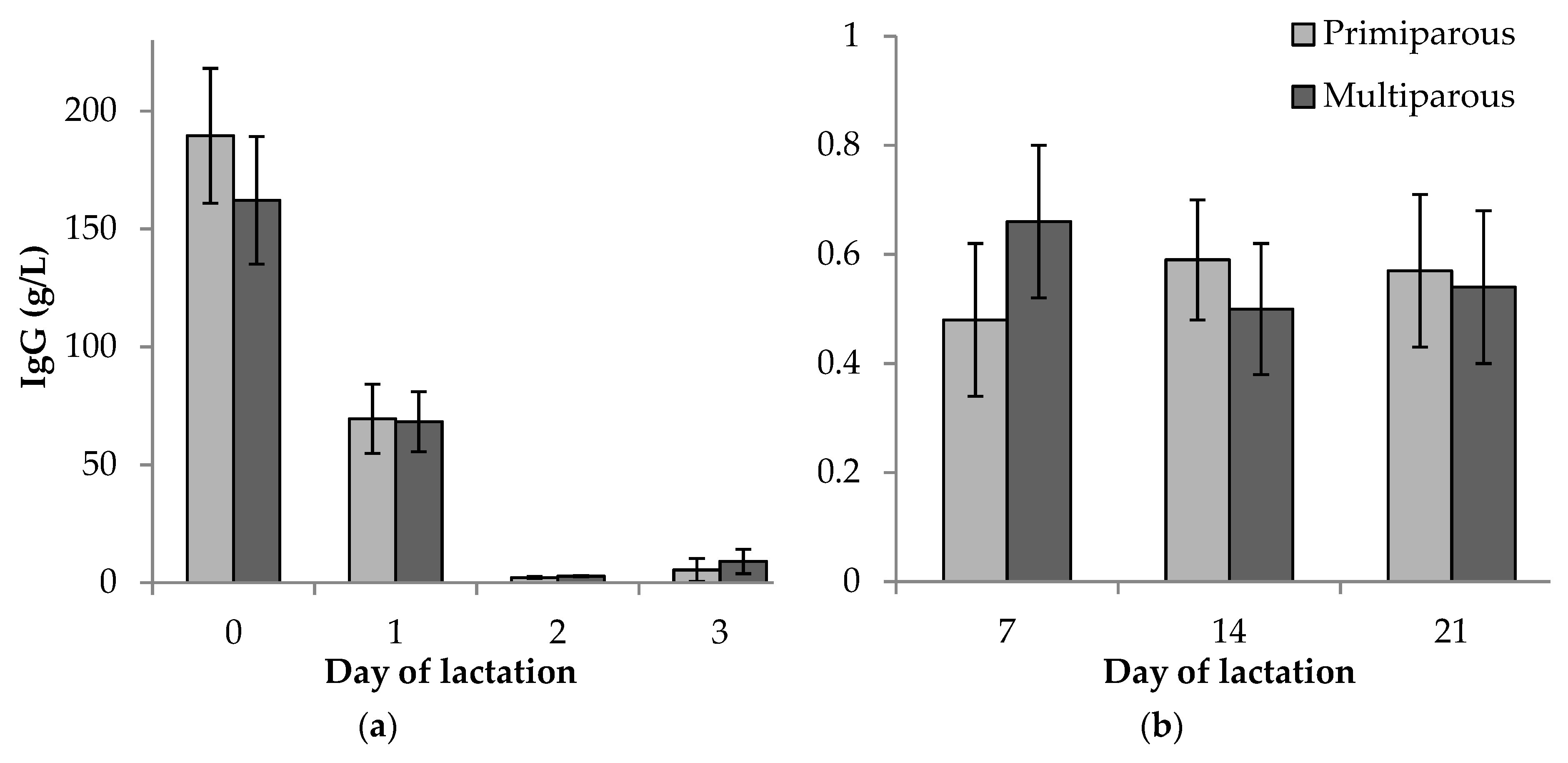
3.2. Immunoglobulin G
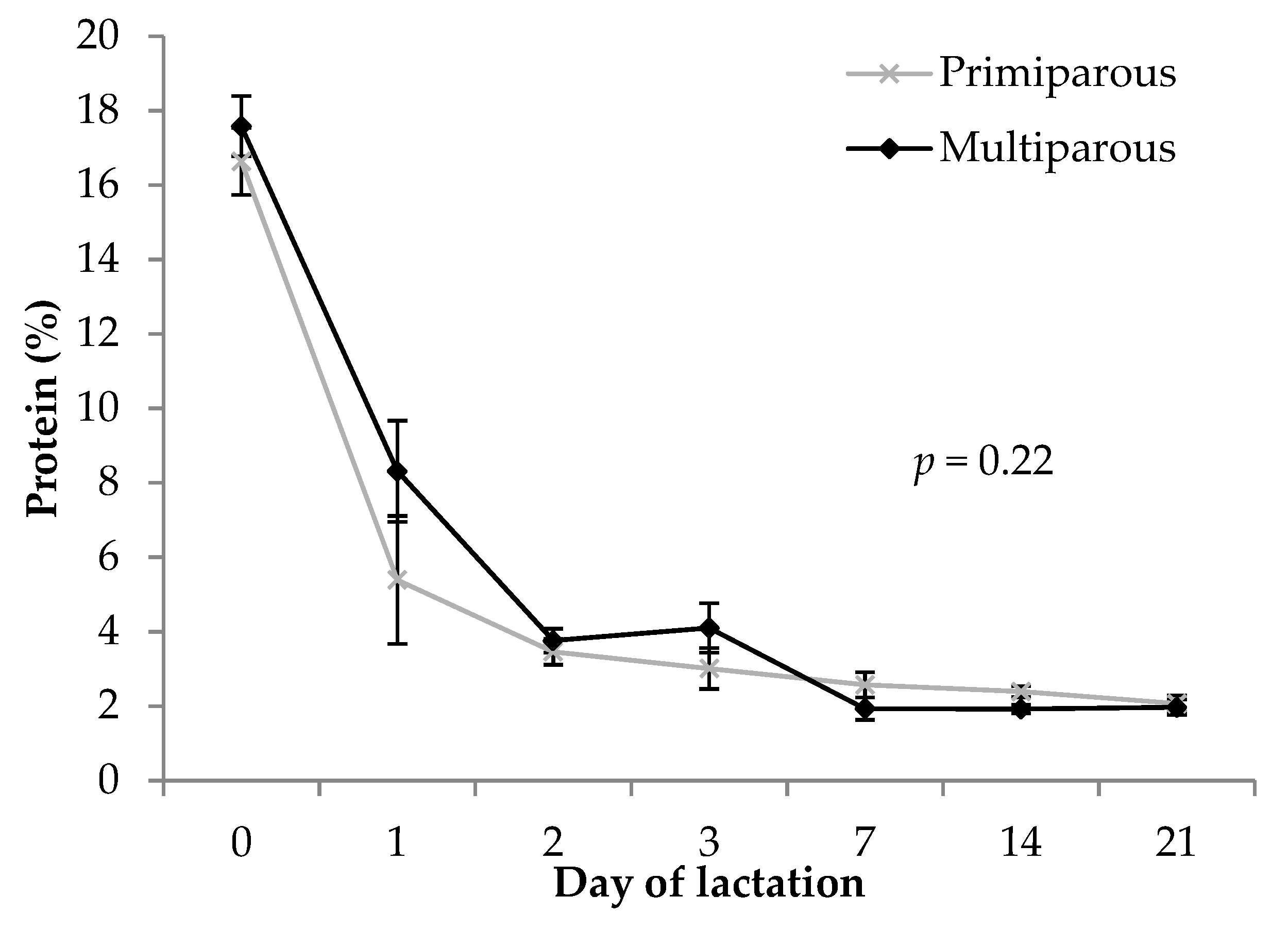
3.3. Protein
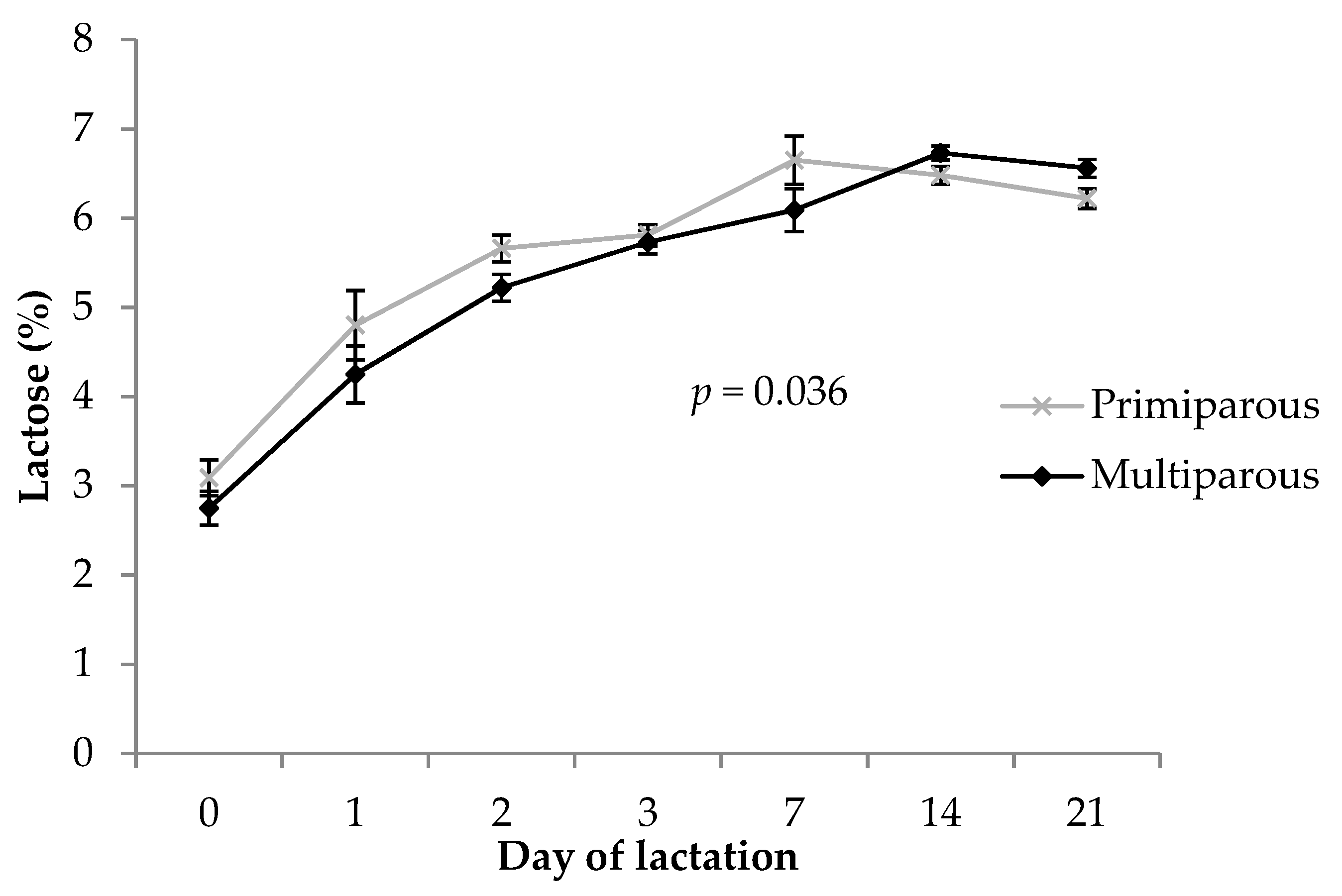
3.4. Lactose
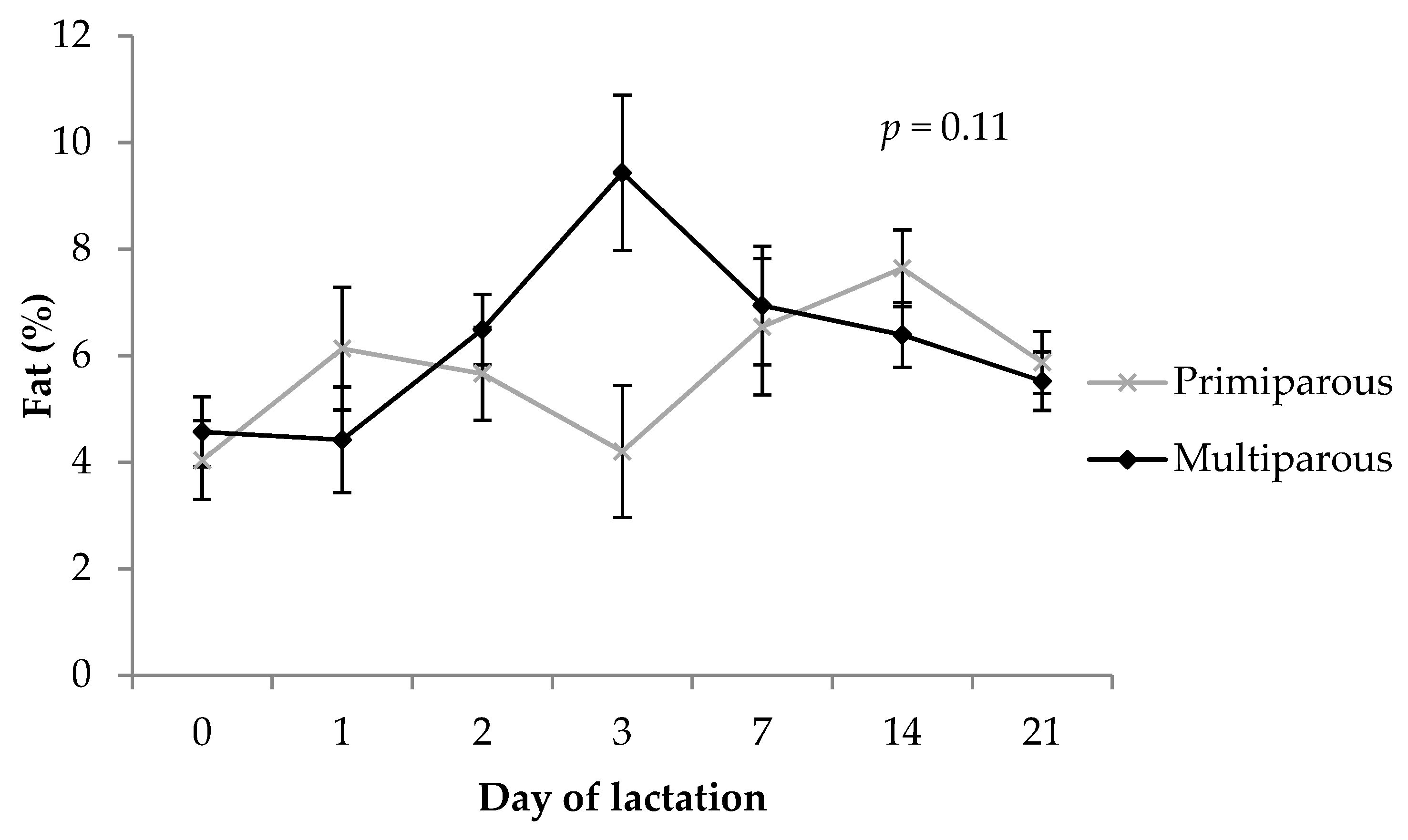
3.5. Fat
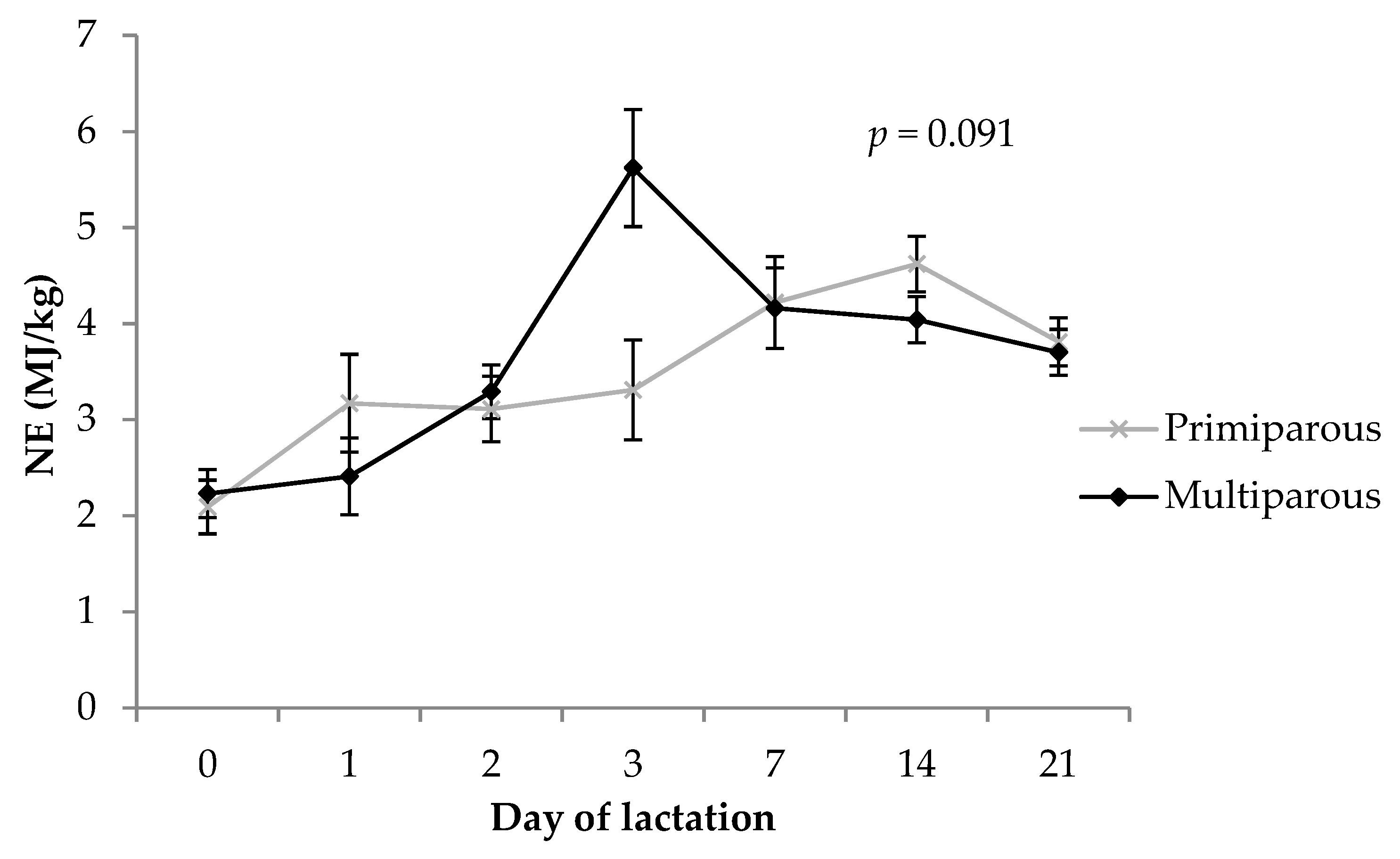
3.6. Net Energy Content
4. Discussion
5. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Craig, J.R.; Collins, C.L.; Bunter, K.L.; Cottrell, J.J.; Dunshea, F.R.; Pluske, J.R. Poorer lifetime growth performance of gilt progeny compared with sow progeny is largely due to weight differences at birth and reduced growth in the preweaning period, and is not improved by progeny segregation after weaning. J. Anim. Sci. 2017, 95, 4904–4916. [Google Scholar] [CrossRef] [PubMed]
- Carney-Hinkle, E.E.; Tran, H.; Bundy, J.W.; Moreno, R.; Miller, P.S.; Burkey, T.E. Effect of dam parity on litter performance, transfer of passive immunity, and progeny microbial ecology. J. Anim. Sci. 2013, 91, 2885–2893. [Google Scholar] [CrossRef] [PubMed]
- Holyoake, P.K. Dam parity affects the performance of nursery pigs. In Proceedings of the 19th International Pig Veterinary Society Congress, Copenhangen, Denmark, 16–19 July 2006; p. 149. [Google Scholar]
- Farmer, C.; Quesnel, H. Nutritional, hormonal, and environmental effects on colostrum in sows. J. Anim. Sci. 2009, 87, 56–64. [Google Scholar] [CrossRef] [PubMed]
- Theil, P.; Nielsen, M.; Sørensen, M.; Lauridsen, C. Lactation, milk and suckling. In Nutritional Physiology of Pigs; Bach Knudsen, K.E., Kjeldsen, N.J., Poulsen, H.D., Jensen, B.B., Eds.; Danish Pig Research Centre: Copenhagen, Denmark, 2012; pp. 1–47. [Google Scholar]
- King, R.H. Factors that influence milk production in well-fed sows. J. Anim. Sci. 2000, 78, 19–25. [Google Scholar] [CrossRef]
- Balzani, A.; Cordell, H.J.; Sutcliffe, E.; Edwards, S.A. Sources of variation in udder morphology of sows. J. Anim. Sci. 2016, 94, 394–400. [Google Scholar] [CrossRef] [PubMed]
- Pluske, J.R.; Williams, I.H.; Zak, L.J.; Clowes, E.J.; Cegielski, A.C.; Aherne, F.X. Feeding lactating primiparous sows to establish three divergent metabolic states: III. Milk production and pig growth. J. Anim. Sci. 1998, 76, 1165–1171. [Google Scholar] [CrossRef] [PubMed]
- Friendship, R.M.; O’Sullivan, T.L. Sow health. In The Gestating and Lactating Sow; Farmer, C., Ed.; Wageningen Academic Publishers: Wageningen, The Netherlands, 2015; pp. 409–421. [Google Scholar]
- King, R.H.; Dunkin, A.C. The effect of nutrition on the reproductive performance of first-litter sows. 3. The response to graded increases in food intake during lactation. Anim. Sci. 1986, 42, 119–125. [Google Scholar] [CrossRef]
- Geale, P. Characterisation of Porcine Colostrum and its Impact on Piglet Growth. Ph.D. Thesis, University of Sydney, Sydney, Australia, 2011. [Google Scholar]
- Szyndler-Nędza, M. Coefficients of repeatability for colostrum and milk composition of PLW and PL sows over three consecutive lactations. Livest. Sci. 2016, 185, 56–60. [Google Scholar] [CrossRef]
- Picone, G.; Zappaterra, M.; Luise, D.; Trimigno, A.; Capozzi, F.; Motta, V.; Davoli, R.; Costa, L.N.; Bosi, P.; Trevisi, P. Metabolomics characterization of colostrum in three sow breeds and its influences on piglets’ survival and litter growth rates. J. Anim. Sci. Biotechnol. 2018, 9, 23. [Google Scholar] [CrossRef]
- Bourne, F.J.; Newby, T.J.; Chidlow, J.W. The influence of route of vaccination on the systemic and local immune response in the pig. Res. Vet. Sci. 1975, 18, 244–248. [Google Scholar] [CrossRef]
- Mainau, E.; Temple, D.; Manteca, X. Experimental study on the effect of oral meloxicam administration in sows on pre-weaning mortality and growth and immunoglobulin G transfer to piglets. Prev. Vet. Med. 2016, 126, 48–53. [Google Scholar] [CrossRef] [PubMed]
- Quesnel, H.; Farmer, C.; Devillers, N. Colostrum intake: Influence on piglet performance and factors of variation. Livest. Sci. 2012, 146, 105–114. [Google Scholar] [CrossRef]
- Peters, J.C.; Mahan, D.C. Effects of dietary organic and inorganic trace mineral levels on sow reproductive performances and daily mineral intakes over six parities. J. Anim. Sci. 2008, 86, 2247–2260. [Google Scholar] [CrossRef] [PubMed]
- Inoue, T.; Kitano, K.; Inoue, K. Possible factors influencing the immunoglobulin G concentration in swine colostrum. Am. J. Vet. Res. 1980, 41, 1134–1136. [Google Scholar] [PubMed]
- Wu, W.Z.; Wang, X.Q.; Wu, G.Y.; Kim, S.W.; Chen, F.; Wang, J.J. Differential composition of proteomes in sow colostrum and milk from anterior and posterior mammary glands. J. Anim. Sci. 2010, 88, 2657–2664. [Google Scholar] [CrossRef] [PubMed]
- Atwood, C.S.; Hartmann, P.E. Collection of fore and hind milk from the sow and the changes in milk composition during suckling. J. Dairy Res. 1992, 59, 287–298. [Google Scholar] [CrossRef] [PubMed]
- Farmer, C.; Lessard, M.; Knight, C.H.; Quesnel, H. Oxytocin injections in the postpartal period affect mammary tight junctions in sows. J. Anim. Sci. 2017, 95, 3532–3539. [Google Scholar] [CrossRef]
- Foisnet, A.; Farmer, C.; David, C.; Quesnel, H. Farrowing induction induces transient alterations in prolactin concentrations and colostrum composition in primiparous sows. J. Anim. Sci. 2011, 89, 3048–3059. [Google Scholar] [CrossRef]
- Vallet, J.L.; Miles, J.R. The effect of farrowing induction on colostrum and piglet serum immunocrits is dependent on parity. J. Anim. Sci. 2017, 95, 688–696. [Google Scholar] [CrossRef]
- NHMRC. The Australian Code for the Care and Use of Animals for Scientific Purposes, 8th ed.; National Health and Medical Research Council: Canberra, Australia, 2013. [Google Scholar]
- Forcato, D.O.; Carmine, M.P.; Echeverria, G.E.; Pécora, R.P.; Kivatinitz, S.C. Milk fat content measurement by a simple UV spectrophotometric method: An alternative screening method. J. Dairy Sci. 2005, 88, 478–481. [Google Scholar] [CrossRef]
- MyAssays Ltd. MyCurveFit Online Curve Fitting. Available online: https://mycurvefit.com/ (accessed on 5 February 2018).
- Hansen, A.V.; Strathe, A.B.; Kebreab, E.; France, J.; Theil, P.K. Predicting milk yield and composition in lactating sows: A Bayesian approach. J. Anim. Sci. 2012, 90, 2285–2298. [Google Scholar] [CrossRef] [PubMed]
- Theil, P.K.; Lauridsen, C.; Quesnel, H. Neonatal piglet survival: Impact of sow nutrition around parturition on fetal glycogen deposition and production and composition of colostrum and transient milk. Animal 2014, 8, 1021–1030. [Google Scholar] [CrossRef] [PubMed]
- Hurley, W.L. Composition of sow colostrum and milk. In The Gestating and Lactating Sow; Farmer, C., Ed.; Wageningen Academic Publishers: Wageningen, The Netherlands, 2015; pp. 193–232. [Google Scholar]
- Speer, V.C.; Cox, D.F. Estimating Milk Yield of Sows. J. Anim. Sci. 1984, 59, 1281–1285. [Google Scholar] [CrossRef] [PubMed]
- Beyer, M.; Jentsch, W.; Kuhla, S.; Wittenburg, H.; Kreienbring, F.; Scholze, H.; Rudolph, P.E.; Metges, C.C. Effects of dietary energy intake during gestation and lactation on milk yield and composition of first, second and fourth parity sows. Arch. Anim. Nutr. 2007, 61, 452–468. [Google Scholar] [CrossRef] [PubMed]
- Cottrell, J.J.; Craig, J.; Wijesiriwardana, U.A.; Fothergill, L.; Ringuet, M.T.; O’Hallorhan, K.; Turpin, D.L.; Munoz, L.M.; Collins, C.L.; Furness, J.B.; et al. The Gastrointestinal Tract of Piglets From First Parity Sows Develops More Slowly And Is More Permeable Than Piglets From Later Parity Sows. FASEB J. 2017, 31, 792. [Google Scholar]
- Craig, J.R.; Dunshea, F.R.; Cottrell, J.J.; Wijesiriwardana, U.A.; Furness, J.B.; Pluske, J.R. Comparison of lactose digestive capacity in the small intestine between gilt and sow progeny around birth and weaning. In Proceedings of the 14th International Symposium on Digestive Physiology of Pigs, Brisbane, Australia, 22–24 August 2018. [Google Scholar]
- Da Silva, A.; Dalto, D.; Lozano, A.; De Oliveira, E.; Gavioli, D.; De Oliveira, J.; Romero, N.; Da Silva, C. Differences in muscle characteristics of piglets related to the sow parity. Can. J. Anim. Sci. 2013, 93, 471–475. [Google Scholar] [CrossRef]
- van Wettere, W.H.E.J.; Willson, N.-L.; Pain, S.J.; Forder, R.E.A. Effect of oral polyamine supplementation pre-weaning on piglet growth and intestinal characteristics. Animal 2016, 10, 1655–1659. [Google Scholar] [CrossRef]
- Theil, P.K.; Flummer, C.; Hurley, W.L.; Kristensen, N.B.; Labouriau, R.L.; Sørensen, M.T. Mechanistic model to predict colostrum intake based on deuterium oxide dilution technique data and impact of gestation and prefarrowing diets on piglet intake and sow yield of colostrum. J. Anim. Sci. 2014, 92, 5507–5519. [Google Scholar] [CrossRef]
- Devillers, N.; Farmer, C.; Le Dividich, J.; Prunier, A. Variability of colostrum yield and colostrum intake in pigs. Animal 2007, 1, 1033–1041. [Google Scholar] [CrossRef]
- Quesnel, H. Colostrum production by sows: Variability of colostrum yield and immunoglobulin G concentrations. Animal 2011, 5, 1546–1553. [Google Scholar] [CrossRef]
- Declerck, I.; Sarrazin, S.; Dewulf, J.; Maes, D. Sow and piglet factors determining variation of colostrum intake between and within litters. Animal 2017, 11, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Declerck, I.; Dewulf, J.; Piepers, S.; Decaluwé, R.; Maes, D. Sow and litter factors influencing colostrum yield and nutritional composition. J. Anim. Sci. 2015, 93, 1309–1317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ison, S.H.; Jarvis, S.; Ashworth, C.J.; Rutherford, K.M.D. The effect of post-farrowing ketoprofen on sow feed intake, nursing behaviour and piglet performance. Livest. Sci. 2017, 202, 115–123. [Google Scholar] [CrossRef] [Green Version]
- Kielland, C.; Rootwelt, V.; Reksen, O.; Framstad, T. The association between immunoglobulin G in sow colostrum and piglet plasma. J. Anim. Sci. 2015, 93, 4453–4462. [Google Scholar] [CrossRef] [PubMed]
- Miller, Y.J.; Collins, A.M.; Emery, D.; Begg, D.J.; Smits, R.J.; Holyoake, P.K. Piglet performance and immunity is determined by the parity of both the birth dam and the rearing dam. Anim. Prod. Sci. 2012, 53, 46–51. [Google Scholar] [CrossRef]
- Cabrera, R.A.; Lin, X.; Campbell, J.M.; Moeser, A.J.; Odle, J. Influence of birth order, birth weight, colostrum and serum immunoglobulin G on neonatal piglet survival. J. Anim. Sci. Biotechnol. 2012, 3, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Klobasa, F.; Butler, J.E.; Werhahn, E.; Habe, F. Maternal-neonatal immunoregulation in swine. II. Influence of multiparity on de novo immunoglobulin synthesis by piglets. Vet. Immunol. Immunopathol. 1986, 11, 149–159. [Google Scholar] [CrossRef]
- Bland, I.M.; Rooke, J.A.; Bland, V.C.; Sinclair, A.G.; Edwards, S.A. Appearance of immunoglobulin G in the plasma of piglets following intake of colostrum, with or without a delay in sucking. Anim. Sci. 2003, 77, 277–286. [Google Scholar] [CrossRef]
- Klobasa, F.; Werhahn, E.; Butler, J.E. Composition of Sow Milk During Lactation. J. Anim. Sci. 1987, 64, 1458–1466. [Google Scholar] [CrossRef] [Green Version]
- Craig, J.R.; Cottrell, J.J.; Wijesiriwardana, U.A.; Furness, J.B.; Dunshea, F.R.; Pluske, J. Gilt progeny have lower serum immunoglobulin G (IgG) concentrations than sow progeny, but not as a result of concentrations in colostrum and milk. In Proceedings of the 10th International Conference on Pig Reproduction, Columbia, MO, USA, 11–14 June 2017; p. 42. [Google Scholar]
- Huser, J.S.; Kennett, T.E.; Plush, K.J.; Pitchford, W.S.; Lines, D.S. Neonatal split suckling has no impact on pre-and post-weaning piglet growth. Anim. Prod. Sci. 2015, 55, 1482. [Google Scholar] [CrossRef]
- Devillers, N.; Le Dividich, J.; Farmer, C.; Mounier, A.; Lefebvre, M.; Prunier, A. Origin and consequences of the variability of colostrum production by the sows and of its intake by the piglets. J. Rech. Porc. 2005, 37, 435–442. [Google Scholar]
- Hendrix, W.F.; Kelley, K.W.; Gaskins, C.T.; Hinrichs, D.J. Porcine neonatal survival and serum gamma globulins. J. Anim. Sci. 1978, 47, 1281–1286. [Google Scholar] [CrossRef] [PubMed]
- Rooke, J.A.; Bland, I.M. The acquisition of passive immunity in the new-born piglet. Livest. Prod. Sci. 2002, 78, 13–23. [Google Scholar] [CrossRef]
- Trayhurn, P.; Temple, N.J.; Aerde, J.V. Evidence from immunoblotting studies on uncoupling protein that brown adipose tissue is not present in the domestic pig. Can. J. Physiol. Pharmacol. 1989, 67, 1480–1485. [Google Scholar] [CrossRef] [PubMed]
- Le Dividich, J.; Rooke, J.A.; Herpin, P. Nutritional and immunological importance of colostrum for the new-born pig. J. Agric. Sci. 2005, 143, 469–485. [Google Scholar] [CrossRef]
- Speer, V.C.; Brown, H.; Quinn, L.; Catron, D.V. The Cessation of Antibody Absorption in the Young Pig. J. Immunol. 1959, 83, 632–634. [Google Scholar] [PubMed]
- Le Dividich, J.; Charneca, R.; Thomas, F. Relationship between birth order, birth weight, colostrum intake, acquisition of passive immunity and pre-weaning mortality of piglets. Span. J. Agric. Res. 2017, 15, e0603. [Google Scholar] [CrossRef] [Green Version]
- Ferrari, C.V.; Sbardella, P.E.; Bernardi, M.L.; Coutinho, M.L.; Vaz Jr, I.S.; Wentz, I.; Bortolozzo, F.P. Effect of birth weight and colostrum intake on mortality and performance of piglets after cross-fostering in sows of different parities. Prev. Vet. Med. 2014, 114, 259–266. [Google Scholar] [CrossRef]
- Bandrick, M.; Ariza-Nieto, C.; Baidoo, S.K.; Molitor, T.W. Colostral antibody-mediated and cell-mediated immunity contributes to innate and antigen-specific immunity in piglets. Dev. Comp. Immunol. 2014, 43, 114–120. [Google Scholar] [CrossRef] [Green Version]
- Rooke, J.A.; Carranca, C.; Bland, I.M.; Sinclair, A.G.; Ewen, M.; Bland, V.C.; Edwards, S.A. Relationships between passive absorption of immunoglobulin G by the piglet and plasma concentrations of immunoglobulin G at weaning. Livest. Prod. Sci. 2003, 81, 223–234. [Google Scholar] [CrossRef]
- Baas, T.J.; Christian, L.L.; Rothschild, M.F. Heterosis and recombination effects in Hampshire and Landrace swine: I. Maternal traits. J. Anim. Sci. 1992, 70, 89–98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Werhahn, E.; Klobasa, F.; Butler, J.E. Investigation of some factors which influence the absorption of IgG by the neonatal piglet. Vet. Immunol. Immunopathol. 1981, 2, 35–51. [Google Scholar] [CrossRef]
- Leong, W.S.; Navaratnam, N.; Stankiewicz, M.J.; Wallace, A.V.; Ward, S.; Kuhn, N.J. Subcellular compartmentation in the synthesis of the milk sugars lactose and α-2, 3-sialyllactose. Protoplasma 1990, 159, 144–156. [Google Scholar] [CrossRef]
- Ngo, T.T.; Quiniou, N.; Heugebaert, S.; Paboeuf, F.; Dourmad, J.Y. Effect of parity and number of suckling piglets on milk production of sows. Journees Recherche Porcine 2012, 44, 195–196. [Google Scholar]
- Zak, L.J.; Williams, I.H.; Foxcroft, G.R.; Pluske, J.R.; Cegielski, A.C.; Clowes, E.J.; Aherne, F.X. Feeding lactating primiparous sows to establish three divergent metabolic states: I. Associated endocrine changes and postweaning reproductive performance. J. Anim. Sci. 1998, 76, 1145–1153. [Google Scholar] [CrossRef] [PubMed]
- Clowes, E.J.; Williams, I.H.; Baracos, V.E.; Pluske, J.R.; Cegielski, A.C.; Zak, L.J.; Aherne, F.X. Feeding lactating primiparous sows to establish three divergent metabolic states: II. Effect on nitrogen partitioning and skeletal muscle composition. J. Anim. Sci. 1998, 76, 1154–1164. [Google Scholar] [CrossRef]
- Kemp, B.; Soede, N.M. Reproductive problems in primiparous sows. In Proceedings of the 18th IPVS Congress, Hoya, Germany, 27 June–1 July 2004; pp. 843–848. [Google Scholar]
- Hughes, P.E.; Smits, R.J.; Xie, Y.; Kirkwood, R.N. Relationships among gilt and sow live weight, P2 backfat depth, and culling rates. J. Swine Health Prod. 2010, 18, 301–305. [Google Scholar]
- Hoving, L.L.; Soede, N.M.; Graat, E.A.M.; Feitsma, H.; Kemp, B. Reproductive performance of second parity sows: relations with subsequent reproduction. Livest. Sci. 2011, 140, 124–130. [Google Scholar] [CrossRef]
- Thaker, M.Y.C.; Bilkei, G. Lactation weight loss influences subsequent reproductive performance of sows. Anim. Reprod. Sci. 2005, 88, 309–318. [Google Scholar] [CrossRef]
- Herpin, P.; Damon, M.; Le Dividich, J. Development of thermoregulation and neonatal survival in pigs. Livest. Prod. Sci. 2002, 78, 25–45. [Google Scholar] [CrossRef]
- Mahan, D.C. Relationship of gestation protein and feed intake level over a five-parity period using a high-reproducing sow genotype. J. Anim. Sci. 1998, 76, 533–541. [Google Scholar] [CrossRef] [PubMed]
- Perrin, D.R. 537. The composition of sow’s milk during the course of lactation. J. Dairy Res. 1954, 21, 55–62. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Trait | Number of Observations (n) | |
|---|---|---|
| Primiparous | Multiparous | |
| Protein (%) | 44 | 53 |
| Lactose (%) | 46 | 53 |
| Fat (%) | 44 | 54 |
| NE (MJ/kg) | 43 | 53 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Craig, J.R.; Dunshea, F.R.; Cottrell, J.J.; Wijesiriwardana, U.A.; Pluske, J.R. Primiparous and Multiparous Sows Have Largely Similar Colostrum and Milk Composition Profiles Throughout Lactation. Animals 2019, 9, 35. https://doi.org/10.3390/ani9020035
Craig JR, Dunshea FR, Cottrell JJ, Wijesiriwardana UA, Pluske JR. Primiparous and Multiparous Sows Have Largely Similar Colostrum and Milk Composition Profiles Throughout Lactation. Animals. 2019; 9(2):35. https://doi.org/10.3390/ani9020035
Chicago/Turabian StyleCraig, Jessica R., Frank R. Dunshea, Jeremy J. Cottrell, Udani A. Wijesiriwardana, and John R. Pluske. 2019. "Primiparous and Multiparous Sows Have Largely Similar Colostrum and Milk Composition Profiles Throughout Lactation" Animals 9, no. 2: 35. https://doi.org/10.3390/ani9020035