The Use of Biochemical Measurements to Identify Pre-Slaughter Stress in Pasture Finished Beef Cattle

 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
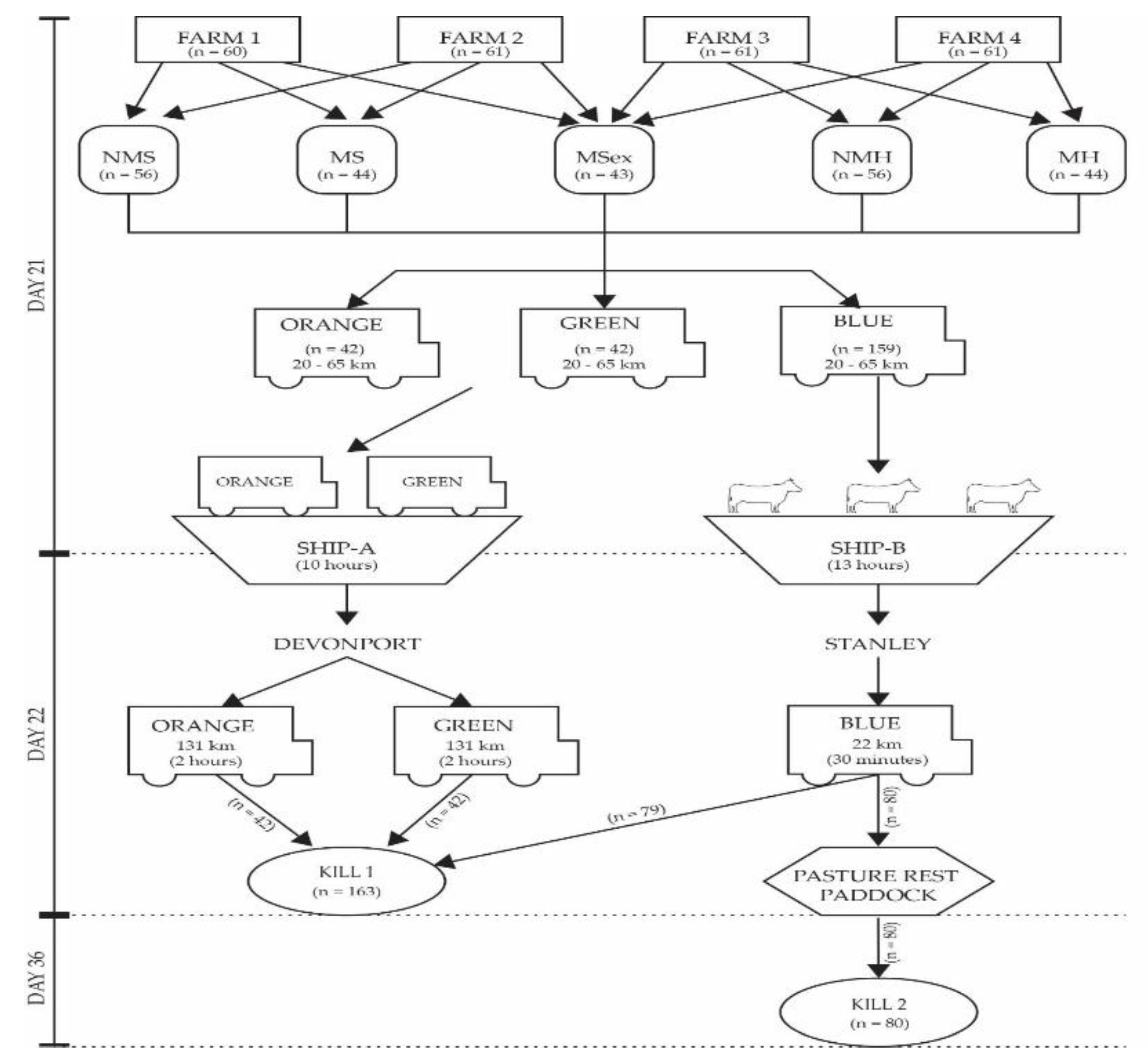
2.1. Cattle Design
2.2. Experiment One Transport methodology
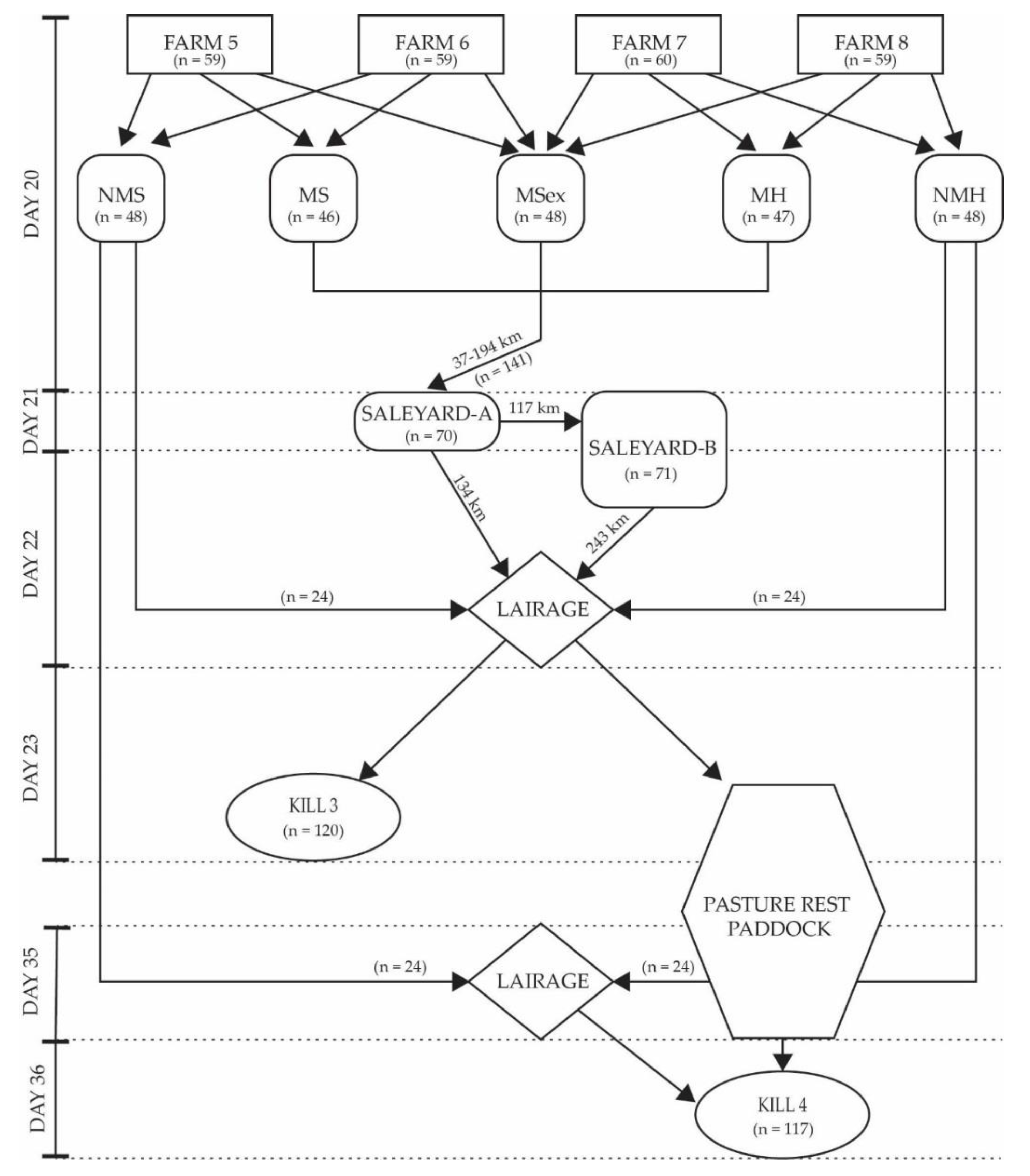
2.3. Experiment Two
2.4. Slaughter and Carcass Measurements
2.5. Blood Sampling and Plasma Analysis
2.6. Muscle Collection and Glycogen Analysis
2.7. Statistical Analysis
3. Results
3.1. Descriptive Statistics
3.2. Treatment One: Mixing
3.2.1. Experiment One
3.2.2. Experiment Two
3.3. Treatment Two: Transport Method
3.3.1. Experiment One
3.3.2. Experiment Two
3.4. Treatment Three: Slaughter Interval
3.4.1. Experiment One
3.4.2. Experiment Two
3.5. Carcass Characteristics
4. Discussion
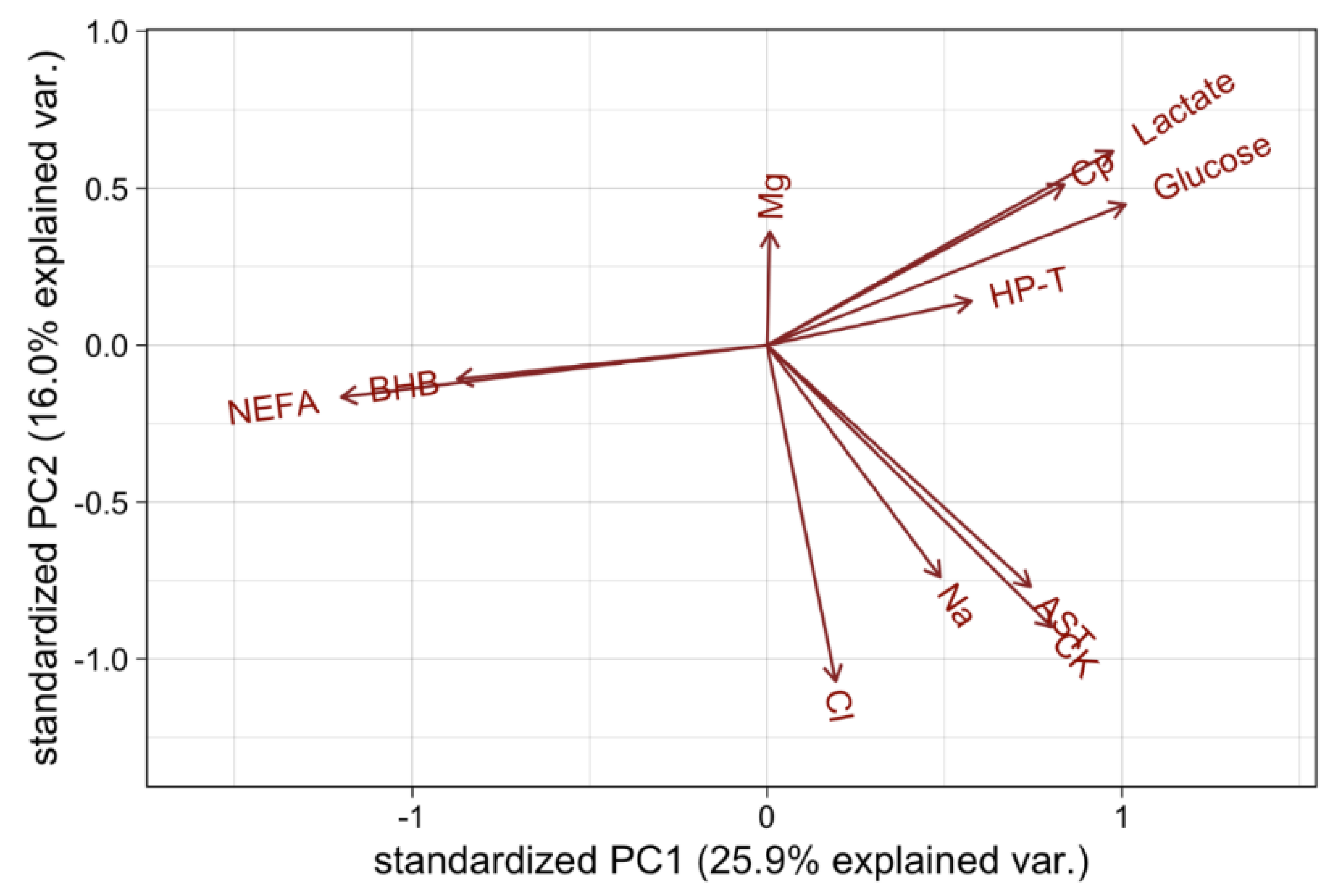
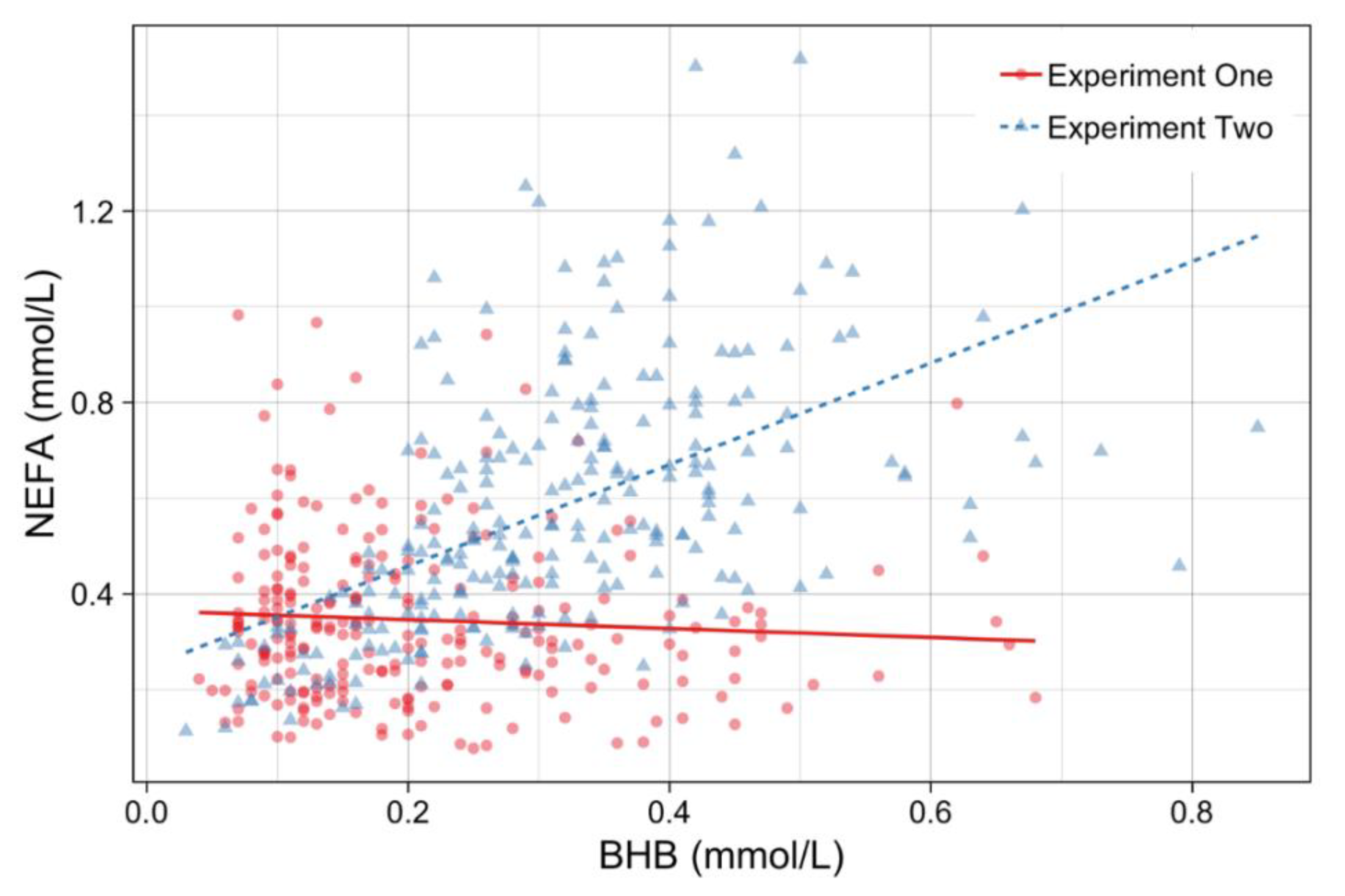
4.1. Correlations and Principle Component Analysis of Blood Metabolites
4.2. The Effect of Mixing and Transport Treatment on Blood Biomarkers
4.3. The Effect of Slaughter Interval on Plasma and Muscle Biomarkers
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Tarrant, P. Animal behaviour and environment in the dark-cutting condition in beef-a review. Ir. J. Food Sci. Technol. 1989, 13, 1–21. [Google Scholar]
- Ferguson, D.M.; Bruce, H.L.; Thompson, J.M.; Egan, A.F.; Perry, D.; Shorthose, W.R. Factors affecting beef palatability—farmgate to chilled carcass. Aust. J. Exp. Agric. 2001, 41, 879–891. [Google Scholar] [CrossRef]
- Ferguson, D.M.; Warner, D.R. Have we underestimated the impact of pre-slaughter stress on meat quality in ruminants? Meat Sci. 2008, 80, 12–19. [Google Scholar] [CrossRef] [PubMed]
- Warner, R.D.; Ferguson, D.M.; Cottrell, J.J.; Knee, B.W. Acute stress induced by the preslaughter use of electric prodders causes tougher beef meat. Aust. J. Exp. Agric. 2007, 47, 782–788. [Google Scholar] [CrossRef]
- Chrousos, G.P. Stressors, stress, and neuroendocrine integration of the adaptive response: The 1997 Hans Selye Memorial Lecture. Ann. N. Y. Acad. Sci. 1998, 851, 311–335. [Google Scholar] [CrossRef] [PubMed]
- Selye, H. The Stress of Life, 2nd ed.; McGraw-Hil: New York, NY, USA, 1956; Volume 5. [Google Scholar]
- Johnson, E.O.; Kamilaris, T.C.; Chrousos, G.P.; Gold, P.G. Mechanisms of stress: A dynamic overview of hormonal and behavioral homeostasis. Neurosci. & Biobehav. Rev. 1992, 16, 115–130. [Google Scholar]
- Chrousos, G.P.; Loriaux, D.L.; Gold, P.W. Introduction: The concept of stress and its historical development. In Mechanisms of Physical and Emotional Stress; Springer: Basel, Switerland, 1988; Volume 245, pp. 3–7. [Google Scholar]
- Moberg, G.P. Biological response to stress: Implications for animal welfare. In The Biology of Animal Stress: Basic Principles and Implications for Animal Welfare; CAB International: Wallingford, UK, 2000; Volume 1, pp. 1–21. [Google Scholar]
- Selye, H. Forty years of stress research: Principal remaining problems and misconceptions. Can. Med. Assoc. J. 1976, 115, 53. [Google Scholar]
- Pethick, D.W.; Harper, G.S.; Dunshea, F.R. Fat metabolism and turnover. In Quantitative Aspects of Ruminant Digestion and Metabolism, 2nd ed.; Dijkstra, J., Fobes, J.M., France, J., Eds.; CAB International: Wallingford, UK, 2005; Volume 13, pp. 345–372. [Google Scholar]
- Seelig, M.S. Consequences of magnesium deficiency on the enhancement of stress reactions; preventive and therapeutic implications (a review). J. Acad. Nutr. Diet. 1994, 13, 429–446. [Google Scholar] [CrossRef]
- Murck, H. Magnesium and affective disorders. Nutr. Neurosci. 2002, 5, 375–389. [Google Scholar] [CrossRef]
- Cannon, W.B. Psychological Trauma: Theory, Research, Practice, and Policy. In Bodily Changes in Pain, Hunger, Fear and Rage: An Account of Recent Research Into the Function of Emotional Excitement. Appleton-Century-Crofts. Professional Curriculum: A Call to Action; D Appleton & Company: New York, NY, USA, 1929; Volume 1, pp. 3–23. [Google Scholar]
- Kuchel, O. Stress and catecholamines. Methods. Achiev. Exp. Pathol. 1991, 14, 80. [Google Scholar]
- Rooyackers, O.E.; Nair, K.S. Hormonal regulation of human muscle protein metabolism. Annu. Rev. Nutr. 1997, 17, 457–485. [Google Scholar] [CrossRef] [PubMed]
- Pethick, D.; Rowe, J. The effect of nutrition and exercise in carcass parameters and the level of glycogen in skeletal muscle of Merino sheep. Aust. J. Agric. Res. 1996, 47, 525–537. [Google Scholar] [CrossRef]
- Gardner, G.; McIntyre, B.; Tudor, G.; Pethick, D. The impact of nutrition on bovine muscle glycogen metabolism following exercise. Aust. J. Agric. Res. 2001, 52, 461–470. [Google Scholar] [CrossRef]
- McVeigh, J.M.; Tarrant, P.V. Glycogen content and repletion rates in beef muscle, effect of feeding and fasting. J. Nutr. 1982, 112, 1306–1314. [Google Scholar] [CrossRef] [PubMed]
- Knee, B.W.; Cummins, L.J.; Walker, P.J.; Kearney, G.A.; Warner, R.D. Reducing dark-cutting in pasture-fed beef steers by high-energy supplementation. Aust. J. Exp. Agric. 2007, 47, 1277–1283. [Google Scholar] [CrossRef]
- Immonen, K.; Ruusunen, M.; Hissa, K.; Puolanne, E. Bovine muscle glycogen concentration in relation to finishing diet, slaughter and ultimate pH. Meat Sci. 2000, 55, 25–31. [Google Scholar] [CrossRef]
- Knee, B.W.; Cummins, L.J.; Walker, P.; Warner, L. Seasonal variation in muscle glycogen in beef steers. Aust. J. Exp. Agric. 2004, 44, 729–734. [Google Scholar] [CrossRef]
- MLA. Meat Standards Australia beef information kit. 2018. Available online: https://www.mla.com.au/globalassets/mla-corporate/marketing-beef-and-lamb/msa_tt_beefinfokit_jul13_lr.pdf (accessed on 24 June 2019).
- Marti, S.; Wilde, R.E.; Moya, D.; Heuston, C.E.; Brown, F.; Schwartzkopf-Genswein, K.S. Effect of rest stop duration during long-distance transport on welfare indicators in recently weaned beef calves. J. Anim. Sci. 2017, 95, 636–644. [Google Scholar] [CrossRef]
- Ferreira, G.B.; Andrade, C.L.; Costa, F.; Freitas, M.Q.; Silva, T.J.; Santos, I.F. Effects of transport time and rest period on the quality of electrically stimulated male cattle carcasses. Meat Sci. 2006, 74, 459–466. [Google Scholar] [CrossRef]
- Knowles, G.; Warriss, P.D.; Brown, S.N.; Edwards, J.E. Effects on cattle of transportation by road for up to 31 hours. Vet. Rec. 1999, 145, 575. [Google Scholar] [CrossRef]
- Loudon, K.M.W.; Lean, I.J.; Pethick, D.W.; Gardner, G.E.; Grubb, I.L.; Evans, A.C.; McGilchrist, P. On farm factors increasing dark cutting in pasture finished beef cattle. Meat Sci. 2018, 144, 110–117. [Google Scholar] [CrossRef] [PubMed]
- Eckersall, P.; Duthie, S.; Safi, S.; Moffatt, D.; Horadagoda, N.; Doyle, S.; Parton, R.; Bennett, D.; Fitzpatrick, J. An automated biochemical assay for haptoglobin: prevention of interference from albumin. Comp. Haematol. Int. 1999, 9, 117–124. [Google Scholar] [CrossRef]
- Siotto, M.; Pasqualetti, P.; Marano, M.; Squitti, R. Automation of o-dianisidine assay for ceruloplasmin activity analyses: usefulness of investigation in Wilson’s disease and in hepatic encephalopathy. J. Neural Transm. 2014, 121, 1281–1286. [Google Scholar] [CrossRef] [PubMed]
- Chan, T.M.; Exton, J.H. A rapid method for the determination of glycogen content and radioactivity in small quantities of tissue or isolated hepatocytes. Anal. Biochem. 1976, 71, 96–105. [Google Scholar] [CrossRef]
- Noll, F. L+ lactate determination. In Methods of Enzymatic Analysis; Bergmeyer, H.U., Ed.; VCH Verlagsgesellschaft: Weinheim, Germany, 1985; pp. 583–588. [Google Scholar]
- R Core Team. A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018; Available online: https://www. R-project. org (accessed on 25 July 2019).
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: Basel, Switerland, 2016. [Google Scholar]
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting linear mixed-effects models using lme4. J. Stat. Softw. 2014, 67, 1–48. [Google Scholar]
- Radostits, O.M.; Gay, C.C.; Blood, D.C.; Hinchcliff, K.W. Veterinary Medicine-a Textbook of the Diseases of Cattle, Horses, Sheep, Pigs and Goats, 9th ed.; WB Saunders Co.: Sydney, Australia, 2000. [Google Scholar]
- Eckersall, P.; Young, F.; McComb, C.; Hogarth, C.; Safi, S.; Fitzpatrick, J.; Nolan, A.; Weber, A.; McDonald, T. Acute phase proteins in serum and milk from dairy cows with clinical mastitis. Vet. Rec. 2001, 148, 35–41. [Google Scholar] [CrossRef] [PubMed]
- Trevisi, E.; Amadori, M.; Cogrossi, S.; Razzuoli, E.; Bertoni, G. Metabolic stress and inflammatory response in high-yielding, periparturient dairy cows. Res. Vet. Sci. 2012, 93, 695–704. [Google Scholar] [CrossRef]
- Laven, R.; Lawrence, K.; Livesey, C. The assessment of blood copper status in cattle: A comparison of measurements of caeruloplasmin and elemental copper in serum and plasma. N. Z. Vet. J. 2007, 55, 171–176. [Google Scholar] [CrossRef]
- Warriss, P. The handling of cattle pre-slaughter and its effects on carcass and meat quality. Appl. Anim. Behav. Sci. 1990, 28, 171–186. [Google Scholar] [CrossRef]
- Polkinghorne, R.; Philpott, J.; Thompson, J. Do extended transport times and rest periods impact on eating quality of beef carcasses? Meat Sci. 2018, 140, 101–111. [Google Scholar] [CrossRef]
- Gruber, S.; Tatum, J.; Engle, T.; Chapman, P.; Belk, K.; Smith, G. Relationships of behavioral and physiological symptoms of preslaughter stress to beef longissimus muscle tenderness 1. J. Anim. Sci. 2010, 88, 1148–1159. [Google Scholar] [CrossRef]
- Pang, D.S.; Boysen, S. Lactate in veterinary critical care: Pathophysiology and management. J. Am. Anim. Hosp. Assoc. 2007, 43, 270–279. [Google Scholar] [CrossRef] [PubMed]
- Bakker, J.; Nijsten, M.W.; Jansen, T.C. Clinical use of lactate monitoring in critically ill patients. Ann. Intensive Care 2013, 3, 12. [Google Scholar] [CrossRef] [PubMed]
- Hermansen, L.; Hultman, E.; Saltin, B. Muscle glycogen during prolonged severe exercise. Acta Physiol. Scand. 1967, 71, 129–139. [Google Scholar] [CrossRef] [PubMed]
- Harman, N.; Pethick, D. The effects of sustained exercise on gluconeogenesis, glycogenolysis and glycogen synthesis in Merino sheep. Aust. J. Agric. Res. 1994, 45, 1189–1202. [Google Scholar] [CrossRef]
- Van Soest, P.J. Nutritional Ecology of the Ruminant, 2nd ed.; Cornell university press: New York, NY, USA, 1994. [Google Scholar]
- Murray, R.K.; Granner, D.K.; Mayes, P.A.; Rodwell, V.W. Harper’s Biochemistry, 24th ed.; Appleton & Lange: Stamford, CT, USA, 1996. [Google Scholar]
- Nelson, D.L.; Lehninger, A.L.; Cox, M.M. Lehninger Principles of Biochemistry; Macmillan: New York, NY, USA, 2008. [Google Scholar]
- Dunshea, F.; King, R. Responses to homeostatic signals in ractopamine-treated pigs. Br. J. Nutr. 1995, 73, 809–818. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sechen, S.J.; Dunshea, F.R.; Bauman, D.E. Somatotropin in lactating cows: Effect on response to epinephrine and insulin. Am. J. Physiol.-Endocrinol. Metab. 1990, 258, E582–E588. [Google Scholar] [CrossRef] [PubMed]
- Ozkan, I.; Ibrahim, C.H. Dehydration, skeletal muscle damage and inflammation before the competitions among the elite wrestlers. J. phys. Ther. Sci. 2016, 28, 162–168. [Google Scholar] [CrossRef] [Green Version]
- Cooke, R.; Bohnert, D. Bovine acute-phase response after corticotrophin-release hormone challenge. J. Anim. Sci. 2011, 89, 252–257. [Google Scholar] [CrossRef]
- Blakley, B.; Hamilton, D. Ceruloplasmin as an indicator of copper status in cattle and sheep. Can. J. Comp. Med. 1985, 49, 405. [Google Scholar]
- Kietzmann, M.; Jablonski, H. On the blocking of stress by magnesium-aspartate hydrochloride in the pig. Prakt. Tierarzt (Germany, FR) 1985, 66, 328–335. [Google Scholar]
- D’souza, D.N.; Warner, R.D.; Leury, B.J.; Dunshea, F.R. The effect of dietary magnesium aspartate supplementation on pork quality. J. Anim. Sci. 1998, 76, 104–109. [Google Scholar] [CrossRef] [PubMed]
- Ryan, M.F. The role of magnesium in clinical biochemistry: An overview. Ann. Clin. Biochem. 1991, 28, 19–26. [Google Scholar] [CrossRef] [PubMed]
- Schonewille, J.T. Magnesium in dairy cow nutrition: An overview. Plant. soil. 2013, 368, 167–178. [Google Scholar] [CrossRef]
- Rayssiguier, Y. Hypomagnesemia resulting from adrenaline infusion in ewes: Its relation to lipolysis. Horm. Metab. Res. Suppl. 1977, 9, 309–314. [Google Scholar] [CrossRef] [PubMed]
- Terashima, Y.; Muzushima, Y.; Yamaoka, R.; Itoh, H. Effects of intravenous infusion of noradrenaline on plasma levels of Ca, inorganic P and Mg in sheep fed high or low calcium diet. Jpn. J. ZooTech. Sci. (Japan) 1983, 54, 52–59. [Google Scholar]
- Yano, H.; Nagaoka, C.; Kawashimai, R. Effects of intravenous glucose or epinephrine injection on plasma concentrations of potassium, magnesium and insulin in sheep. J. Nutr. Sci. Vitaminol. 1984, 30, 535–540. [Google Scholar] [PubMed]
- Whang, R. Magnesium deficiency: Pathogenesis, prevalence, and clinical implications. Am. J. Med. 1987, 82, 24–29. [Google Scholar] [CrossRef]
- Martens, H.; Schweigel, M. Pathophysiology of grass tetany and other hypomagnesemias: Implications for clinical management. Vet. Clin. Food Anim. Pract. 2000, 16, 339–368. [Google Scholar] [CrossRef]
- McGann, A.M.; Hodson, A.W. Delay in cell separation, storage and anticoagulants induced inaccuracies in measuring plasma non-esterified fatty acids. Clin. Chim. Acta 1991, 197, 265–270. [Google Scholar] [CrossRef]
- Morris, J.D.; Fernandez, J.M.; Chapa, A.M.; Gentry, L.R.; Thorn, K.E.; Weick, T.M. Effects of sample handling, processing, storage, and hemolysis on measurements of key energy metabolites in ovine blood. Small Rumin. Res. 2002, 43, 157–166. [Google Scholar] [CrossRef]
- Rogiers, V. Stability of the long chain non-esterified fatty acid pattern in plasma and blood during different storage conditions. Clin. Chim. Acta 1978, 84, 49–54. [Google Scholar] [CrossRef]
- Jones, D.G. Stability and storage characteristics of enzymes in cattle blood. Res. Vet. Sci. 1985, 38, 301–306. [Google Scholar] [CrossRef]
- Brinc, D.; Chan, M.K.; Venner, A.A.; Pasic, M.D.; Colantonio, D.; Kyriakopolou, L.; Adeli, K. Long-term stability of biochemical markers in pediatric serum specimens stored at− 80 C: A CALIPER Substudy. Clin. Biochem. 2012, 45, 816–826. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, B.; Taillade, B.; Médaille, C.; Palenché, F.; Trumel, C.; Lefebvre, H.P. Effect of repeated freeze-thaw cycles on routine plasma biochemical constituents in canine plasma. Vet. Clin. Pathol. 2006, 35, 339–340. [Google Scholar] [CrossRef] [PubMed]
- Stokol, T.; Nydam, D. Effect of anticoagulant and storage conditions on bovine nonesterified fatty acid and β-hydroxybutyrate concentrations in blood. J. Dairy Sci. 2005, 88, 3139–3144. [Google Scholar] [CrossRef]
- Stewart, S.M. The Influence of Acute Stress and Feed Deprivation on the Meat Quality and Intermediary Metabolism of Australian Lamb; Murdoch University: Perth, Australia, 2017. [Google Scholar]
- Gleeson, M. Effect of heparin and storage on human plasma free fatty acid concentration. Clin. Chim. Acta 1987, 169, 315–318. [Google Scholar] [CrossRef]
- Eckel, R.H. Lipoprotein lipase. N. Engl. J. Med. 1989, 320, 1060–1068. [Google Scholar]
- Goldberg, I.J. Lipoprotein lipase and lipolysis: Central roles in lipoprotein metabolism and atherogenesis. J. Lipid Res. 1996, 37, 693–707. [Google Scholar]
- Yang, J.Y.; Kim, T.K.; Koo, B.S.; Park, B.H.; Park, J.W. Change of plasma lipoproteins by heparin-released lipoprotein lipase. Exp. Mol. Med. 1999, 31, 60. [Google Scholar] [CrossRef]
- Menéndez, L.G.; Fernández, A.L.; Enguix, A.; Ciriza, C.; Amador, J. Effect of storage of plasma and serum on enzymatic determination of non-esterified fatty acids. Ann. Clin. Biochem. 2001, 38, 252–255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeevanandam, M.; Hsu, Y.C.; Ramias, L.; Schiller, W.R. A rapid, automated micromethod for measuring free fatty acids in plasma/serum. Clin. Chem. 1989, 35, 2228–2231. [Google Scholar] [PubMed]
- Warriss, P.D.; Kestin, S.C.; Brown, S.N.; Wilkins, L.J. The time required for recovery from mixing stress in young bulls and the prevention of dark cutting beef. Meat Sci. 1984, 10, 53–68. [Google Scholar] [CrossRef]
- Kenny, F.J.; Tarrant, P.V. The behaviour of young Friesian bulls during social re-grouping at an abattoir. Influence of an overhead electrified wire grid. Appl. Anim. Behav. Sci. 1987, 18, 233–246. [Google Scholar] [CrossRef]
- Kenny, F.J.; Tarrant, P.V. The reaction of young bulls to short-haul road transport. Appl. Anim. Behav. Sci. 1987, 17, 209–227. [Google Scholar] [CrossRef]
- Buckham Sporer, K.R.; Weber, P.S.D.; Burton, J.L.; Earley, B.; Crowe, M.A. Transportation of young beef bulls alters circulating physiological parameters that may be effective biomarkers of stress. J. Anim. Sci 2008, 86, 1325–1334. [Google Scholar] [CrossRef] [PubMed]
- Tarrant, P.V.; Kenny, F.J.; Harrington, D.; Murphy, M. Long distance transportation of steers to slaughter: Effect of stocking density on physiology, behaviour and carcass quality. Livest. Prod. Sci. 1992, 30, 223–238. [Google Scholar] [CrossRef]
- Parkinson, T.; Vermunt, J.; Malmo, J. Diseases of Cattle in Australasia: A Comprehensive Textbook; New Zealand Veterinary Association Foundation for Continuing Education: Wellington, New Zealand, 2010. [Google Scholar]
- McVeigh, J.M.; Tarrant, P.V.; Harrington, M.G. Behavioral stress and skeletal muscle glycogen metabolism in young bulls. J. Anim. Sci. 1982, 54, 790–795. [Google Scholar] [CrossRef]
- Moberg, G.P. Animal stress. In Biological Response to Stress: Key to Assessment of Animal Well-Being? Springer: Basel, Switerland, 1985; pp. 27–49. [Google Scholar]
- Reineke, E.L. Overview of Diabetes Treatment in Animals. In Nutritional and Therapeutic Interventions for Diabetes and Metabolic Syndrome; Elsevier: San Diego, CA, USA, 2012; pp. 499–511. [Google Scholar]
- Mudroň, P.; Rehage, J.; Sallmann, H.P.; Höltershinken, M.; Scholz, H. Stress Response in Dairy Cows Related to Different Blood Glucose. Acta Vet. Brno 2005, 74, 37–42. [Google Scholar] [CrossRef] [Green Version]
- Vincent, J.L.; Dufaye, P.; Berré, J.; Leeman, M.; Degaute, J.P.; Kahn, R.J. Serial lactate determinations during circulatory shock. Crit. Care Med. 1983, 11, 449–451. [Google Scholar] [CrossRef]
- Stockman, C.A.; Collins, T.; Barnes, A.L.; Miller, D.; Wickham, S.L.; Beatty, D.T.; Blache, D.; Wemelsfelder, F.W.; Fleming, P.A. Qualitative behavioural assessment and quantitative physiological measurement of cattle naïve and habituated to road transport. Anim. Prod. Sci. 2011, 51, 240–249. [Google Scholar] [CrossRef]
- Warner, R.D.; Dunshea, F.R.; Gutzke, D.; Lau, J.; Kearney, G. Factors influencing the incidence of high rigor temperature in beef carcasses in Australia. Anim. Prod. Sci. 2014, 54, 363–374. [Google Scholar] [CrossRef] [Green Version]
- Gardner, G.E.; McIntyre, B.L.; Tudor, G.D.; Pethick, D.W. Nutritional influences on muscle glycogen recovery following exercise in sheep and cattle. Recent Adv. Ani. Nutr. Aust. 2001, 13, 145–151. [Google Scholar]
- Guyton, A.C. Textbook of Medical Physiology, 8th ed.; W.B. Saunders Company: Philadelphia, PA, USA, 1991. [Google Scholar]
- Thrall, M.A.; Weiser, G.; Allison, R.; Campbell, T. Veterinary Hematology and Clinical Chemistry; John Wiley & Sons: Toronto, ON, Canada, 2012. [Google Scholar]
- Cray, C.; Zaias, J.; Altman, N.H. Acute phase response in animals: A review. Com. Med. 2009, 59, 517–526. [Google Scholar]
- Alsemgeest, S.P.M.; Lambooy, I.E.; Wierenga, H.K.; Dieleman, S.J.B.; Meerkerk, B.; van Ederen, A.M.; Niewold, T.A. Influence of physical stress on the plasma concentration of serum amyloid-a (SAA) and haptoglobin (HP) in calves. Vet. Q. 1995, 17, 9–12. [Google Scholar] [CrossRef] [PubMed]
- Arthington, J.D.; Eicher, S.D.; Kunkle, W.E.; Martin, F.G. Effect of transportation and commingling on the acute-phase protein response, growth, and feed intake of newly weaned beef calves. J. Anim. Sci. 2003, 81, 1120–1125. [Google Scholar] [CrossRef]
- Karki, N. The urinary excretion of noradrenaline and adrenaline in different age groups, its diurnal variation and the effect of muscular work on it. Acta physiol. Scand. Suppl. 1956, 39, 1–96. [Google Scholar]
- McVeigh, J.M.; Tarrant, P.V. Effect of propranolol on muscle glycogen metabolism during social regrouping of young bulls. J. Anim. Sci. 1983, 56, 71–80. [Google Scholar] [CrossRef]
- Young, B.A. Some physiological costs of cold climates. 1975. Available online: http://agris.fao.org/agris-search/search.do?recordID=US19770190577 (accessed on 25 July 2019).
- Young, B. Cold Stress as it Affects Animal Production 1. J. Anim. Sci. 1981, 52, 154–163. [Google Scholar] [CrossRef]
- Haman, F.; Péronnet, F.; Kenny, G.P.; Massicotte, D.; Lavoie, C.; Weber, J.M. Partitioning oxidative fuels during cold exposure in humans: Muscle glycogen becomes dominant as shivering intensifies. J. Physiol. 2005, 566, 247–256. [Google Scholar] [CrossRef]
- Pethick, D.W.; Cummins, G.; Gardner, G.E.; Knee, B.W.; McDowell, M.; McIntyre, B.L.; Tudor, G.; Walker, P.G.; Warner, W.D. The regulation by nutrition of glycogen level in the muscle of ruminants. Recent Adv. Anim. Nutr. Aust. 1999, 12, 145–151. [Google Scholar]
- Cole, N.A.; Hutcheson, D.P. Influence of prefast feed intake on recovery from feed and water deprivation by beef steers. J. Anim. Sci. 1985, 60, 772–780. [Google Scholar] [CrossRef] [PubMed]
- Rett, B.; Marques, R.S.; Brandão, A.P.; de Sousa, O.A.; Schumaher, T.F.; Cooke, R.F.; Bohnert, D.W. Impact of 24-h feed or water, or both, deprivation on feed intake, metabolic, and inflammatory response in beef heifers. Transl. Anim. Sci. 2018, 2, S95. [Google Scholar] [CrossRef]
- Carroll, J.A.; Reuter, R.R.; Chase, C.C., Jr.; Coleman, S.W.; Riley, D.G.; Spiers, D.E.; Arthington, J.D.; Galyean, M.L. Profile of the bovine acute-phase response following an intravenous bolus-dose lipopolysaccharide challenge. Innate Immun. 2009, 15, 81–89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phillips, W.A.; Juniewicz, P.E. Von Tungeln, D.L. The effect of fasting, transit plus fasting and administration of adrenocorticotropic hormone on the source and amount of weight lost by feeder steers of different ages. J. Anim. Sci. 1991, 69, 2342–2348. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Aschenbach, J.R.; Barreda, D.R.; Penner, G.R. Short-term feed restriction impairs the absorptive function of the reticulo-rumen and total tract barrier function in beef cattle. J. Anim. Sci. 2013, 91, 1685–1695. [Google Scholar] [CrossRef]
- Zhang, S.; Aschenbach, J.R.; Barreda, D.R.; Penner, G.R. Recovery of absorptive function of the reticulo-rumen and total tract barrier function in beef cattle after short-term feed restriction. J. Anim. Sci. 2013, 91, 1696–1706. [Google Scholar] [CrossRef]
- Meltzer, H.Y. Plasma creatine phosphokinase activity, hypothermia, and stress. Am. J. Physiol. Leg. Content 1971, 221, 896–901. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Farm Number | Ship | Not Rested (Kill 1) | Rested 14days (Kill 2) | Total Number of Cattle | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| NMS | NMH | MS | MH | MSex | Total | NMS | NMH | MS | MH | MSex | Total | |||
| Farm 1 | A Orange truck | 6 a | 3 m | 2 s | 11 | 0 | 21 | |||||||
| A Green truck | 6 b | 3 n | 1 t | 10 | 0 | |||||||||
| B Blue truck | 8 c | 8 o | 3 u | 19 | 8 c | 8 v | 4 x | 20 | 39 | |||||
| Farm 2 | A Orange truck | 6 d | 3 m | 1 s | 10 | 0 | 21 | |||||||
| A Green truck | 6e | 3 n | 2 t | 11 | 0 | |||||||||
| B Blue truck | 8 f | 8 o | 4 u | 20 | 8 f | 8 v | 4 x | 20 | 40 | |||||
| Farm 3 | A Orange truck | 6 g | 3 p | 2 s | 11 | 0 | 21 | |||||||
| A Green truck | 6 h | 3 q | 1 t | 10 | ||||||||||
| B Blue truck | 8 i | 8 r | 4 u | 20 | 8 i | 8 w | 4 x | 20 | 40 | |||||
| Farm 4 | A Orange truck | 6 j | 3 p | 1 s | 10 | 21 | ||||||||
| A Green truck | 6 k | 3 q | 2 t | 11 | ||||||||||
| B Blue truck | 8 l | 8 r | 4 u | 20 | 8 l | 8 w | 4 x | 20 | 40 | |||||
| Total Ship-A | 24 | 24 | 12 | 12 | 12 | 84 | 0 | 0 | 0 | 0 | 0 | 0 | 84 | |
| Total Ship-B | 16 | 16 | 16 | 16 | 15 | 79 | 16 | 16 | 16 | 16 | 16 | 80 | 159 | |
| Total King Island | 40 | 40 | 28 | 28 | 27 | 163 | 16 | 16 | 16 | 16 | 16 | 80 | 243 | |
| Time from point of muster to slaughter | 28 hours | 1 hour | ||||||||||||
| Farm Number | Direct/Saleyard | Not Rested | Rested 14 Days | Total Head | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| NMS | NMH | MS | MH | MSex | Total | NMS | NMH | MS | MH | MSex | Total | |||
| Farm 5 | Direct | 6 a 6 b | 12 | 6 w 6 x | 12 | 24 | ||||||||
| Saleyard-A | 3 i 3 j | 2 q 1 r | 9 | 2 ee 3 ff | 1 nn 2 oo | 8 | 17 | |||||||
| Saleyard-B | 3 k 3 l | 2 s 1 t | 9 | 3 hh 3 ii | 1 pp 2 qq | 9 | 18 | |||||||
| Farm 6 | Direct | 6 c 6 d | 12 | 6 y 6 z | 12 | 24 | ||||||||
| Saleyard-A | 3 i 3 j | 1 q 2 r | 9 | 3 ee 2 ff | 2 nn 1 oo | 8 | 17 | |||||||
| Saleyard-B | 3 k 3 l | 1 s 2 t | 9 | 3 hh 3 ii | 2 pp 1 qq | 9 | 18 | |||||||
| Farm 7 | Direct | 6 e6 f | 12 | 6 aa 6 bb | 12 | 24 | ||||||||
| Saleyard-A | 3 m 3 n | 2 q 1 r | 9 | 3 jj 3 kk | 1 nn 2 oo | 9 | 18 | |||||||
| Saleyard-B | 3 o 3 p | 2 s 1 t | 9 | 3 ll 3 mm | 1 pp 2 qq | 9 | 18 | |||||||
| Farm 8 | Direct | 6 g 6 h | 12 | 6 cc 6 dd | 12 | 24 | ||||||||
| Saleyard-A | 3 m 3 n | 1 q 2 r | 9 | 3 jj 3 kk | 2 nn 1 oo | 9 | 18 | |||||||
| Saleyard-B | 3 o 3 p | 1 s 2 t | 9 | 3 ll 2 mm | 2 pp 1 qq | 8 | 17 | |||||||
| Total Direct | 24 | 24 | 48 | 24 | 24 | 48 | 96 | |||||||
| Total Saleyard-A | 12 | 12 | 12 | 36 | 10 | 12 | 12 | 34 | 70 | |||||
| Total Saleyard-B | 12 | 12 | 12 | 36 | 12 | 11 | 12 | 35 | 71 | |||||
| Grand Total | 24 | 24 | 24 | 24 | 24 | 120 | 24 | 24 | 22 | 23 | 24 | 117 | 237 | |
| Time from point of muster to slaughter | 76 hours | 1 hour | ||||||||||||
| Variable | Experiment One (n = 243) | Experiment Two (n = 240) | ||||||
|---|---|---|---|---|---|---|---|---|
| Mean | SD | Min | Max | Mean | SD | Min | Max | |
| Carcass weight (kg) | 270.7 | 20.8 | 206.8 | 322.0 | 265.3 | 55.3 | 188.2 | 425.8 |
| P8 (mm) | 7.7 | 2.0 | 5.0 | 15.0 | 7.5 | 2.7 | 2.0 | 20.0 |
| EMA (cm2) | 70.0 | 8.3 | 50.0 | 96.0 | 68.7 | 10.1 | 50.0 | 140.0 |
| Ribfat (mm) | 6.5 | 2.6 | 2.0 | 14.0 | 5.7 | 3.3 | 1.0 | 21.0 |
| Ossification score | 166.7 | 26.4 | 120.0 | 230.0 | 157.8 | 25.5 | 100.0 | 230.0 |
| MSA Marble Score | 345.6 | 67.0 | 170.0 | 740.0 | 300.5 | 70.1 | 130.0 | 520.0 |
| Location | Slaughter Group | pH > 5.7 | Ribfat < 3mm |
|---|---|---|---|
| Experiment One (direct slaughter) | Kill 1 (n = 163) | 64 | 3 |
| Experiment One (rested) | Kill 2 (n = 80) | 3 | 4 |
| Experiment Two (direct slaughter) | Kill 3 (n = 120) | 20 | 14 |
| Experiment Two (rested) | Kill 4 (n = 120) | 26 | 18 |
| Location | Variable | Mean | SD | Min | Max | Published Normal Basal Concentrations |
|---|---|---|---|---|---|---|
| Experiment One | Glucose (mmol/L) | 7.42 | 1.22 | 3.69 | 14.51 | 1.9–8 a |
| Experiment Two | 6.23 | 0.89 | 4.66 | 10.34 | ||
| Experiment One | Lactate (mmol/L) | 14.48 | 3.30 | 7.15 | 23.48 | 0.6–2.2 a |
| Experiment Two | 12.01 | 2.49 | 5.78 | 21.26 | ||
| Experiment One | NEFA (mmol/L) | 0.35 | 0.17 | 0.08 | 0.98 | <0.4 a |
| Experiment Two | 0.58 | 0.27 | 0.11 | 1.52 | ||
| Experiment One | βHB (mmol/L) | 0.21 | 0.13 | 0.04 | 0.68 | 0.35–0.47 a |
| Experiment Two | 0.31 | 0.14 | 0.03 | 0.85 | ||
| Experiment One | Magnesium (mmol/L) | 0.79 | 0.10 | 0.52 | 1.10 | 0.74–1.10 a |
| Experiment Two | 0.78 | 0.11 | 0.48 | 1.05 | ||
| Experiment One | CK (IU/L) | 1137.86 | 1346.48 | 113.30 | 9384.90 | 35–280 a |
| Experiment Two | 596.79 | 509.82 | 144.4 | 4791.00 | ||
| Experiment One | AST (IU/L) | 113.28 | 71.14 | 58.36 | 636.17 | 78–132 a |
| Experiment Two | 110.66 | 48.58 | 55.57 | 440.57 | ||
| Experiment One | Sodium (mmol/L) | 147.72 | 3.39 | 125.70 | 160.80 | 132–152 a |
| Experiment Two | 147.99 | 3.51 | 128.40 | 155.80 | ||
| Experiment One | Chloride (mmol/L) | 97.85 | 2.86 | 91.50 | 113.60 | 95–110 a |
| Experiment Two | 98.07 | 3.74 | 91.40 | 119.50 | ||
| Experiment One | Haptoglobin (mg/mL) | 0.35 | 0.30 | 0.03 | 1.80 | 0.0–0.2 b |
| Experiment Two | 0.26 | 0.49 | 0.01 | 3.24 | ||
| Experiment One | Ceruloplasmin (IU/L) | 112.02 | 22.17 | 50.00 | 202.00 | 15–68 c |
| Experiment Two | 79.96 | 40.34 | 20.00 | 258.00 | ||
| Experiment One | Muscle Glycogen (g/100g) | 0.95 | 0.20 | 0.40 | 1.56 | |
| Experiment Two | 1.18 | 0.22 | 0.31 | 1.78 |
| Bio-marker | Glucose | Lactate | NEFA | Mg | βHB | Cl | Na | HP-T | CK | AST | Cp | Glycogen | pHu |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Glucose | 1.00 | 0.50 | −0.47 | 0.04 | −0.21 | −0.02 | −0.01 | 0.18 | 0.20 | 0.15 | 0.32 | −0.34 | 0.13 |
| Lactate | 1.00 | −0.46 | 0.11 | −0.24 | −0.22 | 0.27 | 0.21 | 0.01 | 0.06 | 0.29 | −0.17 | 0.02 | |
| NEFA | 1.00 | 0.03 | 0.42 | −0.09 | −0.17 | −0.25 | −0.24 | −0.19 | −0.38 | 0.20 | −0.07 | ||
| Mg | 1.00 | 0.08 | 0.00 | 0.01 | −0.11 | −0.07 | −0.09 | 0.15 | 0.02 | −0.07 | |||
| BHB | 1.00 | −0.02 | −0.07 | −0.12 | −0.19 | −0.18 | −0.31 | 0.22 | −0.15 | ||||
| Cl | 1.00 | 0.50 | 0.00 | 0.25 | 0.09 | −0.03 | −0.11 | 0.07 | |||||
| Na | 1.00 | 0.08 | 0.11 | 0.03 | −0.08 | 0.04 | −0.02 | ||||||
| HP−T | 1.00 | 0.05 | 0.13 | 0.17 | −0.09 | −0.04 | |||||||
| CK | 1.00 | 0.70 | 0.14 | −0.52 | 0.42 | ||||||||
| AST | 1.00 | 0.11 | −0.40 | 0.36 | |||||||||
| Cp | 1.00 | −0.22 | 0.18 | ||||||||||
| Glycogen | 1.00 | −0.54 |
| Biomarker | L lactate | Glucose | CK | AST | βHB | NEFA | Mg | Na | Cl | HP-T | Cp | Glycogen | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Experiment number | 1 | 2 | 1 | 2 | 1 | 2 | 1 | 2 | 1 | 2 | 1 | 2 | 1 | 2 | 1 | 2 | 1 | 2 | 1 | 2 | 1 | 2 | 1 | 2 |
| Treatment 1 (Mixing) | 0.706 | 0.394 | 0.112 | 0.012 | 0.222 | 0.168 | 0.005 | 0.771 | 0.820 | 0.506 | 0.977 | 0.606 | ||||||||||||
| Treatment 2 (Transport Method) | 0.889 | 0.462 | 0.987 | 0.139 | 0.629 | 0.266 | 0.879 | 0.397 | 0.001 | 0.010 | 0.259 | 0.002 | 0.394 | 0.263 | 0.093 | 0.107 | 0.005 | 0.697 | 0.001 | 0.789 | 0.178 | 0.551 | 0.495 | 0.379 |
| Treatment 3 (Slaughter Interval) | < 0.001 | 0.565 | 0.852 | 0.170 | < 0.001 | 0.002 | 0.001 | 0.781 | 0.061 | < 0.001 | 0.233 | 0.627 | 0.211 | 0.145 | 0.235 | 0.754 | < 0.001 | 0.601 | < 0.001 | 0.007 | 0.643 | 0.547 | < 0.001 | 0.305 |
| pHu | 0.120 | 0.488 | 0.886 | 0.316 | <0.001 | < 0.001 | < 0.001 | < 0.001 | 0.417 | 0.737 | 0.615 | 0.453 | 0.273 | 0.317 | 0.671 | 0.749 | 0.539 | 0.777 | 0.944 | 0.419 | 0.153 | 0.607 | < 0.001 | < 0.001 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Loudon, K.M.W.; Tarr, G.; Pethick, D.W.; Lean, I.J.; Polkinghorne, R.; Mason, M.; Dunshea, F.R.; Gardner, G.E.; McGilchrist, P. The Use of Biochemical Measurements to Identify Pre-Slaughter Stress in Pasture Finished Beef Cattle. Animals 2019, 9, 503. https://doi.org/10.3390/ani9080503
Loudon KMW, Tarr G, Pethick DW, Lean IJ, Polkinghorne R, Mason M, Dunshea FR, Gardner GE, McGilchrist P. The Use of Biochemical Measurements to Identify Pre-Slaughter Stress in Pasture Finished Beef Cattle. Animals. 2019; 9(8):503. https://doi.org/10.3390/ani9080503
Chicago/Turabian StyleLoudon, Kate M.W., Garth Tarr, David W. Pethick, Ian J. Lean, Rod Polkinghorne, Maddison Mason, Frank R. Dunshea, Graham E. Gardner, and Peter McGilchrist. 2019. "The Use of Biochemical Measurements to Identify Pre-Slaughter Stress in Pasture Finished Beef Cattle" Animals 9, no. 8: 503. https://doi.org/10.3390/ani9080503