
Nanozybiotics: Nanozyme-Based Antibacterials against Bacterial Resistance


1
CAS Engineering Laboratory for Nanozyme, Institute of Biophysics, Chinese Academy of Sciences, Beijing 100101, China
2
College of Life Sciences, Graduate School of University of Chinese Academy of Sciences, Beijing 100049, China
3
Nanozyme Medical Center, School of Basic Medical Sciences, Zhengzhou University, Zhengzhou 450001, China
*
Author to whom correspondence should be addressed.
Antibiotics 2022, 11(3), 390; https://doi.org/10.3390/antibiotics11030390
Submission received: 8 February 2022
/
Revised: 4 March 2022
/
Accepted: 7 March 2022
/
Published: 15 March 2022
(This article belongs to the Special Issue Alternative Approaches to Treating Antimicrobial Resistant Infections - 2nd Volume)
Abstract
:Infectious diseases caused by bacteria represent a global threat to human health. However, due to the abuse of antibiotics, drug-resistant bacteria have evolved rapidly and led to the failure of antibiotics treatment. Alternative antimicrobial strategies different to traditional antibiotics are urgently needed. Enzyme-based antibacterials (Enzybiotics) have gradually attracted interest owing to their advantages including high specificity, rapid mode-of-action, no resistance development, etc. However, due to their low stability, potential immunogenicity, and high cost of natural enzymes, enzybiotics have limitations in practical antibacterial therapy. In recent years, many nanomaterials with enzyme-like activities (Nanozymes) have been discovered as a new generation of artificial enzymes and perform catalytic antibacterial effects against bacterial resistance. To highlight the progress in this field of nanozyme-based antibacterials (Nanozybiotics), this review discussed the antibacterial mechanism of action of nanozybiotics with a comparison with enzybiotics. We propose that nanozybiotics may bear promising applications in antibacterial therapy, due to their high stability, rapid bacterial killing, biofilm elimination, and low cost.
1. Introduction
The diseases caused by bacterial infection represent a serious threat to human health. Antibiotics, as common chemical reagents against bacteria, have been widely used in human patients. However, the resistance to antibiotics is a growing and serious problem, with bacterial infection returning as a potential global threat. According to the Centers for Disease Control and Prevention (CDC), drug-resistant bacteria causes nearly 23,000 deaths per year on average in the United States [1]. The emergence of bacterial resistance is the result of its own multiple drug-resistant mechanisms, which greatly increases the difficulty of providing effective antibacterials and reduces the therapeutic effect of antibiotics. Therefore, it is necessary to develop new antimicrobial strategies while avoiding bacterial resistance.
In recent years, researchers have been devoted to discovering and developing new drug targets and antimicrobial methods. Common antimicrobial strategies include antimicrobial peptides, bacteriophage therapy, antimicrobial enzymes and so on. Antimicrobial peptides (AMPs) are generally polycationic amphiphilic peptides with less than 50 amino acids found in a wide variety of life forms [2]. The cationic AMPs display bactericidal action by disrupting the bacterial anionic cell membrane and by binding to intercellular targets, causing bacteria death [3]. Though AMPs are active against a broad spectrum of bacteria, the clinical translation of AMPs is limited by high manufacturing costs, unfavorable pharmacokinetics and cytotoxicity in vivo [4,5]. Bacteriophages (phages) are viruses that infect bacteria and propagate in their bacterial host, resulting in cell lysis. Some phages also produce depolymerases that hydrolyze biofilm extracellular polymers. Phage therapy shows great efficiency in lysing specific bacteria and degrading the biofilm matrix [6]. However, treatment of bacterial infections with phages bears some disadvantages, including narrow host range, bacterial resistance to phages and inactivation by the immune system [6,7]. Moreover, the phage cocktails strategy is limited by complicated procedures and strong individual specificity [8].
However, enzymatic therapy can overcome almost all obstacles mentioned above. Enzymatic therapy is a treatment that uses materials with catalytic activity to catalyze critical biochemical reactions, fulfilling therapeutic effects in vivo to treat various complications [9]. Enzymatic methods mainly include the application of natural enzyme-based antibacterials. Some natural enzymes, such as lysozyme or deoxyribonuclease, can hydrolyze the cell structure of bacteria and thus exhibit excellent antibacterial performance as enzymes-based antibacterials (Enzybiotics). They are highly specific, effective, and fast-acting. However, the proteinaceous character of enzybiotics results in low stability and potential immunogenicity, which limits their practical applications [10]. To overcome the limitations of proteinaceous enzymes, artificial enzyme mimics may represent a better choice. Recently, nanozymes, a kind of nanomaterials or nanostructures with enzyme-like activity, have attracted much attention as a new generation of enzyme mimics [11]. It has been reported that nanozymes with peroxidase-like or oxidase-like activity could produce a great number of reactive oxygen species (ROS) to fight bacteria [12,13,14,15]. Furthermore, nanozymes with DNase-like activity also prevent spread of drug resistance by degradation of bacterial drug resistance genes [16,17]. Typically, compared with natural enzymes, nanozymes show the characteristics of being more stable, economical, and practical [11]. Therefore, nanozyme-based antibacterial alternatives (Nanozybiotics) may have greater application potential in the antibacterial field.
Herein, to understand and develop antibacterial strategies, we review the common mechanisms of bacterial drug resistance. Subsequently, the mechanisms and applications of enzybiotics and nanozybiotics as antibacterial agents, classified according to enzymatic activity, are summarized, including the combined strategies of nanozybiotics through enzymatic activities and specific physicochemical properties (e.g., photothermic or photodynamic therapy). We hope that this review can provide ideas for the development of a practical enzymatic therapy to combat the challenges of drug resistance in bacterial infections.
2. Non-Antibiotic Strategy Is Needed to Fight against Rapid Evolution of Bacterial Resistance
Antibiotics are considered one of the greatest achievements in the history of medicine, laying the foundation for the prevention and treatment of bacterial infection [18]. However, bacteria have also developed a variety of drug-resistant mechanisms to respond to them [19,20] (Figure 1). In order to maintain long-term effective antibacterial drugs, preventing the formation of super resistant bacteria, broadening the development avenues of antibacterial drugs, and a profound analysis of bacteria resistant mechanisms is necessary. Thus, here we briefly introduce the main mechanisms of bacterial drug resistance, including genetic, biochemical, and metabolic mechanisms.
Genetic mechanisms enact drug resistance through gene mutations leading to a change of drug target in bacteria which can survive treatment with antibiotics [19]. Moreover, other strain bacteria can acquire these drug-resistant genes through vertical transmission of chromosomes, horizontal transmission of plasmids or transposons or acquisition of exogenous drug-resistant genes by integrons [21]. Biochemical mechanisms are a more common style of antidrug. It mainly resists antibiotic treatment by target site mutation, antibiotic inactivation, impaired permeability of antibiotics, and efflux of antibiotics [22]. In addition, bacteria within a biofilm are a great challenge in the treatment of bacterial infection. Biofilms are bacterial self-generated protective communities, with an extracellular polymeric substance (EPS) matrix mainly composed of extracellular bacterial DNA (eDNA), exopolysaccharides, proteins and enzymes [23], and protects bacteria from attack by extraneous antibiotics. Besides, with the in-depth research on the mechanism of bacterial drug resistance, it has been noticed that the change in bacterial metabolic pathways could also affect susceptibility to antibiotics. To overcome drug resistance of bacteria, non-antibiotic strategies are needed to avoid the above issues.
3. Enzybiotics Are Catalytic Antibacterials Based on Enzymes against Drug-Resistant Bacteria
The overuse and misuse of traditional antibiotics have led to the emergence of drug resistance worldwide. Hitherto, there is no current development of new antibiotic classes by pharmaceutical companies. Thus, it is urgent to develop non-antibiotic alternatives to fight against drug-resistant bacteria. Recently, enzybiotics represent a promising class of antibiotics alternatives. Enzybiotics are derived from natural enzymes with antibacterial performance, usually working by degrading the bacterial cell structure. They are characterized by a rapid and unique mode of action, a high specificity of killing pathogens, and a low probability for developing new bacterial resistance [10].
Enzybiotics with antibacterial effects are mainly classified into three categories: peptidoglycan hydrolases, proteases, and nuclease (antibacterial mechanisms shown in Figure 2). Firstly, peptidoglycan hydrolase is a general term for many kinds of enzymes which catalyze the hydrolysis of peptidoglycan, a major component of Gram-positive bacterial cell wall, and can be obtained from various sources, for example, bacteriophages (lysins or endolysins) and bacteria (bacteriocins and autolysins) [24]. The modular structure of lysins contains an N-terminal enzymatically active domain (EAD) connected via a flexible linker sequence to a C-terminal cell wall binding domain (CBD). The EAD is evolutionarily conserved and responsible for the peptidoglycan lytic activity of the enzyme [25]. As a bacteriocin whose antibacterial activity has been studied most thoroughly both in vitro and in vivo, lysostaphin eradicates planktonic and quiescent bacteria as well as inhibiting Staphylococcus aureus (S. aureus) growth in the biofilms as a result of hydrolyzing the penta-glycine cross bridges of S. aureus peptidoglycan. In another words, lysostaphin kills both dividing, non-dividing and encapsulated and unencapsulated S. aureus strains [26,27]. In addition, lysozymes are also an example of peptidoglycan hydrolase-based enzybiotics from hen egg and human. Though lysozymes exert poor killing effects on Gram-negative bacteria due to the thin layer of peptidoglycan protected by outer lipid membranes (Figure 2a) [28,29], additional modification can improve it [30,31]. Secondly, proteases are a kind of enzymes present in all forms of life, and degrade proteins by catalyzing the hydrolytic cleavage of the peptide chain. The major function of proteases in living organisms is the cleavage of proteins resulting in the degradation of damaged, misfolded, and potentially harmful proteins and therefore providing the cell with amino acids essential for the synthesis of new proteins. Recently, the pharmaceutical application of proteases was concerned. The purified protease from bacteria could be used for various purposes such as antibacterial activity against clinical pathogens as well as degrading slime and biofilms to limit bacteria. One of the most widely used proteases is subtilisin which is derived from Bacillus species. Subtilisin is a non-specific serine protease that provides the preferred cleavage on the carboxyl side of hydrophobic amino acid residues [32]. In addition, several plant-derived proteases possess antibacterial activity. For example, papain and bromelain showed bactericidal and effective inhibitory activity against a variety of pathogens [33,34]. Thirdly, nuclease, as the name suggests, are proteins that exhibit the enzymatic activity of degrading nucleic acids. They are very common and play an important role in biological activities. Moreover, the potential application of nuclease has also been considered in the antibacterial and antibiofilm fields. For example, Eller et al. devised a strategy that showed combination of RNase 1 and a secretory peptide, LL-37, displayed extraordinary antimicrobial activity [35]. Banu et al. investigated the anti-biofilm effect of marine bacterial DNase (MBD) by targeting the eDNA present in the biofilms of Candida spp. [36].
Enzybiotics show highly antibacterial properties owing to its rapid and unique mode of action and specificity of killing pathogens. Considering the working features of enzybiotics that destroy bacterial cell structures, enzybiotics not only kill metabolically active bacteria, but also those in a dormant state, even when embedded in biofilm (Figure 3). Moreover, this feature also ensures enzybiotics bear a low probability to develop bacterial resistance. These advantages indicate that enzybiotics are very ideal antibacterial agents. However, enzybiotics have a short half-life and raise neutralizing antibodies in vivo because of their proteinaceous nature, which seriously limits their clinical transformation. Moreover, the widespread application of enzybiotics in industrial processes is considered restricted due to poor stability of enzymes outside optimum operating conditions (pH, temperature, salts, surfactants) and high loading requirements [37]. Although immobilized enzymes and additional modification can improve the stability and antibacterial effect of enzybiotics to a certain extent [30,31,37,38], it increases the complexity and cost of preparation. Therefore, it might boost the catalytic therapy in antibacterial applications if the above problems are solved.
4. Nanozybiotics Are Catalytic Antibacterials Based on Nanozymes with Enzyme-like Activities
Nanozymes are a new generation of artificial enzymes which have unique physicochemical properties and enzyme-like catalytic activity. They catalyze the substrate of natural enzymes under physiological conditions and follow similar enzymatic reaction kinetics [39,40,41,42]. The enzyme-like catalytic activity comes from the intrinsic nano-structure of the nanozyme itself, without the need to introduce additional catalytic functional groups or natural enzymes.
Nanozyme was first reported in 2007 by Gao et al. [43] who found that the ferromagnetic–oxide nanoparticles (Fe3O4 MNPs) had an intrinsic peroxidase-like activity, could catalyze the peroxidase substrate 3,3,5,5-tetramethylbenzidine (TMB), o-phenylenediamine (OPD) and di-azo-aminobenzene (DAB), systematically studied from the perspective of enzymology characteristics of nanomaterials, established the response measurement standards, and used it as a substitute for natural peroxidase in immunoassay. Compared with natural enzymes and other artificial enzymes, nanozymes have better stability, lower cost and adjustable catalytic activity [44,45]. Taking inspiration from natural enzymes–enzybiotics as antibacterial reagents [46,47], we propose use of nanozybiotics to term nanozymes with antibacterial activity. Compared to enzybiotics, nanozybiotics have a broad spectrum and high durability, which is superior for tackling the challenge of antibiotic resistance of bacteria. Moreover, a variety of nanozybiotics have been developed for antibacterial application (partly summarized in Table 1). Therefore, in this section, the antibacterial mechanisms and applications of nanozybiotics based on nanozymes modeled by multiple enzyme-like activities are systematically introduced.
4.1. Peroxidase-like Nanozymes
Peroxidase (POD), such as horseradish peroxidase (HRP), is a kind of hemosiderin enzyme that catalyzes the decomposition of H2O2. It has many applications in wastewater treatment, environmental science, biosensing and organic synthesis. Since the peroxidase-like activity of Fe3O4 MNPs was first reported in 2007, other nanozymes with such activity have been discovered successively (Table 1), and the application of nanozymes has also been extended to many important fields including antibacterial treatment.
Peroxidases catalyze the decomposition of H2O2 and produce free hydroxyl radicals (·OH) with strong oxidation. Hydroxyl radicals are one of the most destructive reactive oxygen species (ROS). It not only decomposes nucleic acid, protein, polysaccharide, and other bacterial biofilm components, but also destroys the structural integrity of bacteria, leading to their death (Figure 4a,c) [122]. Typically, high concentrations of H2O2 (3% or more) are required for sterilization, but higher concentrations of H2O2 may cause cytotoxicity to normal physiological tissues [123]. In contrast, lower concentration of H2O2 (0.5% or less) is needed in the presence of the nanozyme to reach the great antibacterial efficacy [122,124]. So far, many nanozymes with peroxidase-like activity have been regarded as promising antibacterial nanozybiotics [59,110,125,126,127]. For example, iron–oxide nanozymes (IONzymes) were demonstrated to suppress the survival of intracellular Salmonella Enteritidis (S. Enteritidis) via its intrinsic peroxidase-like activity [125].
Moreover, Sun et al. showed that graphene quantum dots (GQDs) with peroxidase-like activity, in the presence of low concentration of H2O2, could produce a large amount of ·OH and effectively kill Gram-negative E. coli and Gram-positive S. aureus. Accordingly, GQDs-Band-Aids were designed to prevent wound infection and promote wound healing [128]. Li et al. prepared one efficient peroxidase-like Fe3C/N-doped graphitic carbon nanomaterial (Fe3C/N-C), which could enable the decomposition of H2O2 to ·OH, resulting in higher broad-spectrum antimicrobial activity than H2O2 alone. Compared with the HRP, Fe3C/N-C exhibited excellent catalytic activity in a wide range of pH (1.0–11.0) and temperature (25–70 °C) [56]. In this model, the Fe3C/N-C nanozyme endowed efficient antibacterial treatment (E. coli and S. aureusas) of wound infection in vivo even in the presence of a much lower-concentration of H2O2 (1.0 mM). Thus, accelerated wound healing in vivo can be achieved without using high concentrations of H2O2.
The antibacterial capacity of nanozybiotics is closely associated with their composition and nanostructure by adjusting enzyme mimic abilities [129]. Xi et al. [108] designed two types of copper/carbon nanozymes including two Cu states (Cu0 and Cu2+). They found that the copper/carbon nanozymes displayed multi-enzyme activities and their antibacterial mechanisms depended on Cu states. Hollow carbon spheres (HCSs) modified with CuO (CuO-HCSs) nanozymes would induce Gram-negative bacteria death (E. coli and P. aeruginosa) when releasing Cu2+. While Cu-HCSs nanozymes killing both Gram-positive (Salmonella typhimurium, S. typhimurium) and Gram-negative bacteria (E. coli and P. aeruginosa) were based on POD-like activity which is responsible for ROS generation.
Given the acidic microenvironment in bacterial accumulation due to large amounts of organic acid produced during anaerobic fermentation, Yu et al. [130] developed a novel acid-responsive ROS generator for biofilm removal by fabricating Zeolitic imidazolate framework-8 (ZIF-8) co-encapsulating the antibacterial ligand (lysine carbon dots, Lys-CDs) and targeted drug (folic acid, FA). The synthesized ZIF-8@Lys-CD@FA nanozybiotics showed peroxidase-like activity in an acid environment, and produced extremely active hydroxyl radicals to effectively destroy mature biofilms, resulting in the significantly improved bacteriostatic rate against E. coli and S. aureus. In addition, in the acidic environments, S. aureus and E. coli were more sensitive to ZIF-8@Lys-CD@FA with MIC values at 32 μg mL−1 and 62.5 μg mL−1 at pH 5.5, and 62.5 μg mL−1 and 125 μg mL−1 at pH 6.5, respectively. Moreover, the pH-dependent catalytic activity of nanozybiotics may limit their antimicrobial application under neutral pH. Therefore, the enhanced catalysis ability of citrate modified with Fe3O4 NPs has been developed by using ATP as a synergist [131]. In this work, Fe3O4 NPs exhibited superior antibacterial performance against E. coli and Bacillus subtilis (B. subtilis, Gram-positive) in the presence of H2O2 under a neutral pH environment with the assistance of ATP.
Recently, catalytic nanozybiotics were shown to disrupt biofilms but lacked a stabilizing coating required for clinical applications. Dextran-coated iron–oxide nanozymes (Dex-NZM) [132] displayed strong peroxidase-like activity at acidic pH values, targeted biofilms with high specificity, and prevented severe caries without impacting surrounding oral tissues in vivo. The Km values for H2O2 were found to be 27 μM and 2.5 mM for Dex-NZM and HRP, respectively. Besides, Dex-NZM/H2O2 treatment significantly reduced the onset and severity of caries lesions (vs control or either Dex-NZM or H2O2 alone) without adverse effects on gingival tissues or oral microbiota diversity.
4.2. Oxidase-like Nanozymes
Oxidase is an important enzyme that catalyzes redox reaction of oxygen. In oxidase-catalyzed reactions, the molecular oxygen (O2) is oxidized and converted to H2O or H2O2 (in certain cases to superoxide radicals, O2−) (Figure 4b) [40]. The active free radicals produced simultaneously in the catalytic process exert antibacterial activity. Due to its strong reactivity, H2O2 directly oxidizes the outer structure of bacteria, destroys the permeability barrier of bacteria, and leads to an imbalance of the electrochemical balance between internal and external substances of bacteria, resulting in bacterial death [47]. Similarly, O2− can react directly with nucleic acids, proteins, etc., causing bacterial death.
In particular, the oxidase- and peroxidase-like activities have shown to be major mechanisms of the antimicrobial efficiency for noble-metal-based nanozymes [133]. For example, Tao et al. [133] found firstly AuNPs as the oxidase mimic and then constructed the AuNPs supported on bifunctionalized mesoporous silica (MSN) (MSN-AuNPs) to achieve dual enzyme activities similar to those of peroxidase and oxidase. They evaluated the antibacterial potency and found that MSN-AuNPs not only exhibited striking antibacterial properties against both Gram-negative (E. coli) and Gram-positive (S. aureus) bacteria, but also degraded the existing biofilm and prevented formation of new biofilm efficiently.
In addition, Ge Fang and co-workers found that palladium (Pd) nanocrystals exhibit facet-dependent oxidase- and peroxidase-like activities that endow them with excellent antibacterial properties [134]. In this work, Pd cubes with higher activities killed Gram-positive drug-resistant Staphylococcus aureus (S. aureus) and Enterococcus faecalis, while Pd octahedrons displaying stronger penetration into bacterial membranes exerted higher antibacterial activity for Gram-negative Escherichia coli (E. coli) and Salmonella enteritidis. Pd-based nanostructures also exhibit excellent oxidase-like activity [134]. It is reported that Pd@Ir octahedra demonstrates powerful bactericidal activity against both E. coli and S. aureus due to significant enhancement of oxidation byproducts [111]. While the application of noble-metals is limited due to high prices, a study reported that terbium oxide nanoparticles (Tb4O7 NPs) with oxidase-like activity at acidic pH values can be easily synthesized with low cost [110]. Moreover, Tb4O7 NPs were able to quickly oxidize a series of organic substrates and produce hydroxyl radicals. Not surprisingly, Tb4O7 NPs exhibited potent antimicrobial activity against both S. aureus and E. coli. When the concentration of Tb4O7 NPs increased to 100 μg/mL, nearly 90% of the S. aureus were killed. The application of the antibacterial activities of Tb4O7 NPs also were validated in a wound infection mouse model.
Recently, He et al. [135] synthesized bamboolike nitrogen-doped carbon nanotubes encapsulating cobalt nanoparticles (N-CNTs@Co) which showed 12.1 times higher oxidase-mimicking activity than that of the most reported CeO2. They demonstrated that N-CNTs@Co exerted a great antibacterial effect against Gram-positive (S. aureus) and Gram-negative (E. coli) in vitro and in vivo by catalyzing oxygen to produce a large number of ROS under acidic condition. Notably, during antibacterial experiments (20 days), neither S. aureus nor E. coli developed resistance to N-CNTs@Co.
All of the above indicate that nanozybiotics with oxidase-like activity bear great development potential in the field of antibacterial application. Moreover, given that oxidase can generate hydrogen peroxide, which is the substrate of peroxidase, it also suggested that the enzyme cascade reaction may be another choice for efficient antibacterial activity for nanozymes with multiple activities.
4.3. Deoxyribonuclease-like Nanozymes
Drug-resistant genes from dead bacteria can remain in the environment and spread to other microbes via horizontal gene transfer. Drug-resistant genes are essentially DNA, a complementary double strand composed of deoxynucleotide units, which are connected by 3′,5′-phosphodiester bonds. As the carrier of life genetic information, DNA is highly stable with a half-life of 521 years in the environment [136,137,138]. Even if ultraviolet, chlorine, ozone, and other physical methods are used, the destructive effect of DNA is not ideal [139,140,141]. However, DNA can be decomposed by natural nucleases, such as deoxyribonuclease (DNase), which acts on 3′,5′-phosphodiester bond (Figure 2c). Therefore, development of antimicrobial materials with the ability to degrade DNA can prevent the dissemination of released drug-resistant genes from dead bacteria (Figure 5a). In addition, considering that eDNA is an important part of biofilm, nanozybiotics based on nanozymes with DNase-like activity also demonstrate an antibiofilm effect (Figure 5b). Recently, artificial nucleases have been developed to mimic the hydrolytic cleavage of phosphodiester bond by taking advantage of multinuclear metal complexes, including transition metals and rare earth elements such as Cu(II), Cr(III), Zn(II), Ce(IV) [142,143,144,145,146]. Among them, cerium complex has attracted considerable attention due to its high catalytic efficiency and good biocompatibility [16,147].
Zhaowei Chen and co-workers designed a DNase-mimetic artificial enzyme (DMAE) for anti-biofilm application. DMAE effectively prevented more than 90% bacterial adhesion and inhibition by degrading eDNA in S. aureus extracellular polymeric substances (EPS) [16]. In addition, imidazolium type poly (ionic liquid) (PIL)/cerium (IV) ion-based electrospun nanofibrous membranes (PIL-Ce) showed a DNase-like activity [17]. The PIL-Ce was able to cleave the phosphodiester bond of BNPP and performed the characteristics of nucleases (Km value = 0.2656 mM). The antibacterial test of PIL-Ce showed the high efficiencies to eradicate bacteria and disintegrate drug-resistant genes. The wound treatment test using MRSA infected mice as the model further demonstrated that PIL-Ce membranes combined both antibacterial and DNase-mimic properties, and may have potential application as a new “green” wound dressing to block the drug resistance spread in a clinical setting. Furthermore, rare earth elements and cerium can be complexed as artificial nucleases with a mechanism of action on the phosphodiester bond. Upon the addition of a Ce-containing complex, two adjacent Ce ions can interact with one phosphodiester bond, and render the phosphate bond susceptible to a nucleophilic species.
4.4. Combination Therapy
Various antibacterial alternatives have been developed to overcome the drug-resistant behavior of microorganisms, such as peptides, metal–sulfide/oxides and carbon-based nanostructures [124,148,149]. However, they still suffer from biotoxicity, high costs, cumbersome preparation processes and/or pollution. Though nanozybiotics, possessing long-term storage, good stability, and tunable catalytic properties, could be applied to construct a range of antibacterial systems, there is still a large space to improve their antimicrobial performance [150]. Therefore, combination therapy seems to potentially be the better option to improve antibacterial effects as much as possible while minimizing side effects [151]. In various antimicrobial therapies, photo-activated strategies such as photothermal therapy (PTT), photodynamic therapy (PDT) have attracted widespread attention, owing to their less-invasive nature, low side effects, and good controllability compared to other antibacterial agents [152,153,154]. The antibacterial mechanism of combination therapy is summarized in Figure 6 with more details.
Recently, Yin et al. [155] reported that polyethylene glycol functionalized MoS2 nanoflowers (PEG-MoS2 NFs) with peroxidase-like catalytic activity and high photothermal conversion efficiency in the near-infrared (NIR) region combined the catalysis with PTT to provide a rapid and great antibacterial effect. In this work, PEG-MoS2 NFs could eliminate both Gram-negative ampicillin resistant Escherichia coli (Ampr E. coli) and Gram-positive endospore-forming Bacillus subtilis (B. subtilis) with a low concentration of H2O2 and 808 nm irradiation. Such a combination not only improved the cell wall damage induced by ·OH, but also minimized the side effects of PTT with a large shortening the treatment time.
Zhang et al. [70] combined photodynamic, photothermal and peroxidase-like enzymatic activities to maximize the antibacterial efficiency by using oxygen-vacancy molybdenum trioxide nanodots (MoO3−x NDs). The synergistic combination of multiple therapies enabled MoO3−x NDs to possess a lower Km value and a higher Vmax value compared with other peroxidase mimics. In vitro antibacterial tests showed that MoO3−x NDs/H2O2/808 nm NIR decreased viability of MRSA and ESBL-producing E. coli, by destroying the bacterial cell surface. In particular, with H2O2 at low concentration (100 μm) and upon NIR exposure, the MoO3−x NDs was heated to the optimum enzymatic temperature of peroxidase-like activity and released utmost ·OH through the intrinsic photothermal effect for killing bacteria and accelerating wound healing.
Sun et al. [118] developed an antibacterial strategy by combining sonodynamic therapy and catalase-like activity of Pd@Pt nanozymes. Sonodynamic therapy (SDT) uses ultrasound to activate acoustic sensitizer and triggers ROS to produce antibacterial effects, which has the advantages of a non-invasive mode and good tissue penetration. A nanoplatform (Pd@Pt-T790) was constructed by bridging Pd@Pt nanoplates with the organic sonosensitizer meso-tetra(4-carboxyphenyl)porphine (T790). The modification of T790 onto Pd@Pt could offer Pd@Pt-T790 a “blocking and activating” enzyme-like activity, namely upon US irradiation, the nanozyme activity was effectively recovered to catalyze the decomposition of endogenous H2O2 into O2. This elaborate strategy was helpful to decrease the potential toxicity and side effects of nanozymes on normal tissues and had potential to realize active, controllable, and disease-loci-specific nanozyme catalytic behavior. Moreover, the anti-bacterial test of the Pd@Pt-T790-based SDT nano-system demonstrated therapeutic effects to eradicate methicillin-resistant S. aureus (MRSA)-induced myositis.
Collectively, compared with enzybiotics, nanozybiotics bear a greater and broader application prospect, which may overcome the limitations of natural enzymes and combine other antibacterial strategies easily (Table 2). In addition, with the characteristics of being more stable, economical, and practical, nanozybiotics are more suitable for widespread application in industrial processes. However, it is worth noting that the catalytic efficiency of nanozybiotics is still not very high compared with natural enzymes, and the types of enzyme activities are fewer. Moreover, it is interesting that enzybiotics usually kill bacteria by directly catalyzing the destruction of bacterial cell structure, whereas nanozybiotics realize antibacterial effects by producing toxic ROS to destroy the cell structure. Thus, to improve the antibacterial performance of nanozybiotics, developing a new catalytic activity and enhancing antibacterial targeting are necessary in future studies.
As an antibacterial strategy, the ultimate goal of nanozybiotics is in vivo application and clinical transformation. However, the current research mainly focuses on in vitro biological experiments or skin surface wound infection models [56,65,73,83,92,93,110], and there are only a few studies on antibacterial application of nanozybiotics in vivo [60,85,96,100,116]. For example, some nanozybiotics are designed to exert an excellent effect on Helicobacter Pylori (H. pylori) eradication in vivo via pH-responsive peroxidase-and oxidase-like activity [60,100,116]. Zhu et al. constructed a cationic chitosan coated ruthenium dioxide nanozyme (QCS-RuO2@RBT, SRT NSs) for the management of biofilm-associated infections, including chronic lung infection [85]. Although nanozybiotics bear biological enzyme activity and have shown excellent therapeutic efficacy in some animal models of in vivo diseases, most nanozymes are inorganic nanomaterials. Due to nanomaterials properties or function varies with the size, composition, morphology and shape, surface modification and surface charge, the studies of biological safety and toxicology is very complex, especially in clinical applications, more special attention and efforts need to be given. Moreover, it represents an important research direction in the field of nanozybiotics in the future.
5. Conclusions and Perspective
Enzymatic therapy represents a promising strategy to combat challenges posed by the drug resistance of bacteria. Natural enzyme-based enzybiotics, originate from nature with great environmental friendliness and exhibiting a high catalytic antibacterial ability. The antibacterial mechanism of enzybiotics usually describes the enzymatic degradation of the bacterial cell structure or biofilms leading to bacterial death. But their disadvantage is that most natural enzymes are unstable in the process of industrial production, which limits their large-scale application and increases costs. Although immobilization and additional modification can improve the stability of natural enzymes to some extent, it increases the production cost and the complexity of production operation. Nanozymes are a kind of nanomaterial with enzyme-like activity, and are economical and stable compared with natural enzymes. At present, nanozyme-based nanozybiotics have shown great antibacterial application prospects against resistant bacteria by mimicking enzyme-like activities of natural enzymes. Their antibacterial performance can be further improved by combining enzyme-like properties with other physiochemical properties of nanozymes such as PTT and PDT. In addition, most antibacterial tests were validated under in vitro environments or topically administered with in vivo models, which means there remains a long road before reaching clinical transformation. Thus, exploring new biocompatible nanozybiotics using enzyme-like nanozymes equipped with multiple antibacterial capabilities and applicable scenarios is of great importance. Altogether, we believe that nanozybiotics based on nanozymes with enzyme-like activity represent a new class of antibiotics alternatives.
Author Contributions
C.Z.: Literature search, Figures, Data collection, Data analysis, Data interpretation, Methodology, Formal analysis, and Writing–Original Draft; Q.W.: Methodology and Writing–Review & Editing; J.J.: Writing–Review & Editing; L.G.: Conceptualization, Resources, Writing–Review & Editing and Funding acquisition. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the National Key R&D Program of China grant 2019YFA0709200, as well as the National Natural Science Foundation of China grant 81930050 (L.G.), Wuxi Science and Technology Plan Project N2020X018, the scientific research project plan of Wuxi Municipal Health Commission was approved and funded M202007.
Acknowledgments
We thank Jiuyang He for language editing and support from the CAS Engineering Laboratory for Nanozymes, Institute of Biophysics, Chinese Academy of Sciences, China.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Jernigan, J.A.; Hatfield, K.M.; Wolford, H.; Nelson, R.E.; Olubajo, B.; Reddy, S.C.; McCarthy, N.; Paul, P.; McDonald, L.C.; Kallen, A.; et al. Multidrug-resistant bacterial infections in us hospitalized patients, 2012–2017. N. Engl. J. Med. 2020, 382, 1309–1319. [Google Scholar] [CrossRef] [PubMed]
- Mahlapuu, M.; Håkansson, J.; Ringstad, L.; Björn, C. Antimicrobial Peptides: An Emerging Category of Therapeutic Agents. Front. Cell. Infect. Microbiol. 2016, 6, 194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Panja, S.; Bharti, R.; Dey, G.; Lynd, N.A.; Chattopadhyay, S. Coordination-Assisted Self-Assembled Polypeptide Nanogels to Selectively Combat Bacterial Infection. ACS Appl. Mater. Interfaces 2019, 11, 33599–33611. [Google Scholar] [CrossRef] [PubMed]
- Hancock, R.E.W.; Sahl, H.G. Antimicrobial and host-defense peptides as new anti-infective therapeutic strategies. Nat. Biotechnol. 2006, 24, 1551–1557. [Google Scholar] [CrossRef] [PubMed]
- Bacalum, M.; Radu, M. Cationic Antimicrobial Peptides Cytotoxicity on Mammalian Cells: An Analysis Using Therapeutic Index Integrative Concept. Int. J. Pept. Res. Ther. 2015, 21, 47–55. [Google Scholar] [CrossRef]
- Donlan, R.M. Preventing biofilms of clinically relevant organisms using bacteriophage. Trends Microbiol. 2009, 17, 66–72. [Google Scholar] [CrossRef]
- Labrie, S.J.; Samson, J.E.; Moineau, S. Bacteriophage resistance mechanisms. Nat. Rev. Microbiol. 2010, 8, 317–327. [Google Scholar] [CrossRef]
- Dedrick, R.M.; Guerrero-Bustamante, C.A.; Garlena, R.A.; Russell, D.A.; Ford, K.; Harris, K.; Gilmour, K.C.; Soothill, J.; Jacobs-Sera, D.; Schooley, R.T.; et al. Engineered bacteriophages for treatment of a patient with a disseminated drug-resistant Mycobacterium abscessus. Nat. Med. 2019, 25, 730–733. [Google Scholar] [CrossRef] [PubMed]
- Datta, S.; Rajnish, K.N.; Doss, C.G.P.; Samuel, S.M.; Selvarajan, E.; Zayed, H. Enzyme therapy: A forerunner in catalyzing a healthy society? Expert Opin. Biol. Ther. 2020, 20, 1151–1174. [Google Scholar] [CrossRef]
- Dams, D.; Briers, Y. Enzybiotics: Enzyme-Based Antibacterials as Therapeutics. In Therapeutic Enzymes: Function and Clinical Implications; Labrou, N., Ed.; Springer International Publishing: Cham, Switzerland, 2019; Volume 1148, pp. 233–253. [Google Scholar]
- Wu, J.; Wang, X.; Wang, Q.; Lou, Z.; Li, S.; Zhu, Y.; Qin, L.; Wei, H. Nanomaterials with enzyme-like characteristics (nanozymes): Next-generation artificial enzymes (II). Chem. Soc. Rev. 2019, 48, 1004–1076. [Google Scholar] [CrossRef]
- Liu, Z.; Zhao, X.; Yu, B.; Zhao, N.; Zhang, C.; Xu, F.-J. Rough Carbon–Iron Oxide Nanohybrids for Near-Infrared-II Light-Responsive Synergistic Antibacterial Therapy. ACS Nano 2021, 15, 7482–7490. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Wang, D.; Wen, J.; Yu, P.; Liu, J.; Li, J.; Chu, H. Chemically Grafted Nanozyme Composite Cryogels to Enhance Antibacterial and Biocompatible Performance for Bioliquid Regulation and Adaptive Bacteria Trapping. ACS Nano 2021, 15, 19672–19683. [Google Scholar] [CrossRef] [PubMed]
- Meng, F.; Peng, M.; Chen, Y.; Cai, X.; Huang, F.; Yang, L.; Liu, X.; Li, T.; Wen, X.; Wang, N.; et al. Defect-rich graphene stabilized atomically dispersed Cu3 clusters with enhanced oxidase-like activity for antibacterial applications. Appl. Catal. B Environ. 2021, 301, 120826. [Google Scholar] [CrossRef]
- Li, Y.; Yu, P.; Wen, J.; Sun, H.; Wang, D.; Liu, J.; Li, J.; Chu, H. Nanozyme-Based Stretchable Hydrogel of Low Hysteresis with Antibacterial and Antioxidant Dual Functions for Closely Fitting and Wound Healing in Movable Parts. Adv. Funct. Mater. 2021, 2110720. [Google Scholar] [CrossRef]
- Chen, Z.; Ji, H.; Liu, C.; Bing, W.; Wang, Z.; Qu, X. A Multinuclear Metal Complex Based DNase-Mimetic Artificial Enzyme: Matrix Cleavage for Combating Bacterial Biofilms. Angew. Chem. Int. Ed. 2016, 55, 10732–10736. [Google Scholar] [CrossRef]
- Luo, Z.; Cui, H.; Guo, J.; Yao, J.; Fang, X.; Yan, F.; Wang, B.; Mao, H. Poly(ionic liquid)/ce-based antimicrobial nanofibrous membrane for blocking drug-resistance dissemination from mrsa-infected wounds. Adv. Funct. Mater. 2021, 31, 2100336. [Google Scholar] [CrossRef]
- Davies, J.; Davies, D. Origins and Evolution of Antibiotic Resistance. Microbiol. Mol. Biol. Rev. 2010, 74, 417–433. [Google Scholar] [CrossRef] [Green Version]
- Blair, J.M.; Webber, M.A.; Baylay, A.J.; Ogbolu, D.O.; Piddock, L.J. Molecular mechanisms of antibiotic resistance. Nat. Rev. Microbiol. 2015, 13, 42–51. [Google Scholar] [CrossRef]
- Abushaheen, M.A.; Muzaheed; Fatani, A.J.; Alosaimi, M.; Mansy, W.; George, M.; Acharya, S.; Rathod, S.; Divakar, D.D.; Jhugroo, C.; et al. Antimicrobial resistance, mechanisms and its clinical significance. Dis. Mon. 2020, 66, 100971. [Google Scholar] [CrossRef]
- HallR, M.; Collis, C.M. Mobile gene cassettes and integrons: Capture and spread of genes by site-specific recombination. Mol. Microbiol. 1995, 15, 593–600. [Google Scholar] [CrossRef]
- Hancock, R. The bacterial outer membrane as a drug barrier. Trends Microbiol. 1997, 5, 37–42. [Google Scholar] [CrossRef]
- Hu, D.; Li, H.; Wang, B.; Ye, Z.; Lei, W.; Jia, F.; Jin, Q.; Ren, K.-F.; Ji, J. Surface-Adaptive Gold Nanoparticles with Effective Adherence and Enhanced Photothermal Ablation of Methicillin-Resistant Staphylococcus aureus Biofilm. ACS Nano 2017, 11, 9330–9339. [Google Scholar] [CrossRef] [PubMed]
- Borysowski, J.; Górski, A. Enzybiotics and Their Potential Applications in Medicine. In Enzybiotics: Antibiotic Enzymes as Drugs and Therapeutics; Villa, T.G., Veiga-Crespo, P., Eds.; Wiley: New York, NY, USA, 2009; pp. 1–26. [Google Scholar]
- Heselpoth, R.D.; Swift, S.M.; Linden, S.B.; Mitchell, M.S.; Nelson, D.C. Enzybiotics: Endolysins and Bacteriocins. In Bacteriophages: Biology, Technology, Therapy; Harper, D., Abedon, S., Burrowes, B., McConville, M., Eds.; Springer International Publishing: Cham, Switzerland, 2017; pp. 1–42. [Google Scholar]
- King, B.F.; Biel, M.L.; Wilkinson, B.J. Facile penetration of the Staphylococcus aureus capsule by lysostaphin. Infect. Immun. 1980, 29, 892–896. [Google Scholar] [CrossRef] [PubMed]
- de Freire Bastos, M.d.C.; Coutinho, B.G.; Varella Coelho, M.L. Lysostaphin: A Staphylococcal Bacteriolysin with Potential Clinical Applications. Pharmaceuticals 2010, 3, 1139–1161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Primo, E.D.; Otero, L.; Ruiz, F.; Klinke, S.; Giordano, W. The disruptive effect of lysozyme on the bacterial cell wall explored by an in-silico structural outlook. Biochem. Mol. Biol. Educ. 2018, 46, 83–90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saito, H.; Sakakibara, Y.; Sakata, A.; Kurashige, R.; Murakami, D.; Kageshima, H.; Saito, A.; Miyazaki, Y. Antibacterial activity of lysozyme-chitosan oligosaccharide conjugates (LYZOX) against Pseudomonas aeruginosa, Acinetobacter baumannii and Methicillin-resistant Staphylococcus aureus. PLoS ONE 2019, 14, e0217504. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, H. On the novel catalytically-independent antimicrobial function of hen egg-white lysozyme: A conformation-dependent activity. Food/Nahrung 1998, 42, 187–193. [Google Scholar] [CrossRef]
- Arima, H.; Ibrahim, H.R.; Kinoshita, T.; Kato, A. Bactericidal action of lysozymes attached with various sizes of hydrophobic peptides to the C-terminal using genetic modification. FEBS Lett. 1997, 415, 114–118. [Google Scholar] [CrossRef] [Green Version]
- Saperas, N.; Fonfría-Subirós, E. Proteolytic Enzymes in Detergents: Evidence of Their Presence through Activity Measurements Based on Electrophoresis. J. Chem. Educ. 2011, 88, 1702–1706. [Google Scholar] [CrossRef] [Green Version]
- Eshamah, H.; Han, I.; Naas, H.; Acton, J.; Dawson, P. Antibacterial effects of natural tenderizing enzymes on different strains of Escherichia coli O157:H7 and Listeria monocytogenes on beef. Meat Sci. 2014, 96, 1494–1500. [Google Scholar] [CrossRef]
- Praveen, N.C.; Rajesh, A.; Madan, M.; Chaurasia, V.R.; Hiremath, N.V.; Sharma, A.M. In vitro Evaluation of Antibacterial Efficacy of Pineapple Extract (Bromelain) on Periodontal Pathogens. J. Int. Oral Health 2014, 6, 96–98. [Google Scholar]
- Eller, C.H.; Raines, R.T. Antimicrobial Synergy of a Ribonuclease and a Peptide Secreted by Human Cells. ACS Infect. Dis. 2020, 6, 3083–3088. [Google Scholar] [CrossRef] [PubMed]
- Banu, S.F.; Thamotharan, S.; Gowrishankar, S.; Pandian, S.K.; Nithyanand, P. Marine bacterial DNase curtails virulence and disrupts biofilms of Candida albicans and non-albicans Candida species. Biofouling 2019, 35, 975–985. [Google Scholar] [CrossRef] [PubMed]
- Sheldon, R.A.; van Pelt, S. Enzyme immobilisation in biocatalysis: Why, what and how. Chem. Soc. Rev. 2013, 42, 6223–6235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iyer, P.V.; Ananthanarayan, L. Enzyme stability and stabilization—Aqueous and non-aqueous environment. Process Biochem. 2008, 43, 1019–1032. [Google Scholar] [CrossRef]
- Wei, H.; Wang, E. Nanomaterials with enzyme-like characteristics (nanozymes): Next-generation artificial enzymes. Chem. Soc. Rev. 2013, 42, 6060. [Google Scholar] [CrossRef]
- Ragg, R.; Tahir, M.N.; Tremel, W. Solids Go Bio: Inorganic Nanoparticles as Enzyme Mimics. Eur. J. Inorg. Chem. 2015, 2016, 1906–1915. [Google Scholar] [CrossRef]
- Gao, L.; Yan, X. Nanozymes: An emerging field bridging nanotechnology and biology. Sci. China Life Sci. 2016, 59, 400–402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, Y.; Ren, J.; Qu, X. Catalytically Active Nanomaterials: A Promising Candidate for Artificial Enzymes. Accounts Chem. Res. 2014, 47, 1097–1105. [Google Scholar] [CrossRef]
- Gao, L.; Zhuang, J.; Nie, L.; Zhang, J.; Zhang, Y.; Gu, N.; Wang, T.; Feng, J.; Yang, D.; Perrett, S.; et al. Intrinsic peroxidase-like activity of ferromagnetic nanoparticles. Nat. Nanotechnol. 2007, 2, 577–583. [Google Scholar] [CrossRef]
- Jiang, D.; Ni, D.; Rosenkrans, Z.T.; Huang, P.; Yan, X.; Cai, W. Nanozyme: New horizons for responsive biomedical applications. Chem. Soc. Rev. 2019, 48, 3683–3704. [Google Scholar] [CrossRef] [PubMed]
- Liang, M.; Yan, X. Nanozymes: From New Concepts, Mechanisms, and Standards to Applications. Accounts Chem. Res. 2019, 52, 2190–2200. [Google Scholar] [CrossRef] [PubMed]
- Laurents, D.; Baldwin, R.L. Characterization of the Unfolding Pathway of Hen Egg White Lysozyme. Biochemistry 1997, 36, 1496–1504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vatansever, F.; De Melo, W.C.; Avci, P.; Vecchio, D.; Sadasivam, M.; Gupta, A.; Chandran, R.; Karimi, M.; Parizotto, N.A.; Yin, R.; et al. Antimicrobial strategies centered around reactive oxygen species—Bactericidal antibiotics, photodynamic therapy, and beyond. FEMS Microbiol. Rev. 2013, 37, 955–989. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, B.; Wang, H.; Wang, W.; Gao, L.; Li, S.; Pan, X.; Wang, H.; Yang, H.; Meng, X.; Wu, Q.; et al. A Single-Atom Nanozyme for Wound Disinfection Applications. Angew. Chem. Int. Ed. 2019, 58, 4911–4916. [Google Scholar] [CrossRef]
- Liu, X.P.; Yan, Z.Q.; Zhang, Y.; Liu, Z.W.; Sun, Y.H.; Ren, J.S.; Qu, X.G. Two-dimensional metal-organic framework/enzyme hybrid nanocatalyst as a benign and m self-activated cascade reagent for in vivo wound healing. Acs Nano 2019, 13, 5222–5230. [Google Scholar] [CrossRef]
- Hwang, G.; Paula, A.J.; Hunter, E.E.; Liu, Y.; Babeer, A.; Karabucak, B.; Stebe, K.; Kumar, V.; Steager, E.; Koo, H. Catalytic antimicrobial robots for biofilm eradication. Sci. Robot. 2019, 4, eaaw2388. [Google Scholar] [CrossRef]
- Sang, Y.; Li, W.; Liu, H.; Zhang, L.; Wang, H.; Liu, Z.; Ren, J.; Qu, X. Construction of Nanozyme-Hydrogel for Enhanced Capture and Elimination of Bacteria. Adv. Funct. Mater. 2019, 29, 1900518. [Google Scholar] [CrossRef]
- Liu, Y.; Lin, A.; Liu, J.; Chen, X.; Zhu, X.; Gong, Y.; Yuan, G.; Chen, L.; Liu, J. Enzyme-Responsive Mesoporous Ruthenium for Combined Chemo-Photothermal Therapy of Drug-Resistant Bacteria. ACS Appl. Mater. Interfaces 2019, 11, 26590–26606. [Google Scholar] [CrossRef]
- Huo, M.; Wang, L.; Zhang, H.; Zhang, L.; Chen, Y.; Shi, J. Construction of Single-Iron-Atom Nanocatalysts for Highly Efficient Catalytic Antibiotics. Small 2019, 15, e1901834. [Google Scholar] [CrossRef]
- Qiu, H.; Pu, F.; Liu, Z.; Liu, X.; Dong, K.; Liu, C.; Ren, J.; Qu, X. Hydrogel-based artificial enzyme for combating bacteria and accelerating wound healing. Nano Res. 2020, 13, 496–502. [Google Scholar] [CrossRef]
- Wang, Y.; Shen, X.; Ma, S.; Guo, Q.; Zhang, W.; Cheng, L.; Ding, L.; Xu, Z.; Jiang, J.; Gao, L. Oral biofilm elimination by combining iron-based nanozymes and hydrogen peroxide-producing bacteria. Biomater. Sci. 2020, 8, 2447–2458. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Ma, W.; Sun, J.; Lin, M.; Niu, Y.; Yang, X.; Xu, Y. Electrochemical generation of Fe3C/N-doped graphitic carbon nanozyme for efficient wound healing in vivo. Carbon 2020, 159, 149–160. [Google Scholar] [CrossRef]
- Wang, T.; Zhang, X.; Mei, L.; Ma, D.; Liao, Y.; Zu, Y.; Xu, P.; Yin, W.; Gu, Z. A two-step gas/liquid strategy for the production of N-doped defect-rich transition metal dichalcogenide nanosheets and their antibacterial applications. Nanoscale 2020, 12, 8415–8424. [Google Scholar] [CrossRef] [PubMed]
- Xi, J.; Wei, G.; Wu, Q.; Xu, Z.; Liu, Y.; Han, J.; Fan, L.; Gao, L. Light-enhanced sponge-like carbon nanozyme used for synergetic antibacterial therapy. Biomater. Sci. 2019, 7, 4131–4141. [Google Scholar] [CrossRef] [PubMed]
- Hu, W.C.; Younis, M.R.; Zhou, Y.; Wang, C.; Xia, X.H. In situ fabrication of ultrasmall gold nanoparticles/2d mofs hybrid as nanozyme for antibacterial therapy. Small 2020, 16, e2000553. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.X.; Chen, Z.T.; Kong, J.L.; Liang, Y.L.; Chen, K.; Chang, Y.A.; Yuan, H.; Wang, Y.J.; Liang, H.J.; Li, J.C.; et al. Fullerenol nanoparticles eradicate helicobacter pylori via ph-responsive peroxidase activity. ACS Appl. Mater. Interfaces 2020, 12, 29013–29023. [Google Scholar] [CrossRef] [PubMed]
- Guo, Z.; Liu, Y.; Zhang, Y.; Sun, X.; Li, F.; Bu, T.; Wang, Q.; Wang, L. A bifunctional nanoplatform based on copper manganate nanoflakes for bacterial elimination via a catalytic and photothermal synergistic effect. Biomater. Sci. 2020, 8, 4266–4274. [Google Scholar] [CrossRef] [PubMed]
- Gao, F.; Li, X.; Zhang, T.; Ghosal, A.; Zhang, G.; Fan, H.M.; Zhao, L. Iron nanoparticles augmented chemodynamic effect by alternative magnetic field for wound disinfection and healing. J. Control. Release 2020, 324, 598–609. [Google Scholar] [CrossRef]
- Zhang, W.; Ren, X.; Shi, S.; Li, M.; Liu, L.; Han, X.; Zhu, W.; Yue, T.; Sun, J.; Wang, J. Ionic silver-infused peroxidase-like metal–organic frameworks as versatile “antibiotic” for enhanced bacterial elimination. Nanoscale 2020, 12, 16330–16338. [Google Scholar] [CrossRef]
- Yim, G.; Kim, C.Y.; Kang, S.; Min, D.-H.; Kang, K.; Jang, H. Intrinsic Peroxidase-Mimicking Ir Nanoplates for Nanozymatic Anticancer and Antibacterial Treatment. ACS Appl. Mater. Interfaces 2020, 12, 41062–41070. [Google Scholar] [CrossRef] [PubMed]
- Liang, M.; Wang, Y.; Ma, K.; Yu, S.; Chen, Y.; Deng, Z.; Liu, Y.; Wang, F. Engineering Inorganic Nanoflares with Elaborate Enzymatic Specificity and Efficiency for Versatile Biofilm Eradication. Small 2020, 16, 2002348. [Google Scholar] [CrossRef] [PubMed]
- Wei, F.; Cui, X.; Wang, Z.; Dong, C.; Li, J.; Han, X. Recoverable peroxidase-like Fe3O4@MoS2-Ag nanozyme with enhanced antibacterial ability. Chem. Eng. J. 2021, 408, 127240. [Google Scholar] [CrossRef] [PubMed]
- Deshmukh, A.R.; Aloui, H.; Kim, B.S. In situ growth of gold and silver nanoparticles onto phyto-functionalized boron nitride nanosheets: Catalytic, peroxidase mimicking, and antimicrobial activity. J. Clean. Prod. 2020, 270, 122339. [Google Scholar] [CrossRef]
- Fang, J.; Wang, H.; Bao, X.; Ni, Y.; Teng, Y.; Liu, J.; Sun, X.; Sun, Y.; Li, H.; Zhou, Y. Nanodiamond as efficient peroxidase mimic against periodontal bacterial infection. Carbon 2020, 169, 370–381. [Google Scholar] [CrossRef]
- Huang, Y.; Liu, Y.; Shah, S.; Kim, D.; Simon-Soro, A.; Ito, T.; Hajfathalian, M.; Li, Y.; Hsu, J.C.; Nieves, L.M.; et al. Precision targeting of bacterial pathogen via bi-functional nanozyme activated by biofilm microenvironment. Biomaterials 2021, 268, 120581. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, D.; Tan, J.; Chang, Z.; Liu, X.; Ma, W.; Xu, Y. Near-Infrared Regulated Nanozymatic/Photothermal/Photodynamic Triple-Therapy for Combating Multidrug-Resistant Bacterial Infections via Oxygen-Vacancy Molybdenum Trioxide Nanodots. Small 2021, 17, e2005739. [Google Scholar] [CrossRef]
- Kumari, N.; Kumar, S.; Karmacharya, M.; Dubbu, S.; Kwon, T.; Singh, V.; Chae, K.H.; Kumar, A.; Cho, Y.-K.; Lee, I.S. Surface-Textured Mixed-Metal-Oxide Nanocrystals as Efficient Catalysts for ROS Production and Biofilm Eradication. Nano Lett. 2020, 21, 279–287. [Google Scholar] [CrossRef] [PubMed]
- Hou, S.; Mahadevegowda, S.H.; Lu, D.; Zhang, K.; Chan-Park, M.B.; Duan, H. Metabolic Labeling Mediated Targeting and Thermal Killing of Gram-Positive Bacteria by Self-Reporting Janus Magnetic Nanoparticles. Small 2021, 17, e2006357. [Google Scholar] [CrossRef]
- Yan, L.; Mu, J.; Ma, P.; Li, Q.; Yin, P.; Liu, X.; Cai, Y.; Yu, H.; Liu, J.; Wang, G.; et al. Gold nanoplates with superb photothermal efficiency and peroxidase-like activity for rapid and synergistic antibacterial therapy. Chem. Commun. 2020, 57, 1133–1136. [Google Scholar] [CrossRef]
- Liu, Y.; Nie, N.; Tang, H.; Zhang, C.; Chen, K.; Wang, W.; Liu, J. Effective Antibacterial Activity of Degradable Copper-Doped Phosphate-Based Glass Nanozymes. ACS Appl. Mater. Interfaces 2021, 13, 11631–11645. [Google Scholar] [CrossRef]
- Wang, T.; Bai, Q.; Zhu, Z.; Xiao, H.; Jiang, F.; Du, F.; Yu, W.W.; Liu, M.; Sui, N. Graphdiyne-supported palladium-iron nanosheets: A dual-functional peroxidase mimetic nanozyme for glutathione detection and antibacterial application. Chem. Eng. J. 2021, 413, 127537. [Google Scholar] [CrossRef]
- Jia, Z.; Lv, X.; Hou, Y.; Wang, K.; Ren, F.; Xu, D.; Wang, Q.; Fan, K.; Xie, C.; Lu, X. Mussel-inspired nanozyme catalyzed conductive and self-setting hydrogel for adhesive and antibacterial bioelectronics. Bioact. Mater. 2021, 6, 2676–2687. [Google Scholar] [CrossRef]
- Zhang, S.; Hao, J.; Ding, F.; Ren, X. Nanocatalyst doped bacterial cellulose-based thermosensitive nanogel with biocatalytic function for antibacterial application. Int. J. Biol. Macromol. 2021, 195, 294–301. [Google Scholar] [CrossRef]
- Wang, X.; Sun, X.; Bu, T.; Wang, Q.; Jia, P.; Dong, M.; Wang, L. In situ fabrication of metal-organic framework derived hybrid nanozymes for enhanced nanozyme-photothermal therapy of bacteria-infected wounds. Compos. Part B Eng. 2021, 229, 109465. [Google Scholar] [CrossRef]
- Feng, Y.; Qin, J.; Zhou, Y.; Yue, Q.; Wei, J. Spherical mesoporous Fe-N-C single-atom nanozyme for photothermal and catalytic synergistic antibacterial therapy. J. Colloid Interface Sci. 2021, 606, 826–836. [Google Scholar] [CrossRef]
- Fan, Y.; Gan, X.; Zhao, H.; Zeng, Z.; You, W.; Quan, X. Multiple application of SAzyme based on carbon nitride nanorod-supported Pt single-atom for H2O2 detection, antibiotic detection and antibacterial therapy. Chem. Eng. J. 2021, 427, 131572. [Google Scholar] [CrossRef]
- Xiao, J.; Hai, L.; Li, Y.; Li, H.; Gong, M.; Wang, Z.; Tang, Z.; Deng, L.; He, D. An Ultrasmall Fe 3 O 4 -Decorated Polydopamine Hybrid Nanozyme Enables Continuous Conversion of Oxygen into Toxic Hydroxyl Radical via GSH-Depleted Cascade Redox Reactions for Intensive Wound Disinfection. Small 2021, 281, 2105465. [Google Scholar] [CrossRef]
- Hou, X.; Zeng, H.; Chi, X.; Hu, X. Pathogen Receptor Membrane-Coating Facet Structures Boost Nanomaterial Immune Escape and Antibacterial Performance. Nano Lett. 2021, 21, 9966–9975. [Google Scholar] [CrossRef]
- Gong, M.; Xiao, J.; Li, H.; Hai, L.; Yang, K.; Li, J.; Wang, Z.; Deng, L.; He, D. Magnetically retained and glucose-fueled hydroxyl radical nanogenerators for H2O2-self-supplying chemodynamic therapy of wound infections. Mater. Sci. Eng. C 2021, 131, 112522. [Google Scholar] [CrossRef]
- Wang, X.; Shi, Q.; Zha, Z.; Zhu, D.; Zheng, L.; Shi, L.; Wei, X.; Lian, L.; Wu, K.; Cheng, L. Copper single-atom catalysts with photothermal performance and enhanced nanozyme activity for bacteria-infected wound therapy. Bioact. Mater. 2021, 6, 4389–4401. [Google Scholar] [CrossRef]
- Zhu, X.; Chen, X.; Jia, Z.; Huo, D.; Liu, Y.; Liu, J. Cationic chitosan@Ruthenium dioxide hybrid nanozymes for photothermal therapy enhancing ROS-mediated eradicating multidrug resistant bacterial infection. J. Colloid Interface Sci. 2021, 603, 615–632. [Google Scholar] [CrossRef]
- Liu, Y.; Huang, Y.; Kim, D.; Ren, Z.; Oh, M.J.; Cormode, D.P.; Hara, A.T.; Zero, D.T.; Koo, H. Ferumoxytol Nanoparticles Target Biofilms Causing Tooth Decay in the Human Mouth. Nano Lett. 2021, 21, 9442–9449. [Google Scholar] [CrossRef]
- Fuentes, K.M.; Onna, D.; Rioual, T.; Huvelle, M.A.L.; Britto, F.; Simian, M.; Sánchez-Domínguez, M.; Soler-Illia, G.J.; Bilmes, S.A. Copper upcycling by hierarchical porous silica spheres functionalized with branched polyethylenimine: Antimicrobial and catalytic applications. Microporous Mesoporous Mater. 2021, 327, 111391. [Google Scholar] [CrossRef]
- Niu, J.; Zhao, C.; Liu, C.; Ren, J.; Qu, X. Bio-Inspired Bimetallic Enzyme Mimics as Bio-Orthogonal Catalysts for Enhanced Bacterial Capture and Inhibition. Chem. Mater. 2021, 33, 8052–8058. [Google Scholar] [CrossRef]
- Cao, J.; Sun, Q.; Shen, A.-G.; Fan, B.; Hu, J.-M. Nano Au@Cu2-xS with near-infrared photothermal and peroxidase catalytic activities redefines efficient antibiofilm-oriented root canal therapy. Chem. Eng. J. 2021, 422, 130090. [Google Scholar] [CrossRef]
- Shan, J.; Li, X.; Yang, K.; Xiu, W.; Wen, Q.; Zhang, Y.; Yuwen, L.; Weng, L.; Teng, Z.; Wang, L. Efficient Bacteria Killing by Cu2WS4 Nanocrystals with Enzyme-like Properties and Bacteria-Binding Ability. ACS Nano 2019, 13, 13797–13808. [Google Scholar] [CrossRef]
- Wang, Y.; Chen, C.; Zhang, D.; Wang, J. Bifunctionalized novel co-v mmo nanowires: Intrinsic oxidase and peroxidase like catalytic activities for antibacterial application. Appl. Catal. B-Environ. 2020, 261, 118256. [Google Scholar] [CrossRef]
- Ma, W.; Zhang, T.; Li, R.; Niu, Y.; Yang, X.; Liu, J.; Xu, Y.; Li, C.M. Bienzymatic synergism of vanadium oxide nanodots to efficiently eradicate drug-resistant bacteria during wound healing in vivo. J. Colloid Interface Sci. 2020, 559, 313–323. [Google Scholar] [CrossRef]
- Wang, W.; Li, B.; Yang, H.; Lin, Z.; Chen, L.; Li, Z.; Ge, J.; Zhang, T.; Xia, H.; Li, L.; et al. Efficient elimination of multidrug-resistant bacteria using copper sulfide nanozymes anchored to graphene oxide nanosheets. Nano Res. 2020, 13, 2156–2164. [Google Scholar] [CrossRef]
- Wang, J.; Wang, Y.; Zhang, D.; Xu, C.J.; Xing, R.E. Dual response mimetic enzyme of novel co4s3/co3o4 composite nanotube for antibacterial application. J. Hazard. Mater. 2020, 392, 122278. [Google Scholar] [CrossRef]
- Shan, J.; Yang, K.; Xiu, W.; Qiu, Q.; Dai, S.; Yuwen, L.; Weng, L.; Teng, Z.; Wang, L. Cu 2 MoS 4 Nanozyme with NIR-II Light Enhanced Catalytic Activity for Efficient Eradication of Multidrug-Resistant Bacteria. Small 2020, 16, 2001099. [Google Scholar] [CrossRef]
- Xu, M.; Hu, Y.; Xiao, Y.; Zhang, Y.; Sun, K.; Wu, T.; Lv, N.; Wang, W.; Ding, W.; Li, F.; et al. Near-Infrared-Controlled Nanoplatform Exploiting Photothermal Promotion of Peroxidase-like and OXD-like Activities for Potent Antibacterial and Anti-biofilm Therapies. ACS Appl. Mater. Interfaces 2020, 12, 50260–50274. [Google Scholar] [CrossRef]
- Li, Z.; Zhang, Y.; Huang, D.; Huang, L.; Zhang, H.; Li, N.; Wang, M. Through quorum sensing, Pseudomonas aeruginosa resists noble metal-based nanomaterials toxicity. Environ. Pollut. 2021, 269, 116138. [Google Scholar] [CrossRef]
- Mu, Q.; Sun, Y.; Guo, A.; Xu, X.; Qin, B.; Cai, A. A bifunctionalized NiCo2O4-Au composite: Intrinsic peroxidase and oxidase catalytic activities for killing bacteria and disinfecting wound. J. Hazard. Mater. 2021, 402, 123939. [Google Scholar] [CrossRef]
- Sun, X.; Dong, M.; Guo, Z.; Zhang, H.; Wang, J.; Jia, P.; Bu, T.; Liu, Y.; Li, L.; Wang, L. Multifunctional chitosan-copper-gallic acid based antibacterial nanocomposite wound dressing. Int. J. Biol. Macromol. 2021, 167, 10–22. [Google Scholar] [CrossRef]
- Yan, L.X.; Wang, B.B.; Zhao, X.; Chen, L.J.; Yan, X.P. A ph-responsive persistent luminescence nanozyme for selective imaging and killing of helicobacter pylori and common resistant bacteria. ACS Appl. Mater. Interfaces 2021, 13, 60955–60965. [Google Scholar] [CrossRef]
- Zhou, Y.; Chen, Z.; Zeng, S.; Wang, C.; Li, W.; Wang, M.; Wang, X.; Zhou, X.; Zhao, X.; Ren, L. Optimization of Nanostructured Copper Sulfide to Achieve Enhanced Enzyme-Mimic Activities for Improving Anti-Infection Performance. ACS Appl. Mater. Interfaces 2021, 13, 53659–53670. [Google Scholar] [CrossRef]
- Zhuang, Q.-Q.; Deng, Q.; He, S.-B.; Chen, Q.-Q.; Peng, H.-P.; Deng, H.-H.; Xia, X.-H.; Chen, W. Bifunctional cupric oxide nanoparticle-catalyzed self-cascade oxidation reactions of ascorbic acid for bacterial killing and wound disinfection. Compos. Part B Eng. 2021, 222, 109074. [Google Scholar] [CrossRef]
- Wang, P.; Peng, L.; Lin, J.; Li, Y.; Luo, Q.; Jiang, S.; Tian, H.; Zhang, Y.; Liu, X.; Liu, J. Enzyme hybrid virus-like hollow mesoporous CuO adhesive hydrogel spray through glucose-activated cascade reaction to efficiently promote diabetic wound healing. Chem. Eng. J. 2021, 415, 128901. [Google Scholar] [CrossRef]
- Xu, Q.; Hua, Y.; Zhang, Y.; Lv, M.; Wang, H.; Pi, Y.; Xie, J.; Wang, C.; Yong, Y. A Biofilm Microenvironment-Activated Single-Atom Iron Nanozyme with NIR-Controllable Nanocatalytic Activities for Synergetic Bacteria-Infected Wound Therapy. Adv. Heal. Mater. 2021, 10, 2101374. [Google Scholar] [CrossRef]
- Zhong, Y.Y.; Wang, T.T.; Lao, Z.T.; Lu, M.L.; Liang, S.; Cui, X.P.; Li, Q.L.; Zhao, S.Q. Au-au/iro2@cu(paba) reactor with tandem enzyme-mimicking catalytic activity for organic dye degradation and antibacterial application. ACS Appl. Mater. Interfaces 2021, 13, 21680–21692. [Google Scholar] [CrossRef]
- Yang, Z.; Fu, X.; Ma, D.; Wang, Y.; Peng, L.; Shi, J.; Sun, J.; Gan, X.; Deng, Y.; Yang, W. Growth Factor-Decorated Ti 3 C 2 MXene/MoS 2 2D Bio-Heterojunctions with Quad-Channel Photonic Disinfection for Effective Regeneration of Bacteria-Invaded Cutaneous Tissue. Small 2021, 17, 2103993. [Google Scholar] [CrossRef]
- Wang, L.; Gao, F.; Wang, A.; Chen, X.; Li, H.; Zhang, X.; Zheng, H.; Ji, R.; Li, B.; Yu, X.; et al. Defect-Rich Adhesive Molybdenum Disulfide/rGO Vertical Heterostructures with Enhanced Nanozyme Activity for Smart Bacterial Killing Application. Adv. Mater. 2020, 32, 2005423. [Google Scholar] [CrossRef]
- Xi, J.Q.; Wei, G.; An, L.F.; Xu, Z.B.; Xu, Z.L.; Fan, L.; Gao, L.Z. Copper/carbon hybrid nanozyme: Tuning catalytic activity by the copper state for antibacterial therapy. Nano Lett. 2019, 19, 7645–7654. [Google Scholar] [CrossRef]
- Li, Y.; Fu, R.; Duan, Z.; Zhu, C.; Fan, D. Adaptive Hydrogels Based on Nanozyme with Dual-Enhanced Triple Enzyme-Like Activities for Wound Disinfection and Mimicking Antioxidant Defense System. Adv. Heal. Mater. 2021, 11, 2101849. [Google Scholar] [CrossRef]
- Li, C.; Sun, Y.; Li, X.; Fan, S.; Liu, Y.; Jiang, X.; Boudreau, M.D.; Pan, Y.; Tian, X.; Yin, J.-J. Bactericidal effects and accelerated wound healing using Tb4O7 nanoparticles with intrinsic oxidase-like activity. J. Nanobiotechnol. 2019, 17, 54. [Google Scholar] [CrossRef]
- Cai, T.; Fang, G.; Tian, X.; Yin, J.-J.; Chen, C.; Ge, C. Optimization of Antibacterial Efficacy of Noble-Metal-Based Core-Shell Nanostructures and Effect of Natural Organic Matter. ACS Nano 2019, 13, 12694–12702. [Google Scholar] [CrossRef]
- Wang, J.; Wang, Y.; Zhang, D.; Chen, C. Intrinsic oxidase-like nanoenzyme co4s3/co(oh)(2) hybrid nanotubes with broad-spectrum antibacterial activity. ACS Appl. Mater. Interfaces 2020, 12, 29614–29624. [Google Scholar] [CrossRef]
- Zhang, W.; Zhao, Y.; Wang, W.; Peng, J.; Li, Y.; Shangguan, Y.; Ouyang, G.; Xu, M.; Wang, S.; Wei, J.; et al. Colloidal Surface Engineering: Growth of Layered Double Hydroxides with Intrinsic Oxidase-Mimicking Activities to Fight against Bacterial Infection in Wound Healing. Adv. Heal. Mater. 2020, 9, 2000092. [Google Scholar] [CrossRef]
- Sharma, S.; Chakraborty, N.; Jha, D.; Gautam, H.K.; Roy, I. Robust dual modality antibacterial action using silver-Prussian blue nanoscale coordination polymer. Mater. Sci. Eng. C 2020, 113, 110982. [Google Scholar] [CrossRef] [PubMed]
- Gao, F.; Shao, T.; Yu, Y.; Xiong, Y.; Yang, L. Surface-bound reactive oxygen species generating nanozymes for selective antibacterial action. Nat. Commun. 2021, 12, 745. [Google Scholar] [CrossRef]
- Zhang, L.F.; Zhang, L.; Deng, H.; Li, H.; Tang, W.T.; Guan, L.Y.; Qiu, Y.; Donovan, M.J.; Chen, Z.; Tan, W.H. In vivo activation of ph-responsive oxidase-like graphitic nanozymes for selective killing of helicobacter pylori. Nat. Commun. 2021, 12, 2002. [Google Scholar] [CrossRef]
- Dong, M.; Sun, X.; Bu, T.; Zhang, H.; Wang, J.; He, K.; Li, L.; Li, Z.; Wang, L. 3D/2D TMSs/TiO2 nanofibers heterojunctions for photodynamic-photothermal and oxidase-like synergistic antibacterial therapy co-driven by VIS and NIR biowindows. Compos. Part B Eng. 2021, 230, 109498. [Google Scholar] [CrossRef]
- Sun, D.; Pang, X.; Cheng, Y.; Ming, J.; Xiang, S.; Zhang, C.; Lv, P.; Chu, C.; Chen, X.; Liu, G.; et al. Ultrasound-Switchable Nanozyme Augments Sonodynamic Therapy against Multidrug-Resistant Bacterial Infection. ACS Nano 2020, 14, 2063–2076. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Tian, F.; Chang, J.; Bai, X.; Yuan, C.; Wang, C.; Neville, A. Haloperoxidase Mimicry by CeO2–x Nanorods of Different Aspect Ratios for Antibacterial Performance. ACS Sustain. Chem. Eng. 2020, 8, 6744–6752. [Google Scholar] [CrossRef]
- Frerichs, H.; Putz, E.; Pfitzner, F.; Reich, T.; Gazanis, A.; Panthofer, M.; Hartmann, J.; Jegel, O.; Heermann, R.; Tremel, W. Nanocomposite antimicrobials prevent bacterial growth through the enzyme-like activity of bi-doped cerium dioxide (ce1-xbixo2-delta). Nanoscale 2020, 12, 21344–21358. [Google Scholar] [CrossRef] [PubMed]
- Xi, J.; An, L.; Wei, G.; Huang, Y.; Li, D.; Fan, L.; Gao, L. Photolysis of methicillin-resistant Staphylococcus aureus using Cu-doped carbon spheres. Biomater. Sci. 2020, 8, 6225–6234. [Google Scholar] [CrossRef] [PubMed]
- Gao, L.; Giglio, K.M.; Nelson, J.L.; Sondermann, H.; Travis, A.J. Ferromagnetic nanoparticles with peroxidase-like activity enhance the cleavage of biological macromolecules for biofilm elimination. Nanoscale 2014, 6, 2588–2593. [Google Scholar] [CrossRef] [Green Version]
- Loo, A.E.K.; Wong, Y.T.; Ho, R.; Wasser, M.; Du, T.; Ng, W.T.; Halliwell, B. Effects of Hydrogen Peroxide on Wound Healing in Mice in Relation to Oxidative Damage. PLoS ONE 2012, 7, e49215. [Google Scholar] [CrossRef] [Green Version]
- Natalio, F.; André, R.; Hartog, A.F.; Stoll, B.; Jochum, K.P.; Wever, R.; Tremel, W. Vanadium pentoxide nanoparticles mimic vanadium haloperoxidases and thwart biofilm formation. Nat. Nanotechnol. 2012, 7, 530–535. [Google Scholar] [CrossRef]
- Shi, S.; Wu, S.; Shen, Y.; Zhang, S.; Xiao, Y.; He, X.; Gong, J.; Farnell, Y.; Tang, Y.; Huang, Y.; et al. Iron oxide nanozyme suppresses intracellular Salmonella Enteritidis growth and alleviates infection in vivo. Theranostics 2018, 8, 6149–6162. [Google Scholar] [CrossRef]
- Karim, N.; Singh, M.; Weerathunge, P.; Bian, P.; Zheng, R.; Dekiwadia, C.; Ahmed, T.; Walia, S.; Della Gaspera, E.; Singh, S.; et al. Visible-Light-Triggered Reactive-Oxygen-Species-Mediated Antibacterial Activity of Peroxidase-Mimic CuO Nanorods. ACS Appl. Nano Mater. 2018, 1, 1694–1704. [Google Scholar] [CrossRef]
- Roudbaneh, S.Z.K.; Kahbasi, S.; Sohrabi, M.J.; Hasan, A.; Salihi, A.; Mirzaie, A.; Niyazmand, A.; Nanakali, N.M.Q.; Shekha, M.S.; Aziz, F.M.; et al. Albumin binding, antioxidant and antibacterial effects of cerium oxide nanoparticles. J. Mol. Liq. 2019, 296, 111839. [Google Scholar] [CrossRef]
- Sun, H.; Gao, N.; Dong, K.; Ren, J.; Qu, X. Graphene Quantum Dots-Band-Aids Used for Wound Disinfection. ACS Nano 2014, 8, 6202–6210. [Google Scholar] [CrossRef]
- Sun, M.; Qian, H.; Liu, J.; Li, Y.; Pang, S.; Xu, M.; Zhang, J. A flexible conductive film prepared by the oriented stacking of Ag and Au/Ag alloy nanoplates and its chemically roughened surface for explosive SERS detection and cell adhesion. RSC Adv. 2017, 7, 7073–7078. [Google Scholar] [CrossRef] [Green Version]
- Yu, M.; Zhang, G.; Li, P.; Lu, H.; Tang, W.; Yang, X.; Huang, R.; Yu, F.; Wu, W.; Xiao, Y.; et al. Acid-activated ROS generator with folic acid targeting for bacterial biofilm elimination. Mater. Sci. Eng. C 2021, 127, 112225. [Google Scholar] [CrossRef]
- Vallabani, N.S.; Vinu, A.; Singh, S.; Karakoti, A. Tuning the ATP-triggered pro-oxidant activity of iron oxide-based nanozyme towards an efficient antibacterial strategy. J. Colloid Interface Sci. 2020, 567, 154–164. [Google Scholar] [CrossRef]
- Naha, P.C.; Liu, Y.; Hwang, G.; Huang, Y.; Gubara, S.; Jonnakuti, V.; Simon-Soro, A.; Kim, D.; Gao, L.; Koo, H.; et al. Dextran-coated iron oxide nanoparticles as biomimetic catalysts for localized and ph-activated biofilm disruption. ACS Nano 2019, 13, 4960–4971. [Google Scholar] [CrossRef]
- Tao, Y.; Ju, E.; Ren, J.; Qu, X. Bifunctionalized Mesoporous Silica-Supported Gold Nanoparticles: Intrinsic Oxidase and Peroxidase Catalytic Activities for Antibacterial Applications. Adv. Mater. 2015, 27, 1097–1104. [Google Scholar] [CrossRef]
- Fang, G.; Li, W.; Shen, X.; Perez-Aguilar, J.M.; Chong, Y.; Gao, X.; Chai, Z.; Chen, C.; Ge, C.; Zhou, R. Differential Pd-nanocrystal facets demonstrate distinct antibacterial activity against Gram-positive and Gram-negative bacteria. Nat. Commun. 2018, 9, 129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, S.; Huang, J.; Zhang, Q.; Zhao, W.; Xu, Z.; Zhang, W. Bamboo-Like Nanozyme Based on Nitrogen-Doped Carbon Nanotubes Encapsulating Cobalt Nanoparticles for Wound Antibacterial Applications. Adv. Funct. Mater. 2021, 31, 2105198. [Google Scholar] [CrossRef]
- Allentoft, M.E.; Collins, M.; Harker, D.; Haile, J.; Oskam, C.; Hale, M.L.; Campos, P.; Samaniego, J.A.; Gilbert, M.; Willerslev, E.; et al. The Half-Life of DNA in bone: Measuring Decay Kinetics in 158 Dated Fossils. In Proceedings of the Royal Society of London; Series B: Biological Sciences; The Royal Society: London, UK, 2012; Volume 279, pp. 4724–4733. [Google Scholar]
- Luong, T.K.N.; Govaerts, I.; Robben, J.; Shestakova, P.; Parac-Vogt, T.N. Polyoxometalates as artificial nucleases: Hydrolytic cleavage of DNA promoted by a highly negatively charged ZrIV-substituted Keggin polyanion. Chem. Commun. 2017, 53, 617–620. [Google Scholar] [CrossRef] [PubMed]
- Mikkola, S.; Lönnberg, T.; Lönnberg, H. Phosphodiester models for cleavage of nucleic acids. Beilstein J. Org. Chem. 2018, 14, 803–837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Furukawa, T.; Jikumaru, A.; Ueno, T.; Sei, K. Inactivation Effect of Antibiotic-Resistant Gene Using Chlorine Disinfection. Water 2017, 9, 547. [Google Scholar] [CrossRef]
- Michael-Kordatou, I.; Karaolia, P.; Fatta-Kassinos, D. The role of operating parameters and oxidative damage mechanisms of advanced chemical oxidation processes in the combat against antibiotic-resistant bacteria and resistance genes present in urban wastewater. Water Res. 2018, 129, 208–230. [Google Scholar] [CrossRef]
- Giannakis, S.; Le, T.-T.M.; Entenza, J.M.; Pulgarin, C. Solar photo-Fenton disinfection of 11 antibiotic-resistant bacteria (ARB) and elimination of representative AR genes. Evidence that antibiotic resistance does not imply resistance to oxidative treatment. Water Res. 2018, 143, 334–345. [Google Scholar] [CrossRef]
- Massoud, S.S.; Perkins, R.S.; Louka, F.R.; Xu, W.; Le Roux, A.; Dutercq, Q.; Fischer, R.C.; Mautner, F.A.; Handa, M.; Hiraoka, Y.; et al. Efficient hydrolytic cleavage of plasmid DNA by chloro-cobalt(ii) complexes based on sterically hindered pyridyl tripod tetraamine ligands: Synthesis, crystal structure and DNA cleavage. Dalton Trans. 2014, 43, 10086–10103. [Google Scholar] [CrossRef] [Green Version]
- Soler, M.; Figueras, E.; Serrano-Plana, J.; González-Bártulos, M.; Massaguer, A.; Company, A.; Martínez, M.Á.; Malina, J.; Brabec, V.; Feliu, L.; et al. Design, Preparation, and Characterization of Zn and Cu Metallopeptides Based on Tetradentate Aminopyridine Ligands Showing Enhanced DNA Cleavage Activity. Inorg. Chem. 2015, 54, 10542–10558. [Google Scholar] [CrossRef]
- Salvio, R.; Volpi, S.; Cacciapaglia, R.; Sansone, F.; Mandolini, L.; Casnati, A. Upper Rim Bifunctional cone-Calix[4 ]arenes Based on a Ligated Metal Ion and a Guanidinium Unit as DNAase and RNAase Mimics. J. Org. Chem. 2016, 81, 4728–4735. [Google Scholar] [CrossRef]
- Piovezan, C.; Jovito, R.; Bortoluzzi, A.J.; Terenzi, H.; Fischer, F.L.; Severino, P.C.; Pich, C.T.; Azzolini, G.G.; Peralta, R.A.; Rossi, L.M.; et al. Heterodinuclear (feznii)-zn-iii-bioinspired complex supported on 3-aminopropyl silica. Efficient hydrolysis of phosphate diester bonds. Inorg. Chem. 2010, 49, 2580–2582. [Google Scholar] [CrossRef] [PubMed]
- Jiang, R.; Xin, Z.; Xu, S.; Shi, H.; Yang, H.; Song, L.; Yan, S.; Luan, S.; Yin, J.; Khan, A.F.; et al. Enzyme-mimicking polymer brush-functionalized surface for combating biomaterial-associated infections. Appl. Surf. Sci. 2017, 423, 869–880. [Google Scholar] [CrossRef]
- Liu, Z.; Wang, F.; Ren, J.; Qu, X. A series of MOF/Ce-based nanozymes with dual enzyme-like activity disrupting biofilms and hindering recolonization of bacteria. Biomaterials 2019, 208, 21–31. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.-Y.; Hua, X.-W.; Wu, F.-G.; Li, B.; Liu, P.; Gu, N.; Wang, Z.; Chen, Z. Synthesis of Ultrastable Copper Sulfide Nanoclusters via Trapping the Reaction Intermediate: Potential Anticancer and Antibacterial Applications. ACS Appl. Mater. Interfaces 2015, 7, 7082–7092. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.-C.; Deokar, A.R.; Liao, J.-H.; Shih, P.-Y.; Ling, Y.-C. Graphene-Based Photothermal Agent for Rapid and Effective Killing of Bacteria. ACS Nano 2013, 7, 1281–1290. [Google Scholar] [CrossRef] [PubMed]
- Natan, M.; Edin, F.; Perkas, N.; Yacobi, G.; Perelshtein, I.; Segal, E.; Homsy, A.; Laux, E.; Keppner, H.; Rask-Andersen, H.; et al. Two are better than one: Combining zno and mgf2 nanoparticles reduces streptococcus pneumoniae and staphylococcus aureus biofilm formation on cochlear implants. Adv. Funct. Mater. 2016, 26, 2473–2481. [Google Scholar] [CrossRef]
- Yin, M.; Li, Z.; Ju, E.; Wang, Z.; Dong, K.; Ren, J.; Qu, X. Multifunctional upconverting nanoparticles for near-infrared triggered and synergistic antibacterial resistance therapy. Chem. Commun. 2014, 50, 10488–10490. [Google Scholar] [CrossRef]
- Dai, T.; Huang, Y.-Y.; Hamblin, M.R. Photodynamic therapy for localized infections—State of the art. Photodiagn. Photodyn. Ther. 2009, 6, 170–188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, B.; Wang, M.; Mikhailovsky, A.; Wang, S.; Bazan, G.C. A Membrane-Intercalating Conjugated Oligoelectrolyte with High-Efficiency Photodynamic Antimicrobial Activity. Angew. Chem. Int. Ed. 2017, 56, 5031–5034. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Shi, S.; Wang, Y.; Yu, S.; Zhu, W.; Zhang, X.; Zhang, D.; Yang, B.; Wang, X.; Wang, J. Versatile molybdenum disulfide based antibacterial composites for in vitro enhanced sterilization and in vivo focal infection therapy. Nanoscale 2016, 8, 11642–11648. [Google Scholar] [CrossRef] [PubMed]
- Yin, W.; Yu, J.; Lv, F.; Yan, L.; Zheng, L.R.; Gu, Z.; Zhao, Y. Functionalized Nano-MoS2 with Peroxidase Catalytic and Near-Infrared Photothermal Activities for Safe and Synergetic Wound Antibacterial Applications. ACS Nano 2016, 10, 11000–11011. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Mechanisms of bacterial resistance. The figure shows a brief overview of intrinsic resistance mechanisms. Firstly, bacteria acquire drug-resistance through gene mutation at the genetic level. The mutated gene can also spread through vertical and horizontal transmission (herein, plasmid for example). Besides, biochemical mechanisms are a more common style of antidrug upon target change, efflux or inactivation. Moreover, the formation of biofilm prevents bacteria from reaching the antibiotic and enhances drug resistance.
Figure 1.
Mechanisms of bacterial resistance. The figure shows a brief overview of intrinsic resistance mechanisms. Firstly, bacteria acquire drug-resistance through gene mutation at the genetic level. The mutated gene can also spread through vertical and horizontal transmission (herein, plasmid for example). Besides, biochemical mechanisms are a more common style of antidrug upon target change, efflux or inactivation. Moreover, the formation of biofilm prevents bacteria from reaching the antibiotic and enhances drug resistance.

Figure 2.
Schematic of the antibacterial mechanism of several typical enzybiotics. (a) Schematic of the action of the lysozyme on Gram-positive and Gram-negative bacteria. (b) Proteinase can degrade proteins by hydrolyzing peptide bonds. (c) Nuclease mainly consists of DNase and RNase, which degrade DNA and RNA, respectively. This figure uses DNase hydrolysis of phosphodiester bound as an example. The black arrow is the hydrolysis site of DNase and the red arrow is the hydrolysis site of RNase (e.g., RNase A family).
Figure 2.
Schematic of the antibacterial mechanism of several typical enzybiotics. (a) Schematic of the action of the lysozyme on Gram-positive and Gram-negative bacteria. (b) Proteinase can degrade proteins by hydrolyzing peptide bonds. (c) Nuclease mainly consists of DNase and RNase, which degrade DNA and RNA, respectively. This figure uses DNase hydrolysis of phosphodiester bound as an example. The black arrow is the hydrolysis site of DNase and the red arrow is the hydrolysis site of RNase (e.g., RNase A family).

Figure 3.
Mechanisms of enzybiotics eradicating bacterial biofilm. The biofilm consists of a combination of glycoproteins, carbohydrates, lipids, eDNA, and bacterial cell. When treating with enzybiotics, different enzymes bind to their respective targets for degradation. In this figure, lysins, proteases and nucleases bind to polysaccharides, proteins and eDNA in the biofilm and degrade them, respectively, thus promoting the disintegration of the biofilm. Besides, lysins also decompose the peptidoglycan, one of the important components of bacteria cell walls, to kill bacteria, especially for Gram-positive species. The peptidoglycan of Gram-negative bacteria is difficult to degrade due to the protection of the outer membrane, while modified lysins may take effect.
Figure 3.
Mechanisms of enzybiotics eradicating bacterial biofilm. The biofilm consists of a combination of glycoproteins, carbohydrates, lipids, eDNA, and bacterial cell. When treating with enzybiotics, different enzymes bind to their respective targets for degradation. In this figure, lysins, proteases and nucleases bind to polysaccharides, proteins and eDNA in the biofilm and degrade them, respectively, thus promoting the disintegration of the biofilm. Besides, lysins also decompose the peptidoglycan, one of the important components of bacteria cell walls, to kill bacteria, especially for Gram-positive species. The peptidoglycan of Gram-negative bacteria is difficult to degrade due to the protection of the outer membrane, while modified lysins may take effect.

Figure 4.
Schematic illustration of the catalytic reaction and antibacterial (and antibiofilm) mechanism of nanozybiotics based on nanozymes with peroxidase- or oxidase-like enzyme activities. (a) Nanozymes with peroxidase-like activity catalyze the reduction of H2O2 and produce free ·OH. (b) Nanozymes with oxidase-like activity catalyze O2 to 1O2, even single oxygen atoms. Both ·OH and 1O2 are strong oxidant, which oxidize the substrate (S) to ox-substrate (Sox), for example, membrane lipid. (c) Nanozymes with peroxidase- or oxidase-like activity destroy the membrane structure or degrade the biofilm matrix to kill bacteria.
Figure 4.
Schematic illustration of the catalytic reaction and antibacterial (and antibiofilm) mechanism of nanozybiotics based on nanozymes with peroxidase- or oxidase-like enzyme activities. (a) Nanozymes with peroxidase-like activity catalyze the reduction of H2O2 and produce free ·OH. (b) Nanozymes with oxidase-like activity catalyze O2 to 1O2, even single oxygen atoms. Both ·OH and 1O2 are strong oxidant, which oxidize the substrate (S) to ox-substrate (Sox), for example, membrane lipid. (c) Nanozymes with peroxidase- or oxidase-like activity destroy the membrane structure or degrade the biofilm matrix to kill bacteria.

Figure 5.
Schematic illustration of the antibacterial and antibiofilm mechanism of nanozybiotics based on nanozymes with deoxyribonuclease-like enzyme activities. Nanozymes with deoxyribonuclease activity catalyze the decomposition of DNA from dead bacteria to prevent the dissemination of released drug-resistant genes (a) and disperse the biofilm by decomposing the eDNA, the essential structural component of biofilm (b).
Figure 5.
Schematic illustration of the antibacterial and antibiofilm mechanism of nanozybiotics based on nanozymes with deoxyribonuclease-like enzyme activities. Nanozymes with deoxyribonuclease activity catalyze the decomposition of DNA from dead bacteria to prevent the dissemination of released drug-resistant genes (a) and disperse the biofilm by decomposing the eDNA, the essential structural component of biofilm (b).

Figure 6.
Schematic illustration of the antibacterial mechanism of combination therapy using nanozybiotics. Here, for example, nanozybiotics based on peroxidase (POD)-like nanozymes can catalyze the decomposition of hydrogen peroxide (H2O2) to produce bactericidal ·OH. With the radiation of light (visible or near-infrared light), photo-activated therapy (PTT and PDT) is active with excellent hyperthermia, singlet oxygen (1O2) and more ·OH production. The produced ·OH and 1O2 interact with bacteria to induce membrane peroxidation and damage cell integrity, making bacteria more vulnerable. In addition, the alternating magnetic field (AMF) exposure also can enhance the catalytic activity of nanozymes and generate more toxic ·OH.
Figure 6.
Schematic illustration of the antibacterial mechanism of combination therapy using nanozybiotics. Here, for example, nanozybiotics based on peroxidase (POD)-like nanozymes can catalyze the decomposition of hydrogen peroxide (H2O2) to produce bactericidal ·OH. With the radiation of light (visible or near-infrared light), photo-activated therapy (PTT and PDT) is active with excellent hyperthermia, singlet oxygen (1O2) and more ·OH production. The produced ·OH and 1O2 interact with bacteria to induce membrane peroxidation and damage cell integrity, making bacteria more vulnerable. In addition, the alternating magnetic field (AMF) exposure also can enhance the catalytic activity of nanozymes and generate more toxic ·OH.


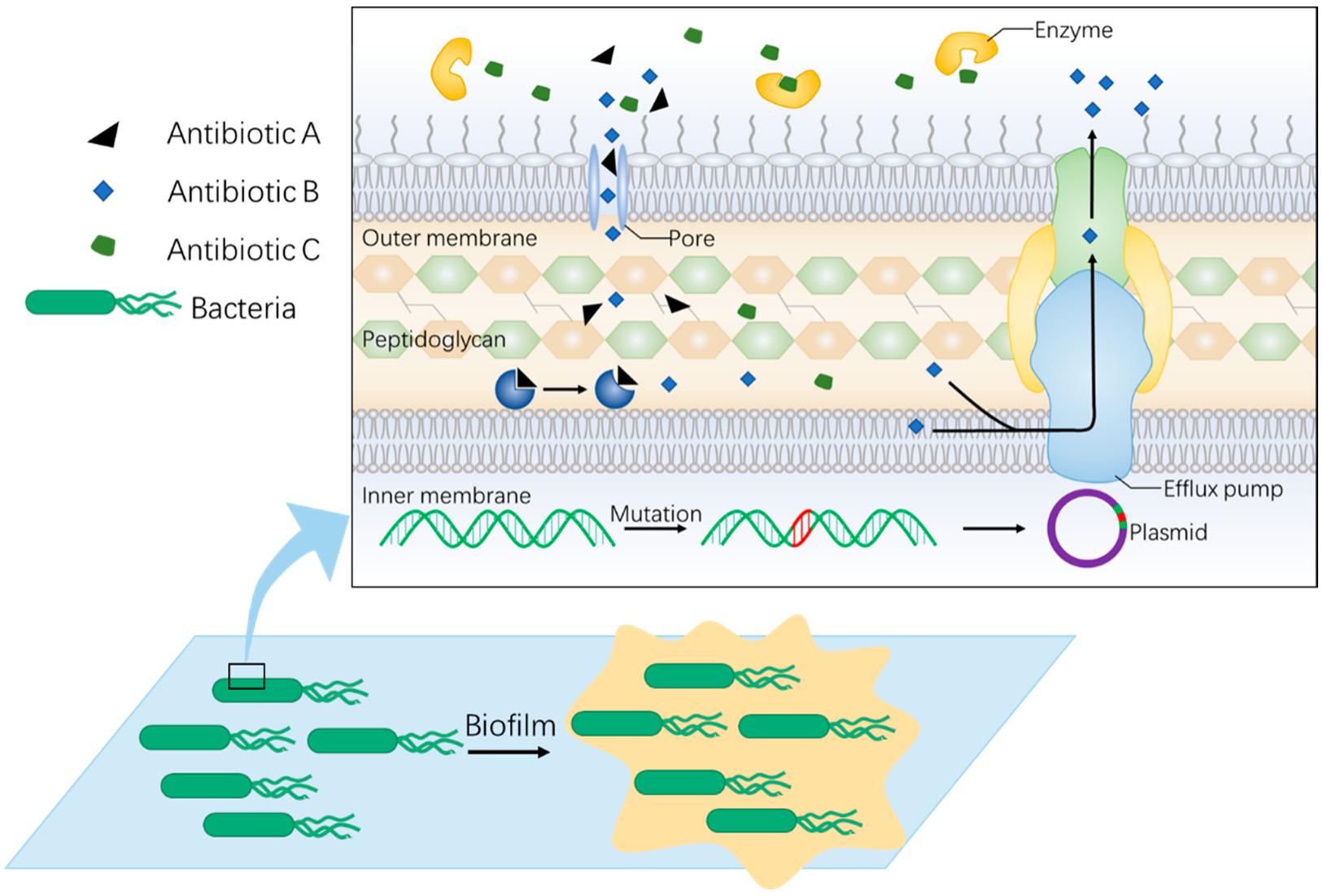{kind=link}
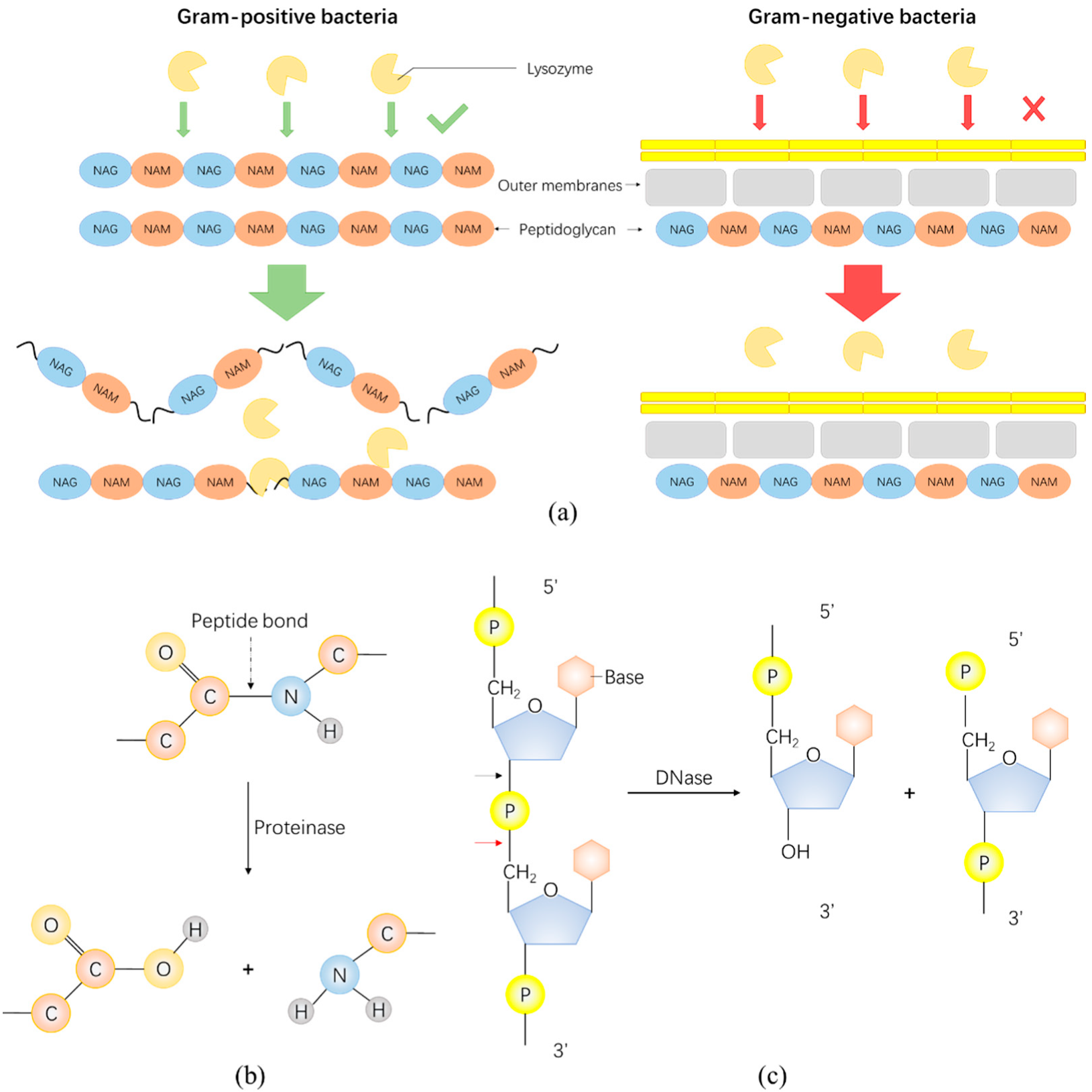{kind=link}
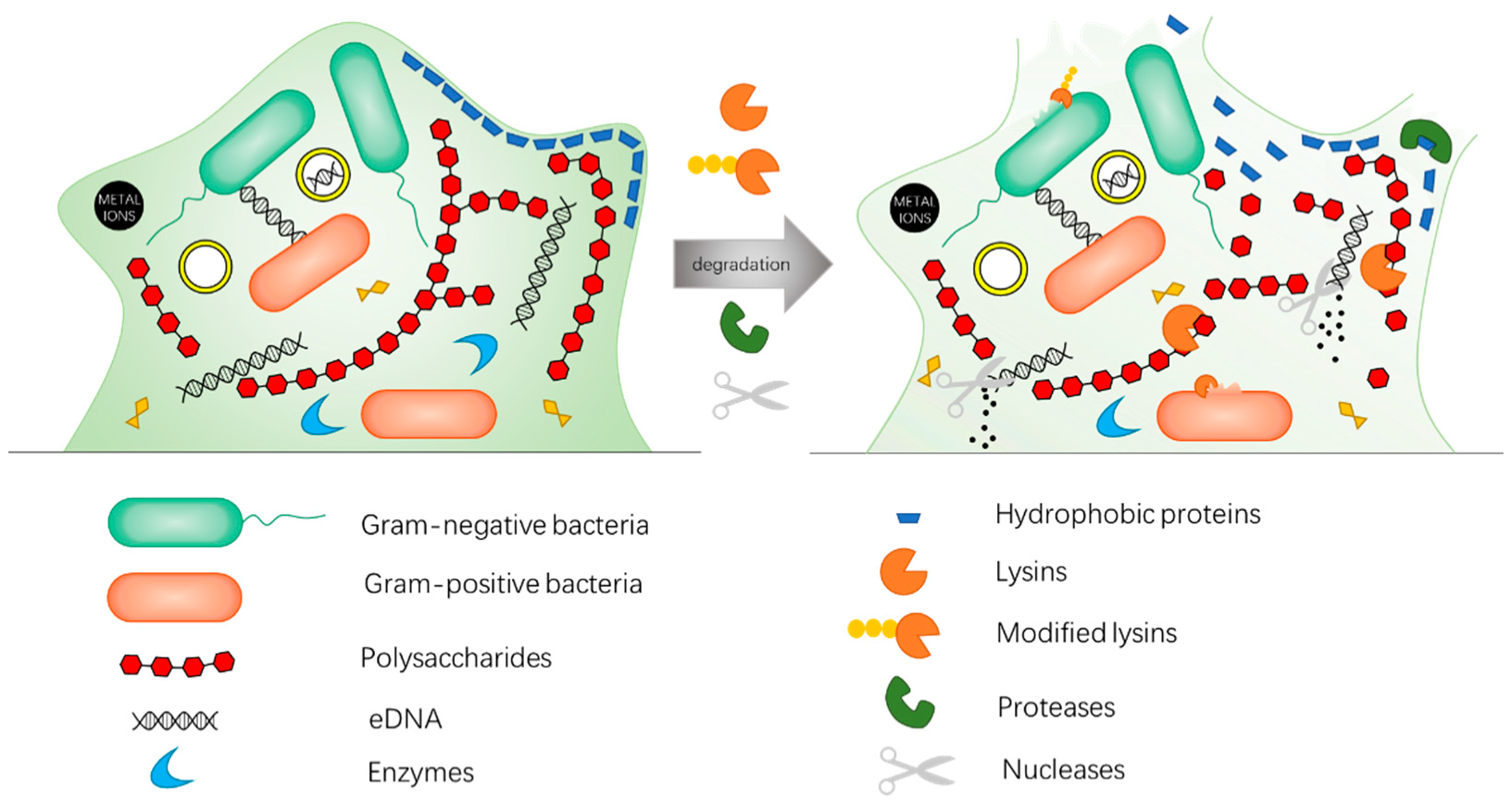{kind=link}
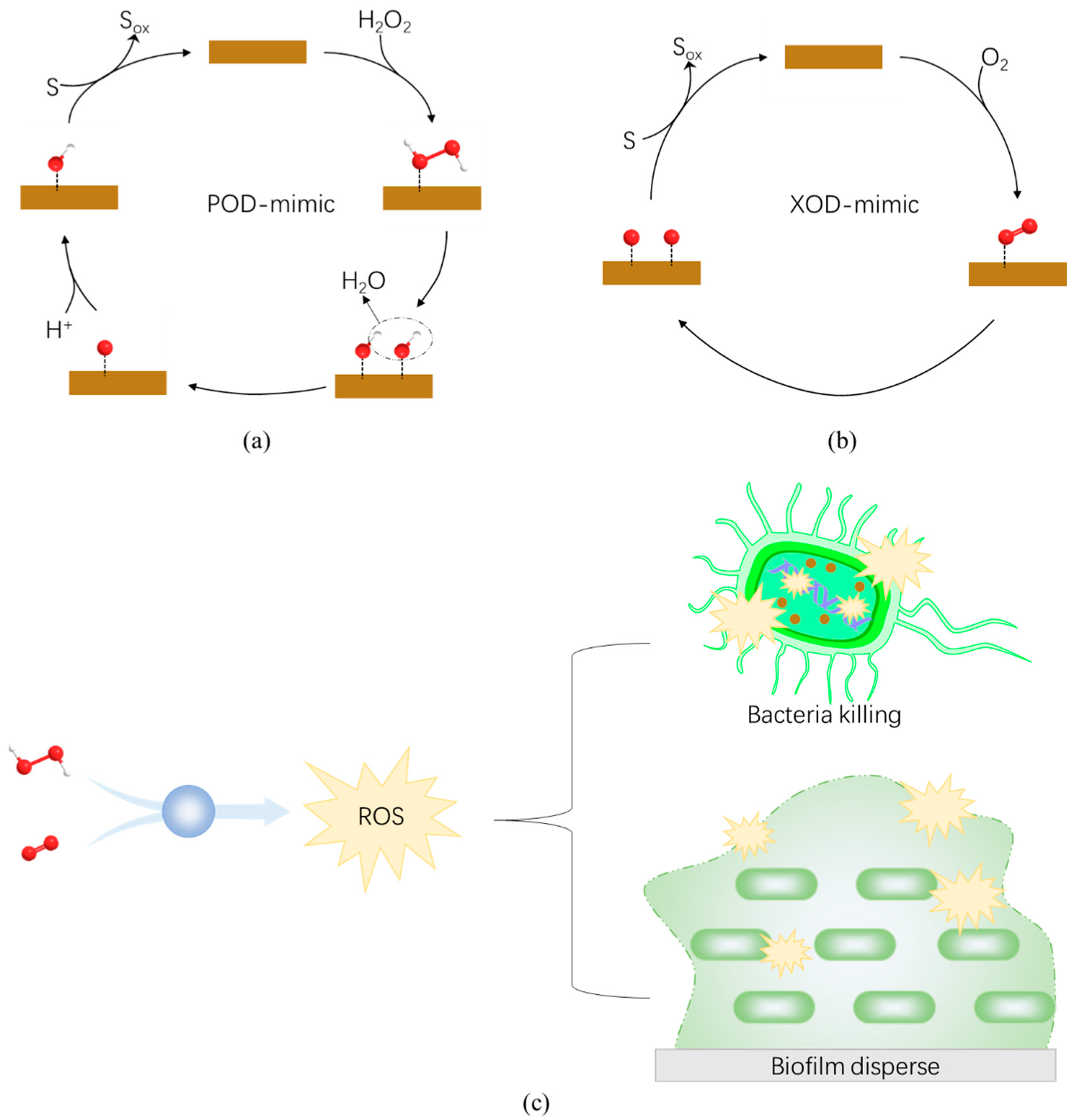{kind=link}
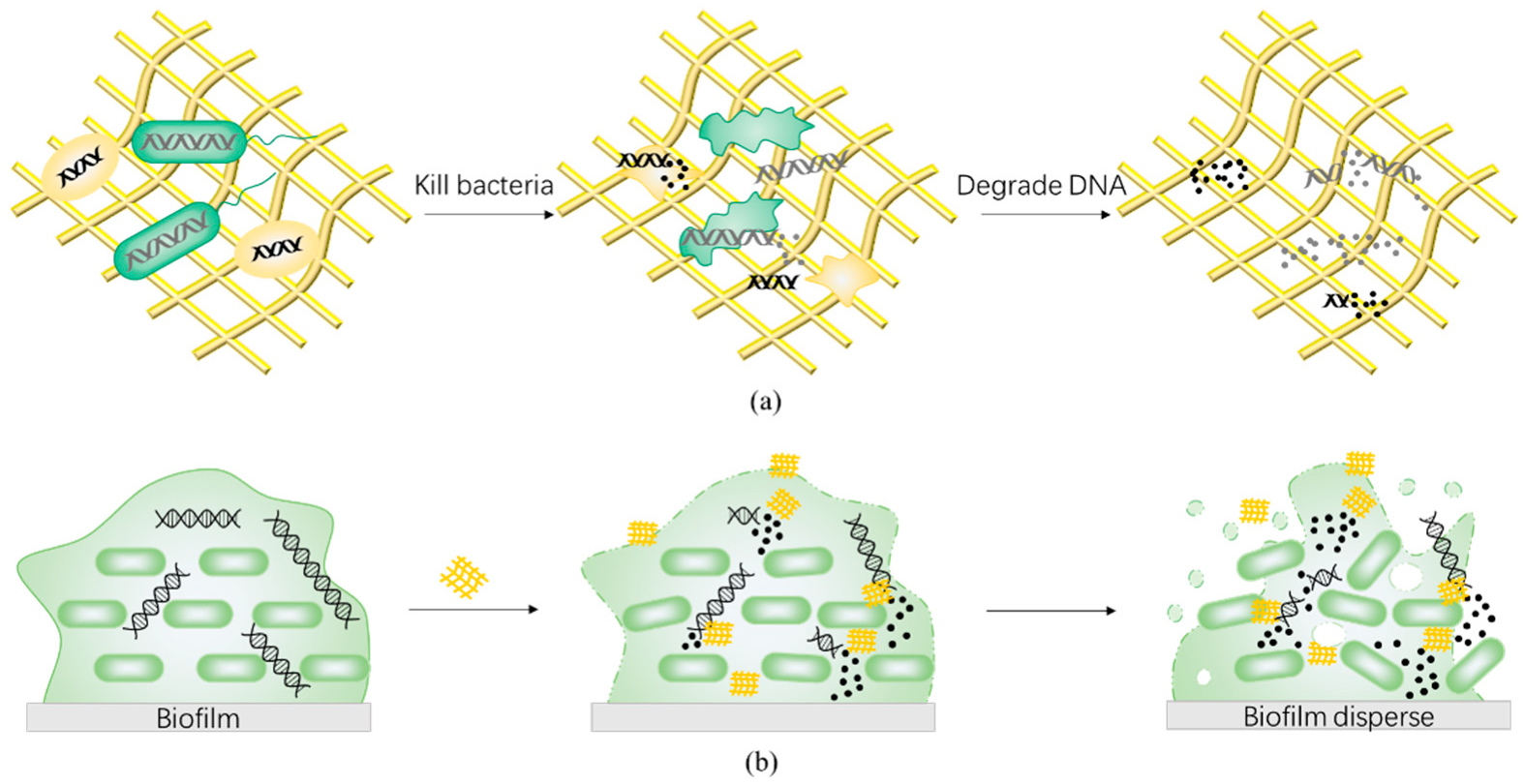{kind=link}
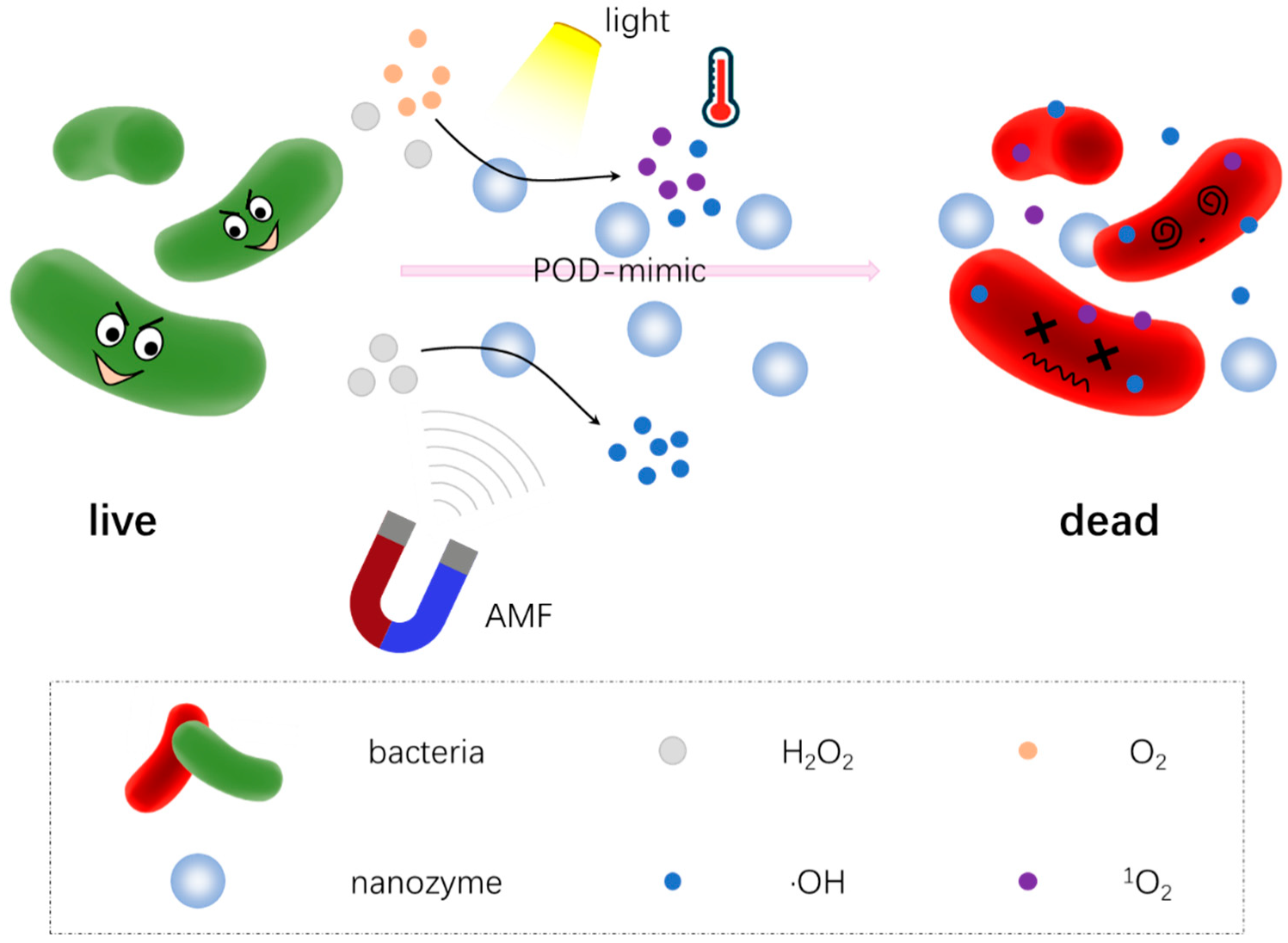{kind=link}
Table 1.
An overview of nanozybiotics as antibacterials.
| Nanozybiotics | Enzyme Like Activity | Pathogens | Ref |
|---|---|---|---|
| PMCS | Peroxidase | Pseudomonas aeruginosa (P. aeruginosa) | [48] |
| 2D Cu-TCPP(Fe) nanosheets | Peroxidase | E. coli, S. aureus | [49] |
| biohybrid CARs | Peroxidase | Streptococcus mutans UA159 | [50] |
| MoS2-hydrogel | Peroxidase | E. coli, S. aureus | [51] |
| AA@Ru@HA-MoS2 | Peroxidase | MDR S. aureus, P. aeruginosa | [52] |
| SAF NCs | Peroxidase | E. coli, S. aureus | [53] |
| hydrogel-based artificial enzyme | Peroxidase | Drug-resistant (DR) S. aureus, DR-E. coli | [54] |
| IONzymes | Peroxidase | Streptococcus mutans (S. mutans) | [55] |
| Fe3C/N-C | Peroxidase | E. coli, S. aureus | [56] |
| N-MoS2, N-WS2 NSs | Peroxidase | ampicillin resistant Escherichia coli (AmprE. coli), endospore-forming Bacillus subtilis (B. subtilis) | [57] |
| N-SCSs | Peroxidase | MDR S. aureus, E. coli | [58] |
| UsAuNPs/MOFs | Peroxidase | E. coli, S. aureus | [59] |
| FNPs | Peroxidase | Helicobacter pylori (H. pylori) | [60] |
| CuMnO2 NFs | Peroxidase | E. coli, S. aureus | [61] |
| AIronNPs | Peroxidase | E. coli, S. aureus | [62] |
| NH2-MIL-88B(Fe)-Ag | Peroxidase | E. coli, S. aureus | [63] |
| IrNPs | Peroxidase | E. coli | [64] |
| CDs@PtNPs | Peroxidase | MRSA | [65] |
| Fe3O4@MoS2-Ag | Peroxidase | E. coli, S. aureus, Bacillus subtilis (B. subtili), MRSA, Candida albicans (C. albicans) | [66] |
| Au-BNNs, Ag-BNNs | Peroxidase | E. coli, S. aureus | [67] |
| oxygenated nanodiamonds (O-NDs) | Peroxidase | Fusobacterium nucleatum (F. nucleatum), Porphyromonas gingivalis (P. gingivalis), S. sanguis | [68] |
| Dex-IONP | Peroxidase | S. mutans | [69] |
| MoO3−x NDs | Peroxidase | MRSA, ESBL-producing E.coli | [70] |
| MTex | Peroxidase | E. coli, S. aureus | [71] |
| Au/MnFe2O4 | Peroxidase | S. aureus, B. subtilis, E. faecalis, S. pyogenes | [72] |
| AuNPTs | Peroxidase | MRSA, E. coli, S. aureus | [73] |
| rough C–Fe3O4 | Peroxidase | MRSA, E. coli, S. aureus | [12] |
| Cu-PBG | Peroxidase | E. coli, S. aureus | [74] |
| PdFe/GDY | Peroxidase | E. coli, S. aureus | [75] |
| ultrasmall TA-Ag nanozyme | Peroxidase | E. coli, Staphylococcus epidermidis (S. epidermidis) | [76] |
| Cu-SA@BCNW/PNI | Peroxidase | E. coli, S. aureus | [77] |
| PEG@Zn/Pt–CN | Peroxidase | E. coli, S. aureus | [78] |
| Fe-N-C SAzyme | Peroxidase | E. coli, S. aureus | [79] |
| SA-Pt/g-C3N4-K | Peroxidase | E. coli, S. aureus, Bacillus cereus, P. aeruginosa | [80] |
| PDA/Fe3O4 | Peroxidase | E. coli, S. aureus | [81] |
| CuFeSe2 | Peroxidase | S. aureus | [82] |
| pFe3O4@GOx | Peroxidase | E. coli, S. aureus | [83] |
| Cu SASs/NPC | Peroxidase | E. coli, MRSA | [84] |
| QCS-RuO2@RBT | Peroxidase | P. aeruginosa | [85] |
| FerIONP | Peroxidase | S. mutans | [86] |
| w-SiO2/CuO | Peroxidase | E. coli | [87] |
| PdCu-Urchin | Peroxidase | E. coli, S. aureus | [88] |
| Au@Cu2−xS NPs | Peroxidase | E. faecalis, Fusobacterium nucleus | [89] |
| Cu2WS4 nanocrystals (CWS NCs) | Peroxidase, oxidase | MDR S. aureus, E. coli | [90] |
| 3CoV-400 | Peroxidase, oxidase | E. coli, Bacillus algicola, Staphylococcus sciuri (S. sciuri), Vibrio harveyi, Pseudoalteromonas | [91] |
| VOxNDs | Peroxidase, oxidase | E. coli, S. aureus | [92] |
| GO NSs, CuS/GO NC | Peroxidase, oxidase | E. coli, S. aureus, MRSA | [93] |
| Co4S3/Co3O4 NTs | Peroxidase, oxidase | E. coli, S. sciuri | [94] |
| Cu2MoS4 NPs | Peroxidase, oxidase | MDR E. coli, MDR S. aureus | [95] |
| WS2QDs | Peroxidase, oxidase | Mu50 (a vancomycin-intermediate Staphylococcus aureus reference strain), E.coli | [96] |
| Pd@NPs | Peroxidase, oxidase | S. aureus, P. aeruginosa | [97] |
| NiCo2O4-Au | Peroxidase, oxidase | E. coli, S. aureus | [98] |
| CS-Cu-GA NCs | Peroxidase, oxidase | E. coli, S. aureus | [99] |
| MSPLNP-Au-CB | Peroxidase, oxidase | Helicobacter pylori (H. pylori), MRSA | [100] |
| CSG-MX | Peroxidase, oxidase | E. coli, S. aureus | [13] |
| Cu2−xS | Peroxidase, oxidase | AmprE. coli 1, S. aureus | [101] |
| CuO NPs/AA | Peroxidase, oxidase | E. coli, S. aureus | [102] |
| HvCuO@GOx | Peroxidase, catalase | E. coli, S. aureus, E. coli with streptomycin resistance (SR-E. coli) | [103] |
| FePN SAzyme | Peroxidase, catalase | E. coli, S. aureus | [104] |
| Au-Au/IrO2@Cu (PABA) | Peroxidase, glucose oxidase (GOx) | E. coli, S. aureus | [105] |
| Ti3C2 MXene/MoS2 (MM) 2D bio-heterojunctions | Peroxidase, glutathione oxidase | E. coli, S. aureus | [106] |
| MoS2/rGO VHS | Peroxidase, oxidase, catalase | E. coli, S. aureus | [107] |
| Cu-HCSs, CuO-HCSs | Peroxidase, catalase, superoxide dismutase | Cu-HCSs: Gram-positive and negative bacteria (S. aureus, S. typhimurium, E. coli, P. aeruginosa) CuO-HCSs: Gram-negative bacteria (S. typhimurium, E. coli, P. aeruginosa) | [108] |
| CNT@MoS2 NSs | Peroxidase, superoxide, catalase | E. coli, S. aureus | [109] |
| MoS2-PDA nanozyme composite hydrogel (MPH) | Peroxidase, catalase, superoxide dismutase | E. coli, S. aureus | [15] |
| Tb4O7 NPs | Oxidase | S. aureus, E. coli | [110] |
| Pd@Ir octahedra (or cubes) | Oxidase | E. coli, S. aureus, Bacillus subtilis, Salmonella enteritidis | [111] |
| Co4S3/Co(OH)2 HNTs | Oxidase | E. coli, P. aeruginosa, S. sciuri, Bacillus | [112] |
| Mn/Ni(OH)x LDHs | Oxidase | E. coli, S. aureus | [113] |
| SPB NCPs | Oxidase | S. aureus, P. aeruginosa | [114] |
| AgPd0.38 | Oxidase | S. aureus, B. subtilis, E. coli, P. aeruginosa, MRSA | [115] |
| PtCo@Graphene | Oxidase | H. pylori | [116] |
| MoS2/TiO2 NFs | Oxidase | E. coli, S. aureus | [117] |
| Cu3/ND@G | Oxidase | E. coli | [14] |
| Pd@Pt-T790 | Catalase | MRSA | [118] |
| DMAE | DNase | S. aureus | [16] |
| PIL-Ce | DNase | E. coli, S. aureus, MRSA | [17] |
| CeO2−x nanorods | Haloperoxidase | E. coli | [119] |
| Ce1−xBixO2−δ | Haloperoxidase | P. aeruginosa, Phaeobacter gallaeciensis | [120] |
| Cu-HCSs | Nuclease/protease | MRSA | [121] |
1 Ampr E. coli: Ampicillin-resistant E. coli.
Table 2.
Comparison between enzybiotics and nanozybiotics for antibacterial application.
| Enzybiotics | Nanozybiotics | |
|---|---|---|
| Derivation | natural enzymes | nanozymes (nanomaterials) |
| Catalytic activity | peptidoglycan hydrolases, proteases, and nuclease | peroxidase, oxidase, catalase, deoxyribonuclease |
| Main antibacterial mechanism | destroy bacterial cell structure | catalyze the production of ROS |
| Application advantages | rapid and unique mode of action, high specificity of kill pathogens, low probability for bacterial resistance development and a proteinaceous nature | economical, stable, with catalytic function without additional modification, easy to integrate a variety of antibacterial strategies |
| Application disadvantages | environmentally sensitive and unstable, high cost, short half-life and immunogenicity of proteins | low enzyme activity, limited types of enzyme catalysis and complicated toxicological profile |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Zhou, C.; Wang, Q.; Jiang, J.; Gao, L. Nanozybiotics: Nanozyme-Based Antibacterials against Bacterial Resistance. Antibiotics 2022, 11, 390. https://doi.org/10.3390/antibiotics11030390
AMA Style
Zhou C, Wang Q, Jiang J, Gao L. Nanozybiotics: Nanozyme-Based Antibacterials against Bacterial Resistance. Antibiotics. 2022; 11(3):390. https://doi.org/10.3390/antibiotics11030390
Chicago/Turabian StyleZhou, Caiyu, Qian Wang, Jing Jiang, and Lizeng Gao. 2022. "Nanozybiotics: Nanozyme-Based Antibacterials against Bacterial Resistance" Antibiotics 11, no. 3: 390. https://doi.org/10.3390/antibiotics11030390
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.