
Multidrug-Resistant Helicobacter pylori Strains: A Five-Year Surveillance Study and Its Genome Characteristics
 , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. Demography of Patients
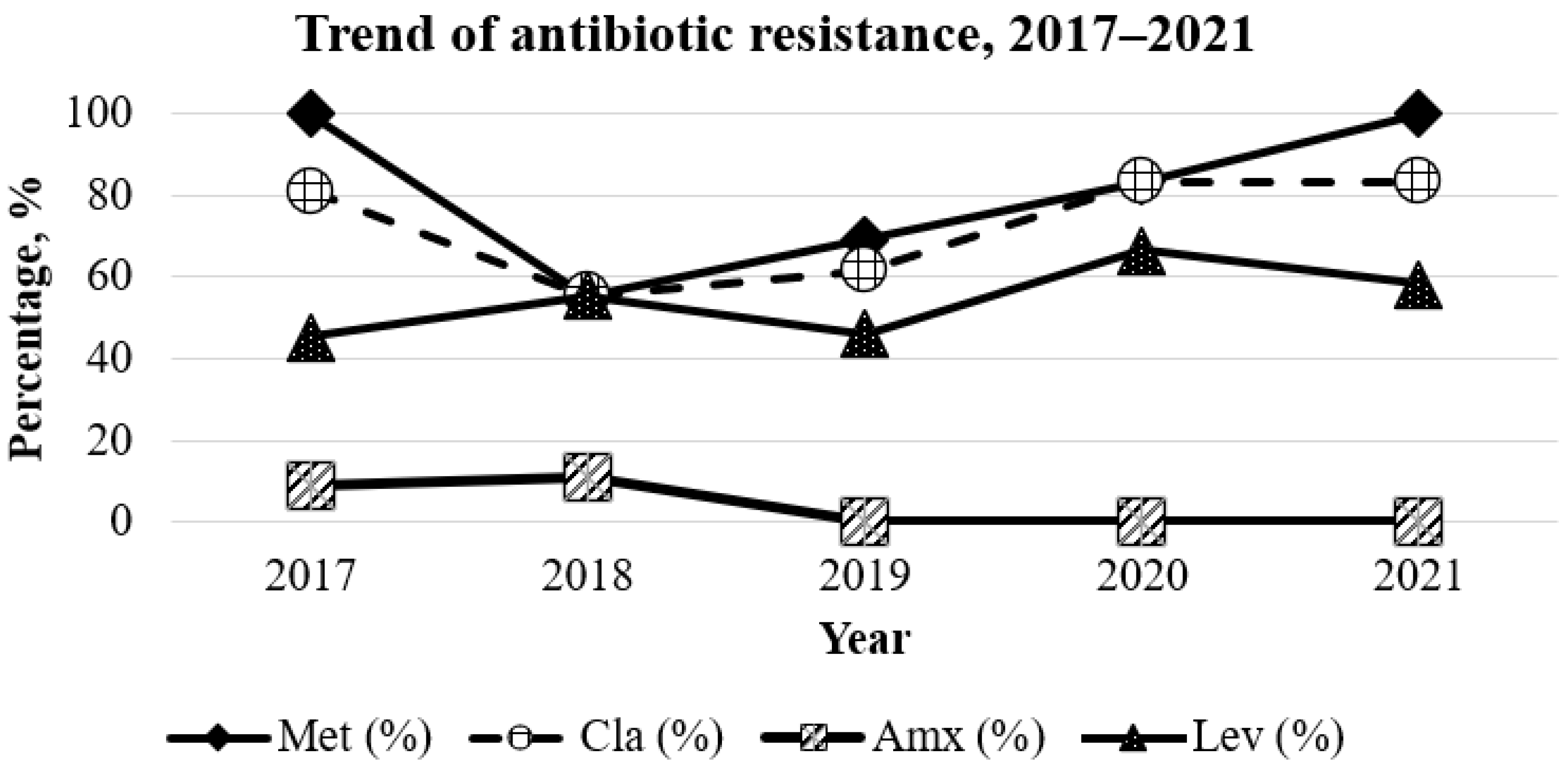
2.2. Antibiotic Resistance Rates
2.3. Minimal Inhibitory Concentration
2.4. Secondary Multidrug Resistance
2.5. Genome Sequencing of Multidrug-Resistant H. pylori
2.6. Virulence Factors Detected in Multidrug-Resistant H. pylori
2.7. Antibiotic-Resistant Genes in Amoxicillin-Resistant H. pylori
2.8. Genotypic and Phenotypic Correlation of the MDR H. pylori Strains
3. Discussion
4. Materials and Methods
4.1. Sample Collection
4.2. Culture of H. pylori
4.3. Antibiotic Susceptibility Testing
4.4. Genome Sequencing of Multidrug-Resistant H. pylori
4.5. Analysis of Antibiotic Resistance Genes and Virulence Factors
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Uemura, N.; Okamoto, S.; Yamamoto, S.; Matsumura, N.; Yamaguchi, S.; Yamakido, M.; Taniyama, K.; Sasaki, N.; Schlemper, R.J. Helicobacter pylori infection and the development of gastric cancer. N. Engl. J. Med. 2021, 345, 784–789. [Google Scholar] [CrossRef]
- Zhang, K.; Yin, J.; Huang, H.; Wang, L.; Guo, L.; Shi, J.; Dai, M. Expenditure and financial burden for stomach cancer diagnosis and treatment in China: A multicenter study. Front. Public Health 2020, 8, 310. [Google Scholar] [CrossRef]
- Chiang, T.H.; Chang, W.J.; Chen, S.L.; Yen, A.M.; Fann, J.C.; Chiu, S.Y.; Chen, Y.R.; Chuang, S.L.; Shieh, C.F.; Liu, C.Y.; et al. Mass eradication of Helicobacter pylori to reduce gastric cancer incidence and mortality: A long-term cohort study on Matsu Islands. Gut 2021, 70, 243–250. [Google Scholar]
- Choi, I.J.; Kim, C.G.; Lee, J.Y.; Kim, Y.I.; Kook, M.C.; Park, B.; Joo, J. Family history of gastric cancer and Helicobacter pylori treatment. N. Engl. J. Med. 2020, 382, 9427–9436. [Google Scholar] [CrossRef]
- Malfertheiner, P.; Megraud, F.; O’Morain, C.A.; Gisbert, J.P.; Kuipers, E.J.; Axon, A.T.; Bazzoli, F.; Gasbarrini, A.; Atherton, J.; Graham, D.Y.; et al. European Helicobacter and Microbiota Study Group and Consensus panel. Management of Helicobacter pylori infection-the Maastricht V/Florence Consensus Report. Gut 2017, 66, 6–30. [Google Scholar] [CrossRef] [Green Version]
- Boyanova, L.; Hadzhiyski, P.; Kandilarov, N.; Markovska, R.; Mitov, I. Multidrug resistance in Helicobacter pylori: Current state and future directions. Expert Rev. Clin. Pharmacol. 2019, 12, 909–915. [Google Scholar] [CrossRef]
- Tacconelli, E.; Carrara, E.; Savoldi, A.; Harbarth, S.; Mendelson, M.; Monnet, D.L.; Pulcini, C.; Kahlmeter, G.; Kluytmans, J.; Carmeli, Y.; et al. WHO Pathogens Priority List Working Group. Discovery, research, and development of new antibiotics: The WHO priority list of antibiotic-resistant bacteria and tuberculosis. Lancet Infect. Dis. 2018, 18, 318–327. [Google Scholar] [CrossRef]
- Goh, K.L.; Parasakthi, N. The racial cohort phenomenon: Seroepidemiology of Helicobacter pylori infection in a multiracial South-East Asian country. Eur. J. Gastroenterol. Hepatol. 2001, 13, 177–183. [Google Scholar] [CrossRef]
- Sukri, A.; Lopes, B.S.; Hanafiah, A. The emergence of multidrug-resistant Helicobacter pylori in Southeast Asia: A systematic review on the trends and intervention strategies using antimicrobial peptides. Antibiotics 2021, 10, 1061. [Google Scholar] [CrossRef]
- Hanafiah, A.; Binmaeil, H.; Raja Ali, R.A.; Rose, I.M.; Lopes, B.S. Molecular characterization and prevalence of antibiotic resistance in Helicobacter pylori isolates in Kuala Lumpur, Malaysia. Infect. Drug Resist. 2019, 12, 3051–3061. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, N.; Zakaria, W.R.; Mohamed, R. Analysis of antibiotic susceptibility patterns of Helicobacter pylori isolates from Malaysia. Helicobacter 2011, 16, 47–51. [Google Scholar] [CrossRef] [PubMed]
- Teh, X.; Khosravi, Y.; Lee, W.C.; Leow, A.H.; Loke, M.F.; Vadivelu, J.; Goh, K.L. Functional and molecular surveillance of Helicobacter pylori antibiotic resistance in Kuala Lumpur. PLoS ONE 2014, 9, e101481. [Google Scholar]
- Tshibangu-Kabamba, E.; Yamaoka, Y. Helicobacter pylori infection and antibiotic resistance—From biology to clinical implications. Nat. Rev. Gastroenterol. Hepatol. 2021, 18, 613–629. [Google Scholar] [CrossRef]
- Globocan 2020. Available online: https://gco.iarc.fr/today/data/factsheets/populations/458-malaysia-fact-sheets.pdf (accessed on 16 April 2022).
- Goh, K.L.; Cheah, P.L.; Md, N.; Quek, K.F.; Parasakthi, N. Ethnicity and H. pylori as risk factors for gastric cancer in Malaysia: A prospective case control study. Am. J. Gastroenterol. 2007, 102, 40–45. [Google Scholar] [CrossRef]
- Sukri, A.; Hanafiah, A.; Kosai, N.R.; Taher, M.M.; Rose, I.M. Distribution of gastric adenocarcinoma subtypes in different ethnicities in Kuala Lumpur, Malaysia. Malays. J. Pathol. 2017, 39, 235–242. [Google Scholar]
- Rasheed, F.; Campbell, B.J.; Alfizah, H.; Varro, A.; Zahra, R.; Yamaoka, Y.; Pritchard, D.M. Analysis of clinical isolates of Helicobacter pylori in Pakistan reveals high degrees of pathogenicity and high frequencies of antibiotic resistance. Helicobacter 2014, 19, 387–399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alfizah, H.; Ramelah, M.; Rizal, A.M.; Anwar, A.S.; Isa, M.R. Association of Malaysian Helicobacter pylori virulence polymorphisms with severity of gastritis and patients’ ethnicity. Helicobacter 2012, 17, 340–349. [Google Scholar] [CrossRef]
- Jiang, F.; Guo, C.G.; Cheung, K.S.; Li, B.; Law, S.; Leung, W.K. Age of eradication and failure rates of clarithromycin-containing triple therapy for Helicobacter pylori: A 15-year population-based study. Helicobacter 2022, 19, 387–399. [Google Scholar] [CrossRef]
- Savoldi, A.; Carrara, E.; Graham, D.Y.; Conti, M.; Tacconelli, E. Prevalence of antibiotic resistance in Helicobacter pylori: A systematic review and meta-analysis in World Health Organization Regions. Gastroenterology 2018, 155, 1372–1382. [Google Scholar] [CrossRef] [Green Version]
- Goderska, K.; Agudo Pena, S.; Alarcon, T. Helicobacter pylori treatment: Antibiotics or probiotics. Appl. Microbiol. Biotechnol. 2018, 102, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Tuan, V.P.; Narith, D.; Tshibangu-Kabamba, E.; Dung, H.D.Q.; Viet, P.T.; Sokomoth, S.; Binh, T.T.; Sokhem, S.; Tri, T.D.; Ngov, S.; et al. A next-generation sequencing-based approach to identify genetic determinants of antibiotic resistance in Cambodian Helicobacter pylori clinical isolates. J. Clin. Med. 2019, 8, 858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tran, T.T.; Nguyen, A.T.; Quach, D.T.; Pham, D.T.; Cao, N.M.; Nguyen, U.T.; Dang, A.N.; Tran, M.A.; Quach, L.H.; Tran, K.T.; et al. Emergence of amoxicillin resistance and identification of novel mutations of the pbp1A gene in Helicobacter pylori in Vietnam. BMC Microbiol. 2022, 22, 45241. [Google Scholar] [CrossRef]
- Han, F.; Liu, S.; Ho, B.; Yan, Z.; Yan, X. Alterations in rdxA and frxA genes and their upstream regions in metronidazole-resistant Helicobacter pylori isolates. Res. Microbiol. 2007, 158, 38–44. [Google Scholar] [CrossRef]
- Chisholm, S.A.; Owen, R.J. Frequency and molecular characteristics of ciprofloxacin- and rifampicin-resistant Helicobacter pylori from gastric infections in the UK. J. Med. Microbiol. 2009, 58, 1322–1328. [Google Scholar] [CrossRef]
- Francesco, V.D.; Zullo, A.; Hassan, C.; Giorgio, F.; Rosania, R.; Ierardi, E. Mechanisms of Helicobacter pylori antibiotic resistance: An updated appraisal. World J. Gastronitest. Pathophysiol. 2011, 2, 35–41. [Google Scholar] [CrossRef]
- Kim, J.M.; Kim, J.S.; Kim, N.; Jung, H.C.; Song, I.S. Distribution of fluoroquinolone MICs in Helicobacter pylori strains from Korean patients. J. Antimicrobiol. Chem. 2005, 56, 965–967. [Google Scholar] [CrossRef] [Green Version]
- Monno, R.; Fumarola, L.; Capolongo, C.; Calia, C.; Pazzani, C.; Ierardi, E.; Miegliotta, G. Susceptibility of Helicobacter pylori to antibiotics including tigecycline. J. Med. Microbiol. Diagn. 2015, S5, 1. [Google Scholar] [CrossRef] [Green Version]
- M100-S21; Performance Standards for Antimicrobial Susceptibility Testing; Twentieth Informational Supplement. MIC Interpretive Standards for Helicobacter pylori. Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2010.
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. fastp: An ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef] [PubMed]
- Wick, R.R.; Judd, L.M.; Gorrie, C.L.; Holt, K.E. Unicycler: Resolving bacterial genome assemblies from short and long sequencing reads. PLoS Comput. Biol. 2017, 13, e1005595. [Google Scholar] [CrossRef] [Green Version]
- Gurevich, A.; Saveliev, V.; Vyahhi, N.; Tesler, G. QUAST: Quality assessment tool for genome assemblies. Bioinformatics 2013, 29, 1072–1075. [Google Scholar] [CrossRef] [Green Version]
- Simão, F.A.; Waterhouse, R.M.; Ioannidis, P.; Kriventseva, E.V.; Zdobnov, E.M. BUSCO: Assessing genome assembly and annotation completeness with single-copy orthologs. Bioinformatics 2015, 31, 3210–3212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seemann, T. Prokka: Rapid prokaryotic genome annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- GenBank. Available online: https://www.ncbi.nlm.nih.gov/genbank/ (accessed on 19 August 2022).
- Alcock, B.P.; Raphenya, A.R.; Lau, T.T.Y.; Tsang, K.K.; Bouchard, M.; Edalatmand, A.; Huynh, W.; Nguyen, A.V.; Cheng, A.A.; Liu, S.; et al. CARD 2020: Antibiotic resistome surveillance with the comprehensive antibiotic resistance database. Nucleic Acids Res. 2020, 48, D517–D525. [Google Scholar] [CrossRef] [PubMed]
- Clustal Omega. Available online: https://www.ebi.ac.uk/Tools/msa/clustalo/ (accessed on 19 August 2022).


{kind=link}
| Overall (n = 218) | p-Value | |
|---|---|---|
| Age: | ||
| Range | 18–77 years | |
| Mean ± SD | 44.5 ± 11.8 years | |
| Age group: | <0.001 * | |
| <60 years | 190 (87.2) | |
| >60 years | 28 (12.8) | |
| Gender, n (%): | ||
| Male | 106 (48.6) | |
| Female | 112 (51.4) | |
| Ethnicity: | <0.001 * | |
| Malay | 33 (15.1) | |
| Chinese | 94 (43.1) | |
| Indian | 48 (22.0) | |
| Other | 43 (20.2) | |
| Positive culture, n (%) | 51 (23.4) |
| Multidrug Resistance | 2017, (n = 11) -%- | 2018, (n = 9) -%- | 2019, (n = 13) -%- | 2020, (n = 6) -%- | 2021, (n = 12) -%- | Total, (n = 41) % |
|---|---|---|---|---|---|---|
| MTZ, CLA | 45.5 | 11.1 | 7.7 | 16.7 | 58.33 | 36.6 |
| MTZ, LEV | 9.1 | 44.4 | 0 | 0 | 8.3 | 14.6 |
| MTX, AMO | 9.1 | 0 | 0 | 0 | 0 | 2.4 |
| MTZ, CLA, LEV | 36.4 | 11.1 | 46.2 | 66.7 | 25 | 43.9 |
| MTZ, CLA, AMO | 0 | 11.1 | 0 | 0 | 0 | 11.1 |
| Variant | Detection of PBP1A Variants | |
|---|---|---|
| HP18045 | HP21041 | |
| V16I | Yes | Yes |
| F125L | Yes | Yes |
| I148L | Yes | Yes |
| G242S | Yes | Yes |
| N322D | Yes | Yes |
| V346L | Yes | No |
| V374L | Yes | No |
| D508E | Yes | Yes |
| D535N | Yes | Yes |
| G595_V596InsG | Yes | No |
| Y604H | Yes | No |
| N608S | Yes | No |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sukri, A.; Hanafiah, A.; Yusoff, H.; Shamsul Nizam, N.A.; Nameyrra, Z.; Wong, Z.; Raja Ali, R.A. Multidrug-Resistant Helicobacter pylori Strains: A Five-Year Surveillance Study and Its Genome Characteristics. Antibiotics 2022, 11, 1391. https://doi.org/10.3390/antibiotics11101391
Sukri A, Hanafiah A, Yusoff H, Shamsul Nizam NA, Nameyrra Z, Wong Z, Raja Ali RA. Multidrug-Resistant Helicobacter pylori Strains: A Five-Year Surveillance Study and Its Genome Characteristics. Antibiotics. 2022; 11(10):1391. https://doi.org/10.3390/antibiotics11101391
Chicago/Turabian StyleSukri, Asif, Alfizah Hanafiah, Hamidah Yusoff, Nur Atiqah Shamsul Nizam, Zarith Nameyrra, Zhiqin Wong, and Raja Affendi Raja Ali. 2022. "Multidrug-Resistant Helicobacter pylori Strains: A Five-Year Surveillance Study and Its Genome Characteristics" Antibiotics 11, no. 10: 1391. https://doi.org/10.3390/antibiotics11101391