
Lipid Oxidation and Colour Stability of Lamb and Yearling Meat (Muscle longissimus lumborum) from Sheep Supplemented with Camelina-Based Diets after Short-, Medium-, and Long-Term Storage

 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Diets, Animals, and Experimental Design
2.2. Slaughter of Animals and Muscle Sample Collection
2.3. Determination of Meat (Muscle LL) Tenderness, Vitamin E Concentration, and Fatty Acid Composition
2.4. Assessment of Colour and Lipid Oxidative Stability of Meat
2.5. Statistical Analyses
3. Results
3.1. Meat Tenderness, Vitamin E Concentration, and Fatty Acid Composition of Meat
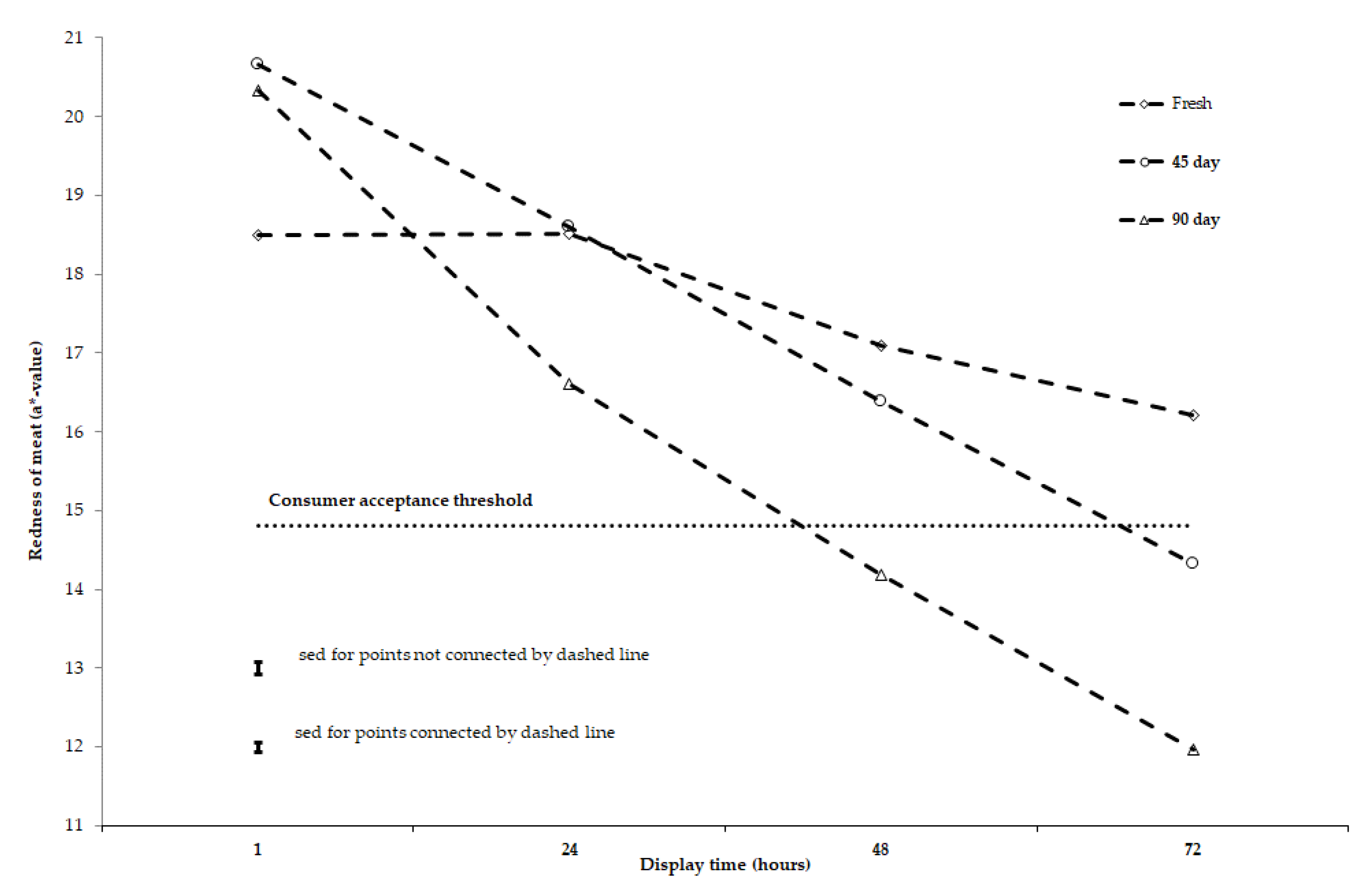
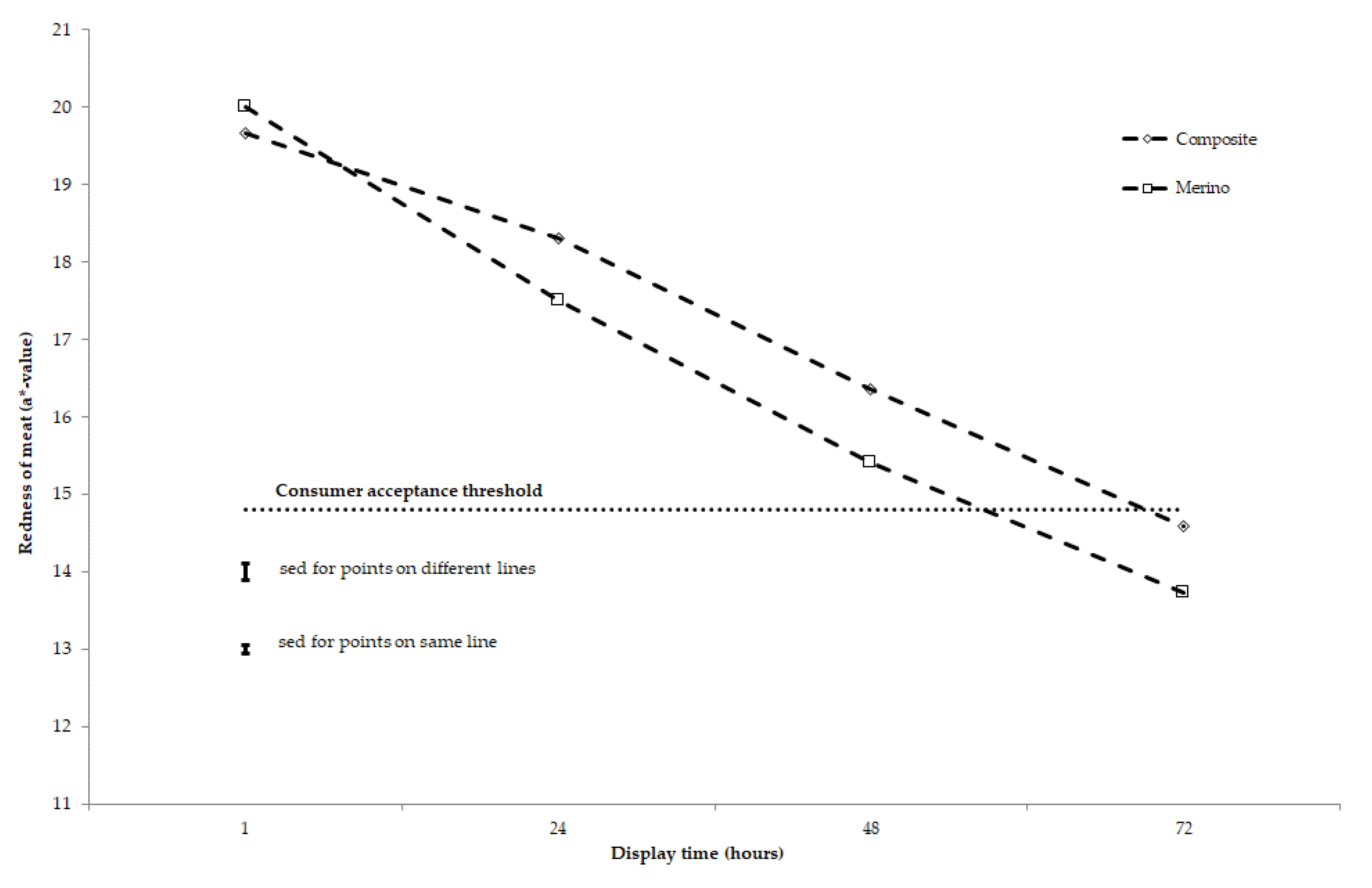
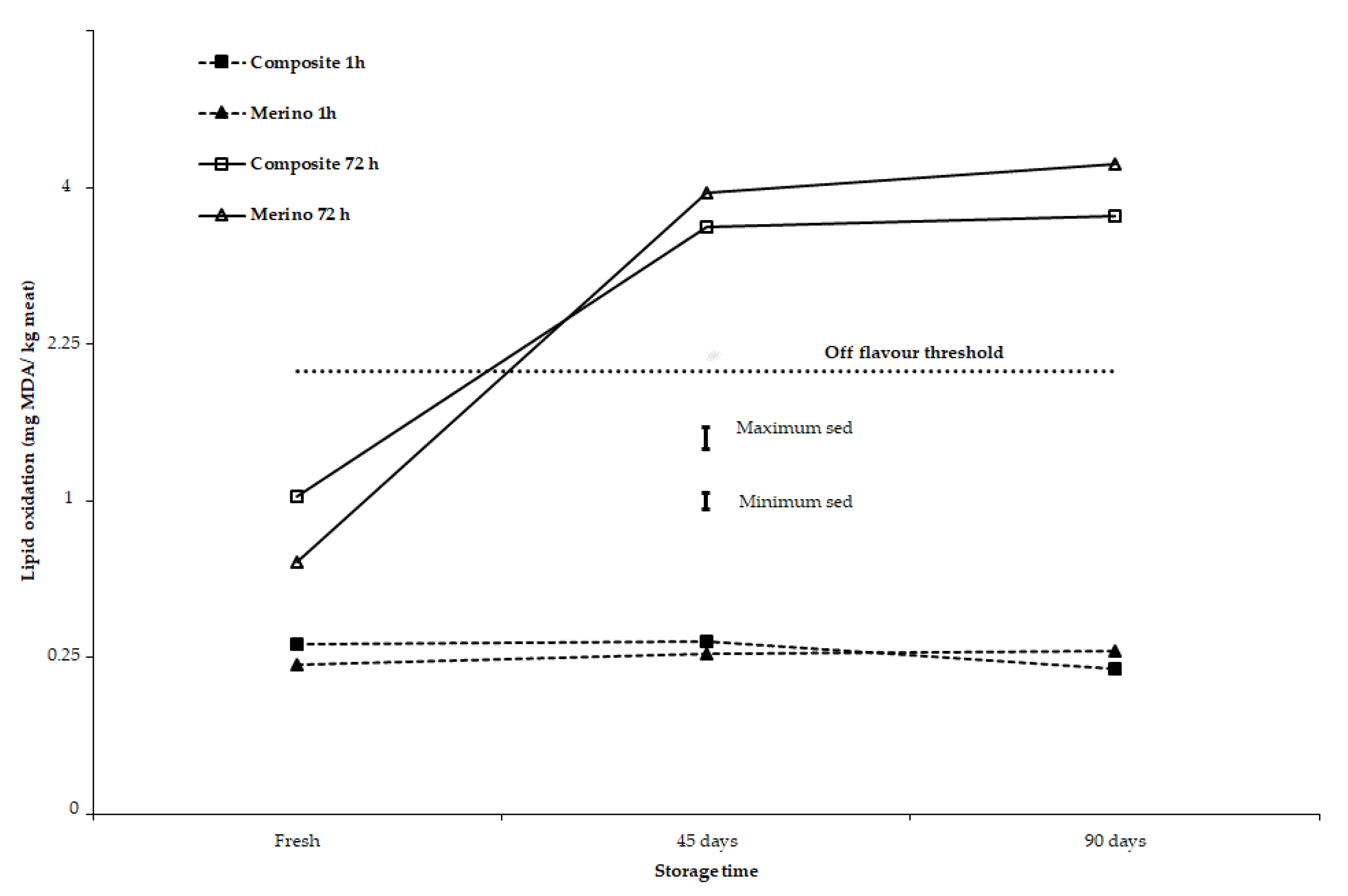
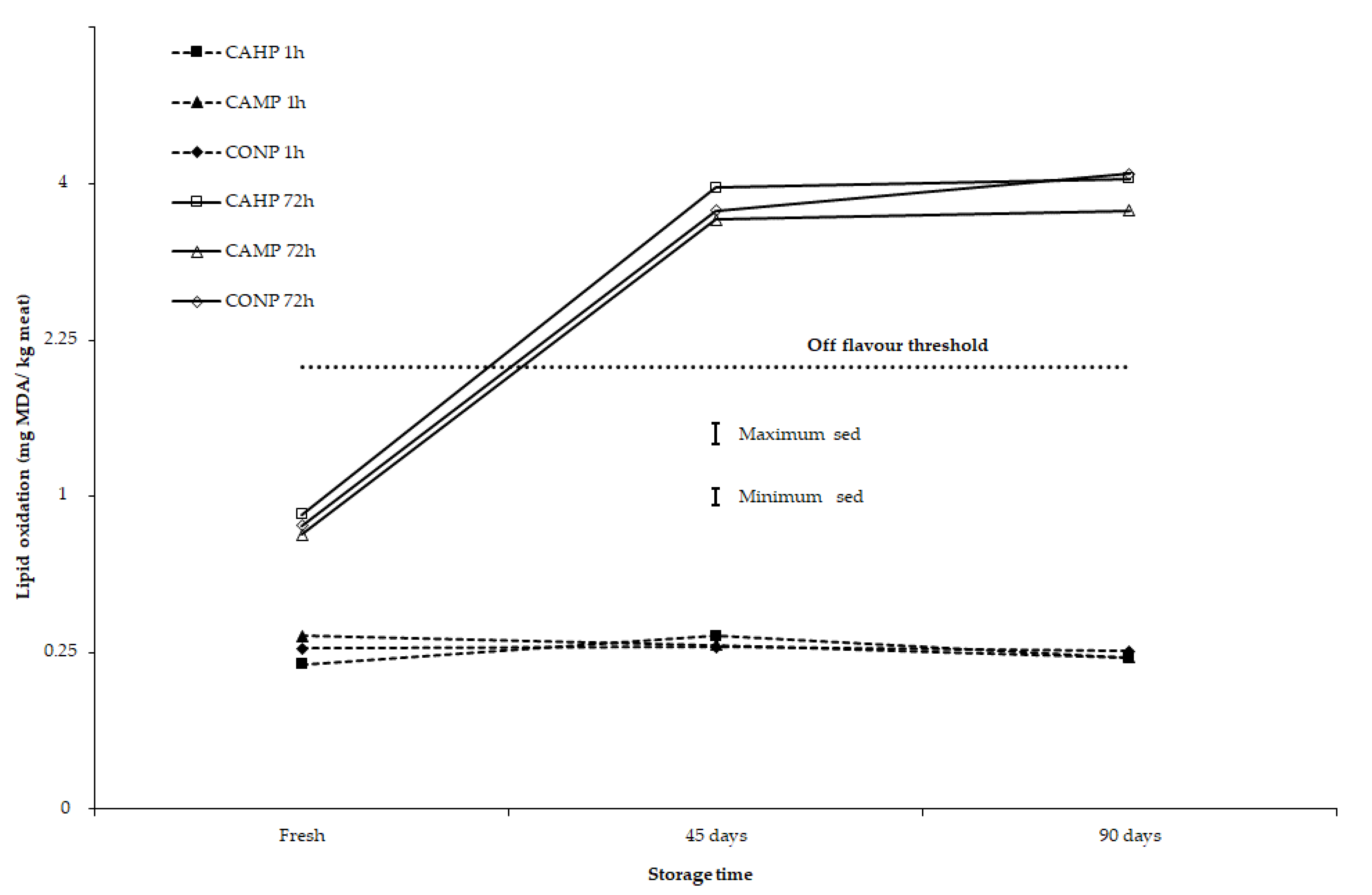
3.2. Stability of Meat Colour and Lipid Oxidation after 2-, 45-, and 90-Day Storage under Chilled to Semi-Frozen Conditions
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Meat and Livestock Australia Global Snapshot-Sheepmeat. Available online: https://www.mla.com.au/globalassets/mla-corporate/prices--markets/documents/os-markets/red-meat-market-snapshots/2020/global-sheepmeat-snapshot-jan2020.pdf (accessed on 12 October 2020).
- Coombs, C.E.O.; Holman, B.W.B.; Friend, M.A.; Hopkins, D.L. Long-term red meat preservation using chilled and frozen storage combinations: A review. Meat Sci. 2017, 125, 84–94. [Google Scholar] [CrossRef] [PubMed]
- Muela, E.; Sañudo, C.; Campo, M.M.; Medel, I.; Beltrán, J.A. Effect of freezing method and frozen storage duration on lamb sensory quality. Meat Sci. 2012, 90, 209–215. [Google Scholar] [CrossRef] [PubMed]
- Williams, P. Nutritional composition of red meat. Nutr. Diet. 2007, 64, S113–S119. [Google Scholar] [CrossRef] [Green Version]
- Ponnampalam, E.N.; Plozza, T.; Kerr, M.G.; Linden, N.; Mitchell, M.; Bekhit, A.E.A.; Jacobs, J.L.; Hopkins, D.L. Interaction of diet and long ageing period on lipid oxidation and colour stability of lamb meat. Meat Sci. 2017, 129, 43–49. [Google Scholar] [CrossRef] [PubMed]
- Scollan, N.; Hocquette, J.F.; Nuernberg, K.; Dannenberger, D.; Richardson, I.; Moloney, A. Innovations in beef production systems that enhance the nutritional and health value of beef lipids and their relationship with meat quality. Meat Sci. 2006, 74, 17–33. [Google Scholar] [CrossRef] [PubMed]
- Meat and Livestock Australia, Shelf Life of Australian Red Meat, 2nd ed.; Meat and Livestock Australia: Sydney, Australia, 2016.
- Coombs, C.E.O.; Holman, B.W.B.; Ponnampalam, E.N.; Morris, S.; Friend, M.A.; Hopkins, D.L. Effects of chilled and frozen storage conditions on the lamb M. longissimus lumborum fatty acid and lipid oxidation parameters. Meat Sci. 2018, 136, 116–122. [Google Scholar] [CrossRef] [PubMed]
- Moriel, P.; Nayigihugu, V.; Cappellozza, B.I.; Gonçalves, E.P.; Krall, J.M.; Foulke, T.; Cammack, K.M.; Hess, B.W. Camelina meal and crude glycerin as feed supplements for developing replacement beef heifers. J. Anim. Sci. 2011, 89, 4314–4324. [Google Scholar] [CrossRef] [Green Version]
- Steppa, R.; Cieślak, A.; Szumacher-Strabel, M.; Bielińska-Nowak, S.; Bryszak, M.; Stanisz, M.; Szkudelska, K. Blood serum metabolic profile and fatty acid composition in sheep fed concentrates with Camelina sativa cake and distillers dried grains with solubles. Small Rumin. Res. 2017, 156, 20–26. [Google Scholar] [CrossRef]
- Cieslak, A.; Stanisz, M.; Wojtowski, J.; Pers-Kamczyc, E.; Szczechowiak, J.; El-Sherbiny, M.; Szumacher-Strabel, M. Camelina sativa affects the fatty acid contents in M. longissimus muscle of lambs. Eur. J. Lipid Sci. Technol. 2013, 115, 1258–1265. [Google Scholar] [CrossRef]
- Gómez-Cortés, P.; Galisteo, O.O.; Avilés Ramirez, C.; Peña Blanco, F.; de la Fuente, M.A.; Núñez Sánchez, N.; Martínez Marín, A.L. Intramuscular fatty acid profile of feedlot lambs fed concentrates with alternative ingredients. Anim. Prod. Sci. 2019, 59, 914–920. [Google Scholar] [CrossRef]
- Noci, F.; Monahan, F.; Moloney, A. The fatty acid profile of muscle and adipose tissue of lambs fed camelina or linseed as oil or seeds. Animal 2011, 5, 134–147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roberts, C.A.; Stuth, J.; Flinn, P. Analysis of forages and feedstuffs. In Near-Infrared Spectroscopy in Agriculture; Roberts, C.A., Workman, J., Jr., Reeves, J.B., III, Eds.; American Society of Agronomy: Madison, WI, USA, 2004; pp. 231–268. [Google Scholar]
- National Health and Medical Research Council. Australian Code for the Care and Use of Animals for Scientific Purposes, 8th ed.; National Health and Medical Research Council: Canberra, Australian, 2013.
- Hopkins, D.L.; Toohey, E.S.; Warner, R.D.; Kerr, M.J.; van de Ven, R. Measuring the shear force of lamb meat cooked from frozen samples: Comparison of two laboratories. Anim. Prod. Sci. 2010, 50, 382–385. [Google Scholar] [CrossRef]
- O’Fallon, J.V.; Busboom, J.R.; Nelson, M.L.; Gaskins, C.T. A direct method for fatty acid methyl ester synthesis: Application to wet meat tissues, oils, and feedstuffs. J. Anim. Sci. 2007, 85, 1511–1521. [Google Scholar] [CrossRef] [Green Version]
- Mestre Prates, J.A.; Gonçalves Quaresma, M.A.; Branquinho Bessa, R.J.; Andrade Fontes, C.M.G.; Mateus Alfaia, C.M.P.; Ingvartsen, K.L. Simultaneous HPLC quantification of total cholesterol, tocopherols and β-carotene in Barrosã-PDO veal. Food Chem. 2006, 94, 469–477. [Google Scholar] [CrossRef]
- Ponnampalam, E.N.; Burnett, V.F.; Norng, S.; Hopkins, D.L.; Plozza, T.; Jacobs, J.L. Muscle antioxidant (vitamin E) and major fatty acid groups, lipid oxidation and retail colour of meat from lambs fed a roughage based diet with flaxseed or algae. Meat Sci. 2016, 111, 154–160. [Google Scholar] [CrossRef] [PubMed]
- Witte, V.; Krause, G.; Bailey, M. A new extraction method for determining 2-thiobarbituric acid values of pork and beef during storage. J. Food Sci. 2006, 35, 582–585. [Google Scholar] [CrossRef]
- VSN International. The Guide to the Genstat Command Language (Release 19), Part 2 Statistics; VSN International: Hemel Hempstead, UK, 2017. [Google Scholar]
- Khliji, S.; van de Ven, R.; Lamb, T.A.; Lanza, M.; Hopkins, D.L. Relationship between consumer ranking of lamb colour and objective measures of colour. Meat Sci. 2010, 85, 224–229. [Google Scholar] [CrossRef]
- Holman, B.W.B.; Coombs, C.E.O.; Morris, S.; Kerr, M.J.; Hopkins, D.L. Effect of long term chilled (up to 5weeks) then frozen (up to 12months) storage at two different sub-zero holding temperatures on beef: 1. Meat quality and microbial loads. Meat Sci. 2017, 133, 133–142. [Google Scholar] [CrossRef]
- Moloney, A.P.; Kennedy, C.; Noci, F.; Monahan, F.J.; Kerry, J.P. Lipid and colour stability of M. longissimus muscle from lambs fed camelina or linseed as oil or seeds. Meat Sci. 2012, 92, 1–7. [Google Scholar] [CrossRef]
- Hopkins, D.L.; Hegarty, R.S.; Walker, P.J.; Pethick, D.W. Relationship between animal age, intramuscular fat, cooking loss, pH, shear force and eating quality of aged meat from sheep. Aust. J. Exp. Agric. 2006, 46, 879–884. [Google Scholar] [CrossRef]
- Pannier, L.; Gardner, G.E.; Pearce, K.L.; McDonagh, M.; Ball, A.J.; Jacob, R.H.; Pethick, D.W. Associations of sire estimated breeding values and objective meat quality measurements with sensory scores in Australian lamb. Meat Sci. 2014, 96, 1076–1087. [Google Scholar] [CrossRef] [PubMed]
- Ponnampalam, E.N.; Hosking, B.J.; Egan, A.R. Rate of carcass components gain, carcass characteristics, and muscle longissimus tenderness in lambs fed dietary protein sources with a low quality roughage diet. Meat Sci. 2003, 63, 143–149. [Google Scholar] [CrossRef]
- Koohmaraie, M.; Seideman, S.C.; Schollmeyer, J.E.; Dutson, T.R.; Babiker, A.S. Factors associated with the tenderness of three bovine muscles. J. Food Sci. 1988, 53, 407–410. [Google Scholar] [CrossRef]
- Whipple, G.; Koohmaraie, M.; Dikeman, M.; Crouse, J.; Hunt, M.; Klemm, R. Evaluation of attributes that affect longissimus muscle tenderness in Bos taurus and Bos indicus cattle. J. Anim. Sci. 1990, 68, 2716–2728. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Avilés Ramirez, C.; Peña Blanco, F.; Horcada, A.; Nuñez Sánchez, N.; Requena Domenech, F.; Guzman Medina, P.; Martínez Marín, A.L. Effects of concentrates rich in by-products on growth performance, carcass characteristics and meat quality traits of light lambs. Anim. Prod. Sci. 2019, 59, 593–599. [Google Scholar] [CrossRef]
- Ponnampalam, E.N.; Behrendt, R.; Kerr, M.G.; Raeside, M.C.; McDonagh, M.B. The influence of the level of ewe gestation nutrition and lamb finishing diet on long-chain polyunsaturated fat concentration, antioxidant and mineral status, and colour stability of meat. Anim. Prod. Sci. 2018, 58, 1481–1487. [Google Scholar] [CrossRef]
- Bellés, M.; Leal, L.N.; Díaz, V.; Alonso, V.; Roncalés, P.; Beltrán, J.A. Effect of dietary vitamin E on physicochemical and fatty acid stability of fresh and thawed lamb. Food Chem. 2018, 239, 1–8. [Google Scholar] [CrossRef]
- González-Calvo, L.; Ripoll, G.; Molino, F.; Calvoa, J.H.; Joya, M. The relationship between muscle α-tocopherol concentration and meat oxidation in light lambs fed vitamin E supplements prior to slaughter. J. Sci. Food Agric. 2015, 95, 103–110. [Google Scholar] [CrossRef]
- Lee, J.H.; Waller, J.C.; Yilmaz, Y.; Melton, S.L. Effect of feeding rumen-protected dietary protein–oil supplements on fatty acid composition and alpha-tocopherol content of blood serum and muscle lipids of lambs. Small Rumin. Res. 2007, 72, 101–110. [Google Scholar] [CrossRef]
- Liu, K.; Ge, S.; Luo, H.; Yue, D.; Yan, L. Effects of dietary vitamin E on muscle vitamin E and fatty acid content in Aohan fine-wool sheep. J. Anim. Sci. Biotechnol. 2013, 4, 21. [Google Scholar] [CrossRef] [Green Version]
- Ponnampalam, E.N.; Burnett, V.F.; Norng, S.; Warner, R.D.; Jacobs, J.L. Vitamin E and fatty acid content of lamb meat from perennial pasture or annual pasture systems with supplements. Anim. Prod. Sci. 2012, 52, 255–262. [Google Scholar] [CrossRef]
- Jose, C.G.; Jacob, R.H.; Pethick, D.W.; Gardner, G.E. Short term supplementation rates to optimise vitamin E concentration for retail colour stability of Australian lamb meat. Meat Sci. 2016, 111, 101–109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ponnampalam, E.N.; Norng, S.; Burnett, V.F.; Dunshea, F.R.; Jacobs, J.L.; Hopkins, D.L. The synergism of biochemical components controlling lipid oxidation in lamb muscle. Lipids 2014, 49, 757–766. [Google Scholar] [CrossRef] [PubMed]
- Ponnampalam, E.N.; Hopkins, D.L.; Butler, K.L.; Dunshea, F.R.; Warner, R.D. Genotype and age effects on sheep meat production 2. Carcass quality traits. Aust. J Exp. Agric. 2007, 47, 1147–1154. [Google Scholar] [CrossRef]
- Fogarty, N.M.; Hopkins, D.L.; Ven, R.V.D. Lamb production from diverse genotypes. 2. Carcass characteristics. Anim. Sci. 2000, 70, 147–156. [Google Scholar] [CrossRef]
- Arnold, R.N.; Arp, S.C.; Scheller, K.K.; Williams, S.N.; Schaefer, D.M. Tissue equilibration and subcellular distribution of vitamin E relative to myoglobin and lipid oxidation in displayed beef. J. Anim. Sci. 1993, 71, 105–118. [Google Scholar] [CrossRef]
- Liu, Q.; Lanari, M.; Schaefer, D.M. A review of dietary Vitamin E supplementation for improvement of beef quality. J. Anim. Sci. 1995, 73, 3131–3140. [Google Scholar] [CrossRef] [PubMed]
- Ponnampalam, E.N.; Butler, K.L.; Burnett, V.F.; McDonagh, M.B.; Jacobs, J.L.; Hopkins, D.L. Aged vacuum packaged lamb cuts are less brown than fresh muscle cuts under simulated retail display. Food Nutr. Sci. 2013, 4, 147–153. [Google Scholar] [CrossRef] [Green Version]
- Ponnampalam, E.N.; Trout, G.R.; Sinclair, A.J.; Egan, A.R.; Leury, B.J. Comparison of the Color Stability and Lipid Oxidative Stability of Fresh and Vacuum Packaged Lamb Muscle Containing Elevated Omega-3 and Omega- 6 Fatty Acid Levels from Dietary Manipulation. Meat Sci. 2001, 58, 151–161. [Google Scholar] [CrossRef]
- Ponnampalam, E.N.; Butler, K.L.; McDonagh, M.B.; Jacobs, J.L.; Hopkins, D.L. Relationship between muscle antioxidant status, forms of iron, polyunsaturated fatty acids and functionality (retail colour) of meat in lambs. Meat Sci. 2012, 90, 297–303. [Google Scholar] [CrossRef]
- Abuelfatah, K.; Zuki, A.B.Z.; Goh, Y.M.; Sazili, A.Q. Effects of enriching goat meat with n−3 polyunsaturated fatty acids on meat quality and stability. Small Rumin. Res. 2016, 136, 36–42. [Google Scholar] [CrossRef]
- Campo, M.M.; Nute, G.R.; Hughes, S.I.; Enser, M.; Wood, J.D.; Richardson, R.I. Flavour perception of oxidation in beef. Meat Sci. 2006, 72, 303–311. [Google Scholar] [CrossRef] [PubMed]
- Nuernberg, K.; Nuernbergb, G.; Endera, K.; Dannenbergera, D.; Schabbelc, W.; Grumbachd, S.; Zuppd, W.; Steinhartc, H. Effect of grass vs. concentrate feeding on the fatty acid profile of different fat depots in lambs. Eur. J. Lipid Sci. Technol. 2005, 107, 737–745. [Google Scholar] [CrossRef]
- Simopoulos, A.P. An increase in the omega-6/omega-3 fatty acid ratio increases the risk for obesity. Nutrients 2016, 8, 128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ponnampalam, E.N.; Kerr, M.G.; Butler, K.L.; Cottrell, J.J.; Dunshea, F.R.; Jacobs, J.L. Filling the out of season gaps for lamb and hogget production: Diet and genetic influence on carcass yield, carcass composition and retail value of meat. Meat Sci. 2019, 148, 156–163. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Dietary Ingredients Used (%) | Camelina Hay (CAHP) 1 | Camelina Meal (CAMP) 2 | Control Pellet (CONP) 3 |
|---|---|---|---|
| Camelina meal | 0 | 8 | 0 |
| Lupins | 30 | 22 | 30 |
| Barley grain | 10 | 20 | 20 |
| Oat grain | 15 | 5 | 5 |
| Oaten hay | 0 | 45 | 45 |
| Camelina–Oaten–Barley hay (33:33:33 w/w/w) | 45 | 0 | 0 |
| Nutritive characteristics of diet | |||
| Dry matter, % | 88.85 | 88.90 | 89.35 |
| Crude protein, % DM | 15.20 | 14.93 | 14.80 |
| Metabolisable energy, MJ/ kg DM | 10.83 | 11.23 | 10.80 |
| Crude fat, % DM | 2.91 | 3.65 | 2.11 |
| Acid detergent fibre, % DM | 19.93 | 17.78 | 19.03 |
| Neutral detergent fibre, % DM | 34.03 | 31.18 | 34.23 |
| Lignin, % DM | 4.20 | 4.13 | 4.30 |
| Phosphorous, % DM | 0.42 | 0.52 | 0.47 |
| Potassium, % DM | 1.42 | 1.52 | 1.47 |
| Sulphur, % DM | 0.21 | 0.26 | 0.25 |
| Vitamin E concentration, mg/kg DM | 4.1 | 4.2 | 4.9 |
| Linolenic acid (n − 3) concentration, mg/100g DM | 367 | 846 | 228 |
| Linoleic acid (n − 6) concentration, mg/100 g DM | 1433 | 1391 | 1357 |
| Effect of Diet | Effect of Animal Type | p-Value | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| CAHP 1 | CAMP 2 | CONP 3 | Sed 4 | Composite | Merino | Sed 4 | Diet | Breed | Interaction of Diet × Breed | |
| Vitamin E (mg/kg meat) | 0.81 | 0.86 | 0.99 | 0.029–0.031 | 0.76 | 1.04 | 0.024 | 4.4 × 10−5 | 1.1 × 10−8 | 0.25 |
| Meat tenderness (N) | 34.9 | 36.3 | 33.1 | 2.55–2.72 | 33.3 | 35.9 | 2.11 | 0.47 | 0.24 | 0.73 |
| Fatty acid content (log10 transformed; back-transformed means in parentheses are mg/100 g meat) | ||||||||||
| C10:0 | 0.68 (4.7) | 0.68 (4.8) | 0.69 (4.9) | 0.025–0.026 | 0.56 (3.6) | 0.81 (6.5) | 0.020 | 0.77 | 4.6 × 10−9 | 0.70 |
| C12:0 | 0.53 (3.4) | 0.56 (3.6) | 0.57 (3.7) | 0.031–0.034 | 0.58 (3.8) | 0.52 (3.3) | 0.026 | 0.47 | 0.036 | 0.74 |
| C14:0 | 1.93 (86) | 1.97 (92) | 1.94 (87) | 0.023–0.024 | 1.88 (76) | 2.01 (103) | 0.023 | 0.41 | 4.4 × 10−6 | 0.42 |
| C14:1 | 0.41 (2.6) | 0.41 (2.6) | 0.43 (2.7) | 0.039–0.042 | 0.32 (2.1) | 0.52 (3.3) | 0.033 | 0.80 | 2.9 × 10−5 | 0.76 |
| C15:0 | 0.98 (9.5) | 0.99 (9.8) | 0.99 (9.8) | 0.026–0.028 | 0.88 (7.6) | 1.09 (12.4) | 0.022 | 0.90 | 1.4 × 10−7 | 0.75 |
| C15:1 | 0.21 (1.6) | 0.12 (1.3) | 0.20 (1.6) | 0.039–0.042 | 0.26 (1.8) | 0.09 (1.2) | 0.032 | 0.091 | 0.00012 | 0.25 |
| C16:0 | 2.98 (960) | 3.01 (1020) | 2.99 (970) | 0.014–0.015 | 2.88 (760) | 3.10 (1270) | 0.012 | 0.23 | 2.5 × 10−11 | 0.13 |
| C16:1 | 1.79 (61) | 1.79 (61) | 1.80 (63) | 0.026–0.027 | 1.65 (44) | 1.94 (86) | 0.021 | 0.83 | 1.8 × 10−9 | 0.70 |
| C17:0 | 1.50 (31) | 1.50 (32) | 1.50 (32) | 0.017–0.018 | 1.37 (23) | 1.63 (42) | 0.014 | 0.90 | 2.6 × 10−11 | 0.65 |
| C18:0 | 2.79 (617) | 2.80 (632) | 2.79 (613) | 0.015–0.016 | 2.70 (503) | 2.88 (764) | 0.013 | 0.69 | 9.3 × 10−10 | 0.34 |
| C18:2n−6 cis | 2.14 (136) | 2.33 (216) | 2.00 (99) | 0.040–0.042 | 1.94 (86) | 2.34 (220) | 0.033 | 2.9 × 10−5 | 5.7 × 10−9 | 0.23 |
| C18:1n-9 cis | 3.20 (1600) | 3.20 (1600) | 3.21 (1610) | 0.011–0.012 | 3.09 (1230) | 3.32 (2090) | 0.009 | 0.90 | 6.0 × 10−13 | 0.019 |
| C20:0 | 0.56 (3.6) | 0.67 (4.7) | 0.53 (3.4) | 0.018–0.019 | 0.52 (3.3) | 0.64 (4.4) | 0.015 | 4.1 × 10−6 | 1.4 × 10−6 | 0.050 |
| C18:3n−6 | −0.24 (0.6) | −0.57 (0.3) | −0.13 (0.7) | 0.051–0.055 | −0.38 (0.4) | −0.20 (0.6) | 0.042 | 1.8 × 10−6 | 0.00087 | 0.12 |
| C18:3n−3 (ALA 5) | 1.43 (27) | 1.74 (55) | 1.40 (25) | 0.020–0.022 | 1.38 (24) | 1.64 (44) | 0.017 | 2.2 × 10−10 | 4.6 × 10−10 | 0.37 |
| C20:4n−6 (AA 6) | 1.51 (32) | 1.45 (28) | 1.54 (35) | 0.009–0.010 | 1.54 (35) | 1.47 (29) | 0.008 | 2.9 × 10−7 | 1.5 × 10−7 | 0.46 |
| C20:5n−3 (EPA 7) | 1.12 (13.1) | 1.56 (14.3) | 1.14 (13.9) | 0.010–0.011 | 1.14 (13.7) | 1.14 (13.8) | 0.008 | 0.0075 | 0.87 | 0.17 |
| C22:5n−3 (DPA 8) | 1.24 (17.3) | 1.26 (18.1) | 1.27 (18.6) | 0.009 | 1.25 (17.9) | 1.26 (18.3) | 0.007 | 0.011 | 0.21 | 0.43 |
| C22:6n−3 (DHA 9) | 0.65 (4.5) | 0.66 (4.5) | 0.71 (5.2) | 0.016–0.017 | 0.62 (4.1) | 0.74 (5.5) | 0.013 | 0.0018 | 2.8 × 10−7 | 0.062 |
| EPA + DHA | 1.25 (18) | 1.28 (19) | 1.28 (19) | 0.010–0.011 | 1.25 (18) | 1.28 (19) | 0.008 | 0.0072 | 0.0016 | 0.094 |
| EPA + DPA + DHA | 1.54 (35) | 1.57 (37) | 1.58 (38) | 0.009 | 1.55 (36) | 1.57 (38) | 0.007 | 0.0052 | 0.011 | 0.17 |
| Total n-6 (Omega-6) | 2.22 (166) | 2.21 (161) | 2.23 (171) | 0.009–0.010 | 2.19 (156) | 2.25 (177) | 0.008 | 0.038 | 4.9 × 10−6 | 0.033 |
| Total n−3 (Omega-3) | 1.80 (62) | 1.97 (94) | 1.81 (64) | 0.013–0.014 | 1.79 (62) | 1.92 (83) | 0.011 | 4.1 × 10−9 | 1.5 × 10−8 | 0.24 |
| n−6/n−3 | 0.43 (2.7) | 0.24 (1.7) | 0.43 (2.7) | 0.011 | 0.41 (2.5) | 0.34 (2.2) | 0.009 | 5.5 × 10−11 | 2.3 × 10−6 | 0.88 |
| ∑ SUFA 10 | 3.23 (1720) | 3.25 (1800) | 3.24 (1720) | 0.013–0.14 | 3.14 (1380) | 3.34 (2200) | 0.011 | 0.32 | 3.3 × 10−11 | 0.16 |
| ∑ MUFA 11 | 3.26 (1800) | 3.28 (1880) | 3.25 (1780) | 0.012–0.013 | 3.14 (1370) | 3.38 (2420) | 0.010 | 0.15 | 5.2 × 10−13 | 0.028 |
| ∑ PUFA 12 | 2.36 (228) | 2.41 (255) | 2.37 (235) | 0.010 | 2.34 (219) | 2.42 (261) | 0.008 | 0.00089 | 1.3 × 10−7 | 0.023 |
| Total fat | 3.57 (3750) | 3.60 (3940) | 3.57 (3750) | 0.012–0.013 | 3.47 (2970) | 3.69 (4880) | 0.010 | 0.18 | 3.2 × 10−12 | 0.077 |
| Terms | Degrees of freedom | Mean Square | F-Value | p-Value |
|---|---|---|---|---|
| Between pens | ||||
| Animal type (AT) | 1 | 19.054 | 9.36 | 0.0085 |
| Diet | 2 | 0.039 | 0.02 | 0.98 |
| Diet. AT | 2 | 0.416 | 0.20 | 0.82 |
| Error | 14 | 2.034 | ||
| Between storage lengths within pens combinations | ||||
| Storage length (ST) | 2 | 82.66 | 145.57 | 1.6 × 10−16 |
| AT. ST | 2 | 0.159 | 0.28 | 0.76 |
| Diet. ST | 4 | 0.781 | 1.37 | 0.27 |
| Diet. AT. ST | 4 | 0.914 | 1.61 | 0.20 |
| Error | 28 | |||
| Between display times within storage lengths within pens | ||||
| Display time (DT) | 1.86 | 361.89 | 2616.4 | 2.9 × 10−71 |
| ST. DT | 3.72 | 35.324 | 255.39 | 4.2 × 10−43 |
| AT. DT | 1.86 | 5.446 | 39.37 | 4.6 × 10−12 |
| AT. ST. DT | 3.72 | 0.103 | 0.75 | 0.56 |
| Diet. DT | 3.72 | 0.085 | 0.61 | 0.64 |
| Diet. ST. DT | 7.44 | 0.055 | 0.40 | 0.91 |
| Diet. AT. DT | 3.72 | 0.600 | 4.34 | 0.0039 |
| Diet. AT. ST. DT | 7.44 | 0.060 | 0.44 | 0.89 |
| Error | 78.17 | |||
| Terms | Degrees of Freedom | Mean Square | F-Value | p-Value |
|---|---|---|---|---|
| Between pens | ||||
| Animal type (AT) | 1 | 0.002 | 0.00 | 0.96 |
| Diet | 2 | 0.011 | 0.17 | 0.85 |
| Diet.AT | 2 | 0.009 | 0.13 | 0.88 |
| Error | 14 | 0.067 | ||
| Between storage lengths within pens combinations | ||||
| Storage times (ST) | 2 | 3.692 | 556.38 | 2.9 × 10−23 |
| AT.ST | 2 | 0.165 | 24.80 | 6.3 ×10-7 |
| Diet.ST | 4 | 0.011 | 1.58 | 0.21 |
| Diet.AT.ST | 4 | 0.009 | 1.32 | 0.29 |
| Error | 26 | 0.007 | ||
| Between display times within storage lengths within pens | ||||
| Display time (DT) | 1 | 36.18 | 1889.7 | 1.5 × 10-36 |
| ST.DT | 2 | 3.69 | 192.9 | 6.8 × 10-22 |
| AT.DT | 1 | 0.010 | 0.54 | 0.47 |
| AT.ST.DT | 2 | 0.064 | 3.32 | 0.046 |
| Diet.DT | 2 | 0.028 | 1.47 | 0.24 |
| Diet.ST.DT | 4 | 0.002 | 0.12 | 0.97 |
| Diet.AT.DT | 2 | 0.020 | 1.05 | 0.36 |
| Diet.AT.ST.DT | 4 | 0.018 | 0.93 | 0.46 |
| Error | 42 | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ponnampalam, E.N.; Butler, K.L.; Muir, S.K.; Plozza, T.E.; Kerr, M.G.; Brown, W.G.; Jacobs, J.L.; Knight, M.I. Lipid Oxidation and Colour Stability of Lamb and Yearling Meat (Muscle longissimus lumborum) from Sheep Supplemented with Camelina-Based Diets after Short-, Medium-, and Long-Term Storage. Antioxidants 2021, 10, 166. https://doi.org/10.3390/antiox10020166
Ponnampalam EN, Butler KL, Muir SK, Plozza TE, Kerr MG, Brown WG, Jacobs JL, Knight MI. Lipid Oxidation and Colour Stability of Lamb and Yearling Meat (Muscle longissimus lumborum) from Sheep Supplemented with Camelina-Based Diets after Short-, Medium-, and Long-Term Storage. Antioxidants. 2021; 10(2):166. https://doi.org/10.3390/antiox10020166
Chicago/Turabian StylePonnampalam, Eric N., Kym L. Butler, Stephanie K. Muir, Tim E. Plozza, Matthew G. Kerr, Wayne G. Brown, Joe L. Jacobs, and Matthew I. Knight. 2021. "Lipid Oxidation and Colour Stability of Lamb and Yearling Meat (Muscle longissimus lumborum) from Sheep Supplemented with Camelina-Based Diets after Short-, Medium-, and Long-Term Storage" Antioxidants 10, no. 2: 166. https://doi.org/10.3390/antiox10020166