
The Impact of Melatonin Supplementation and NLRP3 Inflammasome Deletion on Age-Accompanied Cardiac Damage

 ,
,  , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Animals
2.2. Magnetic Resonance Imaging (MRI)
2.3. Tissue Preparation for Histological Examination
2.4. Transmission Electron Microscopy (TEM)
2.5. Fluorescent Detection of Apoptotic Nuclei
2.6. Morphometrical Analysis
2.7. Real-Time Reverse Transcription Polymerase Chain Reaction (RT-PCR)
2.8. Statistical Analysis
3. Results
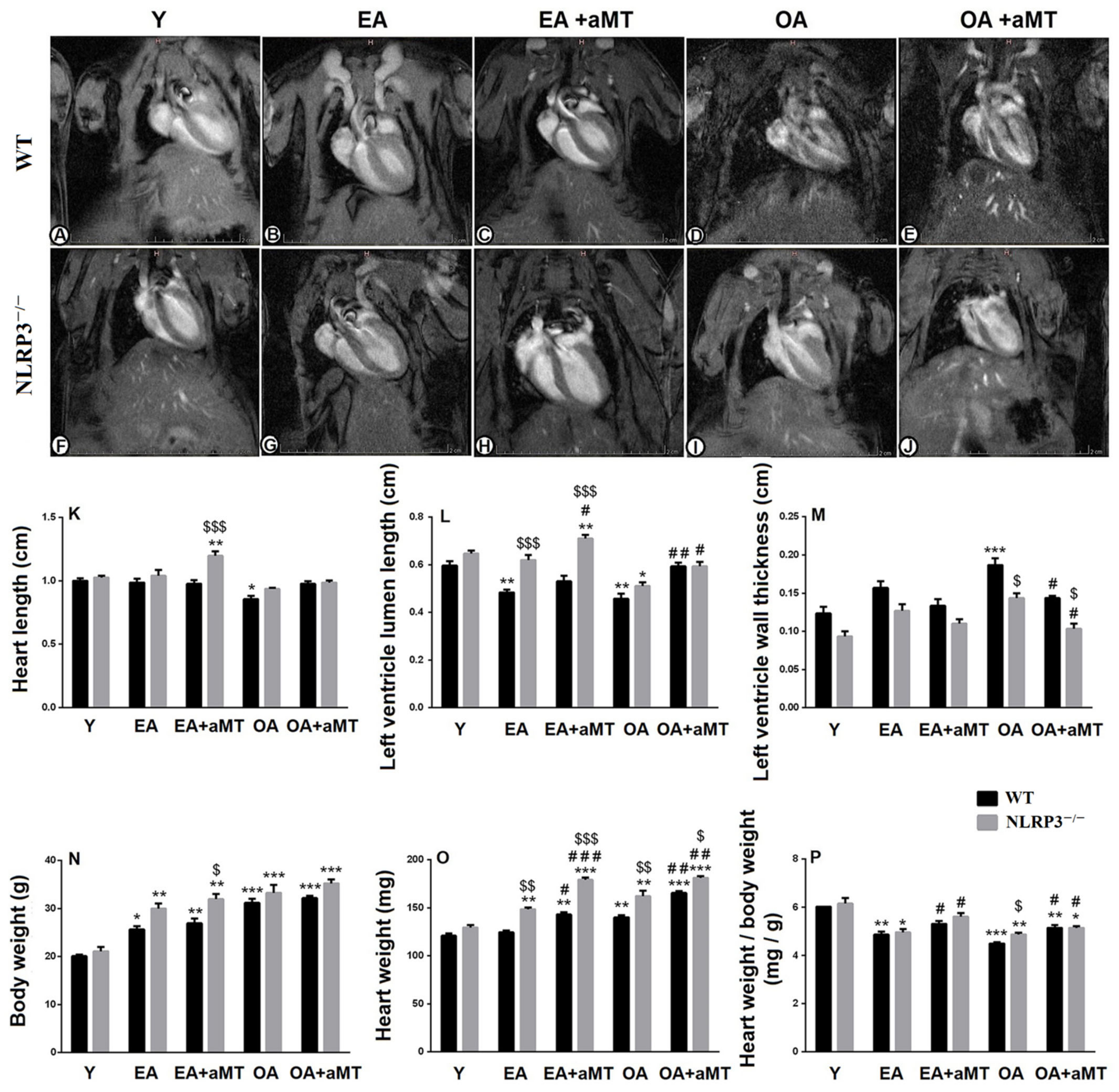
3.1. NLRP3 Absence and Melatonin Administration Restored Left Ventricular Lumen and Inhibited Thickening of Its Wall during Aging
3.2. NLRP3 Deficiency and Melatonin Therapy Enhanced Cardiac Anthropometric Parameters during Aging
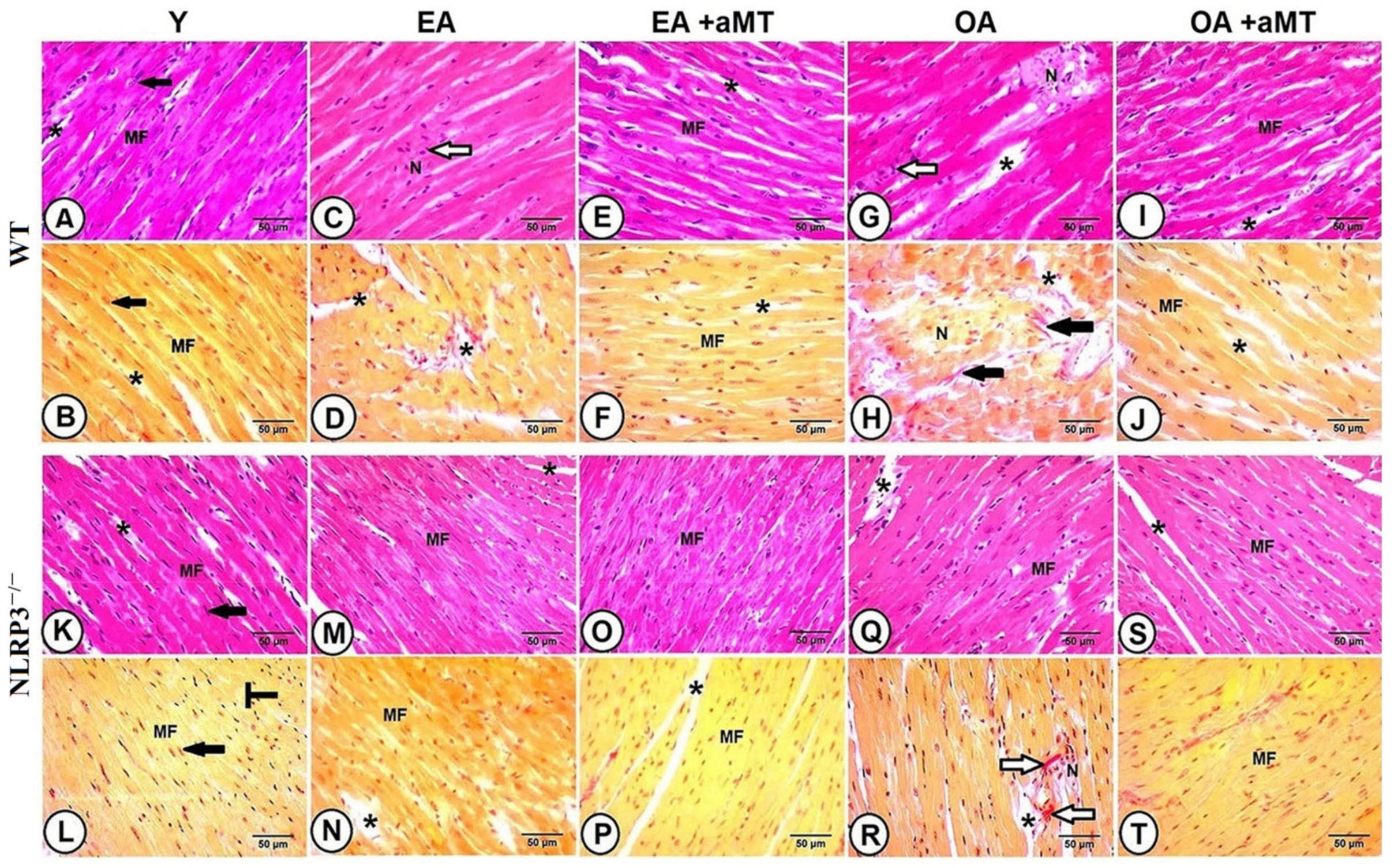
3.3. NLRP3 Deletion and Melatonin Supplementation Reduced Age-Related Histological and Morphometrical Alterations of the Cardiac Myocytes, and Minimized Hypertrophy-Associated Genes as Well as Inflammatory Cytokines Genes
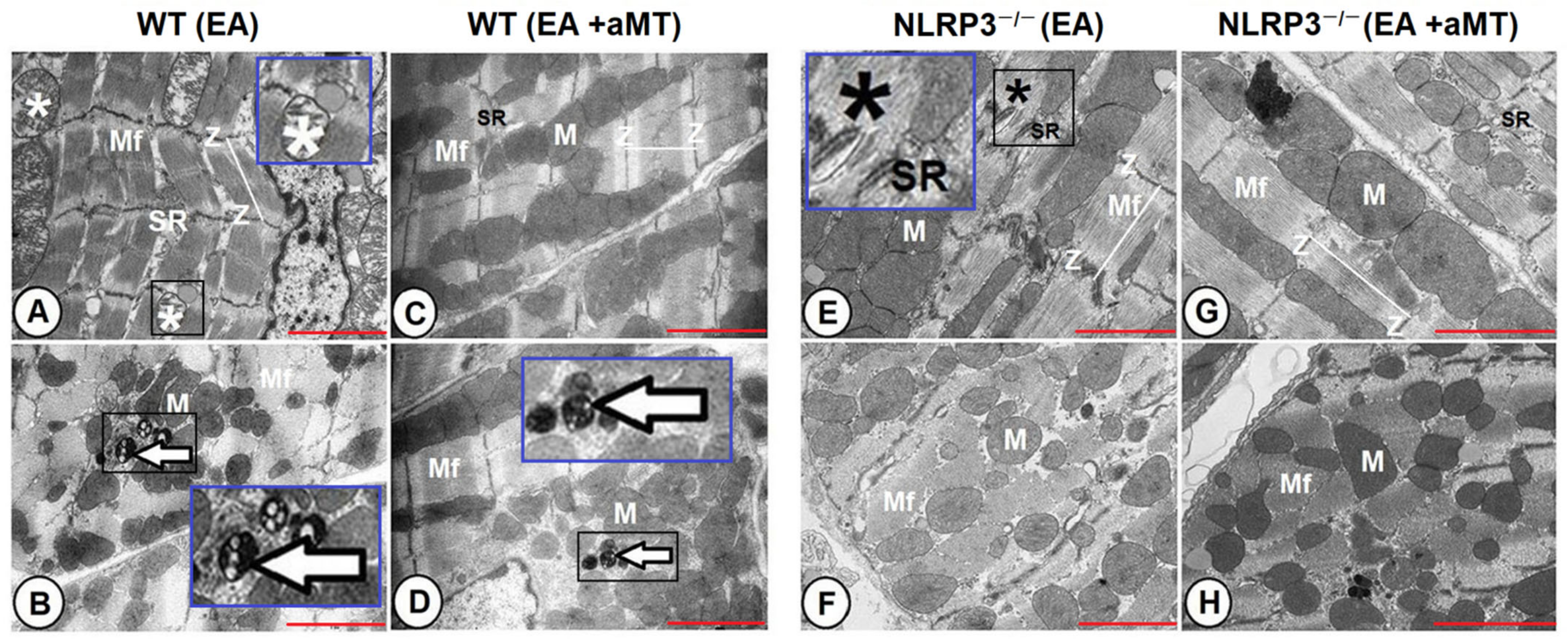
3.4. NLRP3 Ablation and Melatonin Administration Conserved Cardiac Muscle Ultrastructure during Aging
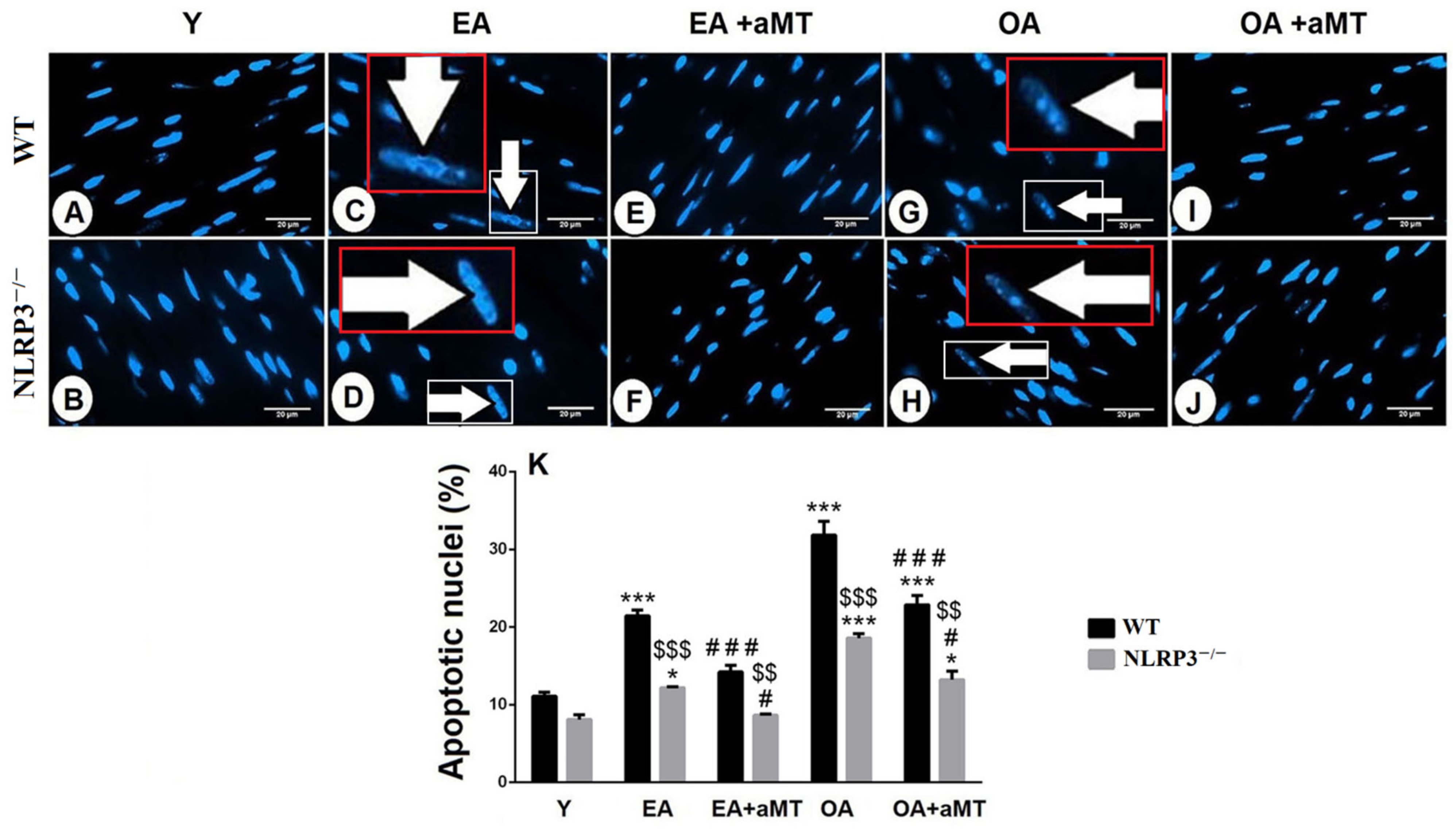
3.5. NLRP3 Absence and Melatonin Treatment Diminished Cardiac Apoptosis during Aging
4. Discussion
5. Conclusions
6. Limitation
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Martin-Cano, F.E.; Camello-Almaraz, C.; Acuña-Castroviejo, D.; Pozo, M.J.; Camello, P.J. Age-related changes in mitochondrial function of mouse colonic smooth muscle: Beneficial effects of melatonin. J. Pineal Res. 2014, 56, 163–174. [Google Scholar] [CrossRef]
- Figueiredo, P.A.; Ferreira, R.M.; Appell, H.J.; Duarte, J.A. Age-induced morphological, biochemical, and functional alterations in isolated mitochondria from murine skeletal muscle. J. Gerontol. A Biol. Sci. Med. Sci. 2008, 63, 350–359. [Google Scholar] [CrossRef] [Green Version]
- Ford, E.S.; Li, C.; Zhao, G.; Pearson, W.S.; Capewell, S. Trends in the prevalence of low risk factor burden for cardiovascular disease among United States adults. Circulation 2009, 120, 1181–1188. [Google Scholar] [CrossRef] [Green Version]
- Gómez-Pardo, E.; Fernández-Alvira, J.M.; Vilanova, M.; Haro, D.; Martínez, R.; Carvajal, I.; Carral, V.; Rodríguez, C.; de Miguel, M.; Bodega, P.; et al. A comprehensive lifestyle peer group-based intervention on cardiovascular risk factors: The randomized controlled fifty-fifty program. J. Am. Coll. Cardiol. 2016, 67, 476–485. [Google Scholar] [CrossRef] [PubMed]
- Finkel, T. The metabolic regulation of aging. Nat. Med. 2015, 21, 1416–1423. [Google Scholar] [CrossRef] [PubMed]
- North, B.J.; Sinclair, D.A. The intersection between aging and cardiovascular disease. Circ. Res. 2012, 110, 1097–1108. [Google Scholar] [CrossRef]
- Visser, M.; Schaap, L.A. Consequences of sarcopenia. Clin. Geriatr Med. 2011, 27, 387–399. [Google Scholar] [CrossRef] [PubMed]
- Curcio, F.; Testa, G.; Liguori, I.; Papillo, M.; Flocco, V.; Panicara, V.; Galizia, G.; Della-Morte, D.; Gargiulo, G.; Cacciatore, F.; et al. Sarcopenia and heart failure. Nutrients 2020, 12, 211. [Google Scholar] [CrossRef] [Green Version]
- Lakatta, E.G. Age-associated cardiovascular changes in health: Impact on cardiovascular disease in older persons. Heart Fail. Rev. 2002, 7, 29–49. [Google Scholar] [CrossRef]
- Treuting, P.M.; Linford, N.J.; Knoblaugh, S.E.; Emond, M.J.; Morton, J.F.; Martin, G.M.; Rabinovitch, P.S.; Ladiges, W.C. Reduction of age—Associated pathology in old mice by overexpression of catalase in mitochondria. J. Gerontol. A Biol. Sci. Med. Sci. 2008, 63, 813–822. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dai, D.F.; Santana, L.F.; Vermulst, M.; Tomazela, D.M.; Emond, M.J.; MacCoss, M.J.; Gollahon, K.; Martin, G.M.; Loeb, L.A.; Ladiges, W.C.; et al. Overexpression of catalase targeted to mitochondria attenuates murine cardiac aging. Circulation 2009, 119, 2789–2797. [Google Scholar] [CrossRef]
- Bullón, P.; Cano-García, F.J.; Alcocer-Gómez, E.; Varela-López, A.; Roman-Malo, L.; Ruiz-Salmerón, R.J.; Quiles, J.L.; Navarro-Pando, J.M.; Battino, M.; Ruiz-Cabello, J.; et al. Could NLRP3-inflammasome be a cardiovascular risk biomarker in acute myocardial infarction patients? Antioxid. Redox Signal. 2017, 27, 269–275. [Google Scholar] [CrossRef] [Green Version]
- Franceschi, C.; Bonafè, M.; Valensin, S.; Olivieri, F.; De Luca, M.; Ottaviani, E.; De Benedictis, G. Inflamm-aging. An evolutionary perspective on immunosenescence. Ann. N.Y. Acad. Sci. 2000, 908, 244–254. [Google Scholar] [CrossRef]
- Liu, D.; Zeng, X.; Li, X.; Mehta, J.L.; Wang, X. Role of NLRP3 inflammasome in the pathogenesis of cardiovascular diseases. Basic Res. Cardiol. 2017, 113, 5. [Google Scholar] [CrossRef]
- McBride, M.J.; Foley, K.P.; D’Souza, D.M.; Li, Y.E.; Lau, T.C.; Hawke, T.J.; Schertzer, J.D. The NLRP3 inflammasome con-tributes to sarcopenia and lower muscle glycolytic potential in old mice. Am. J. Physiol. Endocrinol. Metab. 2017, 313, E222–E232. [Google Scholar] [CrossRef] [Green Version]
- Volt, H.; García, J.A.; Doerrier, C.; Díaz-Casado, M.E.; Guerra-Librero, A.; López, L.C.; Escames, G.; Tresguerres, J.A.; Acuña-Castroviejo, D. Same molecule but different expres-sion: Aging and sepsis trigger NLRP3 inflammasome activation, a target of melatonin. J. Pineal Res. 2016, 60, 193–205. [Google Scholar] [CrossRef]
- Latz, E.; Xiao, T.S.; Stutz, A. Activation and regulation of the inflammasomes. Nat. Rev. Immunol. 2013, 13, 397–411. [Google Scholar] [CrossRef] [PubMed]
- Man, S.M.; Kanneganti, T.D. Regulation of inflammasome activation. Immunol. Rev. 2015, 265, 6–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.; Davies, K.J.A.; Forman, H.J. Oxidative stress response and Nrf2 signaling in aging. Free Radic. Biol. Med. 2015, 88, 314–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmidlin, C.J.; Dodson, M.B.; Madhavan, L.; Zhang, D.D. Redox regulation by NRF2 in aging and disease. Free Radic. Biol. Med. 2019, 134, 702–707. [Google Scholar] [CrossRef]
- Acuña-Castroviejo, D.; Escames, G.; Venegas, C.; Díaz-Casado, M.E.; Lima-Cabello, E.; López, L.C.; Rosales-Corral, S.; Tan, D.X.; Reiter, R.J. Extrapineal melatonin: Sources, regulation, and potential functions. Cell. Mol. Life Sci. 2014, 71, 2997–3025. [Google Scholar] [CrossRef] [PubMed]
- García, J.A.; Volt, H.; Venegas, C.; Doerrier, C.; Escames, G.; López, L.C.; Acuña-Castroviejo, D. Disruption of the NF-κB/NLRP3 connection by melatonin requires retinoid-related orphan receptor-α and blocks the septic response in mice. FASEB J. 2015, 29, 3863–3875. [Google Scholar] [CrossRef] [PubMed]
- Hossain, M.F.; Wang, N.; Chen, R.; Li, S.; Roy, J.; Uddin, M.G.; Li, Z.; Lim, L.W.; Song, Y.Q. Exploring the multifunctional role of melatonin in regulating autophagy and sleep to mitigate Alzheimer's disease neuropathology. Ageing Res. Rev. 2021, 67, 101304. [Google Scholar] [CrossRef] [PubMed]
- Bonmati-Carrion, M.A.; Tomas-Loba, A. Melatonin and Cancer: A Polyhedral Network Where the Source Matters. Antioxidants 2021, 10, 210. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Palanca, P.; Méndez-Blanco, C.; Fondevila, F.; Tuñón, M.J.; Reiter, R.J.; Mauriz, J.L.; González-Gallego, J. Melatonin as an Antitumor Agent against Liver Cancer: An Updated Systematic Review. Antioxidants 2021, 10, 103. [Google Scholar] [CrossRef]
- Cardinali, D.P. Melatonin and healthy aging. Vitam. Horm. 2021, 115, 67–88. [Google Scholar] [CrossRef]
- Ferlazzo, N.; Andolina, G.; Cannata, A.; Costanzo, M.G.; Rizzo, V.; Currò, M.; Ientile, R.; Caccamo, D. Is Melatonin the Cornucopia of the 21st Century? Antioxidants 2020, 9, 1088. [Google Scholar] [CrossRef]
- Leonardo-Mendonça, R.C.; Ocaña-Wilhelmi, J.; de Haro, T.; de Teresa-Galván, C.; Guerra-Hernández, E.; Rusanova, I.; Fernández-Ortiz, M.; Sayed, R.K.A.; Escames, G.; Acuña-Castroviejo, D. The benefit of a supplement with the antioxidant melatonin on redox status and muscle damage in resistance-trained athletes. Appl. Physiol. Nutr. Metab. 2017, 42, 700–707. [Google Scholar] [CrossRef]
- Rodríguez, M.I.; Carretero, M.; Escames, G.; López, L.C.; Maldonado, M.D.; Tan, D.X.; Reiter, R.J.; Acuña-Castroviejo, D. Chronic melatonin treatment prevents age-dependent cardiac mitochondrial dysfunction in senescence-accelerated mice. Free Radic. Res. 2017, 41, 15–24. [Google Scholar] [CrossRef]
- Sayed, R.K.A.; Fernández-Ortiz, M.; Diaz-Casado, M.E.; Rusanova, I.; Rahim, I.; Escames, G.; López, L.C.; Mokhtar, D.M.; Acuña-Castroviejo, D. The protective effect of melatonin against age-associated, sarcopenia-dependent tubular aggregate formation, lactate depletion, and mitochondrial changes. J. Gerontol. A Biol. Sci. Med. Sci. 2018, 73, 1330–1338. [Google Scholar] [CrossRef]
- Sayed, R.K.A.; Fernández-Ortiz, M.; Diaz-Casado, M.E.; Aranda-Martínez, P.; Fernández-Martínez, J.; Guerra-Librero, A.; Escames, G.; López, L.C.; Alsaadawy, R.M.; Acuña-Castroviejo, D. Lack of NLRP3 inflammasome activation reduces age-dependent sarcopenia and mitochondrial dysfunction, favoring the prophylactic effect of melatonin. J. Gerontol. A Biol. Sci. Med. Sci. 2019, 74, 1699–1708. [Google Scholar] [CrossRef] [PubMed]
- Obayashi, K.; Saeki, K.; Maegawa, T.; Iwamoto, J.; Sakai, T.; Otaki, N.; Kataoka, H.; Kurumatani, N. Melatonin secretion and muscle strength in elderly individuals: A cross-sectional study of the HEIJO-KYO cohort. J. Gerontol. A Biol. Sci. Med. Sci. 2016, 71, 1235–1240. [Google Scholar] [CrossRef]
- Zhong, J.; Liu, Y. Melatonin and age-related cardiovascular diseases. Aging Med. 2018, 1, 197–203. [Google Scholar] [CrossRef]
- Rahim, I.; Djerdjouri, B.; Sayed, R.K.; Fernández-Ortiz, M.; Fernández-Gil, B.; Hidalgo-Gutiérrez, A.; López, L.C.; Escames, G.; Reiter, R.J.; Acuña-Castroviejo, D. Melatonin administration to wild-type mice and non-treated NLRP3 mutant mice share similar inhibition of the inflammatory response during sepsis. J. Pineal Res. 2017, 63, e12410. [Google Scholar] [CrossRef]
- López, A.; García, J.A.; Escames, G.; Venegas, C.; Ortiz, F.; López, L.C.; Acuña-Castroviejo, D. Melatonin protects the mitochondria from oxidative damage reducing oxygen consumption, membrane potential, and superoxide anion production. J. Pineal Res. 2009, 46, 188–198. [Google Scholar] [CrossRef]
- Wree, A.; McGeough, M.D.; Peña, C.A.; Schlattjan, M.; Li, H.; Inzaugarat, M.E.; Messer, K.; Canbay, A.; Hoffman, H.M.; Feldstein, A.E. NLRP3 inflammasome activation is required for fibrosis development in NAFLD. J. Mol. Med. 2014, 92, 1069–1082. [Google Scholar] [CrossRef] [Green Version]
- Fernández-Ortiz, M.; Sayed, R.K.A.; Fernández-Martínez, J.; Cionfrini, A.; Aranda-Martínez, P.; Escames, G.; de Haro, T.; Acuña-Castroviejo, D. Melatonin/Nrf2/NLRP3 connection in mouse heart mitochondria during aging. Antioxidants 2020, 9, 1187. [Google Scholar] [CrossRef] [PubMed]
- Ebihara, S.; Marks, T.; Hudson, D.J.; Menaker, M. Genetic control of melatonin synthesis in the pineal gland of the mouse. Science 1986, 231, 491–493. [Google Scholar] [CrossRef]
- Roseboom, P.H.; Namboodiri, M.A.; Zimonjic, D.B.; Popescu, N.C.; Rodriguez, I.R.; Gastel, J.A.; Klein, D.C. Natural melatonin ’knockdown’ in C57BL/6J mice: Rare mechanism truncates serotonin N-acetyltransferase. Brain Res. Mol. Brain Res. 1998, 63, 189–197. [Google Scholar] [CrossRef]
- Khaldy, H.; León, J.; Escames, G.; Bikjdaouene, L.; García, J.J.; Acuña-Castroviejo, D. Circadian rhythms of dopamine and dihydroxyphenyl acetic acid in the mouse striatum: Effects of pinealectomy and of melatonin treatment. Neuroendocrinology 2002, 75, 201–208. [Google Scholar] [CrossRef] [PubMed]
- López, A.; Ortiz, F.; Doerrier, C.; Venegas, C.; Fernández-Ortiz, M.; Aranda, P.; Díaz-Casado, M.E.; Fernández-Gil, B.; Barriocanal-Casado, E.; Escames, G.; et al. Mitochondrial impairment and melatonin protection in parkinsonian mice do not depend of inducible or neuronal nitric oxide synthases. PLoS ONE 2017, 12, e0183090. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, M.I.; Escames, G.; López, L.C.; López, A.; García, J.A.; Ortiz, F.; Sánchez, V.; Romeu, M.; Acuña-Castroviejo, D. Improved mitochondrial function and increased life span after chronic melatonin treatment in senescent prone mice. Exp. Gerontol. 2008, 43, 749–756. [Google Scholar] [CrossRef] [Green Version]
- Venegas, C.; García, J.A.; Escames, G.; Ortiz, F.; López, A.; Doerrier, C.; García-Corzo, L.; López, L.C.; Reiter, R.J.; Acuña-Castroviejo, D. Extrapineal melatonin: Analysis of its subcellular distribution and daily fluctuations. J. Pineal Res. 2012, 52, 217–227. [Google Scholar] [CrossRef]
- Teodoro, B.G.; Baraldi, F.G.; Sampaio, I.H.; Bomfim, L.H.; Queiroz, A.L.; Passos, M.A.; Carneiro, E.M.; Alberici, L.C.; Gomis, R.; Amaral, F.G.; et al. Melatonin prevents mitochondrial dysfunction and insulin resistance in rat skeletal muscle. J. Pineal Res. 2014, 57, 155–167. [Google Scholar] [CrossRef] [PubMed]
- Galley, H.F.; Lowes, D.A.; Allen, L.; Cameron, G.; Aucott, L.S.; Webster, N.R. Melatonin as a potential therapy for sepsis: A phase I dose escalation study and an ex vivo whole blood model under conditions of sepsis. J. Pineal Res. 2014, 56, 427–438. [Google Scholar] [CrossRef] [Green Version]
- Weishaupt, J.H.; Bartels, C.; Pölking, E.; Dietrich, J.; Rohde, G.; Poeggeler, B.; Mertens, N.; Sperling, S.; Bohn, M.; Hüther, G.; et al. Reduced oxidative damage in ALS by high-dose enteral melatonin treatment. J. Pineal Res. 2006, 41, 313–323. [Google Scholar] [CrossRef]
- Toldo, S.; Abbate, A. The NLRP3 inflammasome in acute myocardial infarction. Nat. Rev. Cardiol. 2018, 15, 203–214. [Google Scholar] [CrossRef] [PubMed]
- Marín-Aguilar, F.; Castejón-Vega, B.; Alcocer-Gómez, E.; Lendines-Cordero, D.; Cooper, M.A.; de la Cruz, P.; Andújar-Pulido, E.; Pérez-Alegre, M.; Muntané, J.; Pérez-Pulido, A.J.; et al. NLRP3 Inflammasome Inhibition by MCC950 in Aged Mice Improves Health via Enhanced Autophagy and PPARα Activity. J. Gerontol. A Biol. Sci. Med. Sci. 2020, 75, 1457–1464. [Google Scholar] [CrossRef]
- Boyle, A.J.; Shih, H.; Hwang, J.; Ye, J.; Lee, B.; Zhang, Y.; Kwon, D.; Jun, K.; Zheng, D.; Sievers, R.; et al. Cardiomyopathy of aging in the mammalian heart is characterized by myocardialhypertrophy, fibrosis and a predisposition towards cardiomyocyte apoptosis and autophagy. Exp. Gerontol. 2011, 46, 549–559. [Google Scholar] [CrossRef] [Green Version]
- Carnes, C.A.; Geisbuhler, T.P.; Reiser, P.J. Age-dependent changes in contraction and regional myocardial myosin heavy chain isoform expression in rats. J. Appl. Physiol. 2004, 97, 446–453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pandya, K.; Kim, H.S.; Smithies, O. Fibrosis, not cell size, delineates β-myosin heavy chain reexpression during cardiac hypertrophy and normal aging in vivo. Proc. Natl. Acad. Sci. USA 2006, 103, 16864–16869. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kane, A.E.; Bisset, E.S.; Heinze-Milne, S.; Keller, K.M.; Grandy, S.A.; Howlett, S.E. Maladaptive changes associated with cardiac aging are sex-specific and graded by frailty and inflammation in C57BL/6 mice. J. Gerontol. A Biol. Sci. Med. Sci. 2021, 76, 233–243. [Google Scholar] [CrossRef]
- Huang, N.; Kny, M.; Riediger, F.; Busch, K.; Schmidt, S.; Luft, F.C.; Slevogt, H.; Fielitz, J. Deletion of Nlrp3 protects from inflammation-induced skeletal muscle atrophy. Intensive Care Med. Exp. 2017, 5, 3. [Google Scholar] [CrossRef] [Green Version]
- Hepple, R.T. Impact of aging on mitochondrial function in cardiac and skeletal muscle. Free Radic. Biol. Med. 2016, 98, 177–186. [Google Scholar] [CrossRef] [PubMed]
- Zampino, M.; Spencer, R.G.; Fishbein, K.W.; Simonsick, E.M.; Ferrucci, L. Cardiovascular health and mitochondrial function: Testing an association. J. Gerontol. A Biol. Sci. Med. Sci. 2021, 76, 361–367. [Google Scholar] [CrossRef]
- Liu, F.; Wanigatunga, A.A.; Zampino, M.; Knuth, N.D.; Simonsick, E.M.; Schrack, J.A.; Ferrucci, L. Association of mitochondrial function, substrate utilization, and anaerobic metabolism with age-related perceived fatigability. J. Gerontol. A Biol. Sci. Med. Sci. 2021, 76, 426–433. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Trevino, M.B.; Wang, M.; Gardell, S.J.; Ayala, J.E.; Han, X.; Kelly, D.P.; Goodpaster, B.H.; Vega, R.B.; Coen, P.M. Impaired mitochondrial energetics characterize poor early recovery of muscle mass following hind limb unloading in old mice. J. Gerontol. A Biol. Sci. Med. Sci. 2018, 73, 1313–1322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marín-Aguilar, F.; Lechuga-Vieco, A.V.; Alcocer-Gómez, E.; Castejón-Vega, B.; Lucas, J.; Garrido, C.; Peralta-Garcia, A.; Pérez-Pulido, A.J.; Varela-López, A.; Quiles, J.L.; et al. NLRP3 inflammasome suppression improves longevity and prevents cardiac aging in male mice. Aging Cell 2020, 19, e13050. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Madeo, F.; Zimmermann, A.; Maiuri, M.C.; Kroemer, G. Essential role for autophagy in life span extension. J. Clin. Investig. 2015, 125, 85–93. [Google Scholar] [CrossRef] [Green Version]
- Shirakabe, A.; Ikeda, Y.; Sciarretta, S.; Zablocki, D.K.; Sadoshima, J. Aging and Autophagy in the Heart. Circ. Res. 2016, 118, 1563–1576. [Google Scholar] [CrossRef] [Green Version]
- Taneike, M.; Yamaguchi, O.; Nakai, A.; Hikoso, S.; Takeda, T.; Mizote, I.; Oka, T.; Tamai, T.; Oyabu, J.; Murakawa, T.; et al. Inhibition of autophagy in the heart induces age-related cardiomyopathy. Autophagy 2010, 6, 600–606. [Google Scholar] [CrossRef] [PubMed]
- Salminen, A.; Kaarniranta, K.; Kauppinen, A. Inflammaging: Disturbed interplay between autophagy and inflammasomes. Aging 2012, 4, 166–175. [Google Scholar] [CrossRef] [Green Version]
- Borisov, A.B.; Carlson, B.M. Cell death in denervated skeletal muscle is distinct from classical apoptosis. Anat. Rec. 2000, 258, 305–318. [Google Scholar] [CrossRef]
- Adams, V.; Jiang, H.; Yu, J.; Möbius-Winkler, S.; Fiehn, E.; Linke, A.; Weigl, C.; Schuler, G.; Hambrecht, R. Apoptosis in skeletal myocytes of patients with chronic heart failure is associated with exercise intolerance. J. Am. Coll. Cardiol. 1999, 33, 959–965. [Google Scholar] [CrossRef] [Green Version]
- Pollack, M.; Leeuwenburgh, C. Apoptosis and aging: Role of the mitochondria. J. Gerontol. A Biol. Sci. Med. Sci. 2001, 56, B475–B482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kajstura, J.; Cheng, W.; Sarangarajan, R.; Li, P.; Li, B.; Nitahara, J.A.; Chapnick, S.; Reiss, K.; Olivetti, G.; Anversa, P. Necrotic and apoptotic myocyte cell death in the aging heart of Fischer 344 rats. Am. J. Physiol. 1996, 271, H1215–H1228. [Google Scholar] [CrossRef]
- Acuña-Castroviejo, D.; Noguiera-Navarro, M.; Reiter, R.; Escames, G. Melatonin actions in the heart; more than a hormone. Melatonin Res. 2018, 1, 21–26. [Google Scholar] [CrossRef]
- Sanchez-Hidalgo, M.; Alarcon de la Lastra, C.; Carrascosa-Salmoral, M.P.; Naranjo, M.C.; Gomez-Corvera, A.; Caballero, B.; Guerrero, J.M. Age-related changes in melatonin synthesis in rat extrapineal tissues. Exp. Gerontol. 2009, 44, 328–334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, R.; Ni, L.; Di, X.; Ma, B.; Niu, S.; Rong, Z.; Liu, C. Potential Role of Melatonin as an Adjuvant for Atherosclerotic Carotid Arterial Stenosis. Molecules 2021, 26, 811. [Google Scholar] [CrossRef]
- González, A.; Alonso-González, C.; González-González, A.; Menéndez-Menéndez, J.; Cos, S.; Martínez-Campa, C. Melatonin as an Adjuvant to Antiangiogenic Cancer Treatments. Cancers 2021, 13, 3263. [Google Scholar] [CrossRef]
- Farnoosh, G.; Akbariqomi, M.; Badri, T.; Bagheri, M.; Izadi, M.; saeedi-Boroujeni, A.; Rezaie, E.; Ghaleh, H.E.G.; Aghamallaei, H.; Fasihi-Ramandi, M.; et al. Efficacy of a Low Dose of Melatonin as an Adjunctive Therapy in Hospitalized Patients with COVID-19: A Randomized, Double-blind Clinical Trial. Arch. Med. Res. 2021. [Google Scholar] [CrossRef] [PubMed]
- Shokri, M.; Sajedi, F.; Mohammadi, Y.; Mehrpooya, M. Adjuvant use of melatonin for relieving symptoms of painful diabetic neuropathy: Results of a randomized, double-blinded, controlled trial. Eur. J. Clin. Pharmacol. 2021, 1–15. [Google Scholar] [CrossRef]
- Yang, Y.; Du, J.; Xu, R.; Shen, Y.; Yang, D.; Li, D.; Hu, H.; Pei, H.; Yang, Y. Melatonin alleviates angiotensin-II-induced cardiac hypertrophy via activating MICU1 pathway. Aging 2020, 13, 493–515. [Google Scholar] [CrossRef]
- Molpeceres, V.; Mauriz, J.L.; García-Mediavilla, M.V.; González, P.; Barrio, J.P.; González-Gallego, J. Melatonin is able to reduce the apoptotic liver changes induced by aging via inhibition of the intrinsic pathway of apoptosis. J. Gerontol. A Biol. Sci. Med. Sci. 2007, 62, 687–695. [Google Scholar] [CrossRef] [Green Version]
- Hu, S.; Zhu, P.; Zhou, H.; Zhang, Y.; Chen, Y. Melatonin-induced protective effects on cardiomyocytes against reperfusion injury partly through modulation of IP3R and SERCA2a via activation of ERK1. Arq. Bras. Cardiol. 2018, 110, 44–51. [Google Scholar] [CrossRef] [PubMed]
- Sayed, R.K.; Fernández-Ortiz, M.; Fernández-Martínez, J.; ArandaMartínez, P.; Guerra-Librero, A.; Rodríguez-Santana, C.; de Haro, T.; Escames, G.; Acuña-Castroviejo, D.; Rusanova, I. The Impact of Melatoninand NLRP3 inflammasome on the expression of microRNAs in aged muscle. Antioxidants 2021, 10, 524. [Google Scholar] [CrossRef]
- Martín, M.; Macías, M.; Escames, G.; León, J.; Acuña-Castroviejo, D. Melatonin but not vitamins C and E maintains glutathione homeostasis in t-butyl hydroperoxide-induced mitochondrial oxidative stress. FASEB J. 2000, 14, 1677–1679. [Google Scholar] [CrossRef] [PubMed]
- Escames, G.; León, J.; Macías, M.; Khaldy, H.; Acuña-Castroviejo, D. Melatonin counteracts lipopolysaccharide-induced expression and activity of mitochondrial nitric oxide synthase in rats. FASEB J. 2003, 17, 1–22. [Google Scholar] [CrossRef]
- Acuña-Castroviejo, D.; López, L.C.; Escames, G.; López, A.; García, J.A.; Reiter, R.J. Melatonin-mitochondria interplay in health and disease. Curr. Top. Med. Chem. 2011, 11, 221–240. [Google Scholar] [CrossRef] [PubMed]
- Coto-Montes, A.; Boga, J.A.; Tan, D.X.; Reiter, R.J. Melatonin as a potential agent in the treatment of sarcopenia. Int. J. Mol. Sci. 2016, 17, 1771. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Wild-Type Mice | NLRP3-Knockout Mice | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Y | EA | EA + aMT | OA | OA + aMT | Y | EA | EA + aMT | OA | OA + aMT | |
| Heart length (cm) | 1.00 ± 0.02 | 0.99 ± 0.03 | 0.98 ± 0.03 | 0.86 ± 0.02 | 0.98 ± 0.02 | 1.03 ± 0.01 | 1.04 ± 0.05 | 1.2 ± 0.03 | 0.94 ± 0.01 | 0.99 ± 0.01 |
| Left ventricle lumen length (cm) | 0.6 ± 0.02 | 0.48 ± 0.01 | 0.53 ± 0.02 | 0.46 ± 0.02 | 0.59 ± 0.01 | 0.65 ± 0.01 | 0.62 ± 0.02 | 0.71 ± 0.02 | 0.51 ± 0.01 | 0.59 ± 0.02 |
| Left ventricle wall thickness (cm) | 0.12 ± 0.01 | 0.16 ± 0.008 | 0.13 ± 0.006 | 0.19 ± 0.008 | 0.14 ± 0.003 | 0.09 ± 0.006 | 0.13 ± 0.008 | 0.11 ± 0.005 | 0.14 ± 0.007 | 0.10 ± 0.007 |
| Body weight (g) | 20.07 ± 0.35 | 25.63 ± 0.66 | 26.93 ± 0.97 | 31.17 ± 0.84 | 32.17 ± 0.47 | 21.10 ± 0.92 | 30 ± 1.02 | 31.93 ± 1.14 | 33.27 ± 1.68 | 35.23 ± 0.87 |
| Heart weight (mg) | 120.8 ± 2.3 | 124.4 ± 1.7 | 142.9 ± 2.6 | 140.1 ± 2.2 | 165.3 ± 2.1 | 129.5 ± 2.5 | 148.4 ± 1.9 | 179.0 ± 2.3 | 161.9 ± 5.9 | 181.1 ± 2 |
| Heart weight/body weight (mg/g) | 6.02 ± 0.01 | 4.86 ± 0.13 | 5.32 ± 0.12 | 4.5 ± 0.05 | 5.14 ± 0.12 | 6.15 ± 0.23 | 4.96 ± 0.14 | 5.62 ± 0.15 | 4.88 ± 0.07 | 5.15 ± 0.08 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sayed, R.K.A.; Fernández-Ortiz, M.; Rahim, I.; Fernández-Martínez, J.; Aranda-Martínez, P.; Rusanova, I.; Martínez-Ruiz, L.; Alsaadawy, R.M.; Escames, G.; Acuña-Castroviejo, D. The Impact of Melatonin Supplementation and NLRP3 Inflammasome Deletion on Age-Accompanied Cardiac Damage. Antioxidants 2021, 10, 1269. https://doi.org/10.3390/antiox10081269
Sayed RKA, Fernández-Ortiz M, Rahim I, Fernández-Martínez J, Aranda-Martínez P, Rusanova I, Martínez-Ruiz L, Alsaadawy RM, Escames G, Acuña-Castroviejo D. The Impact of Melatonin Supplementation and NLRP3 Inflammasome Deletion on Age-Accompanied Cardiac Damage. Antioxidants. 2021; 10(8):1269. https://doi.org/10.3390/antiox10081269
Chicago/Turabian StyleSayed, Ramy K. A., Marisol Fernández-Ortiz, Ibtissem Rahim, José Fernández-Martínez, Paula Aranda-Martínez, Iryna Rusanova, Laura Martínez-Ruiz, Reem M. Alsaadawy, Germaine Escames, and Darío Acuña-Castroviejo. 2021. "The Impact of Melatonin Supplementation and NLRP3 Inflammasome Deletion on Age-Accompanied Cardiac Damage" Antioxidants 10, no. 8: 1269. https://doi.org/10.3390/antiox10081269