
Growth, Health, and Gut Microbiota of Female Pacific White Shrimp, Litopenaeus vannamei Broodstock Fed Different Phospholipid Sources

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Diets
2.2. Growth Trial and Sampling
2.3. Antioxidant Capacity Related Parameter Assays
2.4. Immunity Related Parameters Assay
2.5. Gut Microbiota Analysis
2.6. Gut Microbiota and Biochemical Indexes Association Analysis
2.7. Statistical Analysis
3. Results
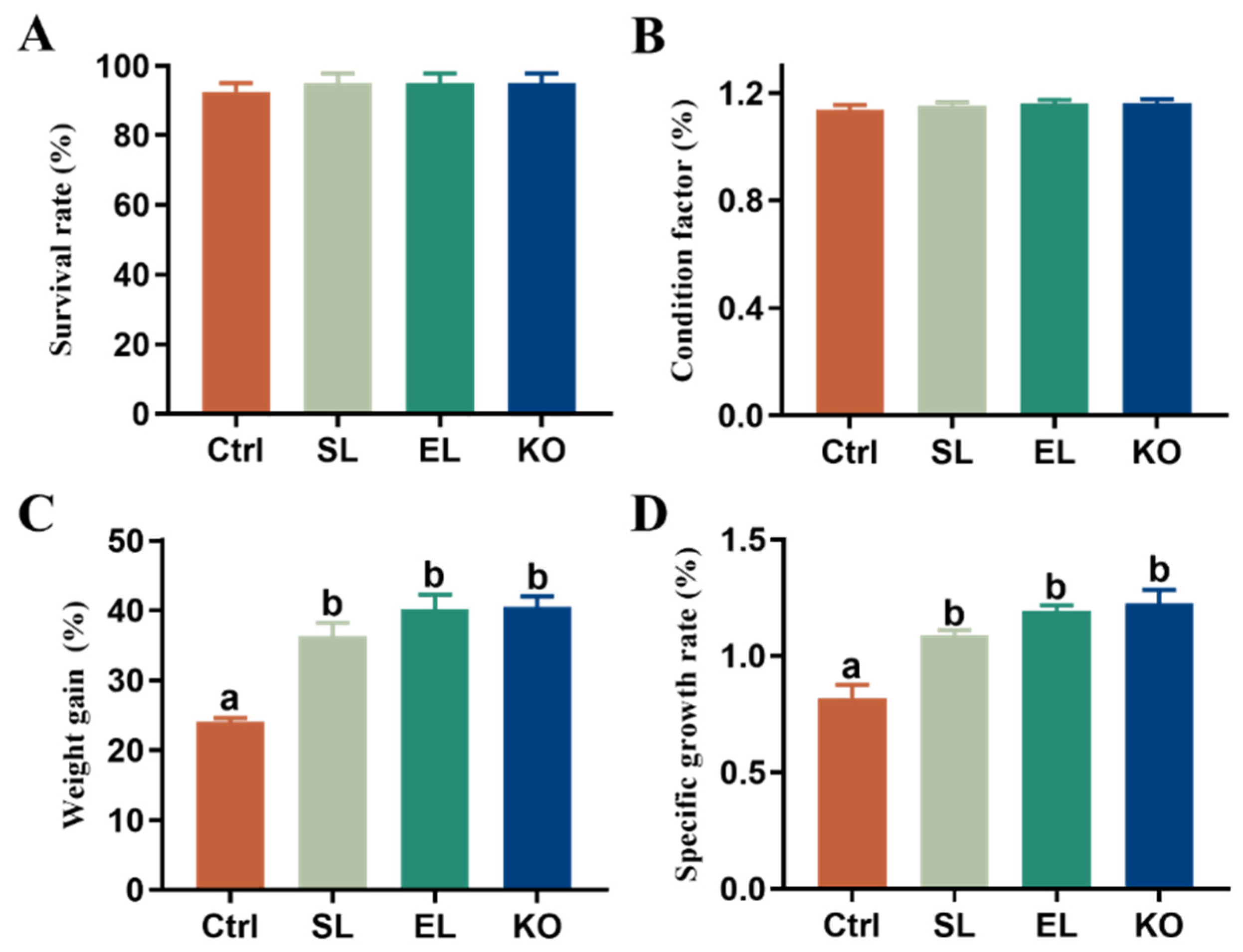
3.1. Growth Performance
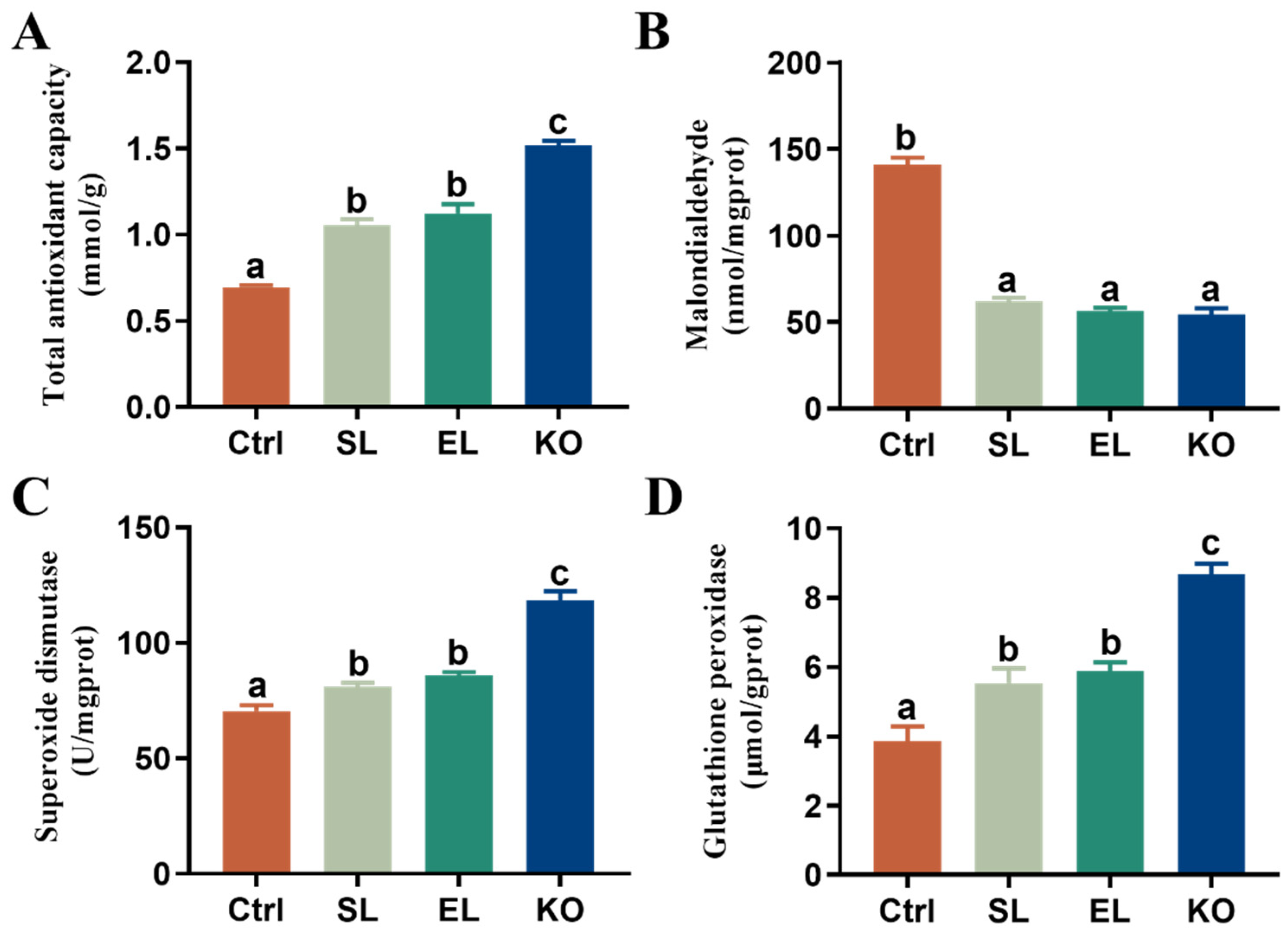
3.2. Antioxidant Capacity
3.3. Immune Responses
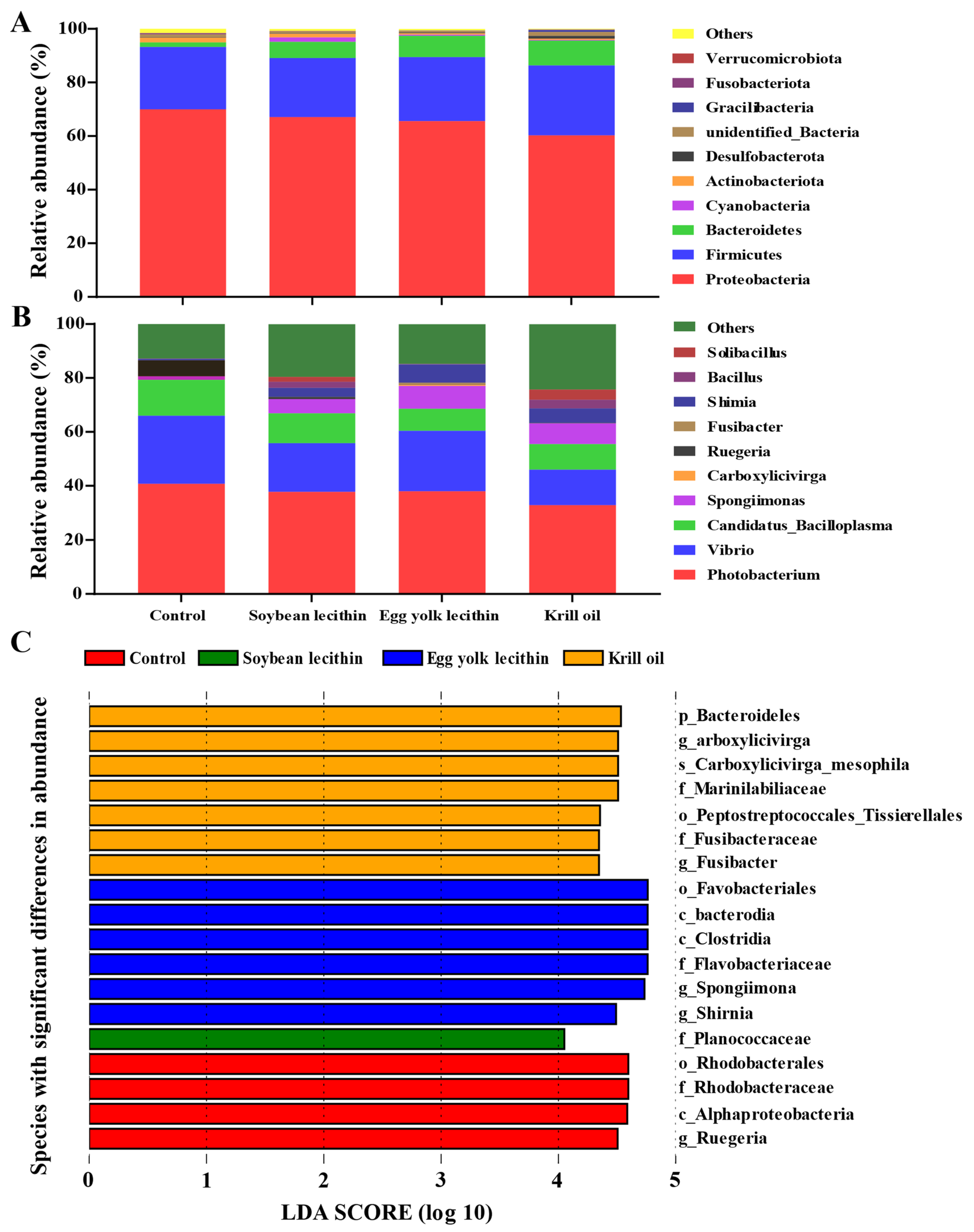
3.4. Gut Microbiota Analysis
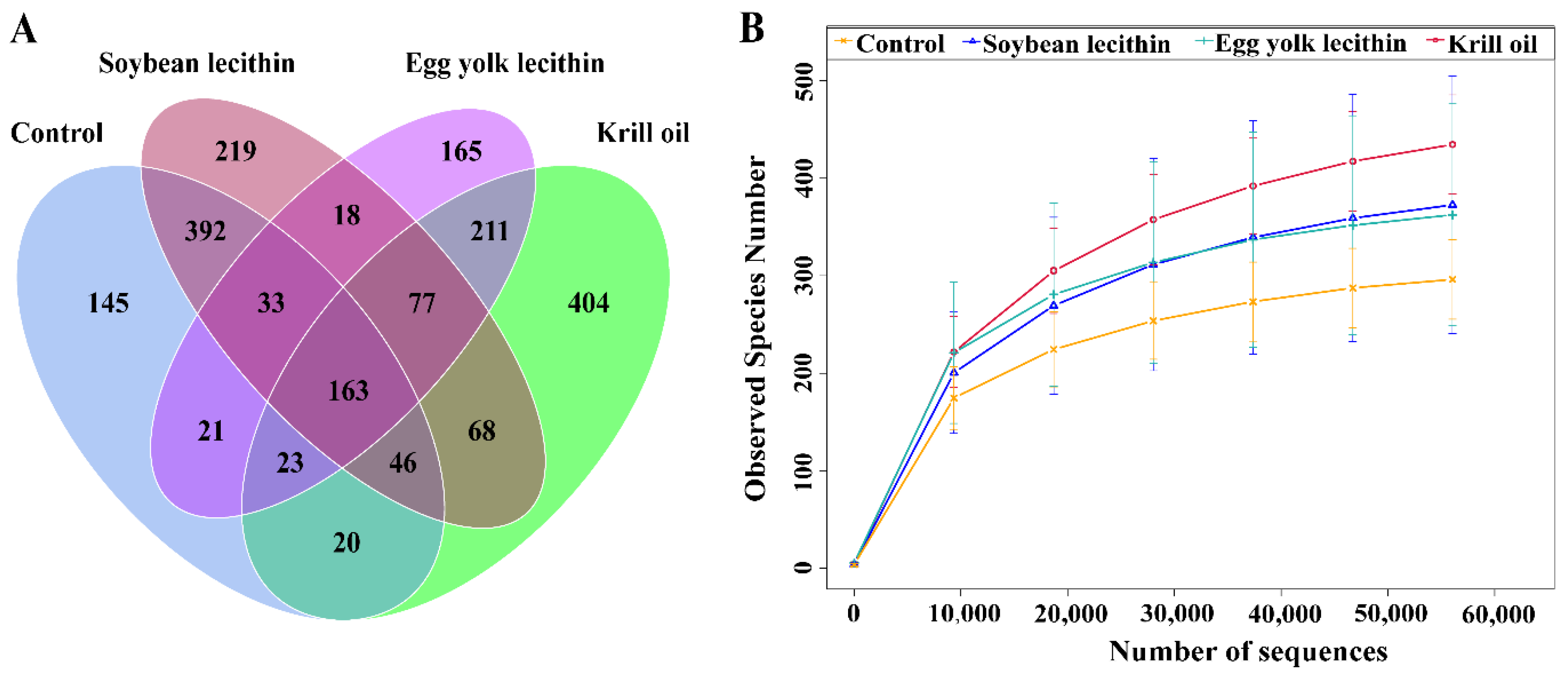
3.4.1. Composition of Gut Microbial Community
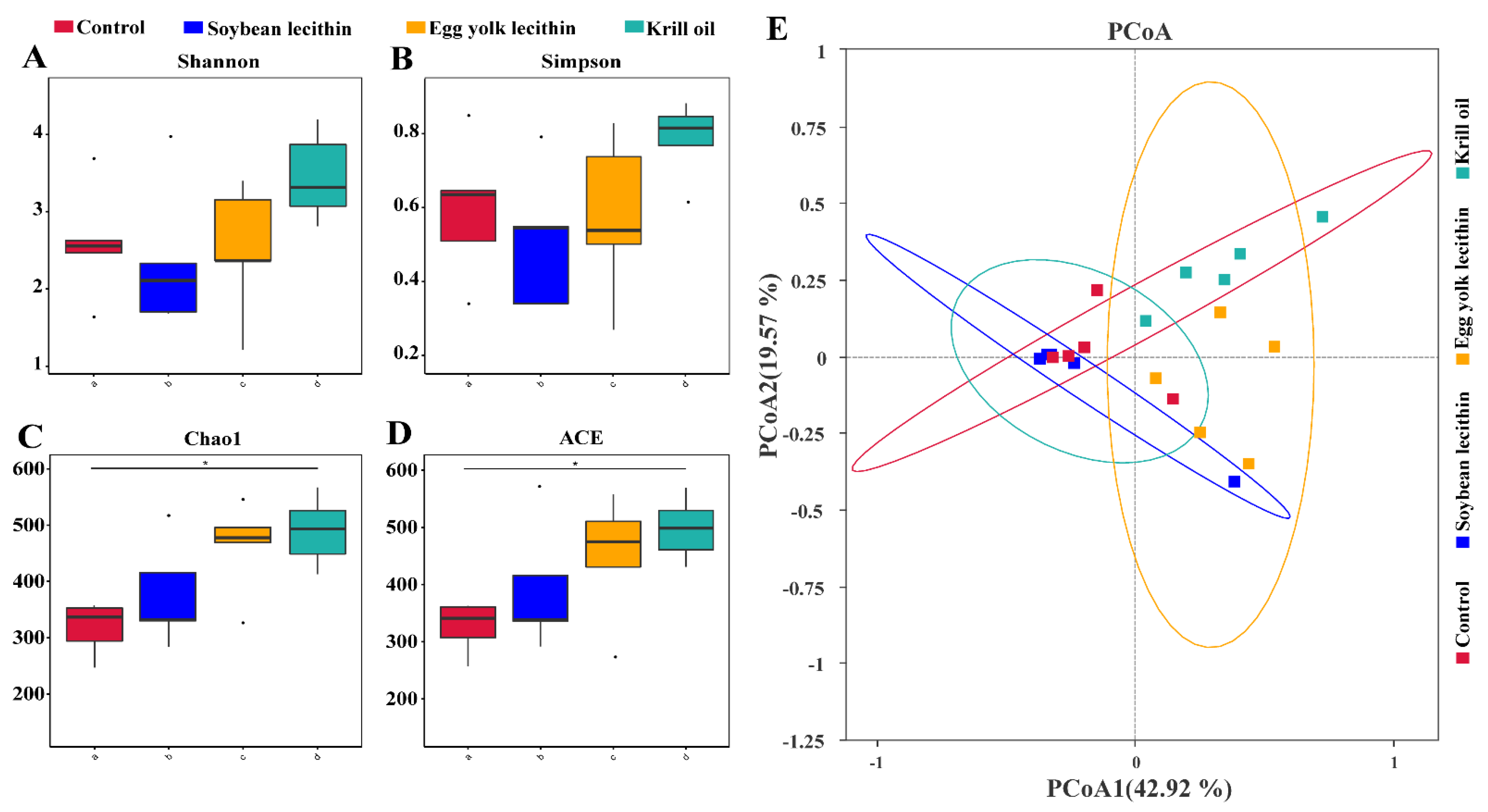
3.4.2. Diversity of Gut Microbial
3.4.3. Gut Microbiota Functional Prediction
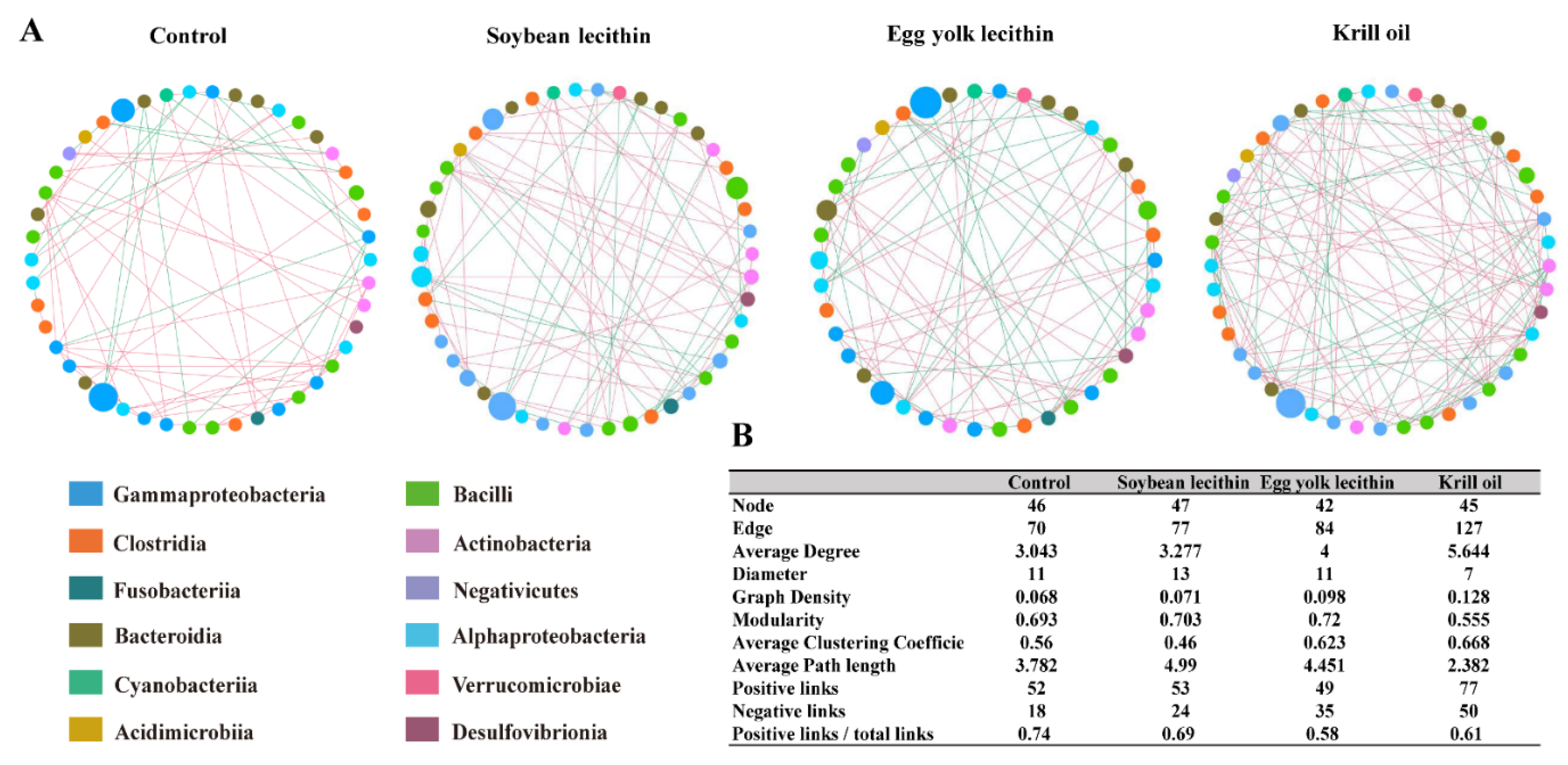
3.4.4. Gut Microbiota Network
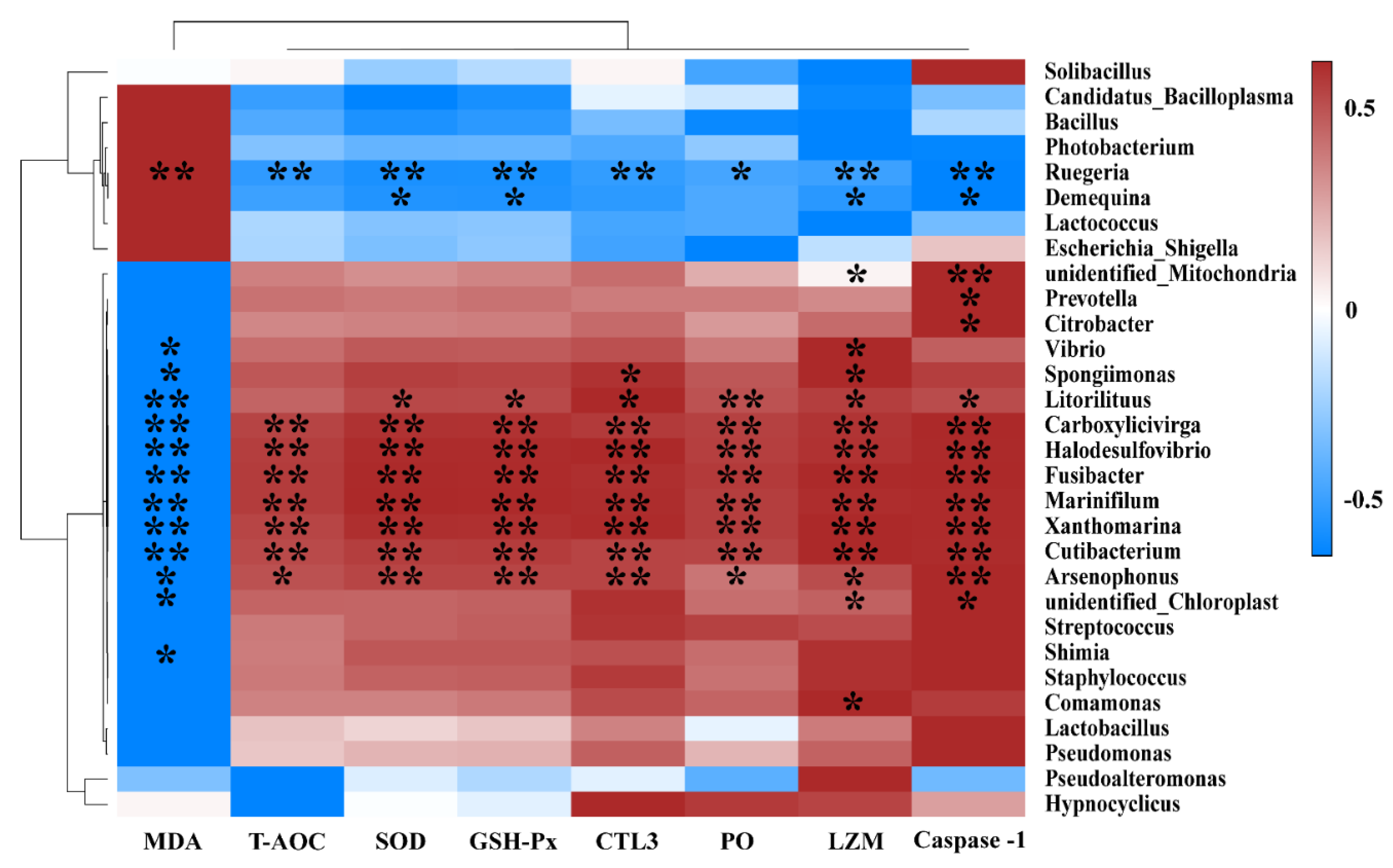
3.5. Gut Microbiota and Biochemical Indexes Association Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Garibay-Valdez, E.; Cicala, F.; Martinez-Porchas, M.; Gómez-Reyes, R.; Vargas-Albores, F.; Gollas-Galván, T.; Martínez-Córdova, L.R.; Calderón, K. Longitudinal Variations in the Gastrointestinal Microbiome of the White Shrimp, Litopenaeus Vannamei. PeerJ 2021, 9, e11827. [Google Scholar] [CrossRef] [PubMed]
- Thitamadee, S.; Prachumwat, A.; Srisala, J.; Jaroenlak, P.; Salachan, P.V.; Sritunyalucksana, K.; Flegel, T.W.; Itsathitphaisarn, O. Review of Current Disease Threats for Cultivated Penaeid Shrimp in Asia. Aquaculture 2016, 452, 69–87. [Google Scholar] [CrossRef]
- Anderson, J.L.; Valderrama, D.; Jory, D.E. Goal 2019: Global Shrimp Production Review. Glob. Aquac. Alliance 2019. Available online: https://www.aquaculturealliance.org/advocate/goal-2019-globalshrimp-production-review/ (accessed on 14 July 2020).
- Servin Arce, K.; de Souza Valente, C.; do Vale Pereira, G.; Shapira, B.; Davies, S.J. Modulation of the Gut Microbiota of Pacific White Shrimp (Penaeus vannamei Boone, 1931) by Dietary Inclusion of a Functional Yeast Cell Wall-Based Additive. Aquac. Nutr. 2021, 27, 1114–1127. [Google Scholar] [CrossRef]
- Hoa, N.D.; Wouters, R.; Wille, M.; Thanh, V.; Dong, T.K.; Van Hao, N.; Sorgeloos, P. A Fresh-Food Maturation Diet with an Adequate HUFA Composition for Broodstock Nutrition Studies in Black Tiger Shrimp Penaeus monodon (Fabricius, 1798). Aquaculture 2009, 297, 116–121. [Google Scholar] [CrossRef]
- Wouters, R.; Molina, C.; Lavens, P.; Calderón, J. Lipid Composition and Vitamin Content of Wild Female Litopenaeus vannamei in Different Stages of Sexual Maturation. Aquaculture 2001, 198, 307–323. [Google Scholar] [CrossRef] [Green Version]
- Vijayan, K.K.; Raj, V.S.; Balasubramanian, C.P.; Alavandi, S.V.; Sekhar, V.T.; Santiago, T.C. Polychaete Worms-a Vector for White Spot Syndrome Virus (WSSV). Dis. Aquat. Org. 2005, 63, 107–111. [Google Scholar] [CrossRef] [Green Version]
- Wouters, R.; Piguave, X.; Bastidas, L.; Calderón, J.; Sorgeloos, P. Ovarian Maturation and Haemolymphatic Vitellogenin Concentration of Pacific White Shrimp Litopenaeus vannamei (Boone) Fed Increasing Levels of Total Dietary Lipids and HUFA. Aquac. Res. 2001, 32, 573–582. [Google Scholar] [CrossRef] [Green Version]
- Xu, H.; Zhang, Y.; Luo, K.; Meng, X.; Luan, S.; Cao, B.; Chen, B.; Liang, M.; Kong, J. Arachidonic Acid in Diets for Early Maturation Stages Enhances the Final Reproductive Performances of Pacific White Shrimp (Litopenaeus vannamei). Aquaculture 2017, 479, 556–563. [Google Scholar] [CrossRef]
- Maneii, K.; Oujifard, A.; Ghasemi, A.; Mozanzadeh, M.T. Reproductive Performance and Vitellogenin mRNA Transcript Abundance in the Hepatopancreas of Female Litopenaeus vannamei Fed Diets with Different Soy Lecithin Content. Anim. Reprod. Sci. 2019, 211, 106228. [Google Scholar] [CrossRef]
- Mirheydari, S.-M.; Matinfar, A.; Emadi, H. Relation between Reproductive and Biologic Performance of Litopenaeus vannamei Females Broodstock Fed Different Dietary Highly Unsaturated Fatty Acid (HUFA) Levels. World Appl. Sci. J. 2014, 32, 123–132. [Google Scholar]
- Arshadi, A.; Gharaei, A.; Mirdar Harijani, J. Effect of Dietary Vitamin E on Reproductive Performance and Vitellogenin Gene Expression in Broodstock of Litopenaeus vannamei. Iran. J. Fish. Sci. 2020, 19, 2475–2492. [Google Scholar]
- Du, S.; Hu, C.; Shen, Q. Effect of Dietary Ascorbic Acid Levels on Reproductive Performance of Shrimp, Litopenaeus vannamei (Boone), Broodstock. J. Shellfish. Res. 2004, 23, 251–255. [Google Scholar]
- Maggioni, D.S.; Andreatta, E.R.; Hermes, E.M.; Barracco, M.A. Evaluation of Some Hemato-Immunological Parameters in Female Shrimp Litopenaeus vannamei Submitted to Unilateral Eyestalk Ablation in Association with a Diet Supplemented with Superdoses of Ascorbic Acid as a Form of Immunostimulation. Aquaculture 2004, 241, 501–515. [Google Scholar] [CrossRef]
- Corral-Rosales, D.C.; Cruz-Suárez, L.E.; Ricque-Marie, D.; Rodríguez-Jaramillo, C.; Palacios, E. Modulation of Reproductive Exhaustion Using Ulva Clathrata in Pacific White Shrimp Litopenaeus vannamei (Boone, 1931) Broodstock during Commercial Maturation. Aquac. Res. 2018, 49, 3711–3722. [Google Scholar] [CrossRef]
- Cavalli, R.O.; Menschaert, G.; Lavens, P.; Sorgeloos, P. Maturation Performance, Offspring Quality and Lipidcomposition of Macrobrachium rosenbergii Females fed Increasing Levels of Dietary Phospholipids. Aquac. Int. 2000, 8, 41–58. [Google Scholar] [CrossRef]
- Wang, L.; Zuo, D.; Lv, W.; Li, J.; Wang, Q.; Zhao, Y. Effects of Dietary Soybean Lecithin on Gonadal Development and Vitellogenin mRNA Expression in the Female Redclaw Crayfish Cherax quadricarinatus (von Martens) at First Maturation. Aquac. Res. 2013, 44, 1167–1176. [Google Scholar] [CrossRef]
- Sui, L.Y.; Wu, X.G.; Wille, M.; Cheng, Y.X.; Sorgeloos, P. Effect of Dietary Soybean Lecithin on Reproductive Performance of Chinese Mitten Crab Eriocheir sinensis (H. Milne-Edwards) Broodstock. Aquac. Int. 2009, 17, 45–56. [Google Scholar] [CrossRef]
- Li, X.; Wang, J.; Han, T.; Hu, S.; Jiang, Y.; Wang, C. Effect of Dietary Phospholipids Levels and Sources on Growth Performance, Fatty Acid Composition of the Juvenile Swimming Crab, Portunus trituberculatus. Aquaculture 2014, 430, 166–172. [Google Scholar] [CrossRef]
- Lin, Z.; Qi, C.; Han, F.; Chen, X.; Qin, C.; Wang, C.; Wang, X.; Qin, J.; Chen, L. Selecting Suitable Phospholipid Source for Female Eriocheir sinensis in Pre-Reproductive Phase. Aquaculture 2020, 528, 735610. [Google Scholar] [CrossRef]
- Liang, X.; Luo, X.; Lin, H.; Han, F.; Qin, J.G.; Chen, L.; Xu, C.; Li, E. Effects and Mechanism of Different Phospholipid Diets on Ovary Development in Female Broodstock Pacific White Shrimp, Litopenaeus vannamei. Front. Nutr. 2022, 9, 830934. [Google Scholar] [CrossRef] [PubMed]
- Arshadi, A.; Yavari, V.; Oujifard, A.; Mousavi, S.M.; Gisbert, E.; Mozanzadeh, M.T. Dietary Nucleotide Mixture Effects on Reproductive and Performance, Ovary Fatty Acid Profile and Biochemical Parameters of Female Pacific Shrimp Litopenaeus vannamei. Aquac. Nutr. 2018, 24, 515–523. [Google Scholar] [CrossRef] [Green Version]
- Ashida, M. Purification and Characterization of Pre-Phenoloxidase from Hemolymph of the Silkworm Bombyx Mori. Arch. Biochem. Biophys. 1971, 144, 749–762. [Google Scholar] [CrossRef]
- Hultmark, D.; Steiner, H.; Rasmuson, T.; Boman, H.G. Insect Immunity. Purification and Properties of Three Inducible Bactericidal Proteins from Hemolymph of Immunized Pupae of Hyalophora Cecropia. Eur. J. Biochem. 1980, 106, 7–16. [Google Scholar] [CrossRef] [PubMed]
- Pfaffl, M.W.; Tichopad, A.; Prgomet, C.; Neuvians, T.P. Determination of Stable Housekeeping Genes, Differentially Regulated Target Genes and Sample Integrity: BestKeeper–Excel-Based Tool Using Pair-Wise Correlations. Biotechnol. Lett. 2004, 26, 509–515. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Shan, L.; Li, Y.; Zheng, S.; Wei, Y.; Shang, Y. Analysis of the Bacterial Floral Structure and Diversity of Xuanwei Ham by 16S rDNA Sequencing. J. Food Saf. 2020, 40, e12800. [Google Scholar] [CrossRef]
- Mori, H.; Maruyama, F.; Kato, H.; Toyoda, A.; Dozono, A.; Ohtsubo, Y.; Nagata, Y.; Fujiyama, A.; Tsuda, M.; Kurokawa, K.E.N. Design and Experimental Application of a Novel Non-Degenerate Universal Primer Set That Amplifies Prokaryotic 16S rRNA Genes with a Low Possibility to Amplify Eukaryotic rRNA Genes. DNA Res. 2014, 21, 217–227. [Google Scholar] [CrossRef]
- Wang, J.T.; Han, T.; Li, X.Y.; Hu, S.X.; Jiang, Y.D.; Wang, C.L. Effects of Dietary Phosphatidylcholine (PC) Levels on the Growth, Molt Performance and Fatty Acid Composition of Juvenile Swimming Crab, Portunus trituberculatus. Anim. Feed. Sci. Technol. 2016, 216, 225–233. [Google Scholar] [CrossRef]
- Feng, S.; Cai, Z.; Zuo, R.; Mai, K.; Ai, Q. Effects of Dietary Phospholipids on Growth Performance and Expression of Key Genes Involved in Phosphatidylcholine Metabolism in Larval and Juvenile Large Yellow Croaker, Larimichthys Crocea. Aquaculture 2017, 469, 59–66. [Google Scholar] [CrossRef]
- Lin, Z.; Han, F.; Lu, J.; Guo, J.; Qi, C.; Wang, C.; Xiao, S.; Bu, X.; Wang, X.; Qin, J.; et al. Influence of Dietary Phospholipid on Growth Performance, Body Composition, Antioxidant Capacity and Lipid Metabolism of Chinese Mitten Crab, Eriocheir sinensis. Aquaculture 2020, 516, 734653. [Google Scholar] [CrossRef]
- Xu, X.; Ji, H.; Belghit, I.; Liland, N.S.; Wu, W.; Li, X. Effects of Black Soldier Fly Oil Rich in n-3 HUFA on Growth Performance, Metabolism and Health Response of Juvenile Mirror Carp (Cyprinus carpio Var. Specularis). Aquaculture 2021, 533, 736144. [Google Scholar] [CrossRef]
- An, W.; He, H.; Dong, X.; Tan, B.; Yang, Q.; Chi, S.; Zhang, S.; Liu, H.; Yang, Y. Regulation of Growth, Fatty Acid Profiles, Hematological Characteristics and Hepatopancreatic Histology by Different Dietary n-3 Highly Unsaturated Fatty Acids Levels in the First Stages of Juvenile Pacific White Shrimp (Litopenaeus vannamei). Aquac. Rep. 2020, 17, 100321. [Google Scholar] [CrossRef]
- Kim, M.G.; Yang, I.; Lee, H.S.; Lee, J.Y.; Kim, K. Lipid-Modifying Effects of Krill Oil vs Fish Oil: A Network Meta-Analysis. Nutr. Rev. 2020, 78, 699–708. [Google Scholar] [CrossRef] [PubMed]
- Azarm, H.M.; Kenari, A.A.; Hedayati, M. Effect of Dietary Phospholipid Sources and Levels on Growth Performance, Enzymes Activity, Cholecystokinin and Lipoprotein Fractions of Rainbow Trout (Oncorhynchus mykiss) Fry. Aquac. Res. 2013, 44, 634–644. [Google Scholar] [CrossRef]
- Chen, Y.-Y.; Sim, S.S.; Chiew, S.L.; Yeh, S.-T.; Liou, C.-H.; Chen, J.-C. Dietary Administration of a Gracilaria Tenuistipitata Extract Produces Protective Immunity of White Shrimp Litopenaeus vannamei in Response to Ammonia Stress. Aquaculture 2012, 370, 26–31. [Google Scholar] [CrossRef]
- Hamza, N.; Mhetli, M.; Khemis, I.B.; Cahu, C.; Kestemont, P. Effect of Dietary Phospholipid Levels on Performance, Enzyme Activities and Fatty Acid Composition of Pikeperch (Sander lucioperca) Larvae. Aquaculture 2008, 275, 274–282. [Google Scholar] [CrossRef] [Green Version]
- Zhao, J.; Ai, Q.; Mai, K.; Zuo, R.; Luo, Y. Effects of Dietary Phospholipids on Survival, Growth, Digestive Enzymes and Stress Resistance of Large Yellow Croaker, Larmichthys crocea Larvae. Aquaculture 2013, 410, 122–128. [Google Scholar] [CrossRef]
- Liu, B.; Zhao, M.; Xue, J.; Gu, Q.; Zhang, X.; Qin, S. Hydrogen Influences HDL-Associated Enzymes and Reduces Oxidized Phospholipids Levels in Rats Fed with a High-Fat Diet. Life Sci. 2021, 267, 118945. [Google Scholar] [CrossRef] [PubMed]
- Heink, A.E.; Parrish, A.N.; Thorgaard, G.H.; Carter, P.A. Oxidative Stress among SOD-1 Genotypes in Rainbow Trout (Oncorhynchus mykiss). Aquat. Toxicol. 2013, 144, 75–82. [Google Scholar] [CrossRef]
- Yu, P.; Liu, Z.; Wu, D.; Chen, M.; Lv, W.; Zhao, Y. Accumulation of Polystyrene Microplastics in Juvenile Eriocheir sinensis and Oxidative Stress Effects in the Liver. Aquat. Toxicol. 2018, 200, 28–36. [Google Scholar] [CrossRef]
- Song, D.; Shi, B.; Ding, L.; Jin, M.; Sun, P.; Jiao, L.; Zhou, Q. Regulation of Dietary Phospholipids on Growth Performance, Antioxidant Activities, Phospholipid Metabolism and Vitellogenesis in Prereproductive Phase of Female Swimming Crabs, Portunus trituberculatus. Aquaculture 2019, 511, 734230. [Google Scholar] [CrossRef]
- Jin, M.; Monroig, Ó.; Lu, Y.; Yuan, Y.; Li, Y.; Ding, L.; Tocher, D.R.; Zhou, Q. Dietary DHA/EPA Ratio Affected Tissue Fatty Acid Profiles, Antioxidant Capacity, Hematological Characteristics and Expression of Lipid-Related Genes but Not Growth in Juvenile Black Seabream (Acanthopagrus schlegelii). PLoS ONE 2017, 12, e0176216. [Google Scholar]
- Li, Y.; Gao, J.; Huang, S. Effects of Different Dietary Phospholipid Levels on Growth Performance, Fatty Acid Composition, PPAR Gene Expressions and Antioxidant Responses of Blunt Snout Bream Megalobrama amblycephala Fingerlings. Fish Physiol. Biochem. 2015, 41, 423–436. [Google Scholar] [CrossRef]
- Gao, J.; Koshio, S.; Wang, W.; Li, Y.; Huang, S.; Cao, X. Effects of Dietary Phospholipid Levels on Growth Performance, Fatty Acid Composition and Antioxidant Responses of Dojo Loach Misgurnus anguillicaudatus Larvae. Aquaculture 2014, 426, 304–309. [Google Scholar] [CrossRef]
- Cai, Z.; Feng, S.; Xiang, X.; Mai, K.; Ai, Q. Effects of Dietary Phospholipid on Lipase Activity, Antioxidant Capacity and Lipid Metabolism-Related Gene Expression in Large Yellow Croaker Larvae (Larimichthys crocea). Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2016, 201, 46–52. [Google Scholar] [CrossRef] [PubMed]
- Che, H.; Li, H.; Song, L.; Dong, X.; Yang, X.; Zhang, T.; Wang, Y.; Xie, W. Orally Administered DHA-Enriched Phospholipids and DHA-Enriched Triglyceride Relieve Oxidative Stress, Improve Intestinal Barrier, Modulate Inflammatory Cytokine and Gut Microbiota, and Meliorate Inflammatory Responses in the Brain in Dextran Sodium Sulfa. Mol. Nutr. Food Res. 2021, 65, 2000986. [Google Scholar] [CrossRef]
- Duan, Y.; Wang, Y.; Liu, Q.; Zhang, J.; Xiong, D. Changes in the Intestine Barrier Function of Litopenaeus vannamei in Response to PH Stress. Fish Shellfish. Immunol. 2019, 88, 142–149. [Google Scholar] [CrossRef] [PubMed]
- Loker, E.S.; Adema, C.M.; Zhang, S.; Kepler, T.B. Invertebrate Immune Systems–Not Homogeneous, Not Simple, Not Well Understood. Immunol. Rev. 2004, 198, 10–24. [Google Scholar] [CrossRef] [PubMed]
- Tong, R.; Pan, L.; Zhang, X.; Li, Y. Neuroendocrine-Immune Regulation Mechanism in Crustaceans: A Review. Rev. Aquac. 2022, 14, 378–398. [Google Scholar] [CrossRef]
- Jiang, H.-F.; Chen, C.; Jiang, X.-Y.; Shen, J.-L.; Ling, F.; Li, P.-F.; Wang, G.-X. Luteolin in Lonicera Japonica Inhibits the Proliferation of White Spot Syndrome Virus in the Crayfish Procambarus clarkii. Aquaculture 2022, 550, 737852. [Google Scholar] [CrossRef]
- Leu, J.-H.; Lin, S.-J.; Huang, J.-Y.; Chen, T.-C.; Lo, C.-F. A Model for Apoptotic Interaction between White Spot Syndrome Virus and Shrimp. Fish Shellfish. Immunol. 2013, 34, 1011–1017. [Google Scholar] [CrossRef] [PubMed]
- Perazzolo, L.M.; Barracco, M.A. The Prophenoloxidase Activating System of the Shrimp Penaeus paulensis and Associated Factors. Dev. Comp. Immunol. 1997, 21, 385–395. [Google Scholar] [CrossRef]
- Zhang, S.-C.; Xu, N. Advance in Study of Animal Lysozymes. Period. Ocean Univ. China 2014, 44, 46–51. [Google Scholar]
- Song, F.; Chen, G.-L.; Lu, K.-C.; Fan, J.-Q.; Yan, M.-T.; He, H.-H.; Lian, Y.-Y.; Zhang, C.-Z.; Chen, Y.-H. Identification and Functional Characterization of a C-Type Lectin Gene from Litopenaeus vannamei That Is Associated with ER-Stress Response. Fish Shellfish. Immunol. 2019, 93, 977–985. [Google Scholar] [CrossRef]
- Han, S.; Liu, Y.; Zhou, Z.; He, S.; Cao, Y.; Shi, P.; Yao, B.; Ringø, E. Analysis of Bacterial Diversity in the Intestine of Grass Carp (Ctenopharyngodon idellus) Based on 16S rDNA Gene Sequences. Aquac. Res. 2010, 42, 47–56. [Google Scholar] [CrossRef] [Green Version]
- Choct, M. Managing Gut Health through Nutrition. Br. Poult. Sci. 2009, 50, 9–15. [Google Scholar] [CrossRef]
- Miao, S.; Wan, W.; Hu, J.; Chang, E.; Zhou, Z.; Zhou, Y.; Sun, L. Dietary Arachidonic Acid Affects the Innate Immunity, Antioxidant Capacities, Intestinal Health and Microbiota in Chinese Mitten Crab (Eriocheir sinensis). Aquaculture 2022, 548, 737635. [Google Scholar] [CrossRef]
- Landsman, A.; St-Pierre, B.; Rosales-Leija, M.; Brown, M.; Gibbons, W. Impact of Aquaculture Practices on Intestinal Bacterial Profiles of Pacific Whiteleg Shrimp Litopenaeus vannamei. Microorganisms 2019, 7, 93. [Google Scholar] [CrossRef] [Green Version]
- Zhou, L.; Chen, C.; Xie, J.; Xu, C.; Zhao, Q.; Qin, J.G.; Chen, L.; Li, E. Intestinal Bacterial Signatures of the “Cotton Shrimp-like” Disease Explain the Change of Growth Performance and Immune Responses in Pacific White Shrimp (Litopenaeus vannamei). Fish Shellfish. Immunol. 2019, 92, 629–636. [Google Scholar] [CrossRef]
- Kang, C.; Wang, B.; Kaliannan, K.; Wang, X.; Lang, H.; Hui, S.; Huang, L.; Zhang, Y.; Zhou, M.; Chen, M. Gut Microbiota Mediates the Protective Effects of Dietary Capsaicin against Chronic Low-Grade Inflammation and Associated Obesity Induced by High-Fat Diet. MBio 2017, 8, e00470-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Liu, L.; Cao, Z.; Li, W.; Li, H.; Lu, C.; Yang, X.; Liu, Y. Gut Microbiota as an “Invisible Organ” That Modulates the Function of Drugs. Biomed. Pharmacother. 2020, 121, 109653. [Google Scholar] [CrossRef] [PubMed]
- Koyama, Y.; Brenner, D.A. Liver Inflammation and Fibrosis. J. Clin. Investig. 2017, 127, 55–64. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, G.; Wang, H.; Cheng, Y.; Liu, H.; Jiang, Z.; Li, P.; Wang, Y. Effects of Different Dissolved Organic Matter on Microbial Communities and Arsenic Mobilization in Aquifers. J. Hazard. Mater. 2021, 411, 125146. [Google Scholar] [CrossRef] [PubMed]
- Orino, K.; Lehman, L.; Tsuji, Y.; Ayaki, H.; Torti, S.V.; Torti, F.M. Ferritin and the Response to Oxidative Stress. Biochem. J. 2001, 357, 241–247. [Google Scholar] [CrossRef]
- Torti, F.M.; Torti, S.V. Regulation of Ferritin Genes and Protein. Blood J. Am. Soc. Hematol. 2002, 99, 3505–3516. [Google Scholar] [CrossRef] [Green Version]
- Pongsomboon, S.; Tang, S.; Boonda, S.; Aoki, T.; Hirono, I.; Yasuike, M.; Tassanakajon, A. Differentially Expressed Genes in Penaeus monodon Hemocytes Following Infection with Yellow Head Virus. BMB Rep. 2008, 41, 670–677. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, G.; Zhang, C.; Wang, Y.; Guo, C.; Sang, F.; Wang, C. Identification and Characterization of a Ferritin Gene Involved in the Immune Defense Response of Scallop Chlamys farreri. Fish Shellfish. Immunol. 2016, 55, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Bäckhed, F.; Ding, H.; Wang, T.; Hooper, L.V.; Gou, Y.K.; Nagy, A.; Semenkovich, C.F.; Gordon, J.I. The Gut Microbiota as an Environmental Factor That Regulates Fat Storage. Proc. Natl. Acad. Sci. USA 2004, 101, 15718–15723. [Google Scholar] [CrossRef] [Green Version]
- Deng, Y.; Jiang, Y.-H.; Yang, Y.; He, Z.; Luo, F.; Zhou, J. Molecular Ecological Network Analyses. BMC Bioinform. 2012, 13, 113. [Google Scholar] [CrossRef] [Green Version]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredients | Experimental Diets | |||
|---|---|---|---|---|
| Ctrl | SL | EL | KO | |
| Fish meal | 200 | 200 | 200 | 200 |
| Casein | 320 | 320 | 320 | 320 |
| Gelatin | 80 | 80 | 80 | 80 |
| Corn starch | 150 | 150 | 150 | 150 |
| Fish oil | 10 | 10 | 10 | 10 |
| Soybean lecithin | 0 | 40 | 0 | 0 |
| Egg yolk lecithin | 0 | 0 | 40 | 0 |
| Krill oil | 0 | 0 | 0 | 40 |
| Cholesterol | 5 | 5 | 5 | 5 |
| Palm oil | 80 | 40 | 40 | 40 |
| Butylated hydroxytoluene | 1 | 1 | 1 | 1 |
| Anhydrous calcium carbonate | 4 | 4 | 4 | 4 |
| Calcium lactate pentahydrate | 4 | 4 | 4 | 4 |
| Choline chloride | 5 | 5 | 5 | 5 |
| Inositol | 0.25 | 0.25 | 0.25 | 0.25 |
| Betaine | 20 | 20 | 20 | 20 |
| Vitamin premix 1 | 10 | 10 | 10 | 10 |
| Mineral premix 2 | 20 | 20 | 20 | 20 |
| Carboxymethyl cellulose | 20 | 20 | 20 | 20 |
| Cellulose | 70.75 | 70.75 | 70.75 | 70.75 |
| Total | 1000 | 1000 | 1000 | 1000 |
| Analyzed proximate composition (%) | ||||
| Moisture | 7.51 | 7.43 | 7.78 | 7.42 |
| Crude protein | 52.33 | 52.15 | 52.50 | 52.47 |
| Crude lipid | 14.28 | 14.40 | 14.05 | 14.22 |
| Ash | 8.31 | 8.30 | 8.28 | 8.31 |
| n-3 PUFAs | 495.31 a | 694.01 c | 568.93 b | 2335.27 d |
| n-6 PUFAs | 335.62 a | 1577.41 d | 577.74 c | 399.4 b |
| Gene | Primer Sequence | Tm (°C) | GC% | GenBank NO |
|---|---|---|---|---|
| β-actin | F: GCAGTCCAACCCGAGAGGAAG | 61.49 | 62.00 | XM_027364954 |
| R: GTGCATCGTCACCAGCGAA | 57.09 | 58.00 | ||
| caspase-1 | F: CGGGTAGGAAGCCCACATATCAA | 59.78 | 52.00 | XM_027356206 |
| R: ACGGCGAAGTCAAAGCCAGAA | 57.59 | 52.00 | ||
| CTL3 | F: ATGTTCTTCGTGCTCCTGCTGT | 57.80 | 50.00 | XM_027356524 |
| R: GCAGTGGTCGTAAATGTTGTG | 55.63 | 48.00 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liang, X.; Luo, X.; Lin, H.; Han, F.; Qin, J.G.; Chen, L.; Xu, C.; Li, E. Growth, Health, and Gut Microbiota of Female Pacific White Shrimp, Litopenaeus vannamei Broodstock Fed Different Phospholipid Sources. Antioxidants 2022, 11, 1143. https://doi.org/10.3390/antiox11061143
Liang X, Luo X, Lin H, Han F, Qin JG, Chen L, Xu C, Li E. Growth, Health, and Gut Microbiota of Female Pacific White Shrimp, Litopenaeus vannamei Broodstock Fed Different Phospholipid Sources. Antioxidants. 2022; 11(6):1143. https://doi.org/10.3390/antiox11061143
Chicago/Turabian StyleLiang, Xiaolong, Xiaolong Luo, Hongxing Lin, Fenglu Han, Jian G. Qin, Liqiao Chen, Chang Xu, and Erchao Li. 2022. "Growth, Health, and Gut Microbiota of Female Pacific White Shrimp, Litopenaeus vannamei Broodstock Fed Different Phospholipid Sources" Antioxidants 11, no. 6: 1143. https://doi.org/10.3390/antiox11061143