
Anti-Inflammatory and Chondroprotective Effects Induced by Phenolic Compounds from Onion Waste Extracts in ATDC-5 Chondrogenic Cell Line
 , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Extracts Obtaining and Phenolic Compound Characterization
2.2. Onion Extracts Antioxidant Properties
2.3. Quantification of Total Phenolic and Flavonoid Content
2.4. Cell Culture and Viability
2.5. Analysis of Cell Cycle and DNA Content
2.6. Evaluation of Cell Apoptosis
2.7. Determination of Intracellular Levels of Reactive Oxygen Species (ROS)
2.8. LPS Treatment and Nitrite Assay
2.9. mRNA Expression Level Analyses
2.10. Immunofluorescence
2.11. Cell Permeability and Pharmacokinetic Properties of Phenolic Compounds
2.12. Statistical Analyses
3. Results and Discussion
3.1. Antioxidant Properties and Phenolic Content of Onion Extracts
3.2. Cellular and Molecular Effects of Onion Extracts in ATDC-5 Cells
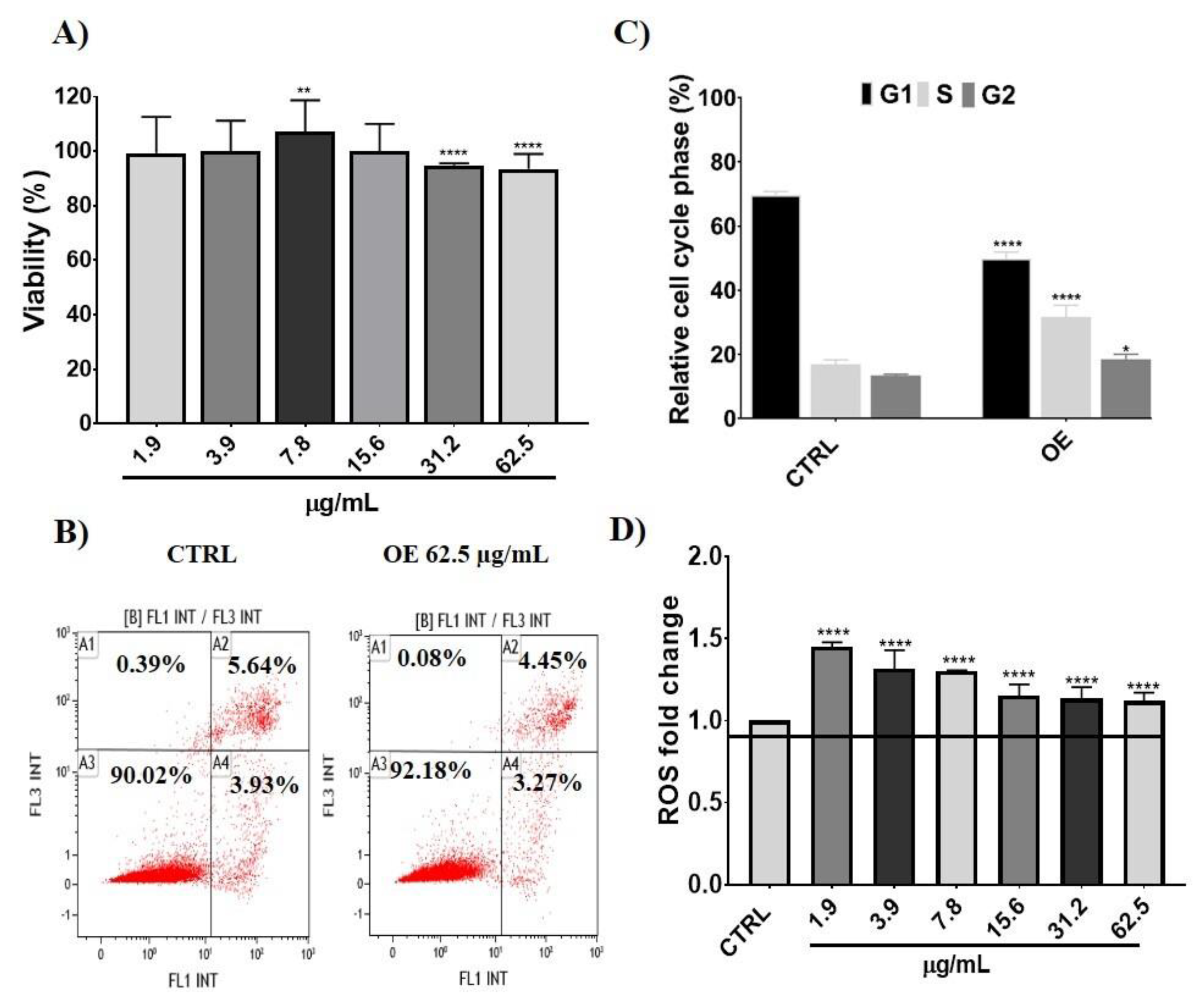
3.2.1. Cellular Effects and Intracellular ROS Levels after Onion Extracts Treatment
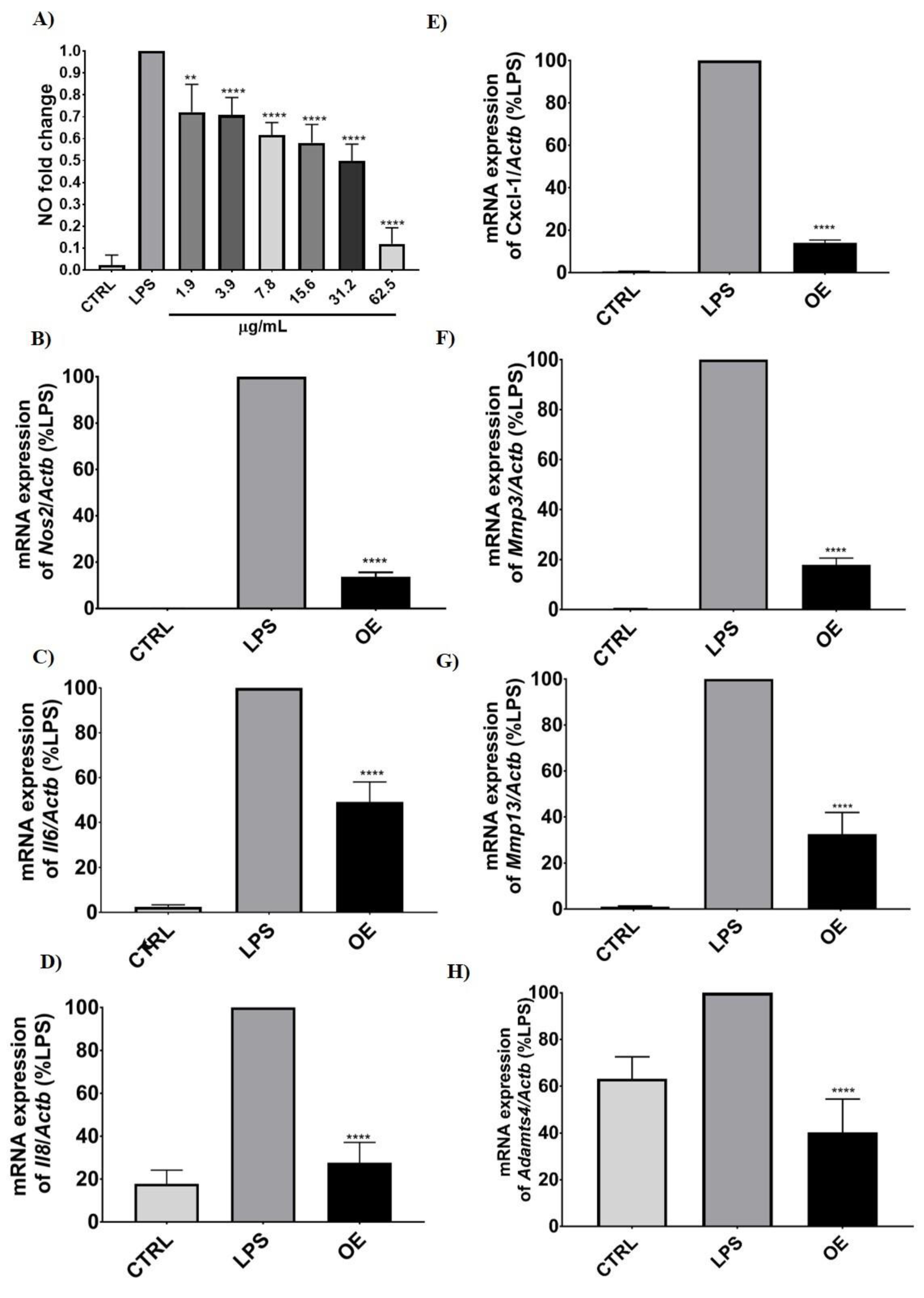
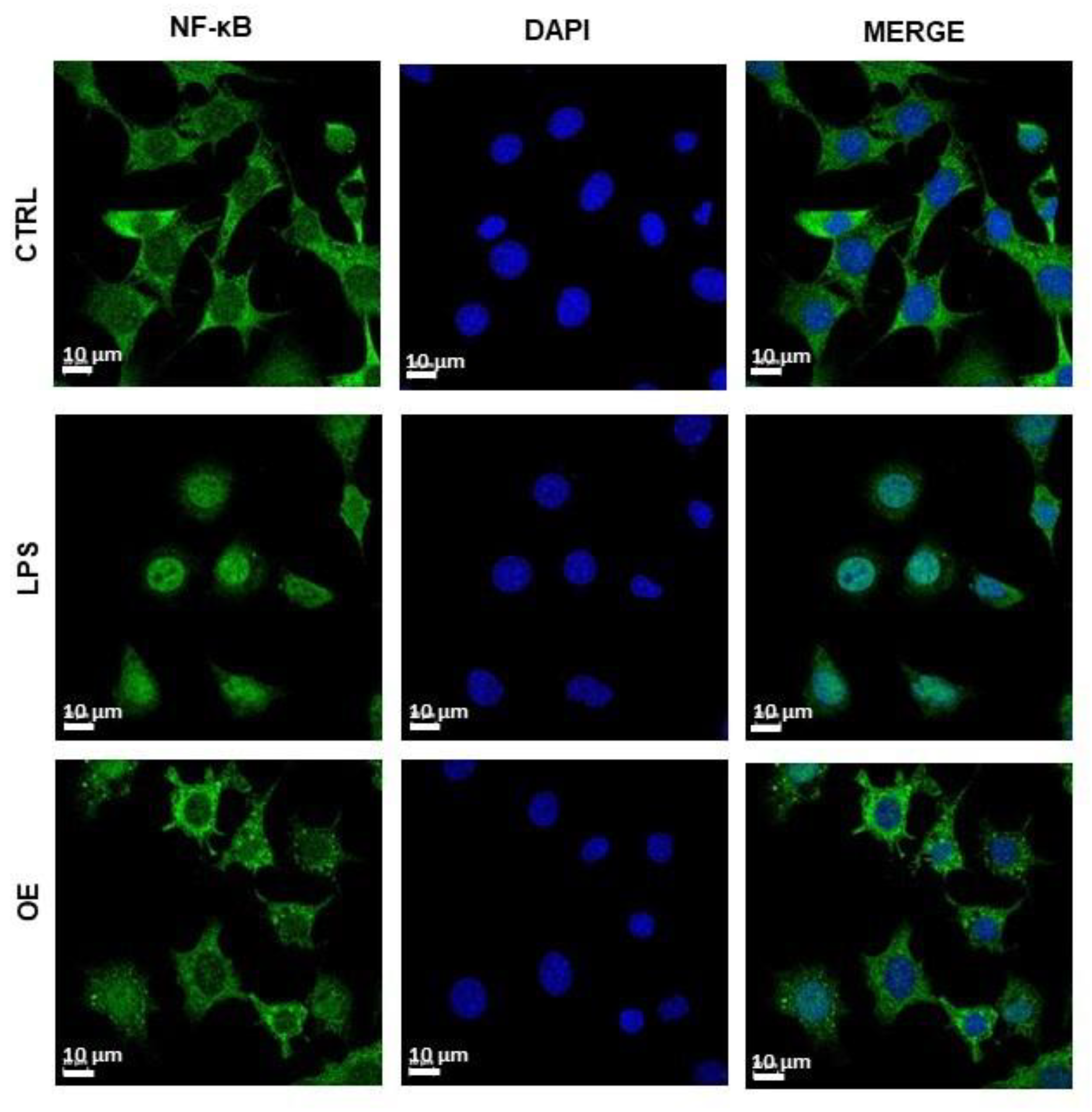
3.2.2. Molecular Changes after LPS and Onion Extracts Treatment
3.3. Membrane Permeability and Pharmacokinetic Properties of Phenolic Compounds
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hitzl, W.; Stamm, T.; Kloppenburg, M.; Ritter, M.; Gaisberger, M.; van der Zee-Neuen, A. Projected Number of Osteoarthritis Patients in Austria for the next Decades–Quantifying the Necessity of Treatment and Prevention Strategies in Europe. BMC Musculoskelet. Disord. 2022, 23, 1–9. [Google Scholar] [CrossRef]
- Turkiewicz, A.; Petersson, I.F.; Björk, J.; Hawker, G.; Dahlberg, L.E.; Lohmander, L.S.; Englund, M. Current and Future Impact of Osteoarthritis on Health Care: A Population-Based Study with Projections to Year 2032. Osteoarthr. Cartil. 2014, 22, 1826–1832. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hootman, J.M.; Helmick, C.G. Projections of US Prevalence of Arthritis and Associated Activity Limitations. Arthritis Rheum. 2006, 54, 226–229. [Google Scholar] [CrossRef] [PubMed]
- Van der Kraan, P.M.; Van den Berg, W.B. Chondrocyte Hypertrophy and Osteoarthritis: Role in Initiation and Progression of Cartilage Degeneration? Osteoarthr. Cartil. 2012, 20, 223–232. [Google Scholar] [CrossRef] [Green Version]
- Gómez, R.; Villalvilla, A.; Largo, R.; Gualillo, O.; Herrero-Beaumont, G. TLR4 Signalling in Osteoarthritis—Finding Targets for Candidate DMOADs. Nat. Rev. Rheumatol. 2014, 11, 159–170. [Google Scholar] [CrossRef] [PubMed]
- Goldring, M.B.; Otero, M.; Plumb, D.A.; Dragomir, C.; Favero, M.; El Hachem, K.; Hashimoto, K.; Roach, H.I.; Olivotto, E.; Borzì, R.M.; et al. Roles of inflammatory and anabolic cytokines in cartilage metabolism: Signals and multiple effectors converge upon mmp-13 regulation in osteoarthritis. Eur. Cell. Mater. 2011, 21, 202. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.T.; Yunus, M.H.M.; Ugusman, A.; Yazid, M.D. Natural Compounds Affecting Inflammatory Pathways of Osteoarthritis. Antioxidants 2022, 11, 1722. [Google Scholar] [CrossRef] [PubMed]
- Qi, W.; Chen, Y.; Sun, S.; Xu, X.; Zhan, J.; Yan, Z.; Shang, P.; Pan, X.; Liu, H. Inhibiting TLR4 Signaling by Linarin for Preventing Inflammatory Response in Osteoarthritis. Aging (Albany NY) 2021, 13, 5369–5382. [Google Scholar] [CrossRef]
- Scotece, M.; Conde, J.; Abella, V.; López, V.; Francisco, V.; Ruiz, C.; Campos, V.; Lago, F.; Gomez, R.; Pino, J.; et al. Oleocanthal Inhibits Catabolic and Inflammatory Mediators in LPS-Activated Human Primary Osteoarthritis (OA) Chondrocytes Through MAPKs/NF-ΚB Pathways. Cell. Physiol. Biochem. 2018, 49, 2414–2426. [Google Scholar] [CrossRef]
- Lim, H.; Park, H.; Kim, H.P. Effects of Flavonoids on Matrix Metalloproteinase-13 Expression of Interleukin-1β-Treated Articular Chondrocytes and Their Cellular Mechanisms: Inhibition of c-Fos/AP-1 and JAK/STAT Signaling Pathways. J. Pharmacol. Sci. 2011, 116, 221–231. [Google Scholar] [CrossRef]
- Imada, K.; Lin, N.; Liu, C.; Lu, A.; Chen, W.; Yano, M.; Sato, T.; Ito, A. Nobiletin, a Citrus Polymethoxy Flavonoid, Suppresses Gene Expression and Production of Aggrecanases-1 and -2 in Collagen-Induced Arthritic Mice. Biochem. Biophys. Res. Commun. 2008, 373, 181–185. [Google Scholar] [CrossRef] [PubMed]
- Wongwichai, T.; Teeyakasem, P.; Pruksakorn, D.; Kongtawelert, P.; Pothacharoen, P. Anthocyanins and Metabolites from Purple Rice Inhibit IL-1β-Induced Matrix Metalloproteinases Expression in Human Articular Chondrocytes through the NF-ΚB and ERK/MAPK Pathway. Biomed. Pharmacother. 2019, 112, 108610. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Gui, Z.; Zhou, Y.; Xia, L.; Lin, K.; Xu, Y. Quercetin Alleviates Rat Osteoarthritis by Inhibiting Inflammation and Apoptosis of Chondrocytes, Modulating Synovial Macrophages Polarization to M2 Macrophages. Free Radic. Biol. Med. 2019, 145, 146–160. [Google Scholar] [CrossRef]
- Liu, F.C.; Lu, J.W.; Chien, C.Y.; Huang, H.S.; Lee, C.C.; Lien, S.B.; Lin, L.C.; Chen, L.W.; Ho, Y.J.; Shen, M.C.; et al. Arthroprotective Effects of Cf-02 Sharing Structural Similarity with Quercetin. Int. J. Mol. Sci. 2018, 19, 1453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, M.J.S.; Bobrowski, P.; Shukla, M.; Gupta, K.; Haqqi, T.M. Chondroprotective Effects of a Proanthocyanidin Rich Amazonian Genonutrient Reflects Direct Inhibition of Matrix Metalloproteinases and Upregulation of IGF-1 Production by Human Chondrocytes. J. Inflamm. 2007, 4, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Z.; Liu, Y.; Lu, Y.; Hou, M.; Shen, X.; Yang, H.; Shi, Q.; Zhang, Y.; He, F.; Zhu, X. Gingko Biloba-Inspired Lactone Prevents Osteoarthritis by Activating the AMPK-SIRT1 Signaling Pathway. Arthritis Res. Ther. 2022, 24, 1–17. [Google Scholar] [CrossRef]
- Yu, W.; Zhu, Y.; Li, H.; He, Y. Injectable Quercetin-Loaded Hydrogel with Cartilage-Protection and Immunomodulatory Properties for Articular Cartilage Repair. ACS Appl. Bio Mater. 2020, 3, 761–771. [Google Scholar] [CrossRef]
- Davidson, R.K.; Green, J.; Gardner, S.; Bao, Y.; Cassidy, A.; Clark, I.M. Identifying Chondroprotective Diet-Derived Bioactives and Investigating Their Synergism. Sci. Rep. 2018, 8, 1–13. [Google Scholar] [CrossRef]
- Akdeniz, B.; Sumnu, G.; Sahin, S. Microencapsulation of Phenolic Compounds Extracted from Onion (Allium Cepa) Skin. J. Food Process. Preserv. 2018, 42, e13648. [Google Scholar] [CrossRef]
- Benito-román, Ó.; Blanco, B.; Sanz, M.T.; Beltrán, S. Subcritical Water Extraction of Phenolic Compounds from Onion Skin Wastes (Allium Cepa Cv. Horcal): Effect of Temperature and Solvent Properties. Antioxidants 2020, 9, 1233. [Google Scholar] [CrossRef]
- Campone, L.; Celano, R.; Piccinelli, A.L.; Pagano, I.; Carabetta, S.; Di Sanzo, R.; Russo, M.; Ibañez, E.; Cifuentes, A.; Rastrelli, L. Response Surface Methodology to Optimize Supercritical Carbon Dioxide/Co-Solvent Extraction of Brown Onion Skin by-Product as Source of Nutraceutical Compounds. Food Chem. 2018, 269, 495–502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Assefa, A.D.; Jeong, Y.J.; Kim, D.J.; Jeon, Y.A.; Lee, J.R.; Ko, H.C.; Baek, H.J.; Sung, J.S. Assessing Phenolic Content and Antioxidant Potential Diversity in Allium Plants Using Multivariate Data Analysis. Hortic. Environ. Biotechnol. 2018, 59, 759–773. [Google Scholar] [CrossRef]
- Wang, T.Y.; Li, Q.; Bi, K.S. Bioactive Flavonoids in Medicinal Plants: Structure, Activity and Biological Fate. Asian J. Pharm. Sci. 2018, 13, 12–23. [Google Scholar] [CrossRef] [PubMed]
- Marrelli, M.; Argentieri, M.P.; Alexa, E.; Meleleo, D.; Statti, G.; Avato, P.; Conforti, F.; Mallamaci, R. Antioxidant Activity and Protective Effect of the Outer Scales Hydroalcoholic Extract of Allium cepa L. Var. Tropea on Toxicity Damage Induced by Cadmium in Caco-2 Cells. Food Chem. Toxicol. 2022, 170, 113495. [Google Scholar] [CrossRef]
- Marrelli, M.; Russo, C.; Statti, G.; Argentieri, M.P.; Meleleo, D.; Mallamaci, R.; Avato, P.; Conforti, F. Phytochemical and Biological Characterization of Dry Outer Scales Extract from Tropea Red Onion (Allium cepa L. Var. Tropea)–A Promising Inhibitor of Pancreatic Lipase. Phytomedicine Plus 2022, 2, 100235. [Google Scholar] [CrossRef]
- Metrani, R.; Singh, J.; Acharya, P.; Jayaprakasha, G.K.; Patil, B.S. Comparative Metabolomics Profiling of Polyphenols, Nutrients and Antioxidant Activities of Two Red Onion (Allium cepa L.) Cultivars. Plants 2020, 9, 1077. [Google Scholar] [CrossRef]
- Hithamani, G.; Kizhakayil, D.; Srinivasan, K. Uptake of Phenolic Compounds from Plant Foods in Human Intestinal Caco-2 Cells. J. Biosci. 2017, 42, 603–611. [Google Scholar] [CrossRef]
- Paesa, M.; Nogueira, D.P.; Velderrain-Rodríguez, G.; Esparza, I.; Jiménez-Moreno, N.; Mendoza, G.; Osada, J.; Martin-Belloso, O.; Rodríguez-Yoldi, M.J.; Ancín-Azpilicueta, C. Valorization of Onion Waste by Obtaining Extracts Rich in Phenolic Compounds and Feasibility of Its Therapeutic Use on Colon Cancer. Antioxidants 2022, 11, 733. [Google Scholar] [CrossRef]
- Lee, S.U.; Lee, J.H.; Choi, S.H.; Lee, J.S.; Ohnisi-Kameyama, M.; Kozukue, N.; Levin, C.E.; Friedman, M. Flavonoid Content in Fresh, Home-Processed, and Light-Exposed Onions and in Dehydrated Commercial Onion Products. J. Agric. Food Chem. 2008, 56, 8541–8548. [Google Scholar] [CrossRef]
- Esparza, I.; Moler, J.A.; Arteta, M.; Jiménez-moreno, N.; Ancín-azpilicueta, C. Phenolic Composition of Grape Stems from Different Spanish Varieties and Vintages. Biomolecules 2021, 11, 1221. [Google Scholar] [CrossRef]
- Quero, J.; Mármol, I.; Cerrada, E.; Rodríguez-Yoldi, M.J. Insight into the Potential Application of Polyphenol-Rich Dietary Intervention in Degenerative Disease Management. Food Funct. 2020, 11, 2805–2825. [Google Scholar] [CrossRef] [PubMed]
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventós, R.M. [14] Analysis of Total Phenols and Other Oxidation Substrates and Antioxidants by Means of Folin-Ciocalteu Reagent. Methods Enzymol. 1999, 299, 152–178. [Google Scholar] [CrossRef]
- Chandra, S.; Khan, S.; Avula, B.; Lata, H.; Yang, M.H.; Elsohly, M.A.; Khan, I.A. Assessment of Total Phenolic and Flavonoid Content, Antioxidant Properties, and Yield of Aeroponically and Conventionally Grown Leafy Vegetables and Fruit Crops: A Comparative Study. Evid. Based. Complement. Alternat. Med. 2014, 2014, 253875. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sánchez-de-Diego, C.; Mármol, I.; Pérez, R.; Gascón, S.; Rodriguez-Yoldi, M.J.; Cerrada, E. The Anticancer Effect Related to Disturbances in Redox Balance on Caco-2 Cells Caused by an Alkynyl Gold(I) Complex. J. Inorg. Biochem. 2017, 166, 108–121. [Google Scholar] [CrossRef] [Green Version]
- Rahiman, S.S.F.; Morgan, M.; Gray, P.; Shaw, P.N.; Cabot, P.J. Dynorphin 1-17 and Its N-Terminal Biotransformation Fragments Modulate Lipopolysaccharide-Stimulated Nuclear Factor-Kappa B Nuclear Translocation, Interleukin-1beta and Tumor Necrosis Factor-Alpha in Differentiated THP-1 Cells. PLoS ONE 2016, 11, e0153005. [Google Scholar] [CrossRef] [Green Version]
- Velderrain-Rodríguez, G.R.; Quero, J.; Osada, J.; Martín-Belloso, O.; Rodríguez-Yoldi, M.J. Phenolic-Rich Extracts from Avocado Fruit Residues as Functional Food Ingredients with Antioxidant and Antiproliferative Properties. Biomolecules 2021, 11, 977. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Lozano, M.L.; Cesaro, A.; Mazor, M.; Esteve, E.; Berteina-Raboin, S.; Best, T.M.; Lespessailles, E.; Toumi, H. Emerging Natural-Product-Based Treatments for the Management of Osteoarthritis. Antioxidants 2021, 10, 265. [Google Scholar] [CrossRef]
- Sukhikh, S.; Babich, O.; Prosekov, A.; Patyukov, N.; Ivanova, S. Future of Chondroprotectors in the Treatment of Degenerative Processes of Connective Tissue. Pharmaceuticals 2020, 13, 220. [Google Scholar] [CrossRef]
- Luo, L.; Wei, Q.; Liu, L.; Lin, X.; Lin, C.; Zheng, L.; Zhao, J. Protocatechuic Acid Benefits Proliferation and Phenotypic Maintenance of Rabbit Articular Chondrocytes: An in Vitro Study. Exp. Ther. Med. 2015, 9, 1865–1870. [Google Scholar] [CrossRef] [Green Version]
- Roleira, F.M.F.; Tavares-Da-Silva, E.J.; Varela, C.L.; Costa, S.C.; Silva, T.; Garrido, J.; Borges, F. Plant Derived and Dietary Phenolic Antioxidants: Anticancer Properties. Food Chem. 2015, 183, 235–258. [Google Scholar] [CrossRef]
- León-González, A.J.; Auger, C.; Schini-Kerth, V.B. Pro-Oxidant Activity of Polyphenols and Its Implication on Cancer Chemoprevention and Chemotherapy. Biochem. Pharmacol. 2015, 98, 371–380. [Google Scholar] [CrossRef] [PubMed]
- Kabe, Y.; Ando, K.; Hirao, S.; Yoshida, M.; Handa, H. Redox Regulation of NF-KappaB Activation: Distinct Redox Regulation between the Cytoplasm and the Nucleus. Antioxid. Redox Signal. 2005, 7, 395–403. [Google Scholar] [CrossRef] [PubMed]
- Morgan, M.J.; Liu, Z.G. Crosstalk of Reactive Oxygen Species and NF-ΚB Signaling. Cell Res. 2011, 21, 103–115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wongwat, T.; Srihaphon, K.; Pitaksutheepong, C.; Boonyo, W.; Pitaksuteepong, T. Suppression of Inflammatory Mediators and Matrix Metalloproteinase (MMP)-13 by Morus Alba Stem Extract and Oxyresveratrol in RAW 264.7 cells and C28/I2 Human Chondrocytes. J. Tradit. Complement. Med. 2020, 10, 132–140. [Google Scholar] [CrossRef]
- Karabasz, A.; Bzowska, M.; Szczepanowicz, K. Biomedical Applications of Multifunctional Polymeric Nanocarriers: A Review of Current Literature. Int. J. Nanomed. 2020, 15, 8673–8696. [Google Scholar] [CrossRef] [PubMed]
- Guirguis, M.; Bhandari, C.; Li, J.; Eroy, M.; Prajapati, S.; Margolis, R.; Shrivastava, N.; Hoyt, K.; Hasan, T.; Obaid, G. Membrane Composition Is a Functional Determinant of NIR-Activable Liposomes in Orthotopic Head and Neck Cancer. Nanophotonics 2021, 10, 3169–3185. [Google Scholar] [CrossRef] [PubMed]
- Hardjono, S.; Siswandono, S.; Andayani, R. Evaluation of N-Benzoylthiourea Derivatives as Popssible Analgesic Agents by Predicting Their Hysicochemical and Pharmacokinetic Properties, Toxicity, and Analgesic Activity. Indones. J. Biotechnol. 2018, 22, 76. [Google Scholar] [CrossRef] [Green Version]
- Sur, V.P.; Sen, M.K.; Komrskova, K. In Silico Identification and Validation of Organic Triazole Based Ligands as Potential Inhibitory Drug Compounds of SARS-CoV-2 Main Protease. Molecules 2021, 26, 6199. [Google Scholar] [CrossRef]
- Zadorozhnii, P.V.; Kiselev, V.V.; Kharchenko, A.V. In Silico ADME Profiling of Salubrinal and Its Analogues. Future Pharmacol. 2022, 2, 160–197. [Google Scholar] [CrossRef]
- Al-Nour, M.Y.; Ibrahim, M.M.; Elsaman, T. Ellagic Acid, Kaempferol, and Quercetin from Acacia Nilotica: Promising Combined Drug With Multiple Mechanisms of Action. Curr. Pharmacol. Rep. 2019, 5, 255–280. [Google Scholar] [CrossRef]
- Zendulka, O.; Zahradníková, L.; Juřica, J.; Totušek, J. The Influence of Trans-Resveratrol and Quercetin on the Activity of CYP1A2 in Rat. Czech J. Food Sci. 2008, 26, S60–S64. [Google Scholar] [CrossRef] [Green Version]
- Lotha, R.; Sivasubramanian, A. Flavonoids nutraceuticals in prevention and treatment of cancer: A review. Asian J. Pharm. Clin. Res. 2018, 11, 42–47. [Google Scholar] [CrossRef] [Green Version]
- Androutsopoulos, V.; Wilsher, N.; Arroo, R.R.J.; Potter, G.A. Bioactivation of the Phytoestrogen Diosmetin by CYP1 Cytochromes P450. Cancer Lett. 2009, 274, 54–60. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Pharmacokinetic Properties (Property; Units) | Protocatechuic Acid | Ellagic Acid | Vanillic Acid | Quercetin | Quercetin 3-Glucoside | Kaempferol | Isorhamnetin |
|---|---|---|---|---|---|---|---|
| Passive cellular permeability (Absorption; log Papp in 10–6 cm/s) | 0.49 | 0.335 | 0.33 | −0.229 | 0.242 | 0.032 | −0.003 |
| Water solubility (Absorption; log mol/L) | 2.069 | −3.181 | −1.838 | −2.925 | −2.925 | −3.04 | −3 |
| Human intestinal absorption (Absorption; % Absorbed) | 71.174 | 86.684 | 78.152 | 77.207 | 47.999 | 74.29 | 76.014 |
| Skin permeability (Absorption; log Kp) | −2.727 | −2.735 | −2.726 | −2.735 | −2.735 | −2.735 | −2.735 |
| P-glycoprotein substrate (Absorption; Yes/No) | No | Yes | No | Yes | Yes | Yes | Yes |
| P-glycoprotein I inhibitor (Absorption; Yes/No) | No | No | No | No | No | No | No |
| P-glycoprotein II inhibitor (Absorption; Yes/No) | No | No | No | No | No | No | No |
| VDss (human) (Distribution; log L/kg) | −1.298 | 0.375 | −1.739 | 1.559 | 1.846 | 1.274 | 1.123 |
| Fraction unbound (human) (Distribution; Fu) | 0.648 | 0.083 | 0.518 | 0.206 | 0.228 | 0.178 | 0.091 |
| BBB permeability (Distribution; log BB) | 0.683 | −1.272 | −0.38 | −1.098 | −1.688 | −0.939 | −1.135 |
| CNS permeability (Distribution; log PS) | −3.305 | −3.533 | −2.628 | −3.065 | −4.093 | −2.228 | −3.188 |
| CYP2D6 substrate (Metabolism; Yes/No) | No | No | No | No | No | No | No |
| CYP3A4 substrate (Metabolism; Yes/No) | No | No | No | No | No | No | No |
| CYP1A2 inhibitor (Metabolism; Yes/No) | No | Yes | No | Yes | No | Yes | Yes |
| CYP2C19 inhibitor (Metabolism; Yes/No) | No | No | No | No | No | No | No |
| CYP2C9 inhibitor (Metabolism; Yes/No) | No | No | No | No | No | No | No |
| CYP2D6 inhibitor (Metabolism; Yes/No) | No | No | No | No | No | No | No |
| CYP3A4 inhibitor (Metabolism; Yes/No) | No | No | No | No | No | No | No |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Paesa, M.; Ancín-Azpilicueta, C.; Velderrain-Rodríguez, G.; Martin-Belloso, O.; Gualillo, O.; Osada, J.; Rodríguez-Yoldi, M.J.; Mendoza, G. Anti-Inflammatory and Chondroprotective Effects Induced by Phenolic Compounds from Onion Waste Extracts in ATDC-5 Chondrogenic Cell Line. Antioxidants 2022, 11, 2381. https://doi.org/10.3390/antiox11122381
Paesa M, Ancín-Azpilicueta C, Velderrain-Rodríguez G, Martin-Belloso O, Gualillo O, Osada J, Rodríguez-Yoldi MJ, Mendoza G. Anti-Inflammatory and Chondroprotective Effects Induced by Phenolic Compounds from Onion Waste Extracts in ATDC-5 Chondrogenic Cell Line. Antioxidants. 2022; 11(12):2381. https://doi.org/10.3390/antiox11122381
Chicago/Turabian StylePaesa, Mónica, Carmen Ancín-Azpilicueta, Gustavo Velderrain-Rodríguez, Olga Martin-Belloso, Oreste Gualillo, Jesús Osada, Maria Jesús Rodríguez-Yoldi, and Gracia Mendoza. 2022. "Anti-Inflammatory and Chondroprotective Effects Induced by Phenolic Compounds from Onion Waste Extracts in ATDC-5 Chondrogenic Cell Line" Antioxidants 11, no. 12: 2381. https://doi.org/10.3390/antiox11122381