Alteration of the Antioxidant Capacity and Gut Microbiota under High Levels of Molybdenum and Green Tea Polyphenols in Laying Hens

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Preparation of TP
2.3. Birds, Experimental Design, and Sample Collection
2.4. Determination of Metabolic Parameters and Antioxidant Enzyme Activity in Serum
2.5. Determination of Trace Element in Serum, Egg, and Tissues
2.6. DNA Extraction and Intestinal Microbiota Analysis
2.7. Statistical Analysis
3. Results
3.1. Production Performances
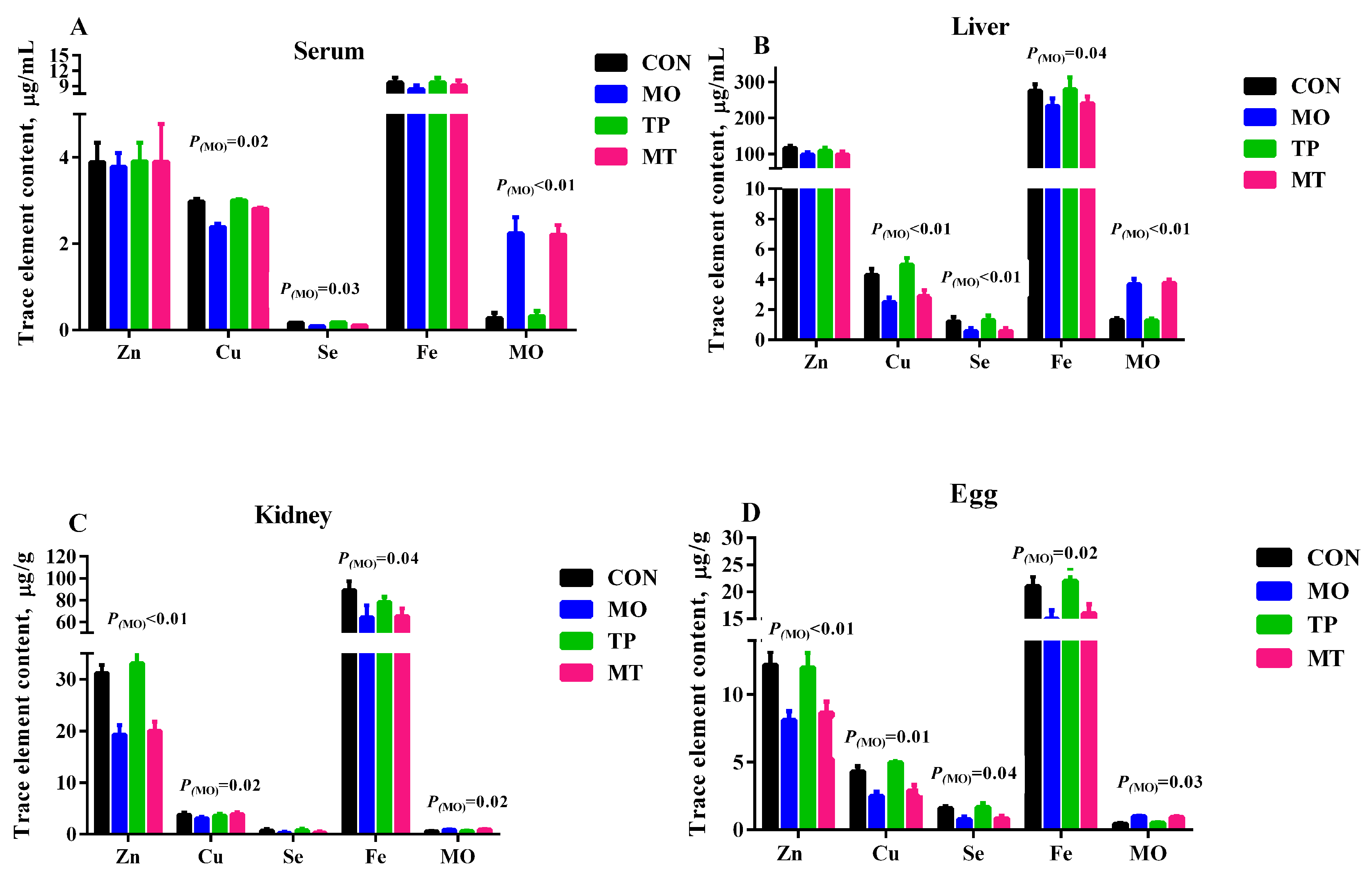
3.2. Trace Element Content in Tissues, Egg, and Serum
3.3. Serum Antioxidant Status Parameters
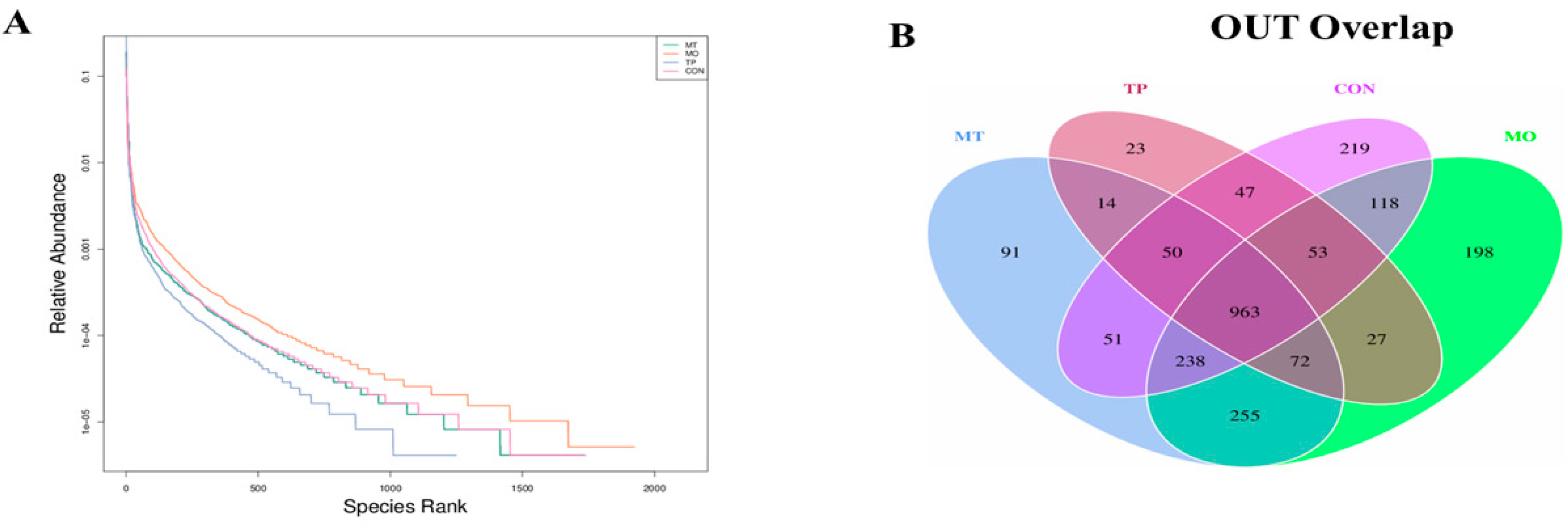
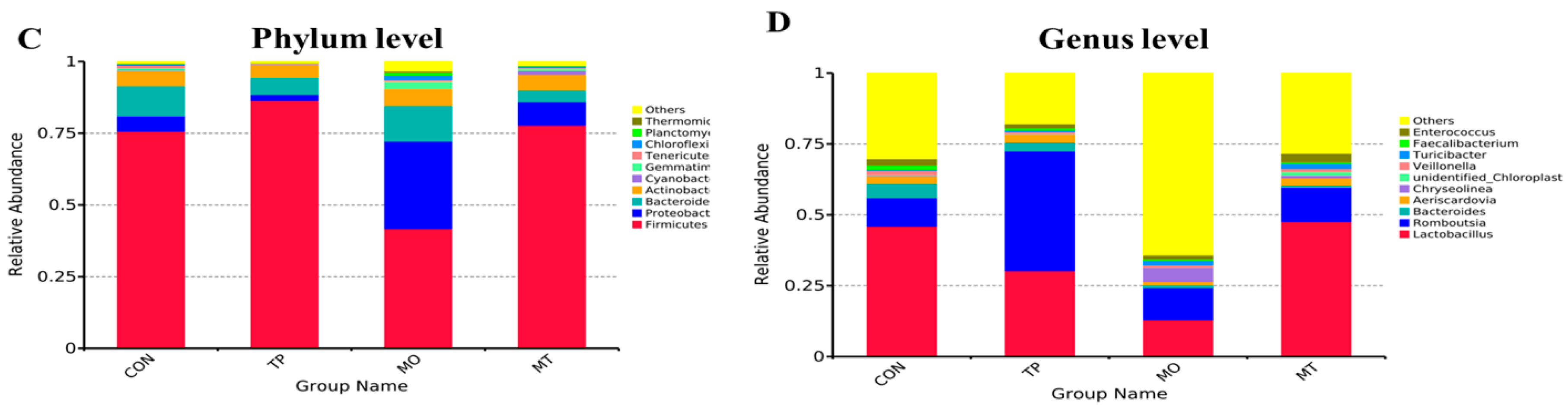
3.4. Microbiota Composition in the Layer Cecum
3.5. Alpha Diversity of Gut Microbiota in the Cecum-Fed High MO and TP
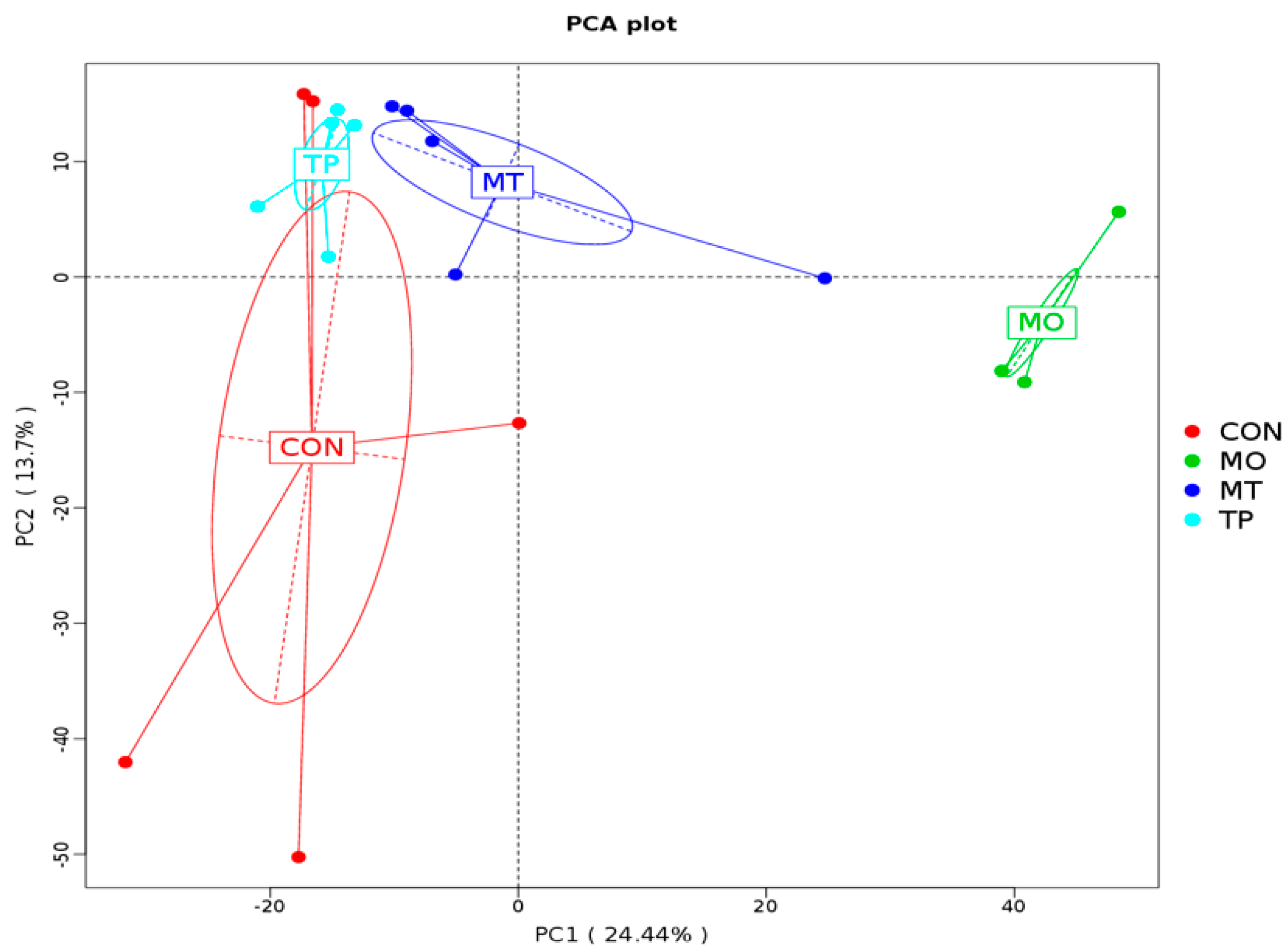
3.6. Beta Diversity of Gut Microbiota in the Cecum Fed High MO and TP
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviation
References
- Trumbo, P.; Yates, A.A.; Schlicker, S.; Poos, M. Dietary reference intakes: Vitamin A, vitamin K, arsenic, boron, chromium, copper, iodine, iron, manganese, molybdenum, nickel, silicon, vanadium, and zinc. J. Am. Diet. Assoc. 2001, 101, 294–301. [Google Scholar] [CrossRef]
- Abramovich, M.; Miller, A.; Yang, H.; Friel, J.K. Molybdenum content of Canadian and US infant formulas. Biol. Trace Elem. Res. 2011, 143, 844–853. [Google Scholar] [CrossRef] [PubMed]
- Barceloux, D.G. Molybdenum. J. Toxicol. Clin. Toxicol. 1999, 37, 231–237. [Google Scholar] [CrossRef] [PubMed]
- Aupperle, H.; Schoon, H.A.; Frank, A. Experimental copper deficiency, chromium deficiency and additional molybdenum supplementation in goats-pathological findings. Acta Vet. Scand. 2001, 42, 311–321. [Google Scholar] [CrossRef] [PubMed]
- Zhou, S.; Zhang, C.; Xiao, Q.; Zhuang, Y.; Gu, X.; Yang, F.; Xing, C.; Hu, G.; Cao, H. Effect of different levels of molybdenum on rumen micriobiota and trace elements changes in tissues from goats. Biol. Trace Elem. Res. 2016, 174, 85–92. [Google Scholar] [CrossRef] [PubMed]
- Liao, Z.Y.; Cao, H.B.; Dai, X.Y.; Xing, C.H.; Xu, X.W.; Nie, H.; Zhang, C.Y. Molybdenum and Cadmium exposure influences the concentration of trace elements in the digestive organs of Shaoxing duck (Anas platyrhyncha). Ecotoxicol. Environ. Saf. 2018, 164, 75–83. [Google Scholar] [CrossRef]
- Tallkvist, J.; Oskarsson, A. Molybdenum in Handbook on the Toxicology of Metals, 4th ed.; Academic Press: Oxford, UK, 2015; Volume II, pp. 1077–1089. [Google Scholar]
- Raisbeck, M.F.; Siemion, R.S.; Smith, M.A. Modest copper supplementation blocks molybdenosis in cattle. J. Vet. Diagn. Investig. 2006, 18, 566–572. [Google Scholar] [CrossRef]
- Yang, F.; Cui, H.; Xiao, J.; Peng, X.; Deng, J.; Zuo, Z. Increased apoptotic lymphocyte population in the spleen of young chickens fed on diets high in molybdenum. Biol. Trace Elem. Res. 2011, 140, 308–316. [Google Scholar] [CrossRef]
- Cao, H.B.; Xia, B.; Zhang, N.M.; Liao, Y.L.; Yang, Z.; Hu, G.L.; Zhang, C.Y. Changes of antioxidant function and the mRNA expression levels of apoptosis genes in duck ovaries caused by molybdenum or/and cadmium. Biol. Trace Elem. Res. 2016, 171, 410–418. [Google Scholar] [CrossRef]
- Xie, B.; Shi, H.; Chen, Q.; Ho, C.T. Antioxidant properties of fractions and polyphenol constituents from green, oolong and black teas. Proc. Natl. Sci. Counc. Repub. China B 1993, 17, 77. [Google Scholar]
- Frei, B.; Higdon, J.V. Antioxidant Activity of Tea Polyphenols in Vivo: Evidence from Animal Studies. J. Nutr. 2003, 133, 3275S–3284S. [Google Scholar] [CrossRef] [PubMed]
- Na, H.K.; Surh, Y.J. Modulation of Nrf2-Mediated antioxidant and detoxifying enzyme induction by the green tea polyphenol EGCG. Food Chem. Toxicol. 2008, 46, 1271–1278. [Google Scholar] [CrossRef] [PubMed]
- Minnelli, C.; Moretti, P.; Fulgenzi, G.; Mariani, P.; Laudadio, E.; Armeni, T.; Galeazzi, R.; Mobbili, G. A Poloxamer-407 modified liposome encapsulating Epigallocatechin-3-Gallate in the presence of magnesium: Characterization and protective effect against oxidative damage. Int. J. Pharm. 2018, 552, 225–234. [Google Scholar] [CrossRef] [PubMed]
- Ariana, M.; Samie, A.; Edriss, M.; Jahanian, R. Effects of powder and extract form of green tea and marigold, and α-Tocopheryl acetate on performance, egg quality and egg yolk cholesterol levels of laying hens in late phase of production. J. Med. Plants Res. 2011, 5, 2710–2716. [Google Scholar]
- Wang, J.; Yuan, Z.; Zhang, K.; Ding, X.; Bai, S.; Zeng, Q.; Peng, H.; Celi, P. Epigallocatechin-3-Gallate protected vanadium-induced eggshell depigmentation via P38MAPK-Nrf2/HO-1 signaling pathway in laying hens. Poult. Sci. 2018, 97, 3109–3118. [Google Scholar] [CrossRef]
- Yuan, Z.H.; Zhang, K.Y.; Ding, X.M.; Luo, Y.H.; Bai, S.P.; Zeng, Q.F.; Wang, J.P. Effect of tea polyphenols on production performance, egg quality, and hepatic antioxidant status of laying hens in Vanadium-Containing diets. Poult. Sci. 2016, 95, 1709–1717. [Google Scholar] [CrossRef]
- Viveros, A.; Chamorro, S.; Pizarro, M.; Arija, I.; Centeno, C.; Brenes, A. Effects of dietary Polyphenol-Rich grape products on intestinal microflora and gut morphology in broiler chicks. Poult. Sci. 2011, 90, 566–578. [Google Scholar] [CrossRef]
- Unno, T.; Sakuma, M.; Mitsuhashi, S. Effect of dietary supplementation of (−)-Epigallocatechin gallate on gut microbiota and biomarkers of colonic fermentation in rats. J. Nutr. Sci. Vitaminol. 2014, 60, 213–219. [Google Scholar] [CrossRef]
- Cheng, M.; Zhang, X.; Miao, Y.; Cao, J.; Wu, Z.; Weng, P. The modulatory effect of (-)-Epigallocatechin 3-O-(3-O-Methyl) gallate (EGCG3″Me) on intestinal microbiota of high fat diet-induced obesity mice model. Food Res. Int. 2017, 92, 9–16. [Google Scholar] [CrossRef]
- Zhang, X.; Zhang, H.; Ho, C.T.; Guo, X.J.; Wu, Z.F.; Weng, P.F.; Yan, M.D.; Cao, J.X. Metagenomics analysis of gut microbiota modulatory effect of green polyphenols by high fat diet-induced obesity mice model. J. Funct. Foods 2018, 46, 268–277. [Google Scholar] [CrossRef]
- NRC. Nutrient Requirements of Poultry, 9th ed.; National Academy Press: Washington, DC, USA, 1994. [Google Scholar]
- Choi, R.; Kim, M.-J.; Sohn, I.; Kim, S.; Kim, I.; Ryu, J.M.; Choi, H.J.; Kim, J.-M.; Lee, S.K.; Yu, J.; et al. Serum trace elements and their associations with breast cancer subgroups in Korean breast cancer patients. Nutrients 2019, 11, 37. [Google Scholar] [CrossRef] [PubMed]
- Fungwe, T.V.; Buddingh, F.; Demick, D.S.; Lox, C.D.; Yang, M.T.; Yang, S.P. The role of dietary molybdenum on estrous activity, fertility, reproduction, and molybdenum and copper enzyme activities of female rats. Nutr. Res. 1990, 10, 515–524. [Google Scholar] [CrossRef]
- Anke, M.K. Molybdenum. In Elements and Their Compounds in the Environment; Merian, E., Anke, M., Ihnat, M., Stoeppler, M., Eds.; Wiley-VCH Verlag: Weinheim, Germany, 2004; pp. 1007–1037. [Google Scholar]
- Novotny, J.A.; Turnlund, J.R. Molybdenum intake influences molybdenum kinetics in men. J. Nutr. 2007, 137, 37–42. [Google Scholar] [CrossRef] [PubMed]
- Xiao, J.; Cui, H.M.; Yang, F.; Peng, X.; Cui, Y. Effect of dietary high molybdenum on the cell cycle and apoptosis of kidney in broiler. Biol. Trace Elem. Res. 2011, 142, 523–531. [Google Scholar] [CrossRef]
- Xia, B.; Cao, H.; Luo, J.; Liu, P.; Guo, X.; Hu, G.; Zhang, C. The Co-Induced effects of molybdenum and cadmium on antioxidants and heat shock proteins in duck kidneys. Biol. Trace Elem. Res. 2015, 168, 261–268. [Google Scholar] [CrossRef]
- Cao, H.; Zhang, M.; Xia, B.; Xiong, J.; Zong, Y.; Hu, G.; Zhang, C. Effects of Molybdenum or/and Cadmium on mRNA expression levels of inflammatory cytokines and HSPs in duck spleens. Biol. Trace Elem. Res. 2016, 170, 237–244. [Google Scholar] [CrossRef]
- Beaulieu, M.; Costantini, D. Biomarkers of oxidative status: Missing tools in conservation physiology. Conserv. Physiol. 2014, 2, cou014. [Google Scholar] [CrossRef]
- Hencken, H. Chemical and physiological behavior of feed carotenoids and their effects on pigmentation. Poult. Sci. 1992, 71, 711–717. [Google Scholar] [CrossRef]
- Leeson, S.; Caston, L. Enrichment of eggs with lutein. Poult. Sci. 2004, 83, 1709–1712. [Google Scholar] [CrossRef]
- Karadas, F.; Grammenidis, E.; Surai, P.F.; Acamovic, T.; Sparks, N.H. Effects of carotenoids from lucerne, marigold and tomato on egg yolk pigmentation and carotenoid composition. Br. Poult. Sci. 2006, 47, 561–566. [Google Scholar] [CrossRef]
- Suksombat, W.; Samitayotin, S.; Lounglawan, P. Effects of conjugated linoleic acid supplementation in layer diet on fatty acid compositions of egg yolk and layer performances. Poult. Sci. 2006, 85, 1603–1609. [Google Scholar] [CrossRef] [PubMed]
- Palace, V.P.; Khaper, N.K.; Qin, Q.N.; Singal, P.K. Antioxidant potentials of vitamin A and carotenoids and their relevance to heart disease. Free Radic. Biol. Med. 1999, 26, 746–761. [Google Scholar] [CrossRef]
- Rajagopalan, K.V. Molybdenum: An essential trace element in human nutrition. Annu. Rev. Nutr. 1988, 8, 401–427. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Luo, J.; Zhang, C.; Cao, H.; Xia, B.; Hu, G. Alterations in antioxidant function and cell apoptosis in duck spleen exposed to molybdenum and/or cadmium. J. Vet. Sci. 2017, 18, 193–200. [Google Scholar] [CrossRef] [Green Version]
- Kehrer, J.P. Free radicals as mediators of tissue injury and disease. Crit. Rev. Toxicol. 1993, 23, 21–48. [Google Scholar] [CrossRef]
- Sagor, M.A.; Tabassum, N.; Potol, M.A.; Alam, M.A. Xanthine oxidase inhibitor, allopurinol, prevented oxidative stress, fibrosis, and myocardial damage in isoproterenol induced aged rats. Oxid. Med. Cell Longev. 2015, 2015, 478039. [Google Scholar] [CrossRef]
- Rio, D.E.; Stewart, A.J.; Pellegrini, N. A review of recent studies on malondialdehyde as toxic molecule and biological marker of oxidative stress. Nutr. Metab. Cardiovasc. Dis. 2005, 15, 316–328. [Google Scholar]
- Celi, P.; Gabai, G. Oxidant/antioxidant balance in animal nutrition and health: The role of protein oxidation. Front. Vet. Sci. 2015, 2, 48. [Google Scholar] [CrossRef]
- Celi, P.; Verlhac, V.; Calvo, E.P.; Schmeisser, J.; Kluenter, A.-M. Biomarkers of gastrointestinal functionality in animal nutrition and health. Anim. Feed Sci. Technol. 2019, 250, 9–31. [Google Scholar] [CrossRef]
- Markesbery, W.R.; Kryscio, R.J.; Lovell, M.A.; Morrow, J.D. Lipid peroxidation is an early event in the brain in amnestic mild cognitive impairment. Ann. Neurol. 2005, 58, 730–735. [Google Scholar] [CrossRef]
- Kiersztan, A.; Winiarska, K.; Drozak, J.; Przedlacka, M.; Wegrzynowicz, M.; Fraczyk, T.; Bryla, J. Differential effects of vanadium, tungsten and molybdenum on inhibition of glucose formation in renal tubules and hepatocytes of control and diabetic rabbits: Beneficial action of melatonin and N-acetylcysteine. Mol. Cell Biochem. 2004, 261, 9–21. [Google Scholar] [CrossRef] [PubMed]
- Bersényi, A.; Berta, E.; Kádár, I.; Glávits, R.; Szilágyi, M.; Fekete, S.G. Effects of high dietary molybdenum in rabbits. Acta Vet. Hung. 2008, 56, 41–55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khandare, A.L.; Suresh, P.; Kumar, P.U.; Lakshmaiah, N.; Manjula, N.; Rao, G.S. Beneficial effect of copper supplementation on deposition of fluoride in bone Influoride- and Molybdenum-Fed rabbits. Calcif. Tissue Int. 2005, 77, 233–238. [Google Scholar] [CrossRef] [PubMed]
- Hosseini, M.J.; Shaki, F.; Ghazi-Khansari, M.; Pourahmad, J. Toxicity of copper on isolated liver mitochondria: Impairment at complexes I, II, and IV leads to increased ROS production. Cell Biochem. Biophys. 2014, 70, 367–381. [Google Scholar] [CrossRef] [PubMed]
- Rastmanesh, R. High polyphenol, low probiotic diet for weight loss because of intestinal microbiota interaction. Chem. Biol. Interact. 2011, 189, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Yilmaz, B.; Li, H. Gut Microbiota and Iron: The Crucial Actors in Health and Disease. Pharmaceuticals 2018, 11, 98. [Google Scholar] [CrossRef] [PubMed]
- Dostal, A.; Fehlbaum, S.; Chassard, C.; Zimmermann, M.B.; Lacroix, C. Low iron availability in continuous in vitro colonic fermentations induces strong dysbiosis of the child gut microbial consortium and a decrease in main metabolites. FEMS Microbiol. Ecol. 2013, 83, 161–175. [Google Scholar] [CrossRef] [PubMed]
- Shin, N.R.; Whon, T.W.; Bae, J.W. Proteobacteria: Microbial signature of dysbiosis in gut microbiota. Trends Biotechnol. 2015, 33, 496–503. [Google Scholar] [CrossRef]
- Guo, X.; Cheng, M.; Zhang, X.; Cao, J.; Wu, Z.; Weng, P. Green tea polyphenols reduce obesity in High-Fat Diet-Induce mice by modulating intestinal microbiota composition. Int. J. Food Sci. Technol. 2017, 52, 1723–1730. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Items 1 | T-AOC, U/mgprot | GSH-ST, U/mgprot | GSH, μmol/gprot | SOD, U/mgprot | MDA, mmol/gprot | GSH-Px, U/mgprot | |
|---|---|---|---|---|---|---|---|
| MO, mg/kg | TP, mg/kg | ||||||
| 0 | 0 | 5.40 ± 0.46 a | 110.61 ± 23.21 a | 89.48 ± 16.31 b | 20.34 ± 4.10 | 4.76 ± 1.21 b | 1706.1 ± 189.2 |
| 0 | 600 | 4.80 ± 0.29 a | 116.05 ± 16.20 a | 106.94 ± 11.32 a | 19.42 ± 2.78 | 5.72 ± 0.78 b | 1616.8 ± 98.3 |
| 100 | 0 | 3.55 ± 0.54 b | 69.35 ± 23.22 b | 47.72 ± 16.20 c | 13.89 ± 5.77 | 8.57 ± 1.32 a | 1469.1 ± 210.1 |
| 100 | 600 | 3.84 ± 0.33 b | 99.95 ± 19.19 a,b | 82.20 ± 17.81 b | 18.42 ± 8.21 | 6.45 ± 1.89 b | 1942.6 ± 245.8 |
| p-Value | 0.06 | 0.03 | <0.01 | 0.28 | 0.04 | 0.73 | |
| p-Value | |||||||
| MO | 0.04 | <0.01 | 0.40 | 0.41 | 0.01 | 0.96 | |
| TP | 0.16 | 0.12 | 0.11 | 0.43 | 0.65 | 0.18 | |
| MO*TP 2 | 0.29 | 0.04 | 0.01 | 0.16 | 0.01 | 0.53 | |
| Items 1 | Firmicutes | Proteobacteria | Bacteroidetes | Actinobacteria | Gemmatimonadetes | Firmicutes/Bacteroidetes | |
|---|---|---|---|---|---|---|---|
| MO, mg/kg | TP, mg/kg | ||||||
| 0 | 0 | 75.68 ± 6.78 a | 5.34 ± 2.18b | 10.52 ± 4.25 | 5.25 ± 1.21 | 0.79 ± 0.09 b | 23.10 ± 8.24 a |
| 0 | 600 | 86.36 ± 8.99 a | 2.15 ± 0.89b | 5.98 ± 2.34 | 4.26 ± 0.78 | 0.10 ± 0.03 b | 19.29 ± 6.10 a |
| 100 | 0 | 41.77 ± 5.78 b | 30.48 ± 2.21a | 12.39 ± 4.21 | 5.85 ± 0.45 | 2.52 ± 0.21 a | 4.53±0.37 b |
| 100 | 600 | 77.78 ± 7.91 a | 8.18 ± 1.21b | 4.12 ± 1.09 | 5.43 ± 0.37 | 0.71 ± 0.10b | 29.46±8.98 a |
| p-Value | <0.01 | <0.01 | 0.28 | 0.86 | <0.01 | 0.01 | |
| p-Value | |||||||
| MO | <0.01 | <0.01 | 0.99 | 0.52 | <0.01 | 0.60 | |
| TP | <0.01 | <0.01 | 0.06 | 0.61 | <0.01 | 0.19 | |
| MO*TP 2 | 0.03 | <0.01 | 0.57 | 0.84 | 0.05 | 0.05 | |
| Items 1 | OTU | Chao1 | Shannon | Simpson | |
|---|---|---|---|---|---|
| MO, mg/kg | TP, mg/kg | ||||
| 0 | 0 | 551.4 ± 109.3 | 1486.1 ± 254.8 | 5.56 ± 0.76 | 0.91 ± 0.07 |
| 0 | 600 | 452.6 ± 145.5 | 1108.6 ± 227.6 | 4.45 ± 0.91 | 0.81 ± 0.11 |
| 100 | 0 | 815.5 ± 178.2 | 1865.7 ± 363.7 | 7.01 ± 0.77 | 0.96 ± 0.38 |
| 100 | 600 | 588.0 ± 139.2 | 1555.4 ± 290.6 | 5.34 ± 0.44 | 0.90 ± 0.06 |
| p-Value | 0.04 | 0.19 | 0.04 | 0.15 | |
| p-Value | |||||
| MO | 0.01 | 0.09 | 0.05 | 0.10 | |
| TP | 0.01 | 0.15 | 0.02 | 0.48 | |
| MO*TP 2 | 0.18 | 0.88 | 0.61 | 0.57 | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, J.; Yang, Z.; Celi, P.; Yan, L.; Ding, X.; Bai, S.; Zeng, Q.; Mao, X.; Feng, B.; Xu, S.; et al. Alteration of the Antioxidant Capacity and Gut Microbiota under High Levels of Molybdenum and Green Tea Polyphenols in Laying Hens. Antioxidants 2019, 8, 503. https://doi.org/10.3390/antiox8100503
Wang J, Yang Z, Celi P, Yan L, Ding X, Bai S, Zeng Q, Mao X, Feng B, Xu S, et al. Alteration of the Antioxidant Capacity and Gut Microbiota under High Levels of Molybdenum and Green Tea Polyphenols in Laying Hens. Antioxidants. 2019; 8(10):503. https://doi.org/10.3390/antiox8100503
Chicago/Turabian StyleWang, Jianping, Zengqiao Yang, Pietro Celi, Lei Yan, Xuemei Ding, Shiping Bai, Qiufeng Zeng, Xiangbing Mao, Bing Feng, Shengyu Xu, and et al. 2019. "Alteration of the Antioxidant Capacity and Gut Microbiota under High Levels of Molybdenum and Green Tea Polyphenols in Laying Hens" Antioxidants 8, no. 10: 503. https://doi.org/10.3390/antiox8100503