Germination Improves the Polyphenolic Profile and Functional Value of Mung Bean (Vigna radiata L.)

 , ,
, ,  ,
,  , , , ,
, , , ,
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Mung Bean Seeds
2.2. Germination Process
2.3. Acetonic Extraction and Extracts’ Preparation
2.4. Determination of Total Polyphenols
2.5. Inhibition of Lipid Peroxidation
2.6. Elemental Composition of Raw and 3-day Germinated Mung Bean Flours and Extracts
2.7. Mass Spectrometry Analysis of Extracts’ Polyphenol Composition
2.8. Cell Culture Experiments
2.9. Extracts’ Preparation for Cell Culture Studies
2.10. Cell Viability and Metabolic Activity Assays in HT29 Cells
2.11. Cell Viability Assay in U937 Cells
2.12. Flow Cytometric Determination of Reactive Oxygen Species (ROS)
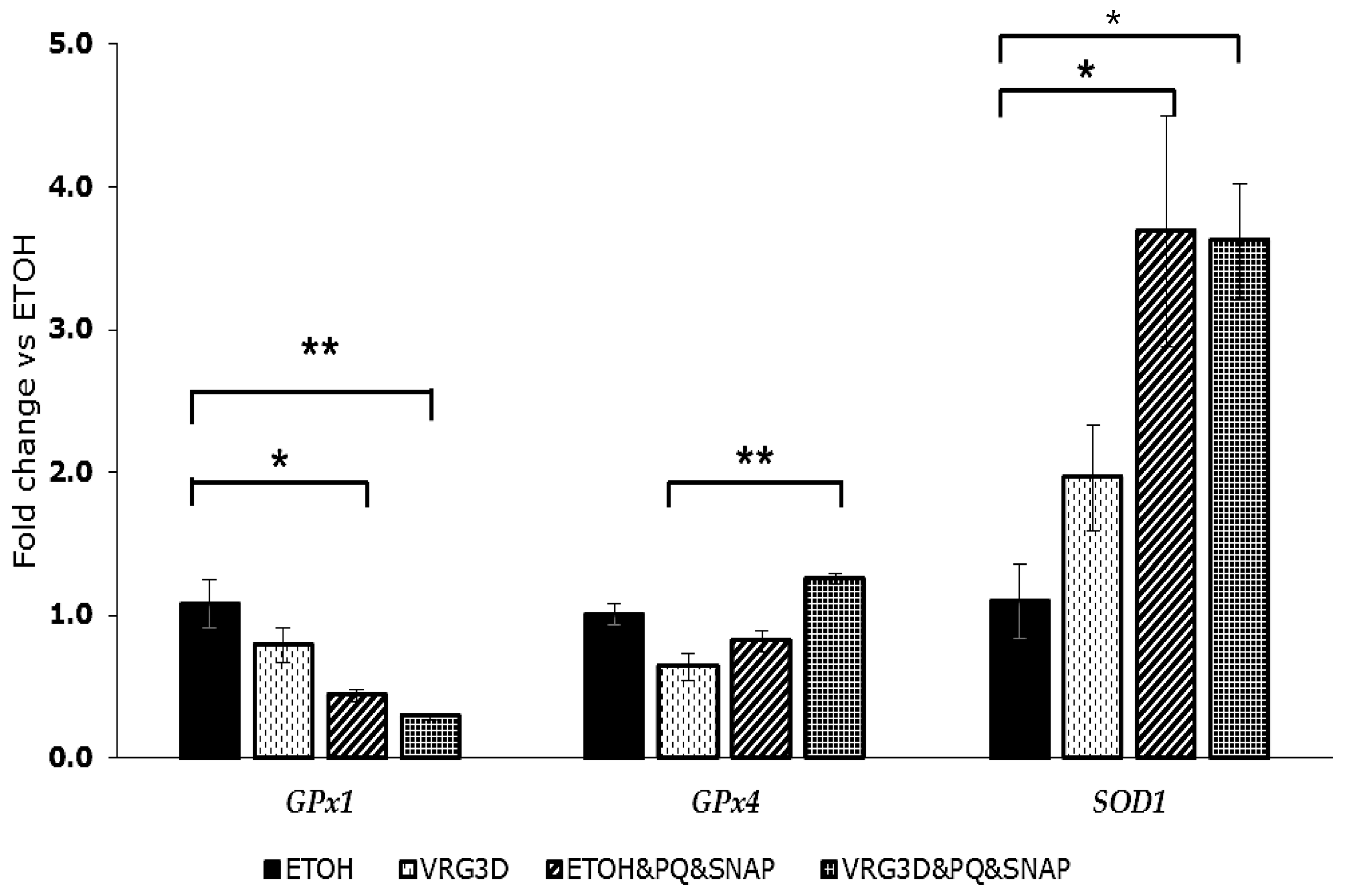
2.13. Quantitative Reverse Transcription Polymerase Chain Reaction Analysis of Antioxidant Enzymes Gene Expression
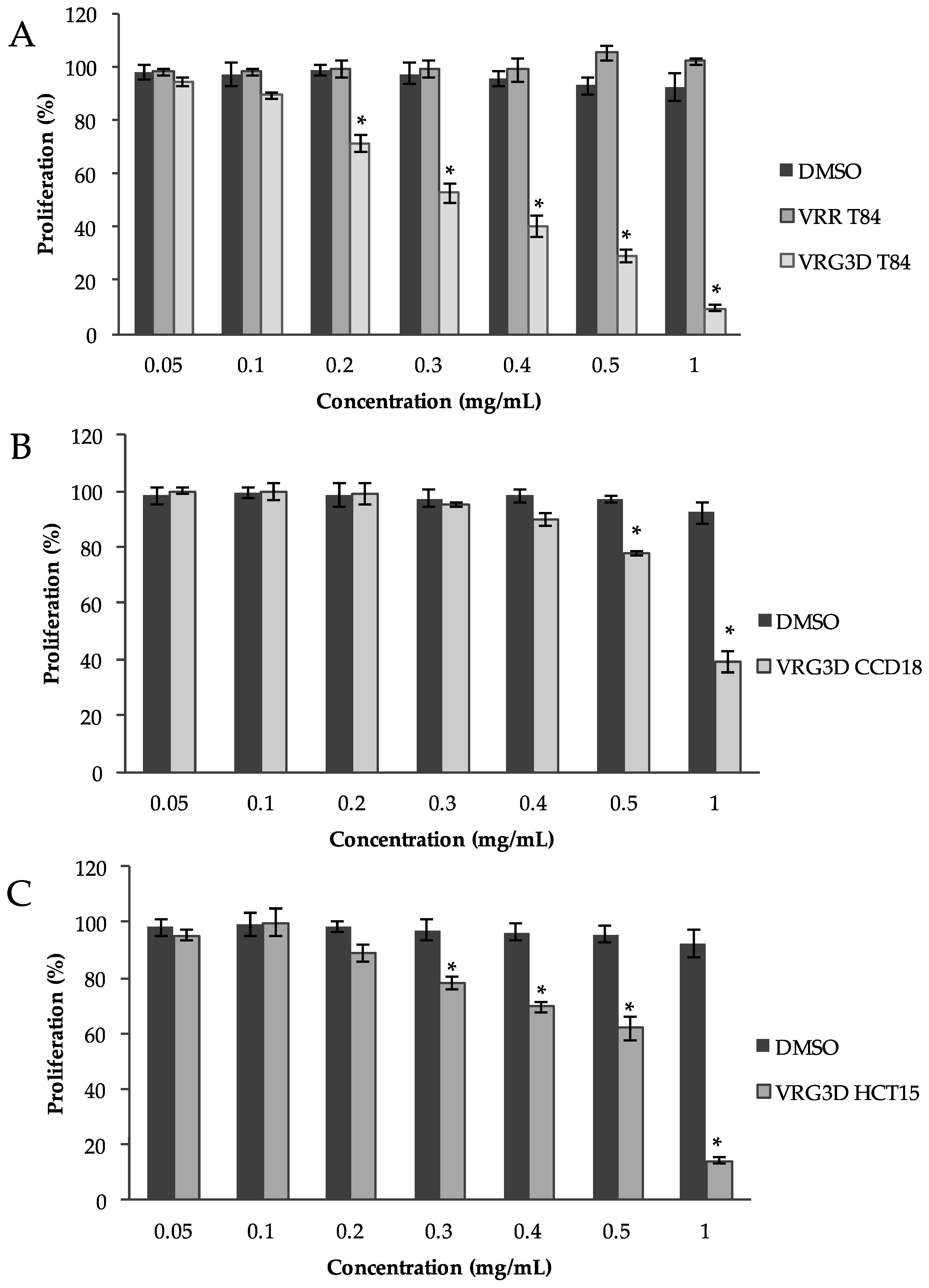
2.14. Cytotoxicity Screening in T84, HCT-15 and CCD-18 Cells
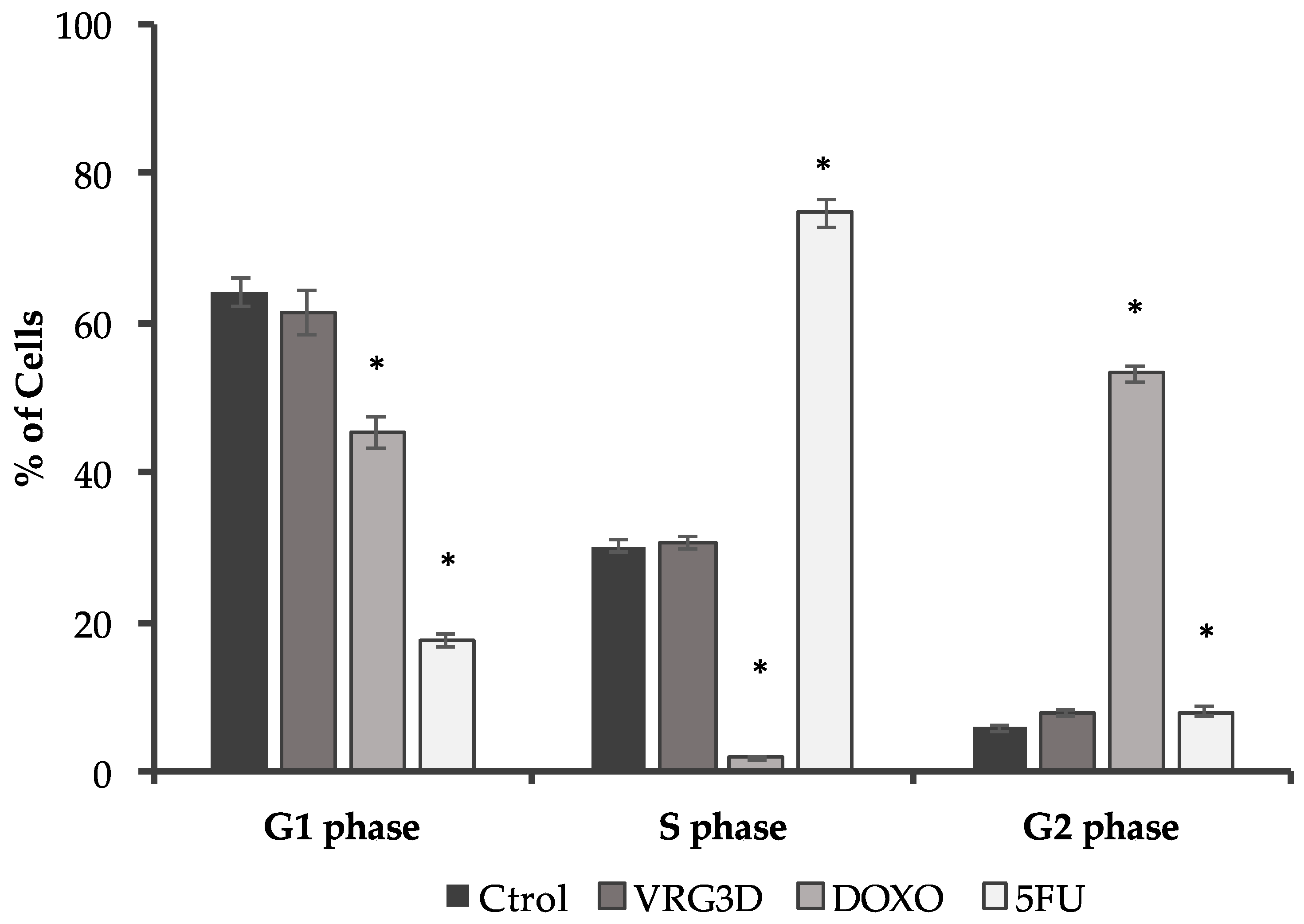
2.15. Flow Cytometric Determination of Cell Cycle
2.16. Statistical Analysis
3. Results
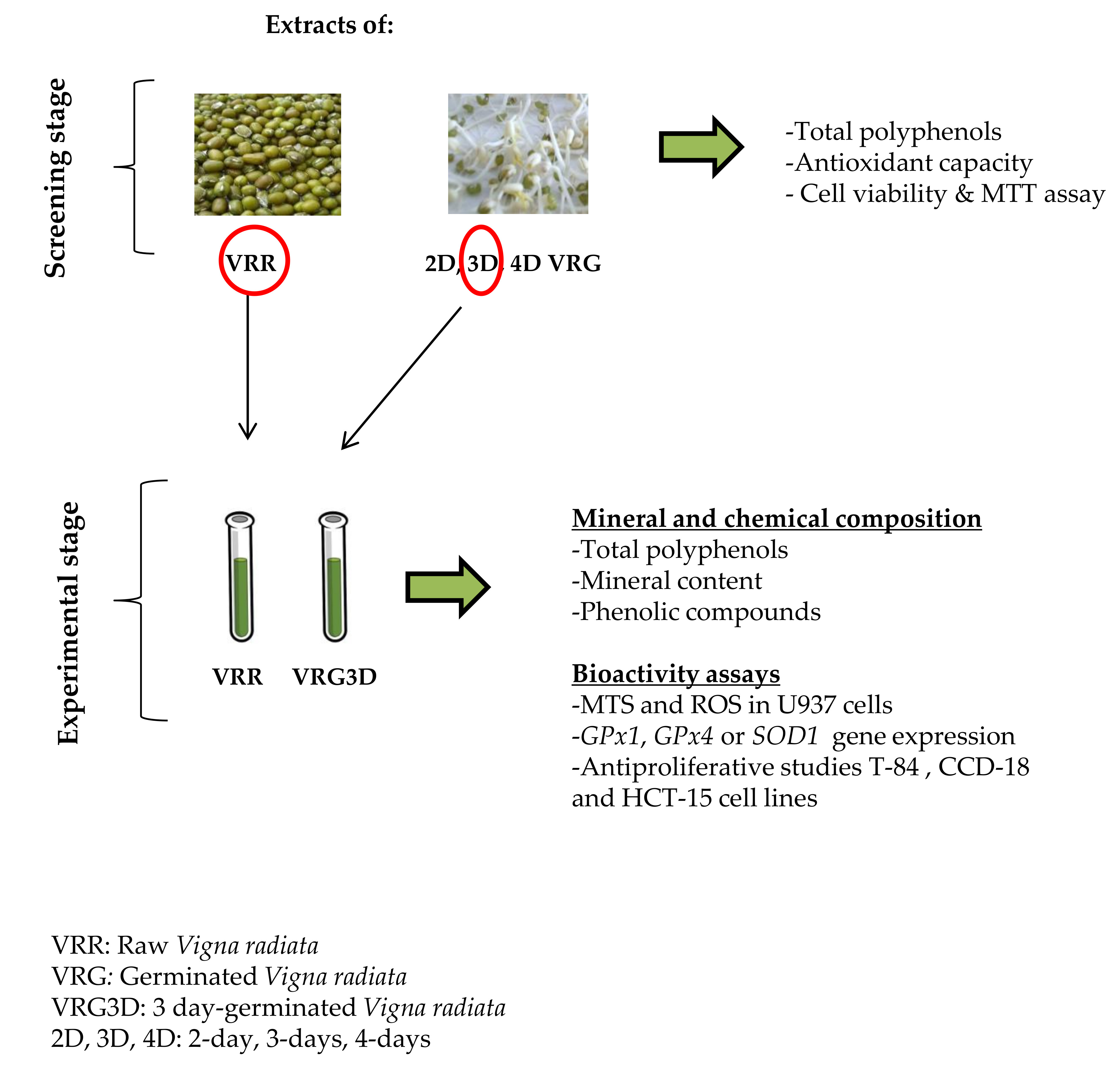
3.1. Screening Stage
3.2. Selection of Optimal Germination Conditions and Analytical Stage
3.2.1. Extraction Yield, Total Polyphenol and Mineral Content of Raw and 3-day-Geminated Mung Bean Extracts
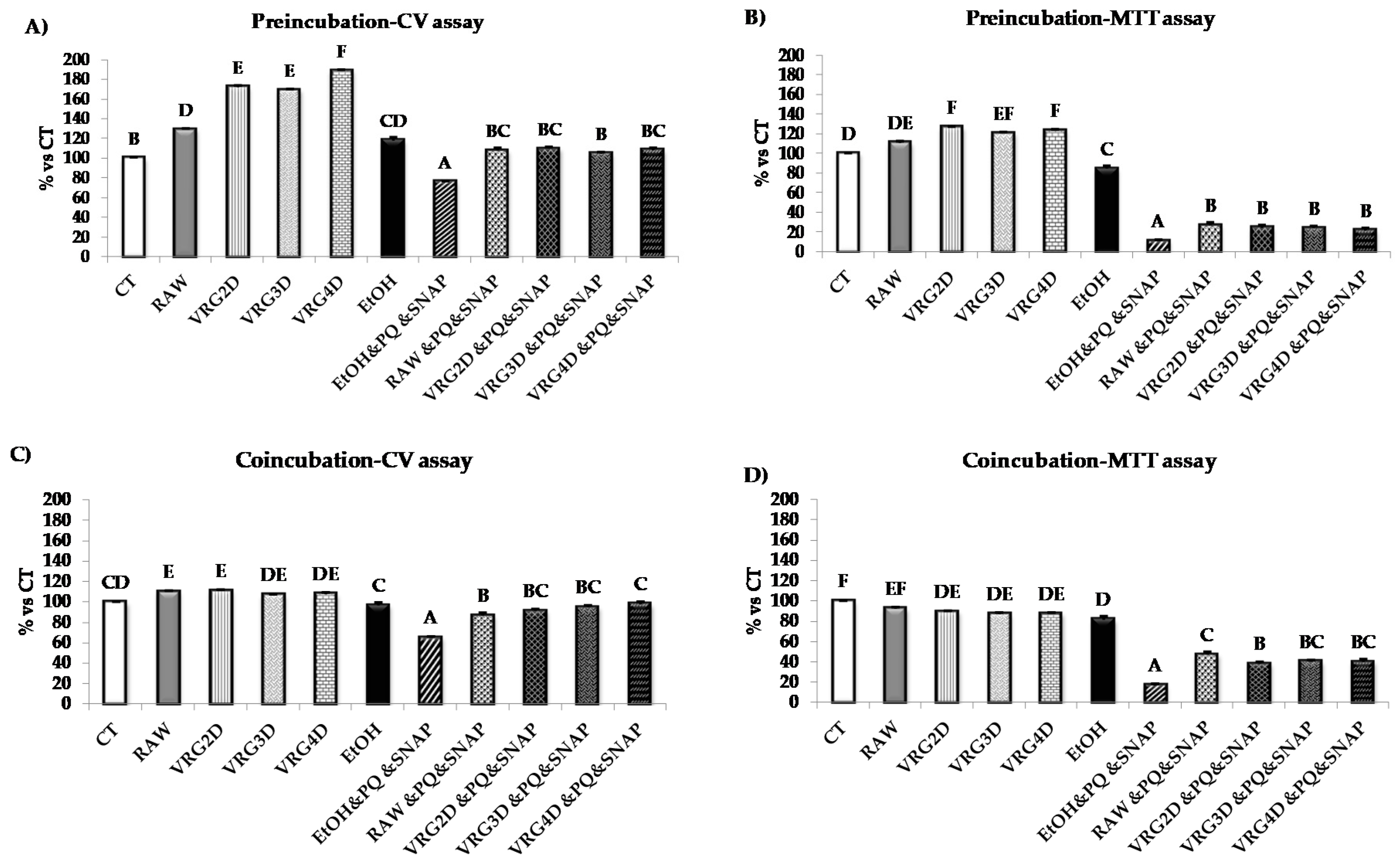
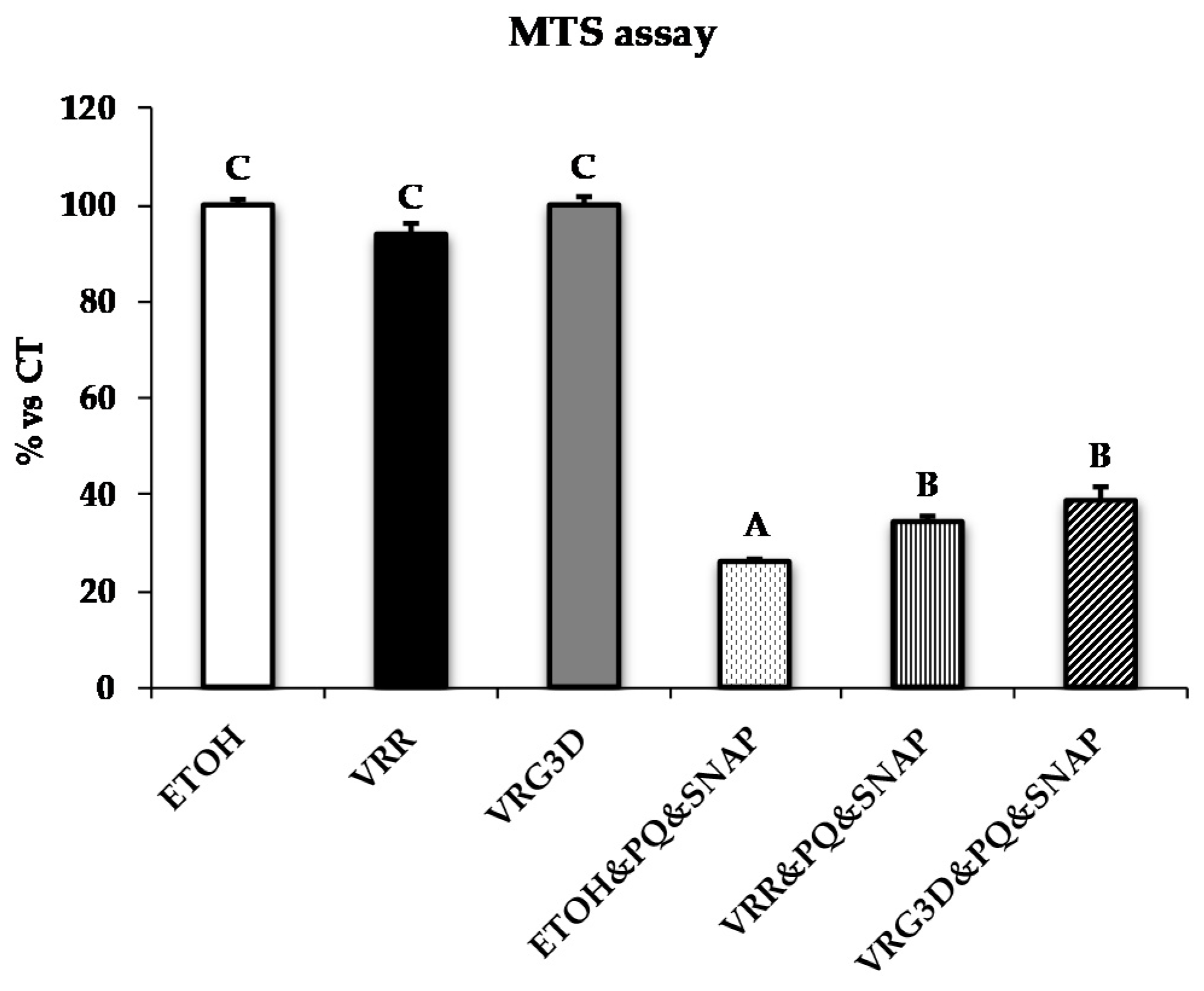
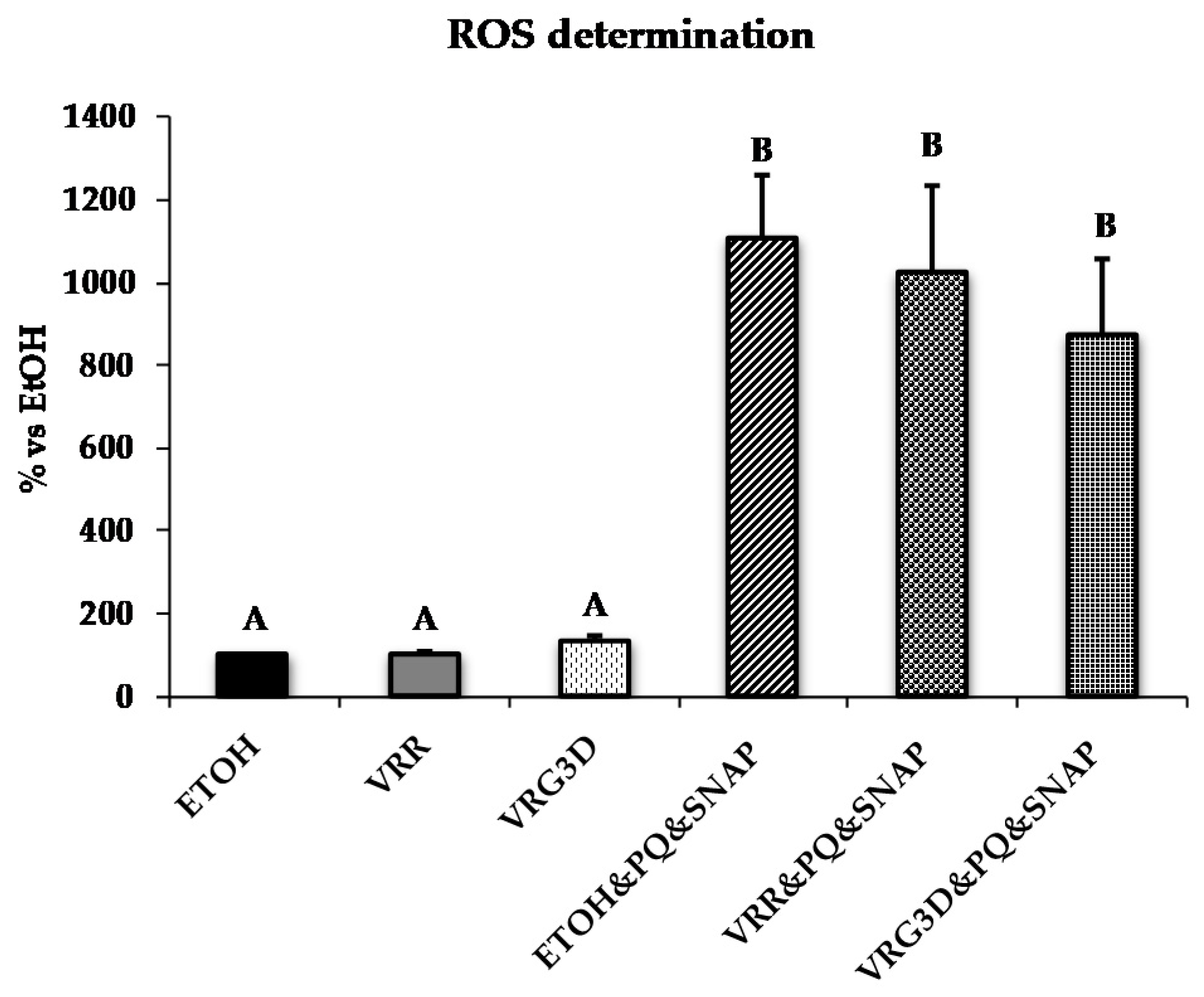
3.2.2. Bioactivity of Raw and 3-day-Geminated Mung Bean Extracts
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Gutiérrez-Uribe, J.A.; Guajardo-Flores, D.; López-Barrios, L. Legumes in the diet. In Encyclopedia of Food and Health; Academic Press: Oxford, UK, 2016; pp. 539–543. ISBN 978-0-12-384953-3. [Google Scholar]
- Bouchenak, M.; Lamri-Senhadji, M. Nutritional quality of legumes, and their role in cardiometabolic risk prevention: A review. J. Med. Food. 2013, 16, 185–198. [Google Scholar] [CrossRef] [PubMed]
- Martínez, R.; López-Jurado, M.; Wanden-Berghe, C.; Sanz-Valero, J.; Porres, J.M.; Kapravelou, G. Beneficial effects of legumes on parameters of the metabolic syndrome: A systematic review of trials in animal models. Br. J. Nutr. 2016, 116, 402–424. [Google Scholar] [CrossRef]
- Ferlay, J.; Colombet, M.; Soerjomataram, I.; Mathers, C.; Parkin, D.M.; Piñeros, M.; Znaor, A.; Bray, F. Estimating the global cancer incidence and mortality in 2018: GLOBOCAN sources and methods. Int. J. Cancer. 2019, 144, 1941–1953. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Cutsem, E.; Köhne, C.-H.; Hitre, E.; Zaluski, J.; Chang Chien, C.-R.; Makhson, A.; D’Haens, G.; Pintér, T.; Lim, R.; Bodoky, G.; et al. Cetuximab and chemotherapy as initial treatment for metastatic colorectal cancer. N. Engl. J. Med. 2009, 360, 1408–1417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chow, F.C.L.; Chok, K.S.H. Colorectal liver metastases: An update on multidisciplinary approach. World. J. Hepatol. 2019, 11, 150–172. [Google Scholar] [CrossRef] [PubMed]
- Milano, A.; Bianco, M.A.; Buri, L.; Cipolletta, L.; Grossi, E.; Rotondano, G.; Tessari, F.; Efthymakis, K.; Neri, M. Metabolic syndrome is a risk factor for colorectal adenoma and cancer: A study in a white population using the harmonized criteria. Therap. Adv. Gastroenterol. 2019, 12, 175628481986783. [Google Scholar] [CrossRef] [Green Version]
- Obidike, O.J.; Rogers, C.R.; Caspi, C.E. Examining colorectal cancer risk awareness and food shelf use among health center patients. J. Racial. Ethn. Health. Disparities 2019, 6, 1021–1029. [Google Scholar] [CrossRef]
- Kawashima, R.; Fujimaki, M.; Ikenoue, Y.; Danjo, K.; Koizumi, W.; Ichikawa, T. Influence of an elemental diet on 5-fluorouracil-induced morphological changes in the mouse salivary gland and colon. Support. Care. Cancer. 2016, 24, 1609–1616. [Google Scholar] [CrossRef]
- Manco, M.; Bottazzo, G.; DeVito, R.; Marcellini, M.; Mingrone, G.; Nobili, V. Nonalcoholic fatty liver disease in children. J. Am. Coll. Nutr. 2008, 27, 667–676. [Google Scholar] [CrossRef]
- Ryan, M.C.; Itsiopoulos, C.; Thodis, T.; Ward, G.; Trost, N.; Hofferberth, S.; O’Dea, K.; Desmond, P.V.; Johnson, N.A.; Wilson, A.M. The Mediterranean diet improves hepatic steatosis and insulin sensitivity in individuals with non-alcoholic fatty liver disease. J. Hepatol. 2013, 59, 138–143. [Google Scholar] [CrossRef]
- Reverri, E.J.; Randolph, J.M.; Steinberg, F.M.; Kappagoda, C.T.; Edirisinghe, I.; Burton-Freeman, B.M. Black beans, fiber, and antioxidant capacity pilot study: Examination of whole foods vs. functional components on postprandial metabolic, oxidative stress, and inflammation in adults with metabolic syndrome. Nutrients 2015, 7, 6139–6154. [Google Scholar] [CrossRef] [PubMed]
- Kapravelou, G.; Martínez, R.; Andrade, A.M.; Sánchez, C.; Chaves, C.L.; López-Jurado, M.; Aranda, P.; Cantarero, S.; Arrebola, F.; Fernández-Segura, E.; et al. Health promoting effects of Lupin (Lupinus albus var. multolupa) protein hydrolyzate and insoluble fiber in a diet-induced animal experimental model of hypercholesterolemia. Food. Res. Int. 2013, 54, 1471–1481. [Google Scholar] [CrossRef]
- Martínez, R.; Kapravelou, G.; Porres, J.M.; Melesio, A.M.; Heras, L.; Cantarero, S.; Gribble, F.M.; Parker, H.; Aranda, P.; López-Jurado, M. Medicago sativa L., a functional food to relieve hypertension and metabolic disorders in a spontaneously hypertensive rat model. J. Funct. Foods 2016, 26, 470–484. [Google Scholar] [CrossRef]
- Huang, X.; Cai, W.; Xu, B. Kinetic changes of nutrients and antioxidant capacities of germinated soybean (Glycine max L.) and mung bean (Vigna radiata L.) with germination time. Food Chem. 2014, 143, 268–276. [Google Scholar] [CrossRef] [PubMed]
- Dahiya, P.K.; Linnemann, A.R.; van Boekel, M.A.J.S.; Khetarpaul, N.; Grewal, R.B.; Nout, M.J.R. Mung bean: Technological and nutritional potential. Crit. Rev. Food Sci. Nutr. 2013, 55, 670–688. [Google Scholar] [CrossRef]
- Anwar, F.; Latif, S.; Przybylski, R.; Sultana, B.; Ashraf, M. Chemical composition and antioxidant activity of seeds of different cultivars of mungbean. J. Food Sci. 2007, 72, S503–S510. [Google Scholar] [CrossRef]
- Yao, Y.; Cheng, X.; Wang, L.; Wang, S.; Ren, G. Biological potential of sixteen legumes in China. Int. J. Mol. Sci. 2011, 12, 7048–7058. [Google Scholar] [CrossRef] [Green Version]
- Urbano, G.; López-Jurado, M.; ławomir Frejnagel, S.; Gómez-Villalva, E.; Porres, J.M.; Frías, J.; Vidal-Valverde, C.; Aranda, P. Nutritional assessment of raw and germinated pea (Pisum sativum L.) protein and carbohydrate by in vitro and in vivo techniques. Nutrition 2005, 21, 230–239. [Google Scholar] [CrossRef]
- Fernandez-Orozco, R.; Frias, J.; Zielinski, H.; Piskula, M.K.; Kozlowska, H.; Vidal-Valverde, C. Kinetic study of the antioxidant compounds and antioxidant capacity during germination of Vigna radiata cv. emmerald, Glycine max cv. jutro and Glycine max cv. merit. Food Chem. 2008, 111, 622–630. [Google Scholar] [CrossRef]
- Hithamani, G.; Srinivasan, K. Bioaccessibility of polyphenols from wheat (Triticum aestivum), sorghum (Sorghum bicolor), green gram (Vigna radiata), and chickpea (Cicer arietinum) as influenced by domestic food processing. J. Agric. Food Chem. 2014, 62, 11170–11179. [Google Scholar] [CrossRef]
- Guo, X.; Li, T.; Tang, K.; Liu, R.H. Effect of germination on phytochemical profiles and antioxidant activity of mung bean sprouts (Vigna radiata). J. Agric. Food Chem. 2012, 60, 11050–11055. [Google Scholar] [CrossRef] [PubMed]
- Doblado, R.; Frías, J.; Vidal-Valverde, C. Changes in vitamin C content and antioxidant capacity of raw and germinated cowpea (Vigna sinensis var. carilla) seeds induced by high pressure treatment. Food Chem. 2007, 101, 918–923. [Google Scholar] [CrossRef]
- Dai, J.; Mumper, R.J. Plant phenolics: Extraction, analysis and their antioxidant and anticancer properties. Molecules 2010, 15, 7313–7352. [Google Scholar] [CrossRef] [PubMed]
- Dewanto, V.; Wu, X.; Adom, K.K.; Liu, R.H. Thermal processing enhances the nutritional value of tomatoes by increasing total antioxidant activity. J. Agric. Food Chem. 2002, 50, 3010–3014. [Google Scholar] [CrossRef]
- Ohkawa, H.; Ohishi, N.; Yagi, K. Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction. Anal. Biochem. 1979, 95, 351–358. [Google Scholar] [CrossRef]
- Kapravelou, G.; Martínez, R.; Andrade, A.M.; Nebot, E.; Camiletti-Moirón, D.; Aparicio, V.A.; Lopez-Jurado, M.; Aranda, P.; Arrebola, F.; Fernandez-Segura, E.; et al. Aerobic interval exercise improves parameters of nonalcoholic fatty liver disease (NAFLD) and other alterations of metabolic syndrome in obese Zucker rats. Appl. Physiol. Nutr. Metab. 2015, 40, 1242–1252. [Google Scholar] [CrossRef] [Green Version]
- Sánchez, C.; Torres, M.; Bermúdez-Peña, M.C.; Aranda, P.; Montes-Bayón, M.; Sanz-Medel, A.; Llopis, J. Bioavailability, tissue distribution and hypoglycaemic effect of vanadium in magnesium-deficient rats. Magnes. Res. 2011, 24, 196–208. [Google Scholar] [CrossRef]
- Martínez, R.; Kapravelou, G.; Donaire, A.; Lopez-Chaves, C.; Arrebola, F.; Galisteo, M.; Cantarero, S.; Aranda, P.; Porres, J.M.; López-Jurado, M. Effects of a combined intervention with a lentil protein hydrolysate and a mixed training protocol on the lipid metabolism and hepatic markers of NAFLD in Zucker rats. Food Funct. 2018, 9, 830–850. [Google Scholar] [CrossRef]
- Fu, Y.; Porres, J.M.; Lei, X.G. Comparative impacts of glutathione peroxidase-1 gene knockout on oxidative stress induced by reactive oxygen and nitrogen species in mouse hepatocytes. Biochem. J. 2001, 359, 687–695. [Google Scholar] [CrossRef]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Meth. 1983, 65, 55–63. [Google Scholar] [CrossRef]
- Bermano, G.; Smyth, E.; Goua, M.; Heys, S.D.; Wahle, K.W.J. Impaired expression of glutathione peroxidase-4 gene in peripheral blood mononuclear cells: A biomarker of increased breast cancer risk. Cancer. Biomark. 2010, 7, 39–46. [Google Scholar] [CrossRef] [PubMed]
- Ukkola, O.; Erkkilä, P.H.; Savolainen, M.J.; Kesäniemi, Y.A. Lack of association between polymorphisms of catalase, copper-zinc superoxide dismutase (SOD), extracellular SOD and endothelial nitric oxide synthase genes and macroangiopathy in patients with type 2 diabetes mellitus. J. Intern. Med. 2001, 249, 451–459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gurib-Fakim, A. Medicinal plants: Traditions of yesterday and drugs of tomorrow. Mol. Aspects. Med. 2006, 27, 1–93. [Google Scholar] [CrossRef] [PubMed]
- Braun, S.; Bitton-Worms, K.; LeRoith, D. The link between the metabolic syndrome and cancer. Int. J. Biol. Sci. 2011, 7, 1003–1015. [Google Scholar] [CrossRef] [PubMed]
- Bahadoran, Z. Potential properties of legumes as important functional foods for management of Type 2 diabetes: A short review. Int. J. Nutr. Food Sci. 2015, 4, 6. [Google Scholar] [CrossRef] [Green Version]
- Messina, V. Nutritional and health benefits of dried beans. Am. J. Clin. Nutr. 2014, 100 (Suppl. 1), 437S–442S. [Google Scholar] [CrossRef] [Green Version]
- Randhir, R.; Shetty, K. Mung beans processed by solid-state bioconversion improves phenolic content and functionality relevant for diabetes and ulcer management. Innov. Food Sci. Emerg. Technol. 2007, 8, 197–204. [Google Scholar] [CrossRef]
- Peng, X.; Zheng, Z.; Cheng, K.-W.; Shan, F.; Ren, G.-X.; Chen, F.; Wang, M. Inhibitory effect of mung bean extract and its constituents vitexin and isovitexin on the formation of advanced glycation endproducts. Food Chem. 2008, 106, 475–481. [Google Scholar] [CrossRef]
- Limón, R.I.; Peñas, E.; Martínez-Villaluenga, C.; Frias, J. Role of elicitation on the health-promoting properties of kidney bean sprouts. LWT. Food Sci. Technol. 2014, 56, 328–334. [Google Scholar] [CrossRef] [Green Version]
- Cao, D.; Li, H.; Yi, J.; Zhang, J.; Che, H.; Cao, J.; Yang, L.; Zhu, C.; Jiang, W. Antioxidant properties of the mung bean flavonoids on alleviating heat stress. PLoS ONE. 2011, 6, e21071. [Google Scholar] [CrossRef] [Green Version]
- Tang, D.; Dong, Y.; Guo, N.; Li, L.; Ren, H. Metabolomic analysis of the polyphenols in germinating mung beans (Vigna radiata) seeds and sprouts. J. Sci. Food Agric. 2014, 94, 1639–1647. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.-K.; Kim, E.-H.; Lee, O.-K.; Park, S.-Y.; Lee, B.; Kim, S.-H.; Park, I.; Chung, I.-M. Variation and correlation analysis of phenolic compounds in mungbean (Vigna radiata L.) varieties. Food Chem. 2013, 141, 2988–2997. [Google Scholar] [CrossRef] [PubMed]
- Porres, J.M.; Aranda, P.; López-Jurado, M.; Urbano, G. Effect of natural and controlled fermentation on chemical composition and nutrient dialyzability from beans (Phaseolus vulgaris L.). J. Agric. Food Chem. 2003, 51, 5144–5149. [Google Scholar] [CrossRef] [PubMed]
- Urbano, G.; López-Jurado, M.; Aranda, C.; Vilchez, A.; Cabrera, L.; Porres, J.M.; Aranda, P. Evaluation of zinc and magnesium bioavailability from pea (Pisum sativum, L.) sprouts. Effect of illumination and different germination periods. Int. J. Food Sci. Technol. 2006, 41, 618–626. [Google Scholar] [CrossRef]
- Treviño, S.; Díaz, A.; Sánchez-Lara, E.; Sanchez-Gaytan, B.L.; Perez-Aguilar, J.M.; González-Vergara, E. Vanadium in biological action: Chemical, pharmacological aspects, and metabolic implications in diabetes mellitus. Biol. Trace Elem. Res. 2019, 188, 68–98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, J.; Wang, X.; Li, S.; Zhu, Y.; Chen, S.; Li, P.; Luo, C.; Huang, Y.; Li, X.; Hu, X.; et al. Interactions between plasma copper concentrations and SOD1 gene polymorphism for impaired glucose regulation and type 2 diabetes. Redox Biol. 2019, 24, 101172. [Google Scholar] [CrossRef] [PubMed]
- Kuo, C.-C.; Moon, K.A.; Wang, S.-L.; Silbergeld, E.; Navas-Acien, A. The association of arsenic metabolism with cancer, cardiovascular disease, and diabetes: A systematic review of the epidemiological evidence. Environ. Health Perspect. 2017, 125, 087001. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Yang, R.; Jin, X.; Chen, Z.; Zhou, Y.; Gu, Z. Effect of germination and incubation on Zn, Fe, and Ca bioavailability values of soybeans (Glycine max L.) and mung beans (Vigna radiata L.). Food Sci. Biotechnol. 2015, 24, 1829–1835. [Google Scholar] [CrossRef]
- Kapravelou, G.; Martínez, R.; Nebot, E.; López-Jurado, M.; Aranda, P.; Arrebola, F.; Cantarero, S.; Galisteo, M.; Porres, J.M. The combined intervention with germinated Vigna radiata and aerobic interval training protocol is an effective strategy for the treatment of Non-Alcoholic Fatty Liver Disease (NAFLD) and other alterations related to the metabolic syndrome in Zucker rats. Nutrients 2017, 9, 774. [Google Scholar] [CrossRef] [Green Version]
- Xu, C.; Arinze, I.J.; Johnson, J.; Tuy, T.T.; Bone, F.; Ernsberger, P.; Massillon, D. Metabolic dysregulation in the SHROB rat reflects abnormal expression of transcription factors and enzymes that regulate carbohydrate metabolism. J. Nutr. Biochem. 2008, 19, 305–312. [Google Scholar] [CrossRef]
- Zengin, M.; Özcan, M.M.; Çetin, Ü.; Gezgin, S. Mineral contents of some aromatic plants, their growth soils and infusions. J. Sci. Food Agric. 2008, 88, 581–589. [Google Scholar] [CrossRef]
- Dimitrijević, D.S.; Kostić, D.A.; Stojanović, G.S.; Mitić, S.S.; Mitić, M.N.; Dorđević, A.S. Phenolic composition, antioxidant activity, mineral content and antimicrobial activity of fresh fruit extracts of Morus alba L. J. Food. Nutr Res. 2014, 53, 22–30. [Google Scholar]
- Then, M.; Szentmihályi, K.; Sárközi, A.; Illés, V.; Forgács, E. Effect of sample handling on alkaloid and mineral content of aqueous extracts of greater celandine (Chelidonium majus L.). J. Chromatogr. A 2000, 889, 69–74. [Google Scholar] [CrossRef]
- Tomlinson, D.J.; Erskine, R.M.; Morse, C.I.; Onambélé, G.L. Body fat percentage, body mass index, fat mass index and the ageing bone: Their singular and combined roles linked to physical activity and diet. Nutrients 2019, 11, 195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aslam, M.N.; Kreider, J.M.; Paruchuri, T.; Bhagavathula, N.; DaSilva, M.; Zernicke, R.F.; Goldstein, S.A.; Varani, J. A mineral-rich extract from the red marine algae Lithothamnion calcareum preserves bone structure and function in female mice on a western-style diet. Calcif. Tissue. Int. 2010, 86, 313–324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yeap, S.K.; Beh, B.K.; Ali, N.M.; Mohd Yusof, H.; Ho, W.Y.; Koh, S.P.; Alitheen, N.B.; Long, K. In vivo antistress and antioxidant effects of fermented and germinated mung bean. Biomed. Res. Int. 2014, 2014, 694842. [Google Scholar] [CrossRef] [Green Version]
- Bhardwaj, M.; Paul, S.; Jakhar, R.; Kang, S.C. Potential role of vitexin in alleviating heat stress-induced cytotoxicity: Regulatory effect of Hsp90 on ER stress-mediated autophagy. Life Sci. 2015, 142, 36–48. [Google Scholar] [CrossRef]
- Hattori, H.; Imai, H.; Furuhama, K.; Sato, O.; Nakagawa, Y. Induction of phospholipid hydroperoxide glutathione peroxidase in human polymorphonuclear neutrophils and HL60 cells stimulated with TNF-α. Biochem. Biophys. Res. Commun. 2005, 337, 464–473. [Google Scholar] [CrossRef]
- Pagmantidis, V.; Méplan, C.; van Schothorst, E.M.; Keijer, J.; Hesketh, J.E. Supplementation of healthy volunteers with nutritionally relevant amounts of selenium increases the expression of lymphocyte protein biosynthesis genes. Am. J. Clin. Nutr. 2008, 87, 181–189. [Google Scholar] [CrossRef] [Green Version]
- Huang, Z.; Rose, A.H.; Hoffmann, P.R. The Role of Selenium in Inflammation and Immunity: From Molecular Mechanisms to Therapeutic Opportunities. Antioxid. Redox. Signal. 2012, 16, 705–743. [Google Scholar] [CrossRef] [Green Version]
- Fridovich, I. Superoxide anion radical (O2−.), superoxide dismutases, and related matters. J. Biol. Chem. 1997, 272, 18515–18517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, F.; Giulivi, C. Superoxide dismutases and their impact upon human health. Mol. Aspects Med. 2005, 26, 340–352. [Google Scholar] [CrossRef] [PubMed]
- Joghatai, M.; Barari, L.; Mousavie Anijdan, S.H.; Elmi, M.M. The evaluation of radio-sensitivity of mung bean proteins aqueous extract on MCF-7, hela and fibroblast cell line. Int. J. Radiat. Biol. 2018, 94, 478–487. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.-K.; Jeong, S.C.; Gorinstein, S.; Chon, S.-U. Total polyphenols, antioxidant and antiproliferative activities of different extracts in mungbean seeds and sprouts. Plant Foods. Hum. Nutr. 2012, 67, 71–75. [Google Scholar] [CrossRef] [PubMed]
- Hafidh, R.R.; Abdulamir, A.S.; Bakar, F.A.; Jalilian, F.A.; Abas, F.; Sekawi, Z. Novel molecular, cytotoxical, and immunological study on promising and selective anticancer activity of mung bean sprouts. BMC Complement. Altern. Med. 2012, 12, 208. [Google Scholar] [CrossRef] [Green Version]
- Xu, B.; Chang, S.K.C. Comparative study on antiproliferation properties and cellular antioxidant activities of commonly consumed food legumes against nine human cancer cell lines. Food Chem. 2012, 134, 1287–1296. [Google Scholar] [CrossRef]
- Chen, Z.; Wang, J.; Liu, W.; Chen, H. Physicochemical characterization, antioxidant and anticancer activities of proteins from four legume species. J. Food Sci. Technol. 2017, 54, 964–972. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.-S.; Lee, Y.-S.; Kim, D.-K. Doxorubicin exerts cytotoxic effects through cell cycle arrest and Fas-mediated cell death. Pharmacology 2009, 84, 300–309. [Google Scholar] [CrossRef]
- Gao, L.; Liu, Q.; Liu, S.; Chen, Z.; Li, C.; Lei, L.; Sun, S.; Li, L.; Liu, J.; Shen, Z. A refined-JinQi-JiangTang tablet ameliorates prediabetes by reducing insulin resistance and improving beta cell function in mice. J. Ethnopharmacol. 2014, 151, 675–685. [Google Scholar] [CrossRef]
- Ali, N.M.; Yeap, S.-K.; Yusof, H.M.; Beh, B.-K.; Ho, W.-Y.; Koh, S.-P.; Abdullah, M.P.; Alitheen, N.B.; Long, K. Comparison of free amino acids, antioxidants, soluble phenolic acids, cytotoxicity and immunomodulation of fermented mung bean and soybean. J. Sci. Food Agric. 2016, 96, 1648–1658. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Accession Number | Primer Sequence |
|---|---|---|
| GPx1 | NG_012264.1 | F: 5′-CACCCAGATGAACGAGCTGC-3′ R: 5′-TGTACCTGCGTAGGGGCACA-3′ |
| GPx4 | NG_050621.1 | F: 5′-ATGCACGAGTTTTCCGCC-3′ R: 5′-CTAGAAATAGTGGGGCAGGTC-3′ |
| SOD1 | NG_008689.1 | F: 5′-GTAAAACGACGCCAGTGAAGCCTTGTTTGAAGAGCTG-3′ R: 5′-AAACCGCGACTAACAATCAAAG-3′ |
| β2-microglobulin | NG_012920.2 | F: 5′-GGCTATCCAGCGTACTCCAAA-3′ R: 5′-CGGCAGGCATACTCATCTTTTT-3′ |
| Screening Stage | ||||
|---|---|---|---|---|
| VRR | VRG2D | VRG3D | VRG4D | |
| Extract yield (mg·mL−1 extract) | 7.59 ± 0.46 A | 8.91 ± 0.36 A | 8.07 ± 0.90 A | 9.36 ± 0.35 A |
| Total polyphenols (μg GAE·mg−1 extract) | 29.44 ± 2.45 A | 24.11 ± 1.64 A | 30.37 ± 0.99 A | 24.93 ± 1.10 A |
| Antioxidant capacity (UAC·mg−1 extract) | 5.41 ± 0.62 A | 6.12 ± 0.93 A | 6.41 ± 0.63 A | 5.65 ± 0.28 A |
| Analytical Stage | ||
|---|---|---|
| VRR | VRG3D | |
| Total yield (g extract·100 g−1 flour) | 7.13 ± 0.23 | 17.53 ± 0.32 *** |
| Extract yield (mg extract·mL−1) | 20.08 ± 1.08 | 53.35 ± 2.96 *** |
| Total polyphenol content (μgGAE·mg−1 extract) | 42.13 ± 2.32 | 61.77 ± 3.20 *** |
| Concentration | Flours | Extracts | ||
|---|---|---|---|---|
| VRR | VRG3D | VRR | VRG3D | |
| Mg (mg·kg−1) | 1348 ± 48.9 | 1404 ± 94.8 | 4252 ± 70.6 | 2128 ± 37.5 ** |
| Ca (mg·kg−1) | 807.2 ± 15.1 | 938.7 ± 54.3 | 404.6 ± 7.8 | 338.7 ± 62.1 |
| Fe (mg·kg−1) | 48.0 ± 1.5 | 47.8 ± 3.9 | 5.36 ± 0.62 | 6.68 ± 0.08 |
| Zn (mg·kg−1) | 22.6 ± 0.8 | 27.5 ± 1.6 | 18.3 ± 0.22 | 9.57 ± 0.34 *** |
| Mn (mg·kg−1) | 11.8 ± 0.51 | 13.3 ± 0.91 | 5.12 ± 0.08 | 5.05 ± 1.01 |
| Cu (mg·kg−1) | 7.11 ± 0.22 | 7.64 ± 0.50 | 13.3 ± 0.02 | 2.90 ± 0.17 *** |
| Se (μg·kg−1) | 53.3 ± 8.3 | 96.0 ± 23.6 | 139.5 ± 19.3 | 63.0 ± 6.9 |
| Co (μg·kg−1) | 42.7 ± 1.7 | 45.3 ± 4.6 | 143.0 ± 1.2 | 73.0 ± 4.6 ** |
| V (μg·kg−1) | 14.7 ± 0.33 | 4.67 ± 1.33 * | 20.0 ± 3.5 | 12.50 ± 1.44 |
| As (μg·kg−1) | 12.0 ± 0.58 | 10.0 ± 0.58 | 10.0 ± 1.15 | 7.50 ± 0.29 |
| VRR: Phenolic Compounds in Raw Mung Bean Extracts. | ||||||
|---|---|---|---|---|---|---|
| Classification | Name | Formula | Mass | Retention Time | Mass Error | Fragments |
| Flavanones | Astilbin | C21H22O11 | 449.108 | 3.01 | −0.4 | 287.076 |
| 235.024 | ||||||
| 197.045 | ||||||
| Isoflavones | Genistein | C15H10O5 | 269.045 | 6.2 | 1.1 | NONE |
| Flavonols | Kaempferol-3-O-rutinose | C27H30O15 | 593.150 | 3.96 | −0.2 | 431.096 |
| 311.056 | ||||||
| - | ||||||
| Quercitin | C21H20O11 | 447.091 | 3.32 | −2.2 | 359.072 | |
| 357.060 | ||||||
| 327.050 | ||||||
| Rutin | C27H30O16 | 609.145 | 3.5 | 611.161 | ||
| Flavones | Vitexin | C21H20O10 | 431.097 | 3.67 | −1.6 | 311.055 |
| 283.060 | ||||||
| - | ||||||
| di-hydrochalcones | Aspalathin | C21H24O11 | 451.123 | 2.03 | −1.3 | 177.018 |
| 137.023 | ||||||
| Phloridzin | C21H24O10 | 435.130 | 4.02 | 1.1 | 315.087 | |
| 167.034 | ||||||
| others | Taxumairol N | C28H42O12 | 569.260 | 4.61 | 0.5 | 527.247 |
| 509.240 | ||||||
| VRG3D: Phenolic Compounds in 3-Day-germinated Mung Bean Extracts. Common phenolic compounds between raw and germinated extracts are underlined. | ||||||
| Flavanones | Astilbin | C21H22O11 | 449.107 | 2.92 | −1.6 | 287.055 |
| 269.044 | ||||||
| Eriodictyol | C15H12O6 | 285.055 | 5.18 | −0.7 | 217.650 | |
| 161.023 | ||||||
| 125.023 | ||||||
| Hesperetin | C16H14O6 | 301.070 | 4.76 | −2.3 | 165.018 | |
| Hesperidin | C28H34O15 | 609.179 | 4.32 | −3.3 | 517.170 | |
| 301.070 | ||||||
| Naringenin | C15H12O5 | 271.060 | 6.12 | −0.4 | 151.0015 | |
| 119.0505 | ||||||
| 107.0126 | ||||||
| Isoflavones | Genistein | C15H10O5 | 269.044 | 6.20 | −1.1 | 133.029 |
| 117.033 | ||||||
| Flavonols | Daidzein | C15H10O4 | 253.050 | 5.15 | - | |
| Kaempferol 3-O-β-d-rutinose | C27H30O15 | 593.150 | 3.96 | −1.0 | 431.096 | |
| Quercitin | C21H20O11 | 447.093 | 3.21 | 0.9 | 357.061 | |
| 285.040 | ||||||
| 253.050 | ||||||
| Rutin | C27H30O16 | 609.145 | 3.50 | −0.8 | 447.0921 | |
| Flavones | Vitexin | C21H20O10 | 431.097 | 3.65 | −0.9 | 311.055 |
| 283.060 | ||||||
| Luteolin | C15H10O6 | 285.0399 | 5.20 | −1.1 | - | |
| Phenolic acids | Ferulic acid | C10H10O4 | 193.050 | 2.10 | −4.7 | 177.055 |
| 161.024 | ||||||
| 4-Methoxycinnamic acid | C10H10O3 | 177.054 | 2.49 | −1.7 | 147.044 | |
| 131.048 | ||||||
| 93.034 | ||||||
| Aspicilin/tri-hydroxy-octadecanoic acid | C18H32O5 | 327.217 | 6.26 | 0.3 | 211.133 | |
| 171.102 | ||||||
| Isoflavanes | Glabridin | C20H20O4 | 323.129 | 10.17 | 2.5 | 203.071 |
| 119.049 | ||||||
| Flavanonol | Phellamurin | C26H30O11 | 517.171 | 5.62 | 0.6 | 355.117 |
| 193.086 | ||||||
| Coumestan | Psoralidin | C20H16O5 | 335.091 | 9.25 | −1.8 | 193.085 |
| others | Comiferylferulate | C20H20O6 | 355.117 | 8.22 | −1.1 | 193.086 |
| 161.023 | ||||||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kapravelou, G.; Martínez, R.; Perazzoli, G.; Sánchez González, C.; Llopis, J.; Cantarero, S.; Goua, M.; Bermano, G.; Prados, J.; Melguizo, C.; et al. Germination Improves the Polyphenolic Profile and Functional Value of Mung Bean (Vigna radiata L.). Antioxidants 2020, 9, 746. https://doi.org/10.3390/antiox9080746
Kapravelou G, Martínez R, Perazzoli G, Sánchez González C, Llopis J, Cantarero S, Goua M, Bermano G, Prados J, Melguizo C, et al. Germination Improves the Polyphenolic Profile and Functional Value of Mung Bean (Vigna radiata L.). Antioxidants. 2020; 9(8):746. https://doi.org/10.3390/antiox9080746
Chicago/Turabian StyleKapravelou, Garyfallia, Rosario Martínez, Gloria Perazzoli, Cristina Sánchez González, Juan Llopis, Samuel Cantarero, Marie Goua, Giovanna Bermano, Jose Prados, Consolación Melguizo, and et al. 2020. "Germination Improves the Polyphenolic Profile and Functional Value of Mung Bean (Vigna radiata L.)" Antioxidants 9, no. 8: 746. https://doi.org/10.3390/antiox9080746