Betaine and Isoquinoline Alkaloids Protect against Heat Stress and Colonic Permeability in Growing Pigs

 , ,
, ,  , , ,
, , , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Experimental Design
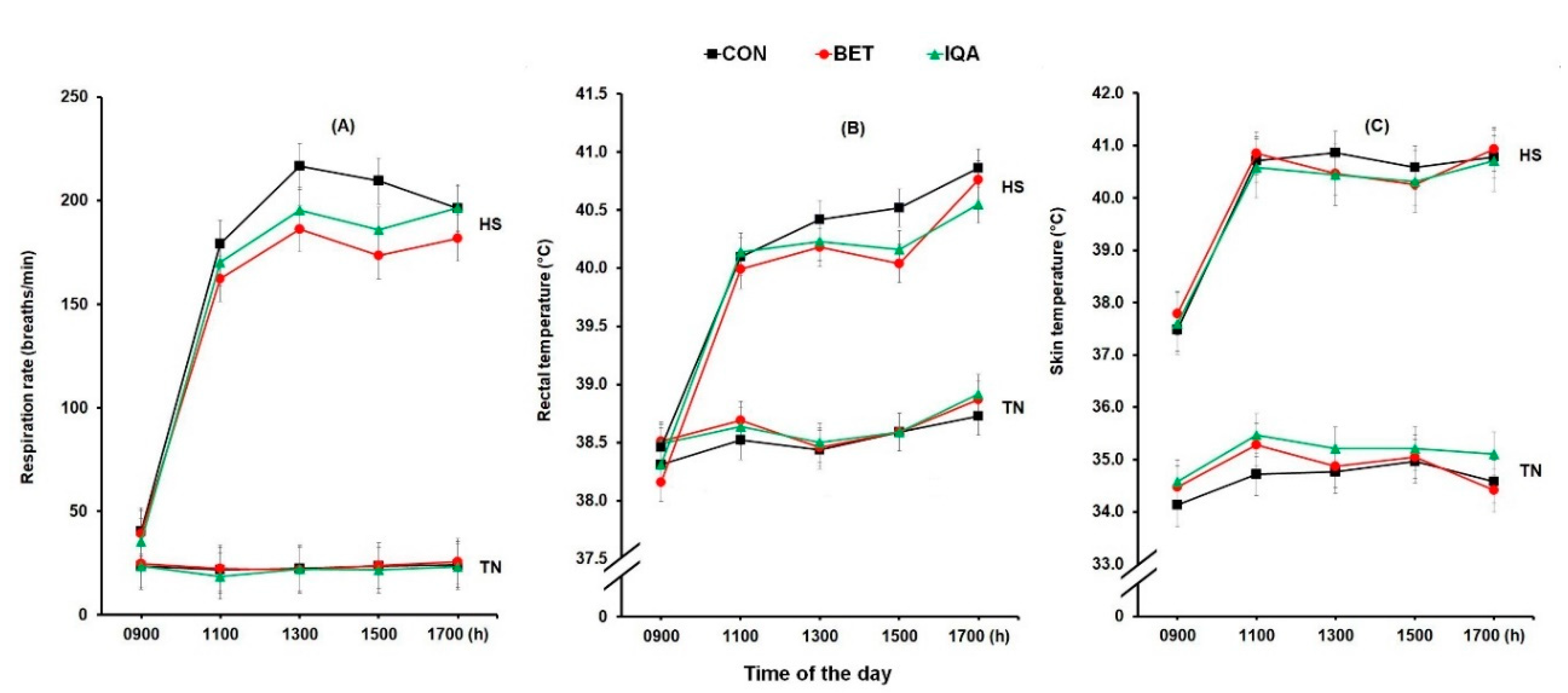
2.2. Physiological Observation
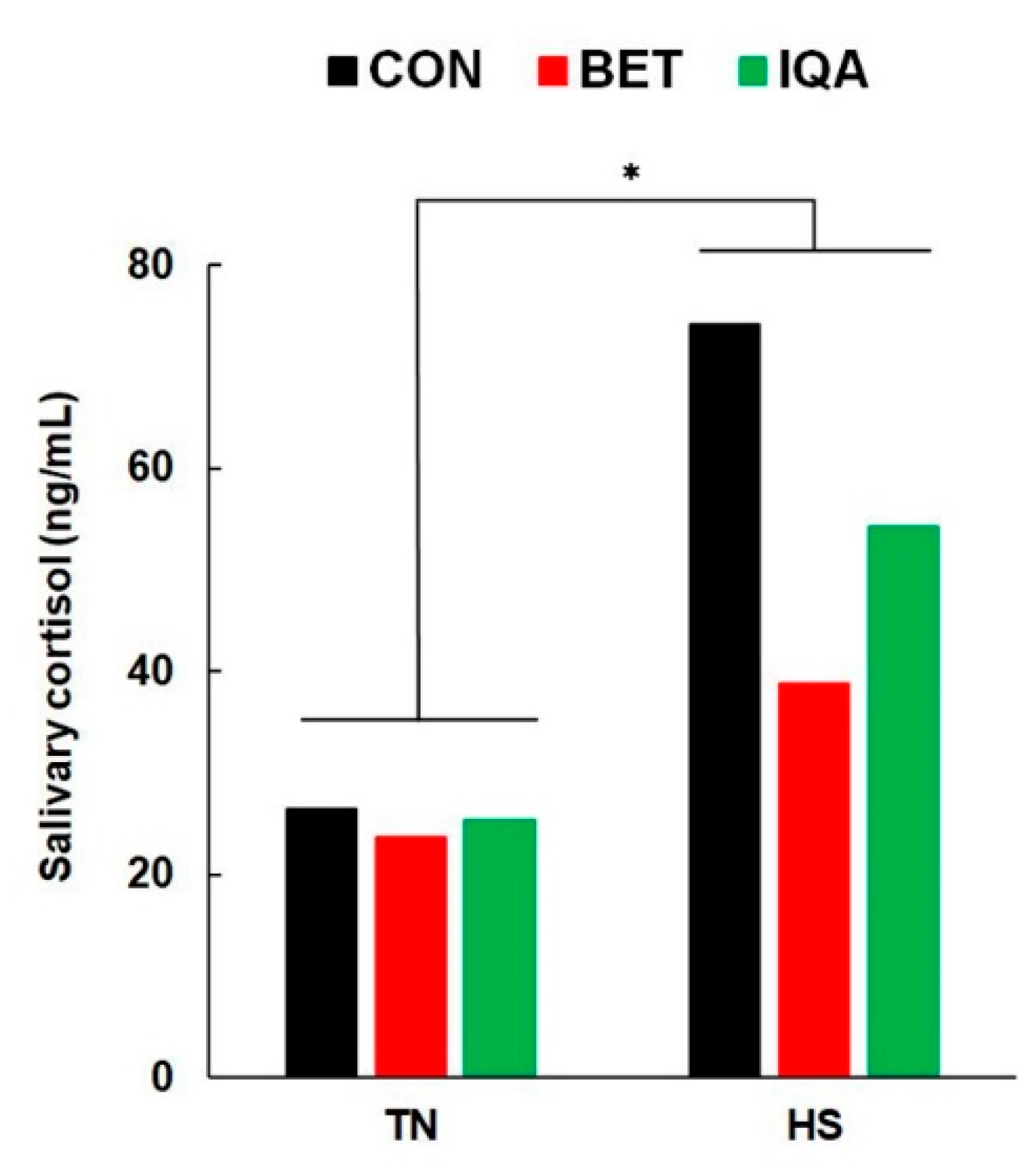
2.3. Saliva Collection and Salivary Cortisol Measurement
2.4. Blood and Tissue Collection, Blood Gas Measurement and Euthanasia
2.5. Plasma Hormones and Metabolites Measurement
2.6. Urinalysis
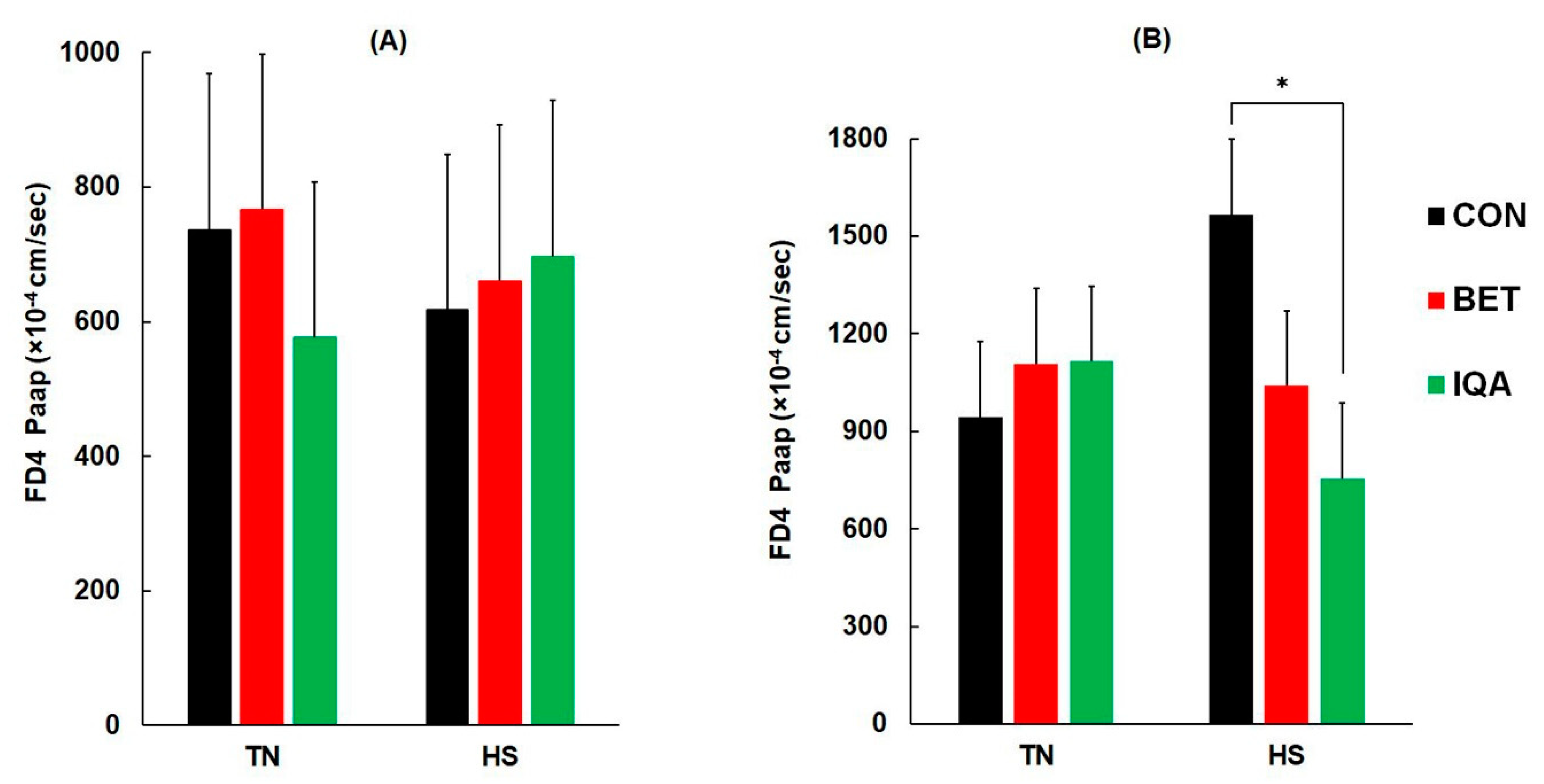
2.7. Tissue Sampling and Intestinal Permeability Measurement
2.8. Measurement of Inflammation Markers by ELISA
2.9. Betaine Distribution
2.10. Test Diet Sanguinarine Concentration
2.11. Statistical Analysis
3. Results
3.1. Physiological Responses
3.2. Blood Oximetry and Electrolytes
3.3. Blood and Urine Biochemistry
3.4. Endocrine Responses to HS
3.5. Inflammatory and Antioxidant Markers
3.6. Intestinal Permeability
3.7. Betaine Distribution
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Renaudeau, D.; Gilbert, H.; Noblet, J. Effect of climatic environment on feed efficiency in swine. In Feed Efficiency in Swine, 2nd ed.; Patience, J.F., Ed.; Wageningen Academic: Wageningen, The Netherlands, 2012; pp. 183–210. [Google Scholar] [CrossRef]
- Bell, A.W.; Hales, J.R.; King, R.B.; Fawcett, A.A. Influence of heat stress on exercise-induced changes in regional blood flow in sheep. J. Appl. Physiol. Respir. Environ. Exerc. Physiol. 1983, 55, 1916–1923. [Google Scholar] [CrossRef] [PubMed]
- Hales, J.R.; Rowell, L.B.; King, R.B. Regional distribution of blood flow in awake heat-stressed baboons. Am. J. Physiol. 1979, 237, H705–H712. [Google Scholar] [CrossRef] [PubMed]
- Wolfenson, D.; Frei, Y.F.; Snapir, N.; Berman, A. Heat stress effects on capillary blood flow and its redistribution in the laying hen. Pflugers Arch. 1981, 390, 86–93. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Yin, P.; Liu, F.; Cheng, G.; Guo, K.; Lu, A.; Zhu, X.; Luan, W.; Xu, J. Effect of heat stress on the porcine small intestine: A morphological and gene expression study. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2010, 156, 119–128. [Google Scholar] [CrossRef] [PubMed]
- Cui, Y.; Gu, X. Proteomic changes of the porcine small intestine in response to chronic heat stress. J. Mol. Endocrinol. 2015, 55, 277–293. [Google Scholar] [CrossRef] [Green Version]
- Pearce, S.C.; Mani, V.; Boddicker, R.L.; Johnson, J.S.; Weber, T.E.; Ross, J.W.; Baumgard, L.H.; Gabler, N.K. Heat stress reduces barrier function and alters intestinal metabolism in growing pigs. J. Anim. Sci. 2012, 90 (Suppl. 4), 257–259. [Google Scholar] [CrossRef]
- Pearce, S.C.; Mani, V.; Boddicker, R.L.; Johnson, J.S.; Weber, T.E.; Ross, J.W.; Rhoads, R.P.; Baumgard, L.H.; Gabler, N.K. Heat stress reduces intestinal barrier integrity and favors intestinal glucose transport in growing pigs. PLoS ONE 2013, 8, e70215. [Google Scholar] [CrossRef]
- Cottrell, J.J.; Liu, F.; Hung, A.T.; DiGiacomo, K.; Chauhan, S.S.; Leury, B.J.; Furness, J.B.; Celi, P.; Dunshea, F.R. Nutritional strategies to alleviate heat stress in pigs. Anim. Prod. Sci. 2015, 55, 1391–1402. [Google Scholar] [CrossRef]
- Moeckel, G.W.; Shadman, R.; Fogel, J.M.; Sadrzadeh, S.M. Organic osmolytes betaine, sorbitol and inositol are potent inhibitors of erythrocyte membrane ATPases. Life Sci. 2002, 71, 2413–2424. [Google Scholar] [CrossRef]
- Shakeri, M.; Cottrell, J.J.; Wilkinson, S.; Ringuet, M.; Furness, J.B.; Dunshea, F.R. Betaine and Antioxidants Improve Growth Performance, Breast Muscle Development and Ameliorate Thermoregulatory Responses to Cyclic Heat Exposure in Broiler Chickens. Animals 2018, 8, 162. [Google Scholar] [CrossRef] [Green Version]
- Shakeri, M.; Cottrell, J.J.; Wilkinson, S.; Le, H.H.; Suleria, H.A.; Warner, R.D.; Dunshea, F.R. Dietary Betaine Reduces the Negative Effects of Cyclic Heat Exposure on Growth Performance, Blood Gas Status and Meat Quality in Broiler Chickens. Agriculture 2020, 10, 176. [Google Scholar] [CrossRef]
- Dunshea, F.R.; Oluboyede, K.; DiGiacomo, K.; Leury, B.J.; Cottrell, J.J. Betaine Improves Milk Yield in Grazing Dairy Cows Supplemented with Concentrates at High Temperatures. Animals 2019, 9, 57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shakeri, M.; Cottrell, J.J.; Wilkinson, S.; Le, H.H.; Suleria, H.A.R.; Warner, R.D.; Dunshea, F.R. Growth Performance and Characterization of Meat Quality of Broiler Chickens Supplemented with Betaine and Antioxidants under Cyclic Heat Stress. Antioxidants 2019, 8, 336. [Google Scholar] [CrossRef] [Green Version]
- Attia, Y.; Abd El-Hamid, A.; Abedalla, A.; Berika, M.; Al-Harthi, M.; Kucuk, O.; Sahin, K.; Abou-Shehema, B. Laying performance, digestibility and plasma hormones in laying hens exposed to chronic heat stress as affected by betaine, vitamin C, and/or vitamin E supplementation. SpringerPlus 2016, 5, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Artuso-Ponte, V.; Moeller, S.; Rajala-Schultz, P.; Medardus, J.J.; Munyalo, J.; Lim, K.; Gebreyes, W.A. Supplementation with Quaternary Benzo(c)phenanthridine Alkaloids Decreased Salivary Cortisol and Salmonella Shedding in Pigs After Transportation to the Slaughterhouse. Foodborne Pathog. Dis. 2015, 12, 891–897. [Google Scholar] [CrossRef]
- Robbins, R.C.; Artuso-Ponte, V.C.; Moeser, A.J.; Morrow, W.M.; Spears, J.W.; Gebreyes, W.A. Effects of quaternary benzo (c) phenanthridine alkaloids on growth performance, shedding of organisms, and gastrointestinal tract integrity in pigs inoculated with multidrug-resistant Salmonella spp. Am. J. Vet. Res. 2013, 74, 1530–1535. [Google Scholar] [CrossRef] [PubMed]
- Tandon, S.; Das, M.; Khanna, S.K. Effect of sanguinarine on the transport of essential nutrients in an everted gut sac model: Role of Na+, K+-ATPase. Nat. Toxins 1993, 1, 235–240. [Google Scholar] [CrossRef]
- Clarke, R.J.; Catauro, M.; Rasmussen, H.H.; Apell, H.-J. Quantitative calculation of the role of the Na+, K+-ATPase in thermogenesis. BBA Bioenerg. 2013, 1827, 1205–1212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.; Kang, B.; Yao, K.; Fu, C.; Zhao, Y. Effects of dietary Macleaya cordata extract on growth performance, immune responses, antioxidant capacity, and intestinal development in weaned piglets. J. Appl. Anim. Res. 2019, 47, 349–356. [Google Scholar] [CrossRef] [Green Version]
- Kosina, P.; Gregorova, J.; Gruz, J.; Vacek, J.; Kolar, M.; Vogel, M.; Roos, W.; Naumann, K.; Simanek, V.; Ulrichova, J. Phytochemical and antimicrobial characterization of Macleaya cordata herb. Fitoterapia 2010, 81, 1006–1012. [Google Scholar] [CrossRef]
- Huynh, T.T.; Aarnink, A.J.; Verstegen, M.W.; Gerrits, W.J.; Heetkamp, M.J.; Kemp, B.; Canh, T.T. Effects of increasing temperatures on physiological changes in pigs at different relative humidities. J. Anim. Sci. 2005, 83, 1385–1396. [Google Scholar] [CrossRef] [PubMed]
- Cottrell, J.J.; Furness, J.B.; Wijesiriwardana, U.A.; Ringuet, M.; Liu, F.; DiGiacomo, K.; Leury, B.J.; Clarke, I.J.; Dunshea, F.R. The Effect of Heat Stress on Respiratory Alkalosis and Insulin Sensitivity in Cinnamon Supplemented Pigs. Animals 2020, 10, 690. [Google Scholar] [CrossRef]
- Tomita, M.; Ohkubo, R.; Hayashi, M. Lipopolysaccharide Transport System across Colonic Epithelial Cells in Normal and Infective Rat. Drug Metab. Pharmacokinet. 2004, 19, 33–40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laryea, M.D.; Steinhagen, F.; Pawliczek, S.; Wendel, U. Simple method for the routine determination of betaine and N,N-dimethylglycine in blood and urine. Clin. Chem. 1998, 44, 1937–1941. [Google Scholar] [CrossRef] [Green Version]
- Xie, H.; Yang, J.; Feng, S.; Cheng, P.; Zeng, J.; Xiong, X. Simultaneous quantitative determination of sanguinarine, chelerythrine, dihydrosanguinarine and dihydrochelerythrine in chicken by HPLC-MS/MS method and its applications to drug residue and pharmacokinetic study. J. Chromatogr. B Analyt. Technol. Biomed. Life Sci. 2015, 985, 124–130. [Google Scholar] [CrossRef] [PubMed]
- Ingram, D. Stimulation of cutaneous glands in the pig. J. Comp. Pathol. 1967, 77, 93–108. [Google Scholar] [CrossRef]
- Hales, J.; Webster, M. Respiratory function during thermal tachypnoea in sheep. J. Physiol. 1967, 190, 241–260. [Google Scholar] [CrossRef] [Green Version]
- Pearce, S.C.; Sanz-Fernandez, M.V.; Hollis, J.H.; Baumgard, L.H.; Gabler, N.K. Short-term exposure to heat stress attenuates appetite and intestinal integrity in growing pigs. J. Anim. Sci. 2014, 92, 5444–5454. [Google Scholar] [CrossRef] [Green Version]
- Shakeri, M.; Cottrell, J.J.; Wilkinson, S.; Zhao, W.; Le, H.H.; McQuade, R.; Furness, J.B.; Dunshea, F.R. Dietary Betaine Improves Intestinal Barrier Function and Ameliorates the Impact of Heat Stress in Multiple Vital Organs as Measured by Evans Blue Dye in Broiler Chickens. Animals 2020, 10, 38. [Google Scholar] [CrossRef] [Green Version]
- Vrublova, E.; Vostalova, J.; Ehrmann, J.; Palikova, I.; Vrbkova, J.; Vacek, J.; Cibicek, N.; Vecera, R.; Ulrichova, J.; Simanek, V. The phytogenic feed additive Sangrovit modulates dextran sulfate sodium-induced colitis in rats. Vet. Med. Praha 2010, 55, 610–618. [Google Scholar] [CrossRef] [Green Version]
- Liu, G.; Guan, G.; Fang, J.; Martínez, Y.; Chen, S.; Bin, P.; Duraipandiyan, V.; Gong, T.; Tossou, M.C.B.; Al-Dhabi, N.A.; et al. Macleaya cordata Extract Decreased Diarrhea Score and Enhanced Intestinal Barrier Function in Growing Piglets. Biomed. Res. Int. 2016, 2016, 1069585. [Google Scholar] [CrossRef] [Green Version]
- Huang, H.; Yao, J.; Liu, K.; Yang, W.; Wang, G.; Shi, C.; Jiang, Y.; Wang, J.; Kang, Y.; Wang, D. Sanguinarine has anthelmintic activity against the enteral and parenteral phases of trichinella infection in experimentally infected mice. Acta Trop. 2020, 201, 105226. [Google Scholar] [CrossRef] [PubMed]
- Lima, T.R.F.; Gallo, S.B.; Rosa, A.F.; Brochado, T.; Bezerra, H.V.A.; Putrino, S.M.; Martins, M.B.; Leme, P.R. Effect of Macleaya cordata and Magnolia officinalis plant extracts on oxidative stress control in lambs fed a high-concentrate diet. Asian australas. J. Anim. Sci. 2019. [Google Scholar] [CrossRef] [PubMed]
- Estrada-Angulo, A.; Aguilar-Hernández, A.; Osuna-Pérez, M.; Núñez-Benítez, V.; Castro-Pérez, B.; Silva-Hidalgo, G.; Contreras-Pérez, G.; Barreras, A.; Plascencia, A.; Zinn, R. Influence of quaternary benzophenantridine and protopine alkaloids on growth performance, dietary energy, carcass traits, visceral mass, and rumen health in finishing ewes under conditions of severe temperature-humidity index. Asian-Australas. J. Anim. Sci. 2016, 29, 652–658. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Li, H.; Yao, H.; Mu, Q.; Zhao, G.; Li, Y.; Hu, H.; Niu, X. Pharmacokinetic and anti-inflammatory effects of sanguinarine solid lipid nanoparticles. Inflammation 2014, 37, 632–638. [Google Scholar] [CrossRef] [PubMed]
- Khadem, A.; Soler, L.; Everaert, N.; Niewold, T.A. Growth promotion in broilers by both oxytetracycline and Macleaya cordata extract is based on their anti-inflammatory properties. Br. J. Nutr. 2014, 112, 1110–1118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kosina, P.; Walterova, D.; Ulrichova, J.; Lichnovsky, V.; Stiborova, M.; Rydlova, H.; Vicar, J.; Krecman, V.; Brabec, M.J.; Simanek, V. Sanguinarine and chelerythrine: Assessment of safety on pigs in ninety days feeding experiment. Food Chem. Toxicol. 2004, 42, 85–91. [Google Scholar] [CrossRef]
- Večeřa, R.; Klejdus, B.; Kosina, P.; Orolin, J.; Stiborova, M.; Smrček, S.; Vičar, J.; Dvořák, Z.; Ulrichova, J.; Kubáň, V. Disposition of sanguinarine in the rat. Xenobiotica 2007, 37, 549–558. [Google Scholar] [CrossRef]
- Wu, Y.; Zhao, N.J.; Cao, Y.; Sun, Z.; Wang, Q.; Liu, Z.Y.; Sun, Z.L. Sanguinarine metabolism and pharmacokinetics study in vitro and in vivo. J. Vet. Pharmacol. Ther. 2020, 43, 208–214. [Google Scholar] [CrossRef]
- Tandon, S.; Das, M.; Khanna, S.K. Biometabolic elimination and organ retention profile of argemone alkaloid, sanguinarine, in rats and guinea pigs. Drug Metab. Dispos. 1993, 21, 194–197. [Google Scholar]
- Straub, K.D.; Carver, P. Sanguinarine, inhibitor of Na-K dependent ATP’ase. Biochem. Biophys. Res. Commun. 1975, 62, 913–922. [Google Scholar] [CrossRef]
- Jankowski, J.; Zduńczyk, Z.; Juśkiewicz, J.; Kozłowski, K.; Lecewicz, A.; Jeroch, H. Gastrointestinal tract and metabolic response of broilers to diets with the Macleaya cordata alkaloid extract. Arch. Geflugelkd. 2009, 73, 95–101. [Google Scholar]
- DiGiacomo, K.; Simpson, S.; Leury, B.J.; Dunshea, F.R. Dietary betaine impacts the physiological responses to moderate heat conditions in a dose dependent manner in sheep. Animals 2016, 6, 51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McKeever, M.; Weir, D.; Molloy, A.; Scott, J. Betaine-homocysteine methyltransferase: Organ distribution in man, pig and rat and subcellular distribution in the rat. Clin. Sci. 1991, 81, 551–556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kettunen, H.; Peuranen, S.; Tiihonen, K.; Saarinen, M. Intestinal uptake of betaine in vitro and the distribution of methyl groups from betaine, choline, and methionine in the body of broiler chicks. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2001, 128, 269–278. [Google Scholar] [CrossRef]
- Schwahn, B.C.; Hafner, D.; Hohlfeld, T.; Balkenhol, N.; Laryea, M.D.; Wendel, U. Pharmacokinetics of oral betaine in healthy subjects and patients with homocystinuria. Br. J. Clin. Pharmacol. 2003, 55, 6–13. [Google Scholar] [CrossRef] [Green Version]
- Gabler, N.; Frouel, S.; Awati, A.; Owusu-Asiedu, A.; Amerah, A.; Patridge, G.; Dunshea, F. Betaine mitigates intestinal permeability in growing pigs induced by heat stress. In Manipulating Pig Production XIV, Proceeding of the Australasian Pig Science Association (APSA), Melbourne, Australia, 24–27 November 2013; Pluske, J., Pluske, J.R., Eds.; Australian Pig Science Association: Melbourne, Australia, 2013; p. 85. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Item | Amount |
|---|---|
| Ingredient, % | |
| Wheat | 50.5 |
| Barley | 7.00 |
| Lupins | 15.0 |
| Millmix 1 | 4.00 |
| Canola meal 38% | 13.8 |
| Soy bean meal 46% | 5.00 |
| Water | 2.00 |
| Tallow mixer | 0.800 |
| Limestone | 1.53 |
| DL-Methionine | 0.060 |
| Lysine | 0.030 |
| Threonine | 0.055 |
| Calculated values | |
| Digestible energy (MJ/kg) | 13.4 |
| Crude protein (%) | 19.0 |
| Lysine (%) | 1.09 |
| Calcium (%) | 0.887 |
| Total phosphorous (%) | 0.533 |
| Parameter 2 | TN | HS | SED | p-Value | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| CON | BET | IQA | CON | BET | IQA | T | D | T × D | ||
| pH | 7.44 | 7.45 | 7.46 | 7.44 | 7.43 | 7.43 | 0.01 | 0.013 | 0.89 | 0.21 |
| pO2 (mmHg) | 66.8 | 68.3 | 66.8 | 50.2 | 44.6 | 43.9 | 3.13 | <0.001 | 0.50 | 0.16 |
| O2 saturation (%) | 93.0 a | 93.7 a | 93.3 a | 85.2 b | 81.9 bc | 78.2 c | 2.01 | <0.001 | 0.096 | 0.035 |
| PCO2 (mmHg) | 48.1 | 46.9 | 47.9 | 49.2 | 46.4 | 48.5 | 1.62 | 0.35 | 0.096 | 0.56 |
| Total CO2 (mM) | 34.6 | 34.3 | 35.4 | 34.6 | 32.9 | 33.3 | 1.10 | 0.074 | 0.43 | 0.37 |
| HCO3 (mM) | 33.1 | 32.9 | 33.9 | 33.1 | 31.5 | 31.8 | 1.05 | 0.054 | 0.44 | 0.35 |
| Sodium (mM) | 142 | 143 | 142 | 143 | 141 | 141 | 1.69 | 0.59 | 0.64 | 0.59 |
| Potassium (mM) | 3.95 | 4.17 | 4.04 | 4.38 | 4.26 | 4.26 | 0.13 | 0.001 | 0.77 | 0.15 |
| Chloride (mM) | 98.2 | 98.0 | 99.1 | 100 | 101 | 99.5 | 1.53 | 0.027 | 0.86 | 0.35 |
| Calcium (mM) | 1.37 | 1.38 | 1.37 | 1.36 | 1.35 | 1.40 | 0.03 | 0.92 | 0.51 | 0.40 |
| Anion gap (mM) | 14.8 | 15.9 | 14.6 | 14.6 | 14.8 | 14.4 | 0.72 | 0.23 | 0.21 | 0.64 |
| Glucose (mM) | 6.23 | 6.58 | 6.26 | 6.52 | 5.96 | 6.81 | 0.39 | 0.66 | 0.68 | 0.10 |
| Lactate (mM) | 1.92 | 1.86 | 1.87 | 1.08 | 1.27 | 1.12 | 0.33 | 0.001 | 0.96 | 0.86 |
| BEb (mM) | 7.83 | 7.89 | 8.76 | 8.29 | 6.23 | 6.38 | 0.93 | 0.029 | 0.32 | 0.088 |
| BEecf (mM) | 9.02 | 9.01 | 10.0 | 9.54 | 7.14 | 7.35 | 1.06 | 0.032 | 0.29 | 0.098 |
| Hwt (%) | 35.4 | 35.1 | 35.3 | 33.2 | 30.7 | 31.2 | 1.34 | <0.001 | 0.31 | 0.45 |
| HgB (g/dL) | 11.8 | 11.9 | 12.0 | 11.3 | 10.5 | 10.6 | 0.44 | <0.001 | 0.50 | 0.25 |
| Parameter | TN | HS | SED | p-Value | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| CON | BET | IQA | CON | BET | IQA | T | D | T × D | ||
| Protein (mg/mL) | 51.3 | 54.4 | 54.1 | 55.7 | 53.7 | 53.2 | 3.60 | 0.59 | 0.98 | 0.47 |
| Albumin (g/L) | 43.0 | 39.5 | 39.9 | 46.0 | 40.5 | 40.4 | 2.87 | 0.33 | 0.048 | 0.78 |
| Bilirubin (µM) | 6.01 | 8.37 | 6.97 | 7.01 | 7.47 | 5.72 | 1.22 | 0.64 | 0.15 | 0.37 |
| Creatinine (mM) | 0.130 | 0.129 | 0.118 | 0.148 | 0.139 | 0.148 | 0.01 | <0.001 | 0.45 | 0.16 |
| Free T3 (pg/mL) | 0.71 | 0.68 | 0.58 | 0.45 | 0.48 | 0.50 | 0.07 | <0.001 | 0.68 | 0.35 |
| Free T4 (pg/mL) | 17.0 | 15.8 | 13.2 | 10.1 | 9.28 | 9.22 | 1.38 | <0.001 | 0.03 | 0.26 |
| Parameter | TN | HS | SED | p-Value | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| CON | BET | IQA | CON | BET | IQA | T | D | T × D | ||
| pH | 5.95 | 6.44 | 6.50 | 6.39 | 6.23 | 6.41 | 0.24 | 0.67 | 0.29 | 0.16 |
| Osmolality (mOsm/kg H2O) | 130 | 210 | 154 | 406 | 227 | 338 | 98 | 0.005 | 0.78 | 0.18 |
| Albumin (g/L) | 0.352 | 0.404 | 0.451 | 0.579 | 0.627 | 0.830 | 0.109 | <0.001 | 0.11 | 0.53 |
| Bilirubin (µM) | 5.78 a | 7.91 ab | 10.9 abc | 17.7 bc | 21.0 c | 5.65 a | 5.16 | 0.014 | 0.33 | 0.030 |
| Creatinine (mM) | 1.85 | 3.69 | 4.93 | 9.49 | 10.3 | 4.92 | 2.73 | 0.002 | 0.63 | 0.15 |
| Parameter | TN | HS | SED | p-Value | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| CON | BET | IQA | CON | BET | IQA | T | D | T × D | ||
| Jejunum | ||||||||||
| IL-1β (pg/mL) | 130 | 115 | 130 | 173 | 253 | 154 | 34.8 | 0.002 | 0.21 | 0.056 |
| IL-8 (pg/mL) | 2965 | 3301 | 3026 | 3470 | 3044 | 3011 | 0.297 | 0.58 | 0.62 | 0.18 |
| TNF-α (pg/mL) | 28.2 | 32.0 | 30.2 | 26.6 | 24.3 | 27.8 | 4.38 | 0.14 | 0.87 | 0.57 |
| Ileum | ||||||||||
| IL-1β (pg/mL) | 228 | 343 | 334 | 402 | 431 | 408 | 62.8 | 0.004 | 0.25 | 0.46 |
| IL-8 (pg/mL) | 2611 | 2706 | 2604 | 2983 | 3111 | 3247 | 310 | 0.012 | 0.81 | 0.80 |
| TNF-α (pg/mL) | 36.3 | 35.3 | 33.2 | 34.0 | 39.4 | 41.2 | 4.78 | 0.29 | 0.75 | 0.30 |
| ESR (mm/h) | 8.3 | 8.7 | 7.8 | 12.2 | 14.1 | 11.3 | 2.10 | 0.001 | 0.45 | 0.83 |
| TAC (mM Trolox) | 1.34 | 1.61 | 1.17 | 1.38 | 1.63 | 1.21 | 0.20 | 0.73 | 0.0205 | 1.00 |
| Parameter | TN | HS | SED | p-Value | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| CON | BET | IQA | CON | BET | IQA | T | D | T × D | ||
| Plasma (µmol/L) | 731 | 791 | 1271 | 949 | 831 | 1293 | 98.4 | 0.088 | <0.001 | 0.28 |
| Urine 2 | 3.23 | 3.69 | 3.20 | 3.33 | 3.86 | 3.19 | 0.22 | 0.50 | 0.002 | 0.83 |
| (µmol/L) | (1714) | (4853) | (1600) | (2128) | (7311) | (1538) | ||||
| Tissue (mg/g) | ||||||||||
| Muscle | 0.50 | 0.74 | 0.51 | 0.54 | 0.74 | 0.65 | 0.14 | 0.50 | 0.084 | 0.80 |
| Liver | 0.94 | 1.54 | 1.14 | 1.38 | 1.56 | 1.22 | 0.18 | 0.070 | 0.007 | 0.20 |
| Kidney | 0.93 | 1.19 | 0.83 | 1.06 | 1.05 | 0.61 | 0.14 | 0.42 | <0.001 | 0.21 |
| Jejunum | 0.34 | 0.45 | 0.36 | 0.40 | 0.52 | 0.33 | 0.12 | 0.15 | <0.001 | 0.19 |
| Ileum | 0.45 | 0.61 | 0.48 | 0.49 | 0.51 | 0.43 | 0.04 | 0.28 | <0.001 | 0.097 |
| Colon | 0.50 | 0.48 | 0.36 | 0.43 | 0.55 | 0.52 | 0.07 | 0.51 | 0.66 | 0.12 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Le, H.H.; Shakeri, M.; Suleria, H.A.R.; Zhao, W.; McQuade, R.M.; Phillips, D.J.; Vidacs, E.; Furness, J.B.; Dunshea, F.R.; Artuso-Ponte, V.; et al. Betaine and Isoquinoline Alkaloids Protect against Heat Stress and Colonic Permeability in Growing Pigs. Antioxidants 2020, 9, 1024. https://doi.org/10.3390/antiox9101024
Le HH, Shakeri M, Suleria HAR, Zhao W, McQuade RM, Phillips DJ, Vidacs E, Furness JB, Dunshea FR, Artuso-Ponte V, et al. Betaine and Isoquinoline Alkaloids Protect against Heat Stress and Colonic Permeability in Growing Pigs. Antioxidants. 2020; 9(10):1024. https://doi.org/10.3390/antiox9101024
Chicago/Turabian StyleLe, Hieu Huu, Majid Shakeri, Hafiz Ansar Rasul Suleria, Weicheng Zhao, Rachel Mai McQuade, Deborah Jayne Phillips, Eva Vidacs, John Barton Furness, Frank Rowland Dunshea, Valeria Artuso-Ponte, and et al. 2020. "Betaine and Isoquinoline Alkaloids Protect against Heat Stress and Colonic Permeability in Growing Pigs" Antioxidants 9, no. 10: 1024. https://doi.org/10.3390/antiox9101024