
Evaluation of Classical Mathematical Models of Tumor Growth Using an On-Lattice Agent-Based Monte Carlo Model †

 ,
,
Abstract
:Featured Application
Abstract
1. Introduction
2. Models
2.1. Classical Mathematical Models of Tumor Growth
2.2. On-Lattice Monte Carlo Agent-Based Model
2.3. Statistical Methods
3. Results and Discussion
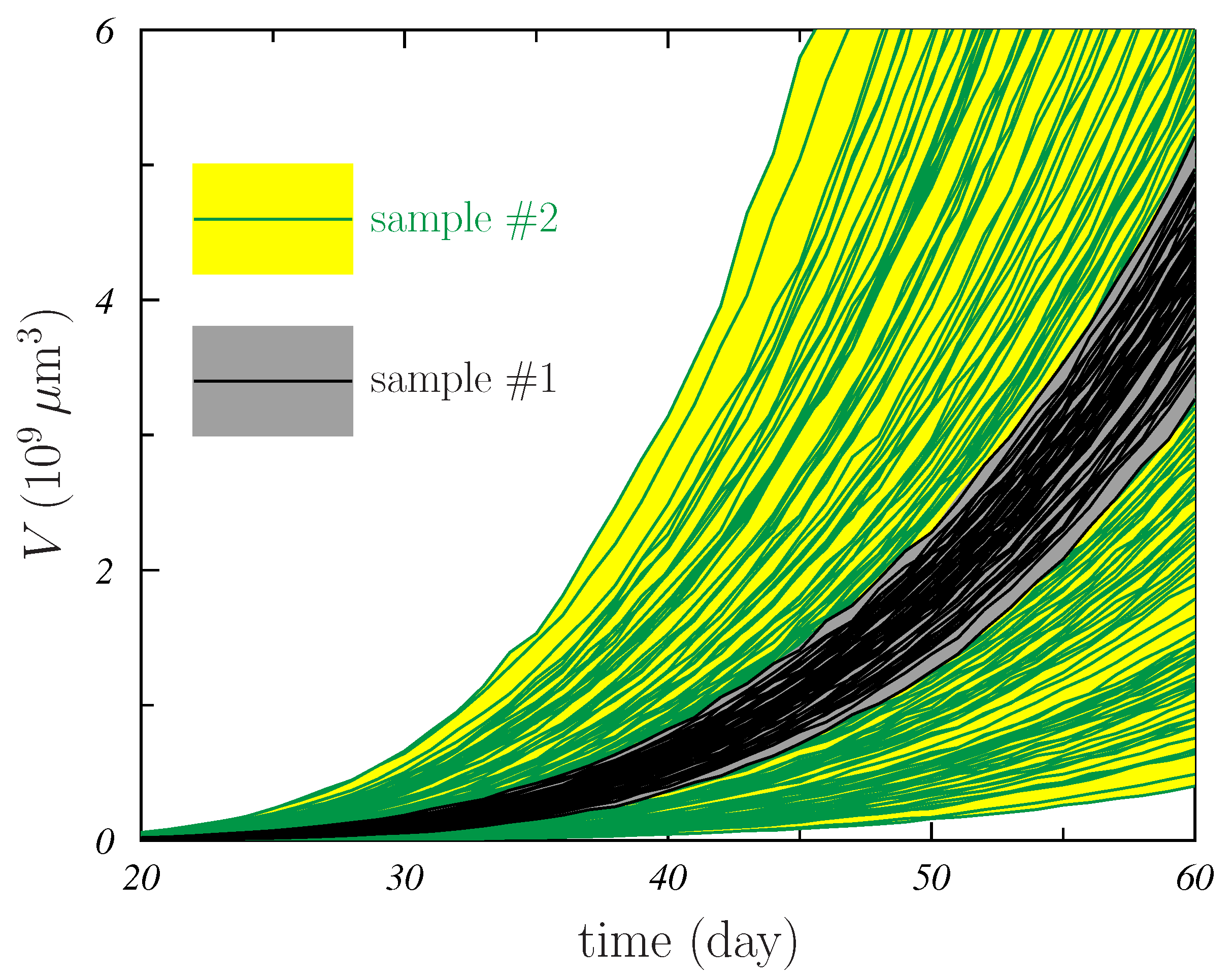
- For all models, the distributions corresponding to sample #2 were wider than those of sample #1. This is seen in Figure 4 and also in Table 1. The sizes of the 95% CI quoted in the table for sample #2 are larger than those of sample #1 by factors between ∼2 for the G-model and ∼5 for the P-model. In addition, the found for sample #2 is larger than the corresponding for sample #1, the increase ranging between and for the G- and B-models, respectively.The variability in the AMB parameters that was considered in the simulation generating sample #2 produces more different growth shapes of the MTS and this results in better (those with smaller values) and worst (those with larger ) fits occuring independently of the specific mathematical model considered.
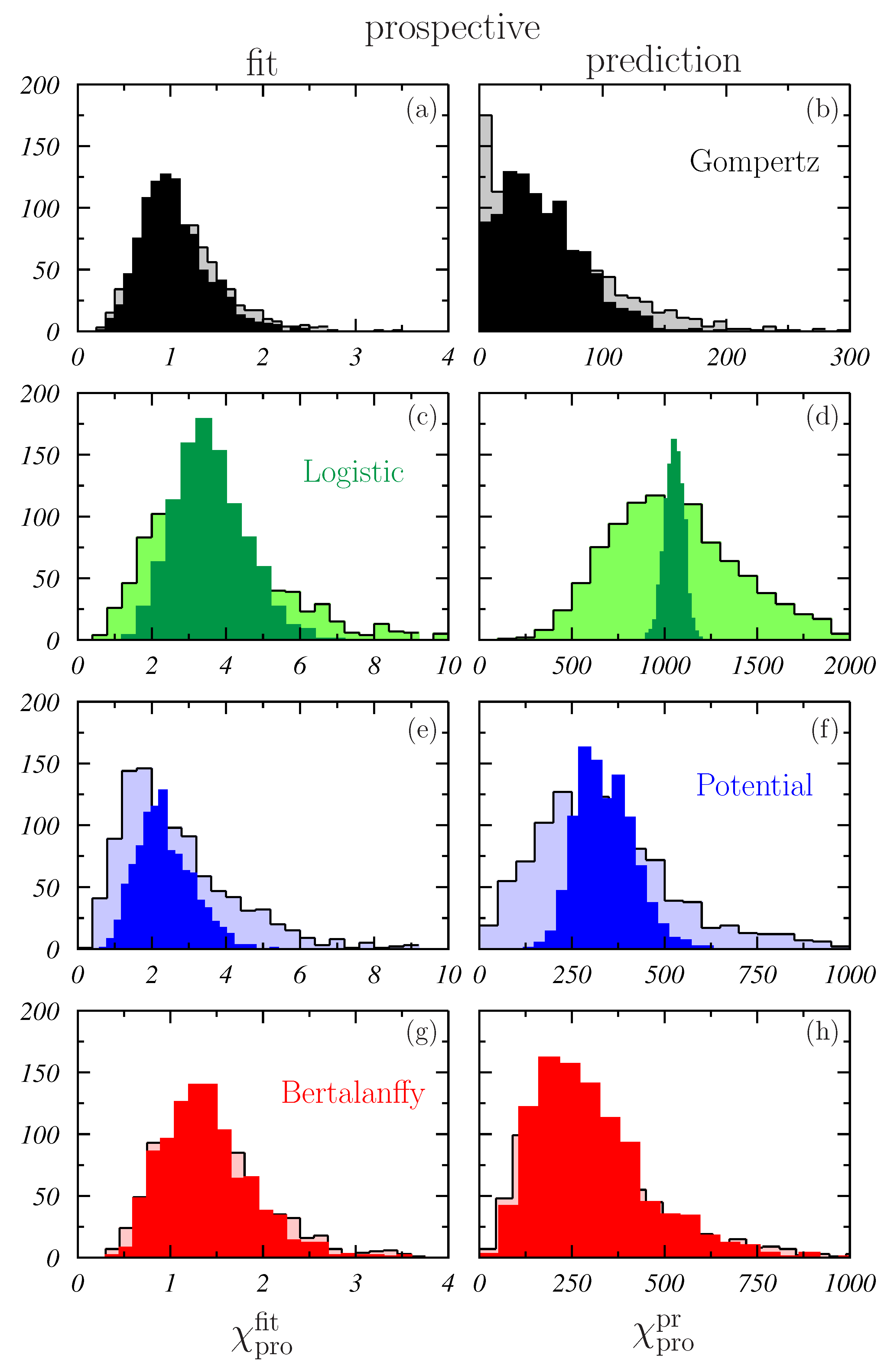
- In general, the best fits to the whole growth curves were provided by the G-model. As shown in Table 1, the obtained for this model are an order of magnitude smaller than those corresponding to the other models considered. In the case of sample#1, the 95% CI obtained for the G-model is clearly below those found for the other models. For sample #2, there is a slight overlap between the CI of the G- and P-models.
- It is noticeable the large values obtained for the L-model. The values are above 170 in both samples, and the respective 95% CI begin at 139 and 56, respectively. This again points out the fact already discussed in connection to Figure 2 about the difficulties of this model to describe the whole MTS growth.
- A curiosity of these results is that all the distributions are skewed to the right (to high ) except that found for the P-model in the case of sample #1 where an almost Gaussian behavior is observed.
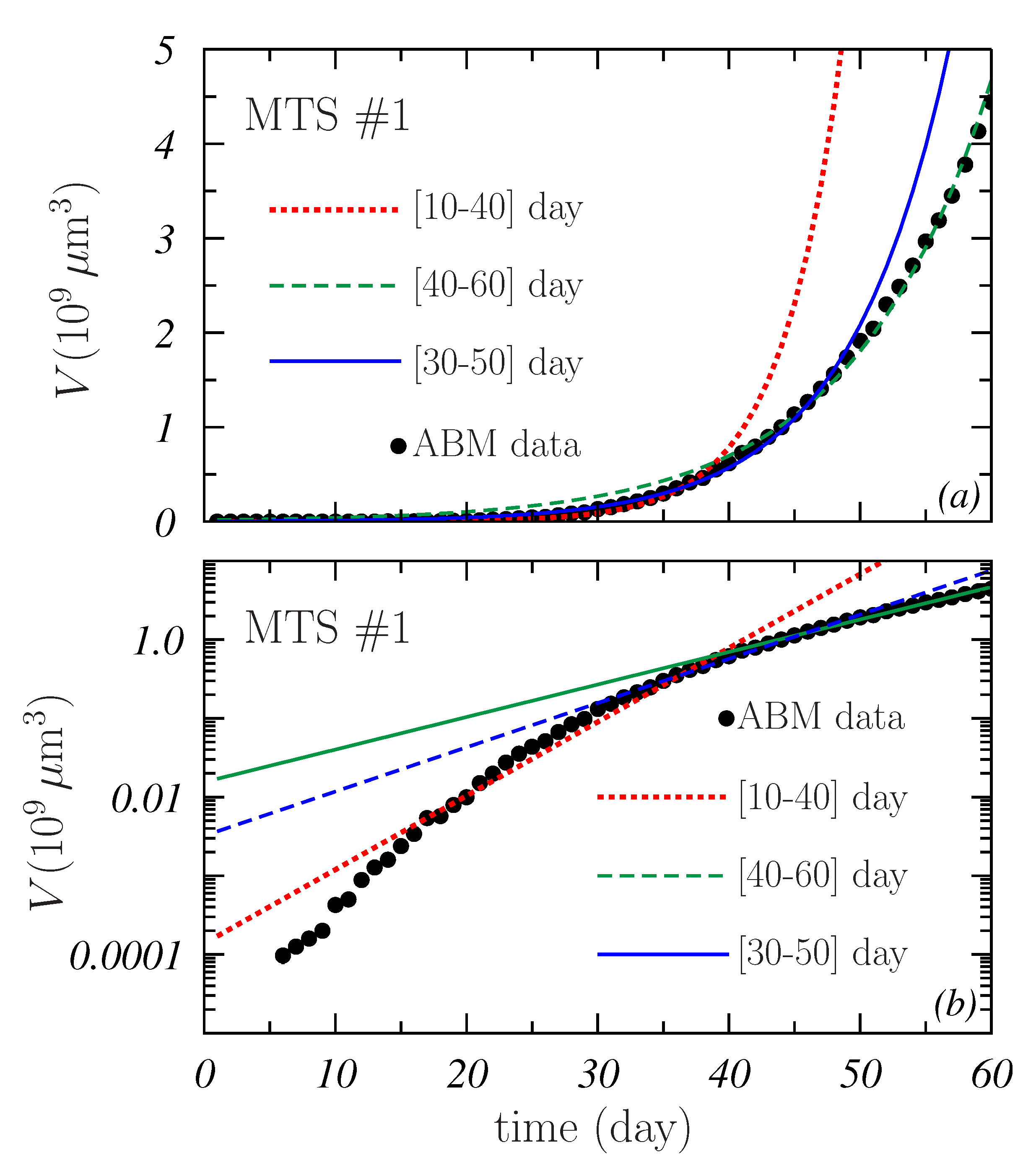
- The best fits were now much better than those found when all volume data were included. The maximum was , which corresponds to the L-model. These relatively low values are due to the fact that the number of data fitted is now 21 instead of the 55 considered before.It is worth noting that also the low limits of the 95% CI of were much lower than in the case of . In all cases, the values obtained are close to 1, with the only exception of the 1.97 found for the L-model in the prospective prediction (see Table 1). This means that, a priori, all the mathematical models could produce a nice description of a more or less short data section of a given spheroid. However, as pointed out in Figure 4, not all the models can describe the whole growth curve of some of the simulated MTSs included in the samples. In this respect, it is important to point out that, in practice, these mathematical models are fitted to a small number of experimental volume data and for few spheroids or tumors in the sample, and the model that better fits the existing data may not be the one that best describes the subsequent growth [34].
- Contrary to what was observed in Figure 4, the distributions obtained for sample #2 are rather similar to those obtained for sample #1. This is corroborated by the results quoted in Table 1 and may indicate that the main ingredient controlling the results of the fit is the number of points to be fitted.An exception to this seems to occur for L- and P-models in the case of the prospective fits (see Figure 6c,e). The possible effect due the region of data fitted is discussed below.
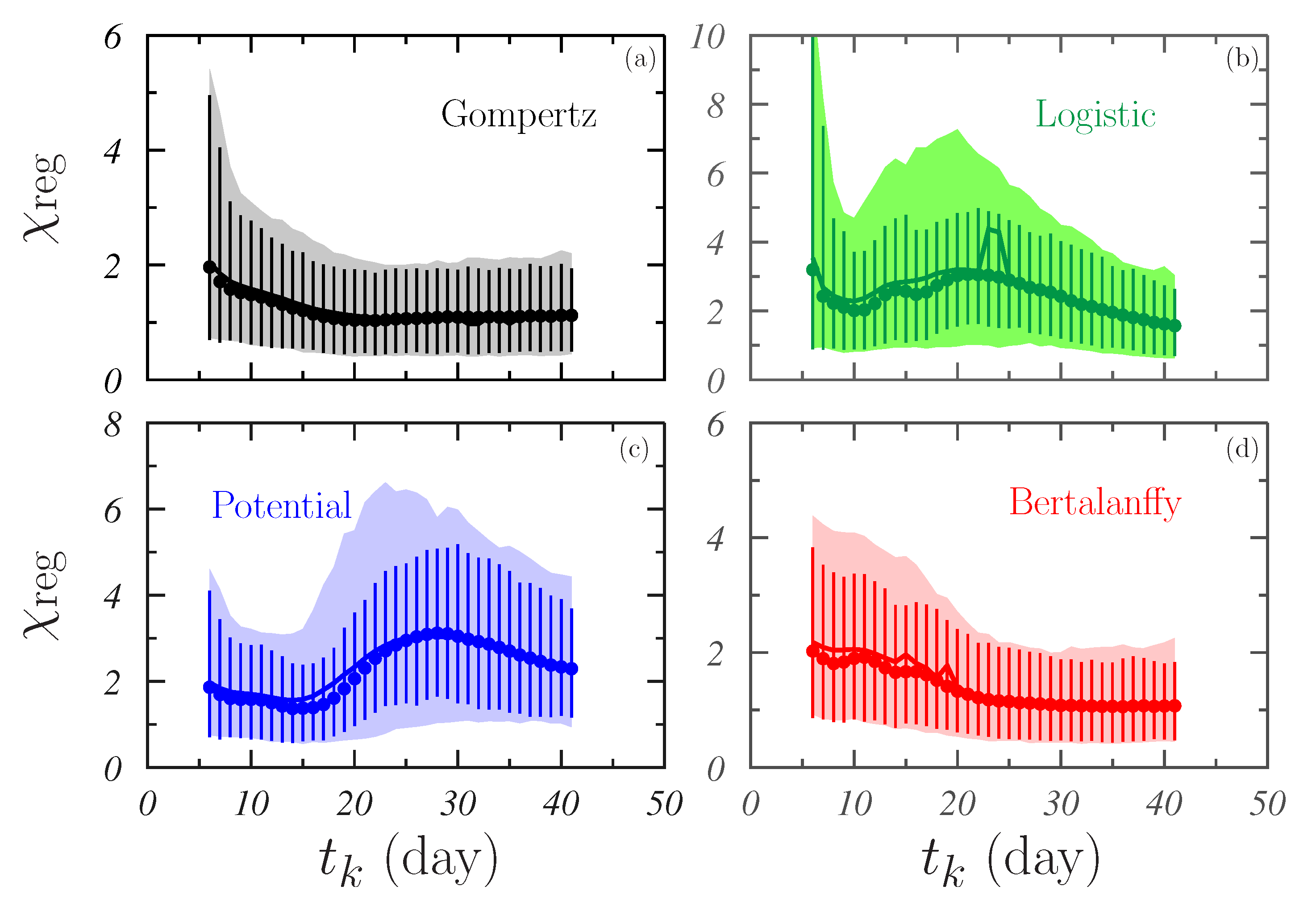
- As can be checked by comparing the right panels in Figure 5 and Figure 6, the models are more efficient in the retrospective predictions than in the prospective predictions. As it can be seen, the distributions obtained in the latter case extend to values that are at least twice as large as those resulting for the former. The worst situation is that of the B-model in which the ratio . This ratio is ∼2 for G- and L-models and ∼5 for the P-model.The retrospective predictive power of the models studied may be of interest in the clinic when the response of tumors to treatments is analyzed. In fact, the evolution of the regression of the irradiated tumors could be similar to looking back at the tumor growth and what has been observed in the present analysis could provide significant hints in that respect. Obviously, treatments may perturb the growth trends and more specific conclusions could be drawn when irradiation is included in the ABM simulation algorithm. Work in this direction is in progress.
- The best results in what refer to retrospective predictions were provided by the B-model for which the average (see Table 1). A slightly larger value, ∼30, was found for the G-model, whereas, for the other two models, much higher values are achieved, in particular for the L-model where .
- The G-model seems to be the most robust in what refers to the prospective predictive capability. In this case, (slightly larger for sample #2 than for sample #1). For P- and B-models, this average was ∼300, while, for the L-model, values larger than 1000 were found.
- In any case, the values obtained for both and indicate that the predictive capabilities of the models considered here are discrete.
- Similar results were found for G- and B-models that showed average values tending to 1 with increasing . The former appeared to be slightly robust in this sense, showing a less noising behavior for day.
- For L- and P-models, a non-negligible dependence on the data region fitted occurred. It is worth pointing out the fluctuating behavior with observed in both cases.
- This also results in the differences between the two samples studied being larger for L- and P-models than for G- and B-models.
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| ABM | agent-based model |
| MTS | multicellular tumor spheroid |
References
- Sutherland, R.M.; Sordat, B.; Bamat, J.; Gabbert, H.; Bourrat, B.; Mueller-Klieser, W. Oxygenation and differentiation in multicellular spheroids of human colon carcinoma. Cancer Res. 1986, 46, 5320–5329. [Google Scholar] [PubMed]
- Cui, X.; Hartanto, Y.; Zhang, H. Advances in multicellular spheroids formation. J. R. Soc. Interface 2017, 14, 20160877. [Google Scholar] [CrossRef]
- Oktem, G.; Vatansever, S.; Ayla, S.; Uysal, A.; Aktas, S.; Karabulut, B.; Bilir, A. Effect of apoptosis and response of extracellular matrix proteins after chemotherapy application on human breast cancer cell spheroids. Oncol. Rep. 2006, 15, 335–340. [Google Scholar] [CrossRef] [PubMed]
- Pampaloni, F.; Reynaud, E.G.; Stelzer, E.H. The third dimension bridges the gap between cell culture and live tissue. Nat. Rev. Mol. Cell Biol. 2007, 8, 839–845. [Google Scholar] [CrossRef] [PubMed]
- Friedrich, J.; Seidel, C.; Ebner, R.; Kunz-Schughart, L.A. Spheroid-based drug screen: Considerations and practical approach. Nat. Protoc. 2009, 4, 309–324. [Google Scholar] [CrossRef]
- Labarbera, D.V.; Reid, B.G.; Yoo, B.H. The multicellular tumor spheroid model for high-throughput cancer drug discovery. Expert Opin. Drug Dis. 2012, 7, 819–830. [Google Scholar] [CrossRef]
- Pacheco-Marín, R.; Melendez-Zajgla, J.; Castillo-Rojas, G.; Mandujano-Tinoco, E.; Garcia-Venzor, A.; Uribe-Carvajal, S.; Cabrera-Orefice, A.; Gonzalez-Torres, C.; Gaytan-Cervantes, J.; Mitre-Aguilar, I.B.; et al. Transcriptome profile of the early stages of breast cancer tumoral spheroids. Sci. Rep. 2016, 6, 23373. [Google Scholar] [CrossRef] [Green Version]
- Mueller-Klieser, W. Three-dimensional cell cultures: From molecular mechanisms to clinical applications. Am. J. Physiol.-Cell Physiol. 1997, 273, C1109–C1123. [Google Scholar] [CrossRef]
- Bates, R.C.; Edwards, N.S.; Yates, J.D. Spheroids and cell survival. Crit. Rev. Oncol. Hematol. 2000, 36, 61–74. [Google Scholar] [CrossRef]
- Mueller-Klieser, W. Tumor biology and experimental therapeutics. Crit. Rev. Oncol. Hematol. 2000, 36, 123–139. [Google Scholar] [CrossRef]
- Kunz-Schughart, L.A.; Freyer, J.P.; Hofstaedter, F.; Ebner, R. The use of 3D cultures for high-throughput screening: The multicellular spheroid model. J. Biomol. Screen. 2004, 9, 273–285. [Google Scholar] [CrossRef] [Green Version]
- Conger, A.D.; Ziskin, M.C. Growth of mammalian multicellular tumor spheroids. Cancer Res. 1983, 43, 556–560. [Google Scholar]
- Freyer, J.P.; Sutherland, R.M. Selective dissociation and characterization of cells from different regions of multicell tumor spheroids. Cancer Res. 1980, 40, 3956–3965. [Google Scholar]
- Guirado, D.; Aranda, M.; Vilches, M.; Villalobos, M.; Lallena, A.M. Dose dependence of the growth rate of multicellular tumour spheroids after irradiation. Br. J. Radiol. 2003, 76, 109–116. [Google Scholar] [CrossRef]
- Guirado, D. Variabilidad en Radiobiología. Ph.D. Thesis, University of Granada, Granada, Spain, 2012. [Google Scholar]
- Wang, C.; Tang, Z.; Zhao, Y.; Yao, R.; Li, L.; Sun, W. Three-dimensional in vitro cancer models: A short review. Biofabrication 2014, 6, 022001. [Google Scholar] [CrossRef]
- McMillan, K.S.; McCluskey, A.G.; Sorensen, A.; Boyd, M.; Zagnoni, M. Emulsion technologies for multicellular tumour spheroid radiation assays. Analyst 2016, 141, 100–110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, Y.; Kim, J.S.; Kim, S.H.; Park, Y.K.; Yu, E.; Kim, K.H.; Seo, E.J.; Oh, H.B.; Lee, H.C.; Kim, K.M.; et al. Patient-derived multicellular tumor spheroids towards optimized treatment for patients with hepatocellular carcinoma. J. Exp. Clin. Cancer Res. 2018, 37, 109. [Google Scholar] [CrossRef]
- Sosa, V.I.; Theys, J.; Groot, A.J.; Barbeau, L.M.O.; Lemmens, A.; Yaromina, A.; Losen, M.; Houben, R.; Dubois, L.; Vooijs, M. Synergistic effects of NOTCH/γ-secretase inhibition and standard of care treatment modalities in non-small cell lung cancer cells. Front. Oncol. 2018, 8, 460. [Google Scholar] [CrossRef]
- Verjans, E.-T.; Doijen, J.; Luyten, W.; Landuyt, B.; Schoofs, L. Three-dimensional cell culture models for anticancer drug screening: Worth the effort? J. Cell. Physiol. 2018, 233, 2993–3003. [Google Scholar] [CrossRef]
- Guirado, D.; Aranda, M.; Ortiz, M.; Mesa, J.A.; Zamora, L.I.; Amaya, E.; Villalobos, M.; Lallena, A.M. Low-dose radiation hyper-radiosensitivity in multicellular tumour spheroids. Br. J. Radiol. 2012, 85, 1398–1406. [Google Scholar] [CrossRef] [Green Version]
- Aranda, M. Los Esferoides Multicelulares como test Predictivo de Radiosensibilidad y Radiocurabilidad Tumoral. Ph.D. Thesis, University of Granada, Granada, Spain, 2003. [Google Scholar]
- Maruŝić, M.; Bajzer, Z.; Freyer, J.P.; Vuk-Pavlović, S. Analysis of growth of multicellular tumour spheroids by mathematical models. Cell Prolif. 1994, 27, 73–94. [Google Scholar] [CrossRef] [PubMed]
- Benzekry, S.; Lamont, C.; Beheshti, A.; Tracz, A.; Ebos, J.M.L.; Hlatky, L.; Hahnfeldt, P. Classical mathematical models for description and prediction of experimental tumor growth. PLoS Comput. Biol. 2014, 10, e1003800. [Google Scholar] [CrossRef] [Green Version]
- Vaidya, V.G.; Alexandro, F.J. Evaluation of some mathematical models for tumor growth. Int. J. Biomed. Comput. 1982, 13, 19–36. [Google Scholar] [CrossRef]
- Olea, N.; Villalobos, M.; Nuñez, J.; Elvira, J.; Ruiz de Almodóvar, J.M.; Pedraza, V. Evaluation of the growth rate of MCF-7 breast cancer multicellular spheroids using three mathematical models. Cell Prolif. 1993, 27, 213–223. [Google Scholar] [CrossRef]
- Wallace, D.I.; Guo, X. Properties of tumor spheroid growth exhibited by simple mathematical models. Front. Oncol. 2013, 3, 51. [Google Scholar] [CrossRef] [Green Version]
- Barbolosi, D.; Freyer, G.; Ciccolini, J.; Iliadis, A. Optimisation de la posologie et des modalités d,administration des agents cytotoxiques à l,aide d,un modèle mathématique. Bull. Cancer 2003, 90, 167–175. [Google Scholar] [PubMed]
- Swierniak, A.; Kimmel, M.; Smieja, J. Mathematical modeling as a tool for planning anticancer therapy. Eur. J. Pharmacol. 2009, 625, 108–121. [Google Scholar] [CrossRef] [Green Version]
- Simeoni, M.; De Nicolao, G.; Magni, P.; Rocchetti, M.; Poggesi, I. Modeling of human tumor xenografts and dose rationale in oncology. Drug Discov. Today Technol. 2013, 10, e365–e372. [Google Scholar] [CrossRef]
- Simeoni, M.; Magni, P.; Cammia, C.; De Nicolao, G.; Croci, V.; Presenti, E.; Germani, M.; Poggesi, I.; Rocchetti, M. Predictive pharmacokinetic-pharmacodynamic modeling of tumor growth kinetics in xenograft models after administration of anticancer agents. Cancer Res. 2004, 64, 1094–1101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ribba, B.; Watkin, E.; Tod, M.; Girard, P.; Grenier, E.; You, B.; Guiraudo, E.; Freyer, G. A model of vascular tumour growth in mice combining longitudinal tumour size data with histological biomarkers. Eur. J. Cancer 2011, 47, 479–490. [Google Scholar] [CrossRef]
- The Royal College of Radiologists. Timely Delivery of Radical Radiotherapy: Guidelines for the Management of Unscheduled Treatment Interruptions, 4th ed.; Ref. No. BFCO(19)1; The Royal College of Radiologists: London, UK, 2019. [Google Scholar]
- Murphy, H.; Jaafari, H.; Dobrovolny, H.M. Differences in predictions of ODE models of tumor growth: A cautionary example. BMC Cancer 2016, 16, 163. [Google Scholar] [CrossRef] [Green Version]
- Bilous, M.; Serdjebi, C.; Boyer, A.; Tomasini, P.; Pouypoudat, C.; Barbolosi, D.; Barlesi, F.; Chomy, F.; Benzekry, S. Quantitative mathematical modeling of clinical brain metastasis dynamics in non-small cell lung cancer. Sci. Rep. 2019, 9, 13018. [Google Scholar] [CrossRef] [Green Version]
- Wei, H.C. Mathematical modeling of tumor growth: The MCF-7 breast cancer cell line. Math. Biosci. Eng. 2019, 16, 6512–6535. [Google Scholar] [CrossRef]
- Vaghi, C.; Rodallec, A.; Fanciullino, R.; Ciccolini, J.; Mochel, J.P.; Mastri, M.; Poignard, C.; Ebos, J.L.M.; Benzekry, S. Population modeling of tumor growth curves and the reduced Gompertz model improve prediction of the age of experimental tumors. PLoS Comput. Biol. 2020, 16, e1007178. [Google Scholar] [CrossRef] [Green Version]
- Nicolò, C.; Périer, C.; Prague, M.; Bellera, C.; MacGrogan, G.; Saut, O.; Benzekry, S. Machine learning and mechanistic modeling for prediction of metastatic relapse in early-stage breast cancer. JCO Clin. Cancer Inform. 2020, 4, 259–274. [Google Scholar] [CrossRef]
- Rockne, R.C.; Hawkins-Daarud, A.; Swanson, K.R.; Sluka, J.P.; Glazier, J.A.; Macklin, P.; Hormuth, D.A., II; Jarrett, A.M.; Lima, E.A.; Oden, J.T.; et al. The 2019 mathematical oncology roadmap. Phys. Biol. 2019, 16, 041005. [Google Scholar] [CrossRef]
- Hadjicharalambous, M.; Wijeratne, P.A.; Vavourakis, V. From tumour perfusion to drug delivery and clinical translation of in silico cancer models. Methods 2021, 185, 82–93. [Google Scholar] [CrossRef] [PubMed]
- Vera, J.; Lischer, C.; Nenov, M.; Nikolov, S.; Lai, X.; Eberhardt, M. Mathematical modelling in biomedicine: A primer for the curious and the skeptic. Int. J. Mol. Sci. 2021, 22, 547. [Google Scholar] [CrossRef] [PubMed]
- Düchting, W.; Lehrig, R.; Rademacher, G.; Ulmer, W. Computer simulation of clinical irradiation schemes applied to in vitro tumor spheroids. Strahlenther. Onkol. 1989, 165, 873–878. [Google Scholar]
- Düchting, W.; Ulmer, W.; Ginsberg, T. Cancer: A challenge for control theory and computer modelling. Eur. J. Cancer 1996, 32A, 1283–1292. [Google Scholar] [CrossRef]
- Al-Dweri, F.M.O.; Guirado, D.; Lallena, A.M. Simulación de programas fraccionados de radioterapia. Estudio del control tumoral y del efecto de la interrupción del tratamiento. Rev. Fís. Méd. 2001, 2, 17–20. [Google Scholar]
- Al-Dweri, F.M.O.; Guirado, D.; Lallena, A.M.; Pedraza, V. Effect on tumour control of time interval between surgery and postoperative radiotherapy: An empirical approach using Monte Carlo Simulation. Phys. Med. Biol. 2004, 49, 2827–2839. [Google Scholar] [CrossRef]
- Powathil, G.; Kohandel, M.; Sivaloganathan, S.; Oza, A.; Milosevic, M. Mathematical modeling of brain tumors: Effects of radiotherapy and chemotherapy. Phys. Med. Biol. 2007, 52, 3291–3306. [Google Scholar] [CrossRef] [PubMed]
- Powathil, G.G.; Gordon, K.E.; Hill, L.A.; Chaplain, M.A.J. Modelling the effects of cell-cycle heterogeneity on the response of a solid tumour to chemotherapy: Biological insights from a hybrid multiscale cellular automaton model. J. Theor. Biol. 2012, 308, 1–19. [Google Scholar] [CrossRef]
- Powathil, G.G.; Adamson, D.J.A.; Chaplain, M.A.J. Towards predicting the response of a solid tumour to chemotherapy and radiotherapy treatments: Clinical insights from a computational model. PLoS Comput. Biol. 2013, 9, e1003120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nikolov, S.; Dimitrov, A.; Vera, J. Hierarchical levels of biological systems and their integration as a principal cause for tumour occurrence. Nonlinear Dyn. Psychol. Life Sci. 2019, 23, 315–329. [Google Scholar]
- Hamis, S.; Powathil, G.G.; Chaplain, M.A.J. Blackboard to bedside: A mathematical modeling bottom-up approach toward personalized cancer treatments. JCO Clin. Cancer Inform. 2019, 3, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Materi, W.; Wishart, D.S. Computational systems biology in cancer: Modeling methods and applications. Gene Regul. Syst. Biol. 2007, 1, 91–110. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Butner, J.D.; Kerketta, R.; Cristini, V.; Deisboeck, T.S. Simulating cancer growth with multiscale agent-based modeling. Semin. Cancer Biol. 2015, 30, 70–78. [Google Scholar] [CrossRef] [Green Version]
- Ruiz-Arrebola, S.; Tornero-López, A.M.; Guirado, D.; Villalobos, M.; Lallena, A.M. An on-lattice agent-based Monte Carlo model simulating the growth kinetics of multicellular tumor spheroids. Phys. Med. 2020, 77, 194–203. [Google Scholar] [CrossRef] [PubMed]
- Yorke, E.D.; Fuks, Z.; Norton, L.; Whitmore, W.; Ling, C.C. Modeling the development of metastases from primary and locally recurrent tumors: Comparison with a clinical data base for prostatic cancer. Cancer Res. 1993, 53, 2987–2993. [Google Scholar]
- Spratt, J.A.; Von Fournier, D.; Spratt, J.S.; Weber, E.E. Decelerating growth and human breast cancer. Cancer 1993, 71, 2013–2019. [Google Scholar] [CrossRef]
- Von Bertalanffy, L. Quantitative laws in metabolism and growth. Q. Rev. Biol. 1957, 32, 217–231. [Google Scholar] [CrossRef]
- West, G.B.; Brown, J.H.; Enquist, B.J. A general model for ontogenetic growth. Nature 2001, 413, 628–631. [Google Scholar] [CrossRef]
- Guiot, C.; Degiorgis, P.G.; Delsanto, P.P.; Gabriele, P.; Deisboeck, T.S. Does tumor growth follow a ‘universal law’? J. Theor. Biol. 2003, 225, 147–151. [Google Scholar] [CrossRef] [Green Version]
- Herman, A.B.; Savage, V.M.; West, G.B. A quantitative theory of solid tumor growth, metabolic rate and vascularization. PLoS ONE 2011, 6, e22973. [Google Scholar] [CrossRef]
- Villalobos, M. Modelos Tumorales en Oncología: Los Esferoides Multicelulares en el Estudio del Cáncer Hormonodependiente. Ph.D. Thesis, University of Granada, Granada, Spain, 1996. [Google Scholar]
- Gong, X.; Lin, C.; Cheng, J.; Su, J.; Zhao, H.; Liu, T.; Wen, X.; Zhao, P. Generation of multicellular tumor spheroids with microwell-based agarose xcaffolds for drug testing. PLoS ONE 2015, 10, e0130348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Press, W.H.; Teukolsky, S.A.; Vetterling, W.T.; Flannery, B.P. Numerical Recipes in FORTRAN. The Art of Scientific Computing; Cambridge University Press: New York, NY, USA, 1995. [Google Scholar]
- Withers, H.R.; Taylor, J.M.; Maciejewski, B. The hazard of accelerated tumor clonogen repopulation during radiotherapy. Acta Oncol. 1988, 27, 131–146. [Google Scholar] [CrossRef]
- Fowler, J.F. 21 years of biologically effective dose. Br. J. Radiol. 2010, 83, 554–568. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Model | All Data | Retrospective | Prospective | ||
|---|---|---|---|---|---|---|
| #1 | G | 3.23 | 1.09 | 29.08 | 1.04 | 50.49 |
| [1.89:6.29] | [0.51:1.82] | [5.47:68.67] | [0.49:1.87] | [3.78:126.70] | ||
| L | 171.49 | 1.77 | 478.86 | 3.58 | 1050.98 | |
| [139.06:258.91] | [0.81:2.95] | [360.00:613.40] | [1.97:5.51] | [956.80:1140.00] | ||
| P | 22.39 | 2.56 | 64.59 | 2.35 | 340.05 | |
| [17.50:27.41] | [1.34:4.18] | [44.49:87.20] | [1.12:3.91] | [212.30:501.50] | ||
| B | 30.94 | 1.08 | 22.99 | 1.40 | 304.76 | |
| [21.74:46.02] | [0.50:1.78] | [4.74:48.63] | [0.64:2.52] | [92.49:694.50] | ||
| #2 | G | 3.79 | 1.11 | 29.76 | 1.11 | 58.79 |
| [1.29:9.83] | [0.45:2.19] | [1.46:94.18] | [0.44:2.12] | [1.35:183.90] | ||
| L | 179.48 | 1.80 | 480.19 | 3.75 | 1061.00 | |
| [56.72:378.29] | [0.74:3.55] | [147.10:959.40] | [1.14:8.28] | [467.80:1833.00] | ||
| P | 24.17 | 2.60 | 65.14 | 2.65 | 340.71 | |
| [6.22:57.37] | [1.12:4.85] | [15.11:139.60] | [0.71:6.36] | [54.31:835.20] | ||
| B | 31.02 | 1.08 | 23.37 | 1.44 | 315.34 | |
| [11.46:64.49] | [0.46:2.17] | [4.18:52.42] | [0.57:2.76] | [67.76:836.80] | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ruiz-Arrebola, S.; Guirado, D.; Villalobos, M.; Lallena, A.M. Evaluation of Classical Mathematical Models of Tumor Growth Using an On-Lattice Agent-Based Monte Carlo Model. Appl. Sci. 2021, 11, 5241. https://doi.org/10.3390/app11115241
Ruiz-Arrebola S, Guirado D, Villalobos M, Lallena AM. Evaluation of Classical Mathematical Models of Tumor Growth Using an On-Lattice Agent-Based Monte Carlo Model. Applied Sciences. 2021; 11(11):5241. https://doi.org/10.3390/app11115241
Chicago/Turabian StyleRuiz-Arrebola, Samuel, Damián Guirado, Mercedes Villalobos, and Antonio M. Lallena. 2021. "Evaluation of Classical Mathematical Models of Tumor Growth Using an On-Lattice Agent-Based Monte Carlo Model" Applied Sciences 11, no. 11: 5241. https://doi.org/10.3390/app11115241