
Generation and Characterization of Novel Bioactive Peptides from Fish and Beef Hydrolysates



1
Department of Food Hygiene and Control, Faculty of Veterinary Medicine, South Valley University, Qena 83522, Egypt
2
Laboratory of Microbial Technology, Division of Systems Bioengineering, Department of Bioscience and Biotechnology, Faculty of Agriculture, Graduate School, Kyushu University, Fukuoka 819-0395, Japan
*
Author to whom correspondence should be addressed.
Appl. Sci. 2021, 11(21), 10452; https://doi.org/10.3390/app112110452
Submission received: 13 October 2021
/
Revised: 3 November 2021
/
Accepted: 4 November 2021
/
Published: 7 November 2021
(This article belongs to the Special Issue Bioactive Molecules in Food)
Abstract
:Bioactive peptides were successfully produced from fish (Gadidae) and beef skeletal muscles after being hydrolyzed for 8 h with pepsin. Subsequently, they were purified using a Sep-Pak C18 cartridge and reversed-phase high-performance liquid chromatography (RP-HPLC). The molecular weights of pure fish and beef peptides were determined to be 2364.4 and 3771.8, respectively. According to Edman degradation, the fish peptide was composed of 21 amino acid residues (F21), while the beef peptide was composed of 34 amino acid residues (B34). F21 and B34 displayed angiotensin-converting enzyme inhibitory activity with a half maximal inhibitory concentration (IC50) values of 7.3 µg/mL and 5.8 µg/mL, respectively. F21 exhibited antioxidant activity with an IC50 value of 389.9 µg/mL, whereas B34 exhibited no antioxidant activity. Moreover, F21 and B34 displayed antimicrobial effects against a wide spectrum of food-borne pathogens and spoilage bacteria. Bioactive peptides derived from muscle proteins are a promising strategy for the production of functional food materials and safe food preservatives.
1. Introduction
Meat proteins are essential food constituents with nutritional and physiological properties. They are rich in essential amino acids, which are required for development of the body as well as the chemical and sensory characteristics of protein-enclosing products. Moreover, meat proteins contain bioactive peptides, which are protein fragments containing certain amino acids. Bioactive peptides are inert in the original protein chains but can be generated by proteolytic digestion. They could pass through the intestine and into the circulation, where they can perform a variety of biological functions. Bioactive peptides have received increasing attention and have been recommended as prophylactic and therapeutic substances for a variety of illnesses [1]. Antimicrobial, angiotensin-converting enzyme (ACE) inhibitory, and antioxidant characteristics have already been reported in a variety of bioactive peptides [2].
Food-derived antibacterial peptides are promising alternatives to harmful chemical preservatives and can be safely used in food preservation [3]. There are multiple reports on bioactive peptides with antimicrobial activity from milk proteins and lactoferricin [4,5]. Lactenin was the first antimicrobial agent isolated from the rennet hydrolysate of milk [6]. Biological activity of lactoferrin-related peptides against bacteria, fungus, and yeast has been documented [7].
ACE participates in the renin–angiotensin mechanism and is important in adjusting blood pressure. ACE also suppresses the vasodilatation job of bradykinin [8]. Various ACE inhibitors were discovered in proteins obtained from food—including meat [9], milk [10], soya bean [11], and pork muscle [12].
Lipid oxidation of meat is a major concern in food safety. Lipid oxidation can damage food quality and liberate harmful compounds, resulting in tumor formation and adverse cardiovascular effects in consumers [13]. Malondialdehyde can be formed during lipid oxidation as a result of peroxide oxidation. Butylated hydroxyanisole and butylated hydroxytoluene are synthetic preservatives that are frequently used to lower fat oxidation and microbial growth in food. However, they have an adverse influence on public health [14]. On the other hand, bioactive peptides liberated from meat can reduce the oxidation of fat in foods, including those generated by papain and actinase E digestion of pork skeletal muscle, as well as papain digestion of marine protein [15]. Various synthetic antioxidants have been developed to control fat oxidation; however, antioxidants derived from protein hydrolysates are considered safer.
Beef and marine muscles are promising sources for bioactive peptides. The aim of this study is to identify bioactive peptides in beef and fish hydrolysates digested with pepsin, as well as evaluate their antimicrobial, antioxidant, and ACE inhibitory properties.
2. Materials and Methods
2.1. Protein Extraction and Enzymatic Hydrolysis
Frozen fish and beef samples were processed to extract the bioactive peptide, as described by Jang and Lee [16], with slight modifications. Briefly, fish (Gadidae) and beef skeletal muscle samples (30 g each) were mixed individually with 200 mL of 0.02 M sodium phosphate buffer (pH 7.4) and centrifuged at 15,770× g for 20 min at 4 °C. The supernatant was collected, and the pH was adjusted by using 0.1 M HCl to the optimal value (pH 3) for the enzyme reaction, and 10 mg of pepsin enzyme (Sigma-Aldrich, St. Louis, MO, USA) was added. After 8 h of digestion at 37 °C, the solution was boiled for 5 min to stop the enzyme action, followed by cooling at 25 °C.
2.2. Purification of Protein Hydrolysates
The pepsin-digested fish and beef samples were concentrated using a SpeedVac concentrator and sterilized using a sterile cellulose acetate membrane filter (0.2 μm, Advantec, Tokyo, Japan) before fractionation and are known as fish and beef hydrolysates, respectively. Furthermore, peptides in the hydrolysates were purified using a Sep-Pak C18 cartridge (Waters, Milford, MA, USA). Elution was conducted using acetonitrile containing 0.1% trifluoroacetic acid (TFA). The eluted fractions were processed in a Speed Vac concentrator (Savant, Farmingdale, NY, USA) to remove the solvent. The fractions were analyzed using an Atlantis dC18 column (4.6 × 150 mm, 5 µm; Waters) in an LC-2000 Plus high-performance liquid chromatography (HPLC) system (JASCO, Tokyo, Japan). The elution program consisted of a gradient of Milli-Q-acetonitrile containing 0.1% TFA at a flow rate of 1 mL/min: 0–45 min, 0–70% acetonitrile. Concentration of proteins in the hydrolysates and the purified peptides were determined using the Pierce bicinchoninic acid (BCA) protein assay kit (Thermo Fisher Scientific, Waltham, MA, USA).
2.3. Mass Spectrometry and Amino Acid Sequencing
The molecular weights of the fish and beef peptides were determined using electrospray ionization time-of-flight mass spectrometry (ESI-TOF MS) with a JMS-T100LC mass spectrometer (JEOL, Tokyo, Japan) according to a previous study [17]. The purest fractions from beef and fish hydrolysates that appeared in the mass spectrum were selected for further analyses.
Amino acid sequencing was carried out by Edman degradation [18] using a PPSQ-31 automated protein sequencer (Shimadzu, Kyoto, Japan), according to the manufacturer’s instructions. Briefly, purified F21 and B34 peptides were concentrated using a SpeedVac concentrator (Thermo Fisher Scientific) and then applied to the protein sequencer. The profile obtained in each cycle of Edman degradation was processed and analyzed automatically by the PPSQ-31 program, and the amino acid sequences were identified.
2.4. Sequence Analysis
Identity exploration of the amino acid sequences of the purified F21 and B34 peptides was conducted using NCBI BLAST (http://www.ncbi.nlm.gov/BLAST (accessed on 10 January 2021)). The secondary structure was predicted using the GOR method accessible via the Prabiserver (https://npsa-prabi.ibcp.fr (accessed on 12 January 2021)).
2.5. Determination of Antimicrobial Activity
Minimum inhibitory concentrations (MICs) were evaluated using the broth micro-dilution method as reported by Wiegand et al. [19] against a panel of indicator strains prepared as follows. Enterococcus faecalis JCM 5803T was cultured in MRS medium (Oxoid, Basingstoke, United Kingdom). Listeria innocua ATCC 33090T, E. coli JM109, Bacillus coagulans JCM 2257T, Salmonella enterica subsp. enterica NBRC 13245T, and Proteus vulgaris F24B isolated from Japanese fish [20] and displaying multi-drug resistance were cultured in tryptic soy broth (BD, Sparks, MD, USA) supplemented with 0.6% yeast extract (Nacalai Tesque, Kyoto, Japan), whereas Pseudomonas putida ATCC 12633T was cultured in Luria-Bertani medium (BD).
Briefly, 90 µL culture of the indicator strain was incubated in the appropriate culture media at the appropriate temperature in the microtiter plate until an optical density (OD620) of 0.11 was reached. Subsequently, 10 μL portions of the two-fold serially diluted hydrolysates, F21 and B34, were prepared and added to the culture. The total volume of each well was 100 µL. Cultures without peptides were used as a control. The microtiter plates were incubated overnight, and bacterial inhibition was measured at OD620 using an Infinite F200 Pro microplate reader (Tecan, Männedorf, Switzerland). MICs were defined as the lowest concentrations that inhibited the growth of indicator bacteria.
2.6. ACE Inhibition Activity Assay
The ACE inhibition activity of the hydrolysates, F21 and B34, was assayed using the ACE kit-WST (Dojindo Laboratories, Kumamoto, Japan), according to the manufacturer’s instructions. Briefly, 20 μL of each sample was added to each well of a microplate, and subsequently 20 μL substrate buffer, deionized water, and enzyme working solution were added to each well and incubated for 1 h at 37 °C. Then, 200 μL of indicator solution was added and the plate was incubated for 10 min at room temperature. The optical density at 450 nm was measured using a Sunrise microplate absorbance reader (Tecan), and the IC50 values were determined via regression line analysis.
2.7. Antioxidant Activity Assay
The antioxidant property of the hydrolysates, F21 and B34 was analyzed using the DPPH antioxidant assay kit (Dojindo Laboratories), based on the manufacturer’s guidelines. Briefly, 20 μL of samples were added to each well of a microtiter plate, followed by 80 μL assay buffer, 100 μL ethanol, and 100 μL DPPH working solution. Then, the plate was incubated at 25 °C for 30 min in the dark. The optical density was measured at 517 nm using a Sunrise microplate absorbance reader. The IC50 value was determined via regression line analysis, and the antioxidant activity was also described as the Trolox equivalent antioxidant capacity (TEAC; IC50 Trolox/IC50 sample).
3. Results
3.1. Generation, Purification, and Structural Analysis of Fish and Beef Peptides
The fish and beef hydrolysates were purified by means of a Sep-Pak C18 cartridge with 65% acetonitrile, followed by reverse-phase HPLC (Figure 1A,B). All RP-HPLC fractions of fish and beef were analyzed by mass spectrometry, and the purest fractions from fish and beef peptides were selected based on clearance of the mass spectra for further study. The molecular weights of F21 and B34 were 2364.4 and 3771.8, respectively (Figure 1C,D). Furthermore, the amino acid sequences of F21 was FKYDSTHGRFHGEVKAEGGKL, while of B34 was TALGGILKKKGHHEAEVKHLAESHANKHKIPVKY as identified by Edman degradation. The observed molecular weights were identical to those calculated according to the amino acid sequences determined. A database search showed that F21 and B34 possessed 100% identity to partial sequences of glyceraldehyde 3-phosphate dehydrogenase (GAPDH), hypothetical protein from Salmo salar (a species of ray-finned fish) with accession number NP_001117033.1, and myoglobin from Bos taurus (cattle) with accession number NP_776306.1, respectively.
3.2. Prediction of Secondary Structure
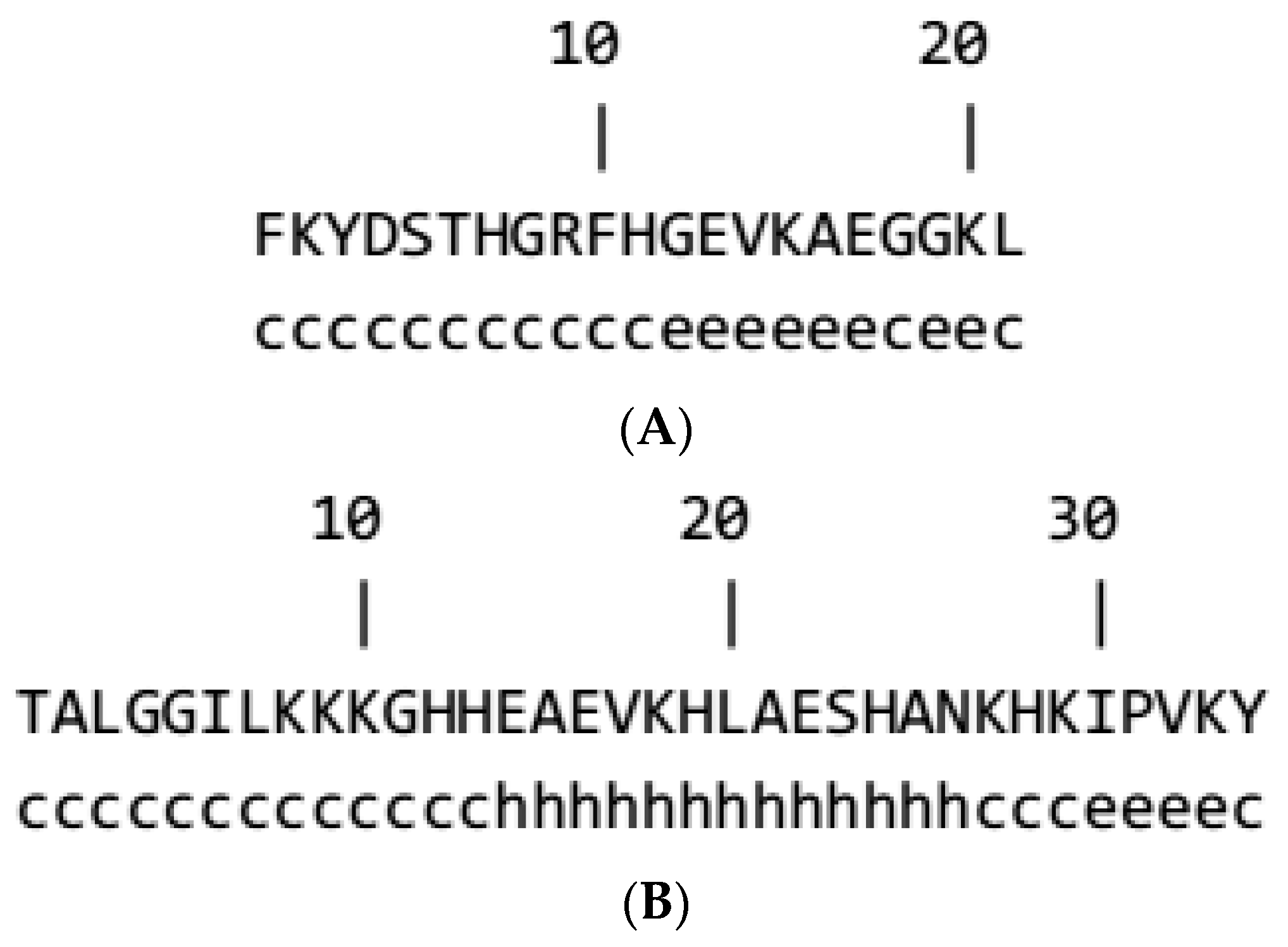
The secondary structures of the F21 and B34 were anticipated to further characterize their structures. The GOR program indicated that extended strands and random coils would probably form in F21 (Figure 2A). The GOR program also suggested that the central region in B34 (amino acids 14–26) could develop an α-helix (Figure 2B).
3.3. Antimicrobial Spectrum
The antimicrobial activities of hydrolysates F21 and B34 are illustrated in Table 1. Fish and beef hydrolysates displayed antimicrobial activity toward various indicator strains, including food pathogens and spoilage bacteria. F21 exhibited antimicrobial activity against a broad spectrum of food microbes. Interestingly, B34 displayed antimicrobial activity against Proteus vulgaris F24B, a multi-drug resistant bacterium.
3.4. ACE Inhibition and Antioxidant Activities
Beef hydrolysates showed stronger ACE inhibitory activity (IC50, 265.6 µg/mL) than fish hydrolysates (IC50 304.5 µg/mL). For the antioxidant property, fish hydrolysates (IC50, 470.4 µg/mL) displayed more potency than the beef hydrolysates (IC50, 531.0 µg/mL). Fish and beef hydrolysates exhibited almost identical TEAC activities (Table 2).
F21 and B34 displayed ACE inhibitory activity with IC50 values of 7.3 and 5.8 µg/mL, respectively, suggesting higher ACE inhibitory activity than the hydrolysates. Antioxidant activity was observed in the F21 (IC50, 389.9 µg/mL; TEAC, 0.17), whereas no activity was detected in B34 (Table 2).
4. Discussion
The rise in food-related diseases—such as heart diseases, hypertension, and obesity—has prompted consumers to seek food products that provide both nutritional and health benefits. In this study, two unreported multifunctional peptides were generated, purified, and identified from fish and beef using a Sep-Pak C18 cartridge, RP-HPLC, ESI-TOF MS, and Edman degradation. Further characterization of the peptides demonstrated that hydrolysis of proteins by enzymes, such as pepsin, can be a good strategy to obtain bioactive peptides and that peptides obtained from fish and beef are prospective sources of bioactive peptides.
Amino acid sequences of F21 showed high similarity to those of GAPDH. GAPDH has a variety of biological functions, including regulation of the sixth stage of glycolysis and the prevention of cell death [21]. Two GAPDH-related peptides, YFGAP and SJGAP—obtained from fish skin—showed antibacterial properties and molecular masses of 3.4 kDa [22,23]. Each peptide was composed of 32 amino acid residues that had not undergone any post-translational modifications. The fish peptide identified in this study had a different amino acid sequence from YFGAP and SJGAP, and GAPDH can be considered a good source of bioactive peptides.
Beef is also considered a potential source of bioactive peptides. In previous studies, small peptide fragments produced by acid whey digestion of beef showed various biological functions, such as immunomodulatory, ACE inhibitory, and antioxidative activities and enhancement of the cardiovascular and nervous systems [24]. Additionally, some other bioactive peptides originating from beef have been identified and characterized [25,26].
The antibacterial activity of bioactive peptides depends on the allocation of hydrophobic and charged residues in the peptide structure [27]. The existence of positively charged residues, as well as an amphipathic structure, allows the peptide to interact with the bacterial membrane via electrostatic intercommunication, similarly to other antimicrobial peptides [28]. Furthermore, the occurrence of aromatic residues, random coils, and helical regions in peptides is frequently linked to biological activity [29]. The fish peptide obtained in the present study lacked helix formation, whereas YFGAP and SJGAP contained one helix and two strands respectively [22,23], suggesting that F21 shows different biological activity from that of the previously reported peptides.
The fish hydrolysates and F21 exhibited broad antimicrobial spectra compared to beef, which can be attributed to differences in the amino acid sequence and secondary structure. Seo et al. [22] reported that the purified fish peptide (YFGAP) exhibited antimicrobial activity against some Gram-negative bacteria, E. coli D31 (minimal effective concentration (MEC), 6.2 μg/mL) and Gram-positive B. subtilis KCTC1021 (MEC, 13 μg/mL), which was similar to that of F21. A few studies have shown peptides extracted from meat with considerable antibacterial activity [30]. Jang et al. [31] reported that certain bioactive peptides extracted from beef demonstrated antimicrobial activity against E. coli, Salmonella typhimurium, and Pseudomonas, which can be considered as a broader antibacterial spectrum than that of B34. However, Keska and Stadnik [32] stated that there was no proof of antimicrobial activity of peptides obtained from cow and pork on Staphylococcus aureus and E. coli. The antimicrobial activity of peptides generated from meat varies greatly depending on the origin and their primary and secondary structures.
The generation and characterization of new ACE inhibitors are required for applications in food and medicine. B34 showed a higher ACE inhibitory action than those reported by Jang and Lee [16]. They reported that ACE inhibitory peptide fractions extracted from beef hydrolysates with IC50 values ranging from 23.11 to 24.15 µg/mL. Furthermore, B34 was more potent than an ACE inhibitory peptide (IC50 28.5 µM) generated by digestion of pork with pepsin [33]. On the other hand, the ACE inhibitory activity of peptide fractions obtained after hydrolysis of skin and bone gelatin of catfish by alcalase (IC50 3.2 μg/mL and 1.3 μg/mL, respectively) were more potent than F21. The amino acid sequence of the peptides with ACE inhibitory activity, the hydrophobic residues containing aliphatic side groups—such as Ile, Gly, Leu, Ala, and Val—at the C-terminus were responsible for enhancing the ACE inhibitory activity due to their increased binding capability with ACE [34]. Furthermore, positively charged residues, such as Arg and Lys, are also implicated in the greater efficacy of ACE inhibitory activity [35]. Additionally, the ACE inhibitory activity of peptides depends on their molecular masses; a study showed that peptides with a molecular mass < 3 kDa displayed greater ACE inhibitory activity than larger peptides [35]. Another general property of ACE inhibitory peptides is the occurrence of hydrophobic amino acid residues at the N-terminals of peptides [36]. B34 has more hydrophobic residues in the N-terminal—including Ala, Leu, and Ile—than the fish peptide, and this illustrates the higher ACE inhibitory activity of the pure beef peptide. However, the precise correlation between the functions of peptides and their structures is still unclear [37].
Free radicals target the main biological compounds, such as DNA, and negatively influence human health, leading to cardiovascular diseases and tumor formation. Utilization of antioxidant peptides derived from natural sources is a promising approach to attenuate the effects of free radicals. F21 exhibited greater antioxidant activity than bioactive peptides derived by enzymatic hydrolysis of tuna head (IC50 value of 1.34 mg/mL) [38] and an antioxidant peptide derived from a ray gelatin hydrolysate (IC50 value of 1.98 mg/mL) [39]. The location of the peptide in the protein composition, hydrophobicity, molecular weight, and amino acid sequence influence the antioxidant action of peptides.
5. Conclusions
In the current work, fish and beef were found to be possible sources of bioactive peptides. Antimicrobial, antihypertensive, and antioxidant peptides generated by enzymatic hydrolysis of fish and beef proteins are promising compounds for applications in food and medicine. Only one peptide obtained from the fish and beef hydrolysates each was characterized here, but the results of the hydrolysates suggest that they contain more bioactive peptides. Further screening of peptides in fish and beef hydrolysates, as well as characterization of their action mechanisms, will improve the likelihood of their application.
Author Contributions
Conceptualization, M.A.M. and T.Z.; Methodology, M.A.M. and T.Z.; Practical work and acquisition of the data, M.A.M. and T.Z.; Analysis and interpretation of the data, M.A.M. and T.Z.; Resources, T.Z.; Data Curation, M.A.M. and T.Z.; Writing—original draft preparation, M.A.M.; Writing—review and editing, M.A.M. and T.Z.; Supervision, T.Z.; Funding acquisition, T.Z. All authors have read and agreed to the published version of the manuscript.
Funding
The research was supported by an Egyptian postdoctoral scholarship awarded to Mohamed Abdelfattah Maky and JSPS KAKENHI Scholarship Number 21H02107.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data sharing not applicable.
Acknowledgments
The authors acknowledge Takashi Nakashima of Kyushu University for his aid in the amino acid sequence analysis.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Marqus, S.; Pirogova, E.; Piva, T.J. Evaluation of the use of therapeutic peptides for cancer treatment. Biomed. Sci. 2017, 24, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Halim, N.R.A.; Yusof, H.M.; Sarbon, N.M. Functional and bioactive properties of fish protein hydrolysates and peptides: A comprehensive review. Trends Food Sci. Technol. 2016, 51, 24–33. [Google Scholar] [CrossRef]
- Przybylski, R.; Firdaous, L.; Chataigne, G.; Dhulster, P.; Nedjar, N. Production of an antimicrobial peptide derived from slaughterhouse byproduct and its potential application on meat as preservative. Food Chem. 2016, 211, 306–313. [Google Scholar] [CrossRef]
- Bellamy, W.; Takase, M.; Yamauchi, K.; Wakabayashi, H.; Kawase, K.; Tomita, M. Identification of the bactericidal domain of lactoferrin. Biochim. Biophys. Acta 1992, 1121, 130–136. [Google Scholar] [CrossRef]
- McCann, K.B.; Shiell, B.J.; Michalski, W.P.; Lee, A.; Wan, J.; Roginsky, H.; Coventry, M.J. Isolation and characterization of antibacterial peptides derived from the f(164–207) region of bovine αS2-casein. Int. J. Dairy Sci. 2005, 15, 133–143. [Google Scholar] [CrossRef]
- Jones, F.S.; Simms, H.S. The bacterial growth inhibitor (lactenin) of milk. J. Exp. Med. 1929, 51, 327–339. [Google Scholar] [CrossRef] [Green Version]
- Korhonen, H.; Pihlanto, A. Bioactive peptides from food proteins. In Handbook of Food Products Manufacturing; Hui, Y.H., Ed.; John Wiley and Sons Inc.: Hoboken, NJ, USA, 2007; pp. 5–37. [Google Scholar]
- Furuta, T.; Miyabe, Y.; Yasui, H.; Kinoshita, Y.; Kishimura, H. Angiotensin I converting enzyme inhibitory peptides derived from phycobiliproteins of dulse Palmaria palmata. Mar. Drugs 2016, 14, 32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vercruysse, L.; Camp, J.V.; Smagghe, G. ACE inhibitory peptides derived from enzymatic hydrolysates of animal muscle protein: A review. J. Agric. Food Chem. 2005, 53, 8106–8115. [Google Scholar] [CrossRef]
- Maruyama, S.; Nakagomi, K.; Tomizuka, N.; Suzuki, H. Angiotensin I-converting enzyme inhibitor derived from an enzymatic hydrolysate of casein. Agric. Biol. Chem. 1985, 49, 1405–1409. [Google Scholar] [CrossRef]
- Okamoto, A.; Hanagata, H.; Kawamura, Y.; Yanagida, F. Anti-hypertensive substances in fermented soybean, natto. Plant Foods Hum. Nutr. 1995, 47, 39–47. [Google Scholar] [CrossRef]
- Arihara, K.; Nakashima, Y.; Mukai, T.; Ishikawa, S.; Itoh, M. Peptide inhibitors for angiotensin I-converting enzyme from enzymatic hydrolysates of porcine skeletal muscle proteins. Meat Sci. 2001, 57, 319–324. [Google Scholar] [CrossRef]
- Lobo, V.; Patil, A.; Phatak, A.; Chandra, N. Free radicals, antioxidants and functional foods: Impact on human health. Pharmacogn. Rev. 2010, 4, 118–126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imaida, K.; Fukushima, S.; Shirai, T.; Ohtani, M.; Nakanishi, K.; Ito, N. Promoting activities of butylated hydroxyanisole and butylated hydroxytoluene on 2-stage urinary carcinogenesis and inhibition of gamma-glutamyl transpeptidase-positivefor development in the liver of rats. Carcinogenesis 1983, 4, 895–899. [Google Scholar] [CrossRef] [PubMed]
- Sohaib, M.; Anjum, F.M.; Sahar, A.; Arshad, M.S.; Rahman, U.U.; Imran, A.; Hussain, S. Antioxidant proteins and peptides to enhance the oxidative stability of meat and meat products: A comprehensive review. Int. J. Food Prop. 2017, 20, 2581–2593. [Google Scholar] [CrossRef]
- Jang, A.; Lee, M. Purification and identification of angiotensin converting enzyme inhibitory peptides from beef hydrolysates. Meat Sci. 2005, 69, 653–661. [Google Scholar] [CrossRef]
- Zendo, T.; Nakayama, J.; Fujita, K.; Sonomoto, K. Bacteriocin detection by liquid chromatography/mass spectrometry for rapid identification. J. Appl. Microbiol. 2008, 104, 499–507. [Google Scholar] [CrossRef]
- Edman, P. Method for determination of the amino acid sequence in peptides. Acta Chem. Scand. 1950, 4, 283–293. [Google Scholar] [CrossRef] [Green Version]
- Wiegand, I.; Hilpert, K.; Hancock, R.E.W. Agar and broth dilution methods to determine the minimal inhibitory concentration (MIC) of antimicrobial substances. Nat. Protoc. 2008, 3, 163–175. [Google Scholar] [CrossRef]
- Maky, M.A.; Ishibashi, N.; Nakayama, J.; Zendo, T. Characterization of the biosynthetic gene cluster of enterocin F4-9, a glycosylated bacteriocin. Microorganisms 2021, 9, 2276. [Google Scholar] [CrossRef]
- Abuine, R.; Rathnayake, A.U.; Byun, H. Biological activity of peptides purified from fish skin hydrolysates. Fish. Aquat. Sci. 2019, 22, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Seo, J.; Lee, M.J.; Go, H.; Park, T.H.; Park, N.G. Purification and characterization of YFGAP, a GAPDH-related novel antimicrobial peptide, from the skin of yellow fin tuna, Thunnus albacares. Fish Shellfish Immunol. 2012, 33, 743–752. [Google Scholar] [CrossRef]
- Seo, J.; Lee, M.J.; Go, H.; Kim, Y.J.; Park, N.G. Antimicrobial function of the GAPDH related antimicrobial peptide in the skin of skipjack tuna, Katsuwonus pelamis. Fish Shellfish Immunol. 2014, 36, 571–581. [Google Scholar] [CrossRef]
- Keska, P.; Wojciak, K.M.; Stadnik, J. Bioactive peptides from beef products fermented by acid whey—In vitro and in silico study. Sci. Agric. 2019, 76, 311–320. [Google Scholar] [CrossRef]
- Ryan, J.T.; Ross, R.P.; Bolton, D.; Fitzgerald, G.F.; Stanton, C. Bioactive peptides from muscle Sources: Meat and fish. Nutrients 2011, 3, 765–791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xing, L.; Liu, R.; Cao, S.; Zhang, W.; Guanghong, Z. Meat protein based bioactive peptides and their potential functional activity: A review. Int. J. Food Sci. 2019, 54, 1956–1966. [Google Scholar] [CrossRef] [Green Version]
- Yeaman, M.R.; Yount, N.Y. Mechanisms of antimicrobial peptide action and resistance. Pharmacol. Rev. 2003, 55, 27–55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shai, Y. Mechanism of the binding, insertion and destabilization of phospholipid bilayer membranes by alpha-helical antimicrobial and cell non-selective membrane-lytic peptides. Biochim. Biophys. Acta 1999, 1462, 55–70. [Google Scholar] [CrossRef] [Green Version]
- Mahmoodani, F.; Ghassem, M.; Babji, A.S.; Yusop, S.M.; Khosrokhavar, R. ACE inhibitory activity of pangasius catfish (Pangasiussutchi) skin and bone gelatin hydrolysate. J. Food Sci. Technol. 2014, 51, 1847–1856. [Google Scholar] [CrossRef] [Green Version]
- Rezaharsamto, B.; Subroto, E. A review on bioactive peptides derived from various sources of meat and meat by-products. Int. J. Sci. Technol. Res. 2019, 8, 3151–3156. [Google Scholar]
- Jang, A.; Jo, C.; Kang, K.S.; Lee, M. Antimicrobial and human cancer cell cytotoxic effect of synthetic angiotensin-converting enzyme (ACE) inhibitory peptides. Food Chem. 2008, 107, 327–336. [Google Scholar] [CrossRef]
- Keska, P.; Stadnik, J. Antimicrobial peptides of meat origin—An in silico and in vitro analysis. Protein Pept. Lett. 2017, 24, 165–173. [Google Scholar] [CrossRef]
- Katayama, K.; Mori, T.; Kawahara, S.; Miake, K.; Kodama, Y.; Sugiyama, M.; Kawamura, Y.; Nakayama, T.; Maruyama, M.; Muguruma, M. Angiotensin-I converting enzyme inhibitory peptide derived from porcine skeletal muscle myosin and its antihypertensive activity in spontaneously hypertensive rats. J. Food Sci. 2007, 72, S702–S706. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.Y.; Hur, S.J. Purification of novel angiotensin converting enzyme inhibitory peptides from beef myofibrillar proteins and analysis of their effect in spontaneously hypertensive rat model. Biomed. Pharmacother. 2019, 116, 1–7. [Google Scholar] [CrossRef]
- Wang, R.; Lu, X.; Sun, Q.; Gao, J.; Ma, L.; Huang, J. Novel ACE inhibitory peptides derived from simulated gastrointestinal digestion in vitro of Sesame (Sesamumindicum L.) protein and molecular docking study. Int. J. Mol. Sci. 2020, 21, 1059. [Google Scholar] [CrossRef] [Green Version]
- Wijesekara, I.; Kim, S. Angiotensin-I-converting enzyme (ACE) inhibitors from marine resources: Prospects in the pharmaceutical industry. Mar. Drugs 2010, 8, 1080–1093. [Google Scholar] [CrossRef] [Green Version]
- Jakubczyk, A.; Karaś, M.; Rybczyńska-Tkaczyk, K.; Zielińska, E.; Zieliński, D. Current trends of bioactive peptides—New sources and therapeutic effect. Foods 2020, 9, 846. [Google Scholar] [CrossRef]
- Yang, P.; Ke, H.; Hong, P.; Zeng, S.; Cao, W. Antioxidant activity of bigeye tuna (Thunnus obesus) head protein hydrolysate prepared with alcalase. Int. J. Food Sci. 2011, 46, 2460–2466. [Google Scholar] [CrossRef]
- Lassoued, I.; Mora, L.; Nasri, R.; Jridi, M.; Toldra, F.; Aristoy, M.; Barkia, A.; Nasri, M. Characterization and comparative assessment of antioxidant and ACE inhibitory activities of thornback ray gelatin hydrolysates. J. Funct. Foods 2015, 13, 225–238. [Google Scholar] [CrossRef]
Figure 1.
Purification and identification of the bioactive peptides from fish and beef. Chromatographic profiles of the fish and beef hydrolysates obtained by RP-HPLC using a dC18 column (A and B, respectively). The peaks marked by arrows show the target peptides termed F21 and B34. ESI-TOF mass spectra showing multiple charged molecular ions of the peptides F21 and B34 (C and D, respectively). The molecular masses of F21 and B34 were calculated based on the most abundant peaks.
Figure 1.
Purification and identification of the bioactive peptides from fish and beef. Chromatographic profiles of the fish and beef hydrolysates obtained by RP-HPLC using a dC18 column (A and B, respectively). The peaks marked by arrows show the target peptides termed F21 and B34. ESI-TOF mass spectra showing multiple charged molecular ions of the peptides F21 and B34 (C and D, respectively). The molecular masses of F21 and B34 were calculated based on the most abundant peaks.

Figure 2.
Prediction of the secondary structure of the F21 and B34 isolated in this study. Secondary structures of F21 (A) and B34 (B) peptides were predicted using the GOR method; c, e, and h represent the secondary structures of random coil, exceeded strand, and helix, respectively.
Figure 2.
Prediction of the secondary structure of the F21 and B34 isolated in this study. Secondary structures of F21 (A) and B34 (B) peptides were predicted using the GOR method; c, e, and h represent the secondary structures of random coil, exceeded strand, and helix, respectively.


{kind=link}
{kind=link}
Table 1.
Antimicrobial spectra of the hydrolysates F21 and B34.
| Indicator Strain | MIC (μg/mL) | |||
|---|---|---|---|---|
| Fish Hydrolysates | F21 | Beef Hydrolysates | B34 | |
| Enterococcus faecalis JCM 5803T | 800 | 46 | 800 | NA3 |
| Listeria innocua ATCC 33090T | 400 | 23 | 400 | NA3 |
| Escherichia coli JM109 | 400 | 46 | 400 | 184 |
| Bacillus coagulans JCM 2257T | 400 | 23 | 400 | NA3 |
| Pseudomonas putida ATCC 12633T | 800 | 23 | NA2 | NA3 |
| Salmonella enterica subsp. enterica NBRC 13245T | 400 | 184 | NA2 | NA3 |
| Proteus vulgaris F24B | 400 | NA1 | 800 | 91.6 |
JCM, Japan Collection of Microorganisms (Wako, Japan); ATCC, American Type Culture Collection (Manassas, VA, USA); NBRC, National Institute of Technology and Evaluation (NITE) Biological Resource Center (Chiba, Japan). NA1 (no activity) > 184 μg/mL, NA2 > 800 μg/mL and NA3 > 184 μg/mL.
Table 2.
ACE inhibitory activity and antioxidant properties of the hydrolysates F21 and B34.
| Sample | IC50 (µg/mL) | ||
|---|---|---|---|
| ACE | Antioxidant | TEAC | |
| Fish hydrolysates | 304.5 | 470.4 | 0.14 |
| F21 | 7.3 | 389.9 | 0.17 |
| Beef hydrolysates | 265.6 | 531.0 | 0.12 |
| B34 | 5.8 | negative | negative |
IC50 was defined as the protein concentration required to inhibit 50% ACE for ACE inhibitory activity and to inhibit 50% of DPPH radicals for antioxidant activity. TEAC indicates the Trolox equivalent antioxidant capacity obtained with the IC50 Trolox/IC50 sample.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Maky, M.A.; Zendo, T. Generation and Characterization of Novel Bioactive Peptides from Fish and Beef Hydrolysates. Appl. Sci. 2021, 11, 10452. https://doi.org/10.3390/app112110452
AMA Style
Maky MA, Zendo T. Generation and Characterization of Novel Bioactive Peptides from Fish and Beef Hydrolysates. Applied Sciences. 2021; 11(21):10452. https://doi.org/10.3390/app112110452
Chicago/Turabian StyleMaky, Mohamed Abdelfattah, and Takeshi Zendo. 2021. "Generation and Characterization of Novel Bioactive Peptides from Fish and Beef Hydrolysates" Applied Sciences 11, no. 21: 10452. https://doi.org/10.3390/app112110452
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.