
Mathematical Modeling to Estimate Photosynthesis: A State of the Art

 ,
,  ,
,  ,
,  ,
,  ,
,  and
and 
Abstract
:1. Introduction
1.1. Photosynthesis Process
Methods for Inferring Photosynthesis
1.2. Mathematical Modeling
- Definitions;
- Systems analysis;
- Modeling;
- Simulation;
- Validation.
2. Techniques
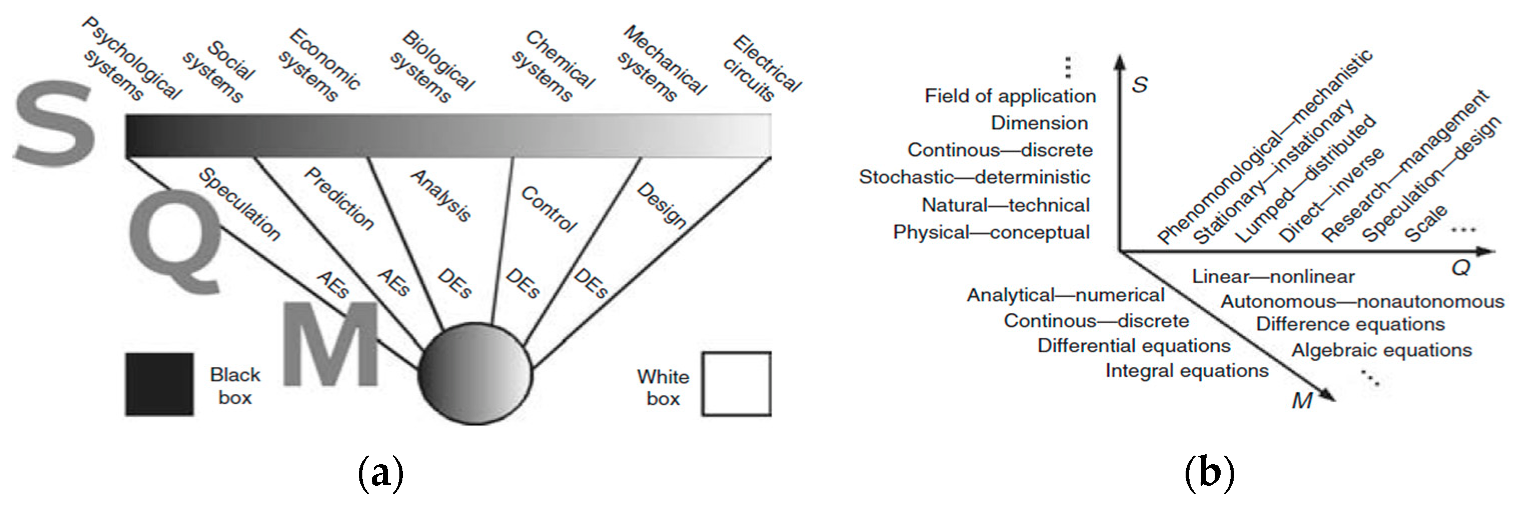
- (a)
- The models had to estimate gross photosynthesis, net photosynthesis, photosynthetic rate, biomass, or carbon assimilation. Wohlfahrt & Gu [50] reviewed the photosynthesis background and the associated terminology; they showed that different definitions and names are used for photosynthesis in literature. Then, as the understanding of photosynthesis has deepened, the terminologies and definitions of photosynthesis have also evolved [51]. In this work, Gross photosynthesis is defined as the total energy that plants fix during the photosynthesis process, while net photosynthesis is the total fixation of CO2 [52]. The photosynthetic rate refers to the amount of CO2 absorbed by a plant per unit of leaf area and unit of time. Biomass results from the net accumulation of assimilated CO2 throughout the growth cycle [53]. Lastly, carbon assimilation is the amount of CO2 stored in the plant. These terms are related to photosynthesis since plants can capture CO2 from the atmosphere and metabolize it through the photosynthetic process to obtain sugars and other compounds that are required for the normal development of their life cycle [26,54]. Then, the series of definitions described above refers to the estimation of photosynthesis.
- (b)
- The articles had to have a validation method to check if the different models used in the existing literature yielded similar results among other accredited models or with a photosynthesis measurement device on the market. This is important because one of the steps for modeling development is validation [55,56]. Validation is defined as the comparison of the model predictions with the observed values of the actual process to determine if the model fits its purpose; the validation results are a necessary step in the acceptance of the model [57,58,59]. Validation can also be defined as a demonstration that a model has acceptable predictive accuracy in that domain within a specific application. The quantitative precision of the predictions is often referred to as the “validity” of the model. Generally, it refers to the ability of a model to accurately predict the results of a particular experimental or observational scenario [44].
- -
- They did not estimate gross photosynthesis, net photosynthesis, photosynthetic rate, biomass, or carbon assimilation; instead, they modeled chlorophyll, fluorescence, or stomatal conductance, among others.
- -
- -
- -
- -
- -
- Some authors present a mathematical formulation of the model; however, they do not present validation or results obtained [70].
- -
- -
- -
3. Discussion
- i.
- To have the minimum number of model parameters with a reasonable error.
- ii.
- To have the simplest form with minimal error.
- iii.
- To be based on physical, chemical, and biological laws as much as possible.
- iv.
- To have the minimum deviation between the predicted and empirical values.
- v.
- To have the minimum output variance.
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| a | Leaf’s age Equation (5) |
| a* | Total concentration of PSUs Equations (14) and (24) |
| A | CO2 assimilation Equation (1); Photosynthetic rate Equations: (10), (12), (19) and (22); Photosynthesis Equation (18); Net photosynthesis Equation (11) |
| Ac | Assimilation rate limited by activation and kinetic properties of the Rubisco enzyme Equations (12) and (22) |
| Aj | Net photosynthesis rate limited by light Equations (12) and (22) |
| An | CO2 assimilation Equation (23); Photosynthetic rate Equations (26) and (38); Photosynthesis Equation (9) |
| age | Age of plant Equation (31) |
| B | Biomass Equation (29) |
| C | Carbon dioxide in the leaf Equations: (1), (10), (19), (25) and (30); Carbon dioxide from the atmosphere Equations (2), (6), (8), (18), (20), (21) and (34) |
| C1 | CO2 Concentration in the leaf Equation (3) |
| Ca | CO2 in the atmosphere Equations (12) and (23) |
| Cbs | CO2 concentration Equation (9) |
| Cc | CO2 partial pressure at the Rubisco carboxylate sites Equation (19) |
| Cc | CO2 offset point Equation (3) |
| Ci | CO2 partial pressures Equations (10) and (11); Internal CO2 concentration in the leaf Equation (23); Carbon dioxide from the atmosphere Equation (22) |
| Cl | Chlorophyll Equation (13) |
| CLA | Cumulative leaf area of entire crown Equation (26) |
| Cs | CO2 concentration on the leaf surface Equation (23) |
| CUO | Accumulated absorption O3 Equation (23) |
| D | Conversion coefficient Pg Equations (6) and (20); Stem diameter Equation (29) |
| DO2 | Dissolved oxygen concentration Equations (35), (36) and (38) |
| E | Irradiance Equations (13) and (32) |
| E1 | Light utilization efficiency Equation (3) |
| Ea | Start of photosynthesis saturation Equation (17) |
| Ec | CO2 utilization efficiency Equation (3) |
| EUR | Radiation use efficiency Equation (34) |
| EXT | Extinction coefficient of PAR inside the cover Equation (34) |
| f | PS2 fraction Equation (32) |
| F′ | Fluorescence Equation (37) |
| f (I, T) | Function of light and temperature in the respiration of the micro algae Equation (37) |
| f (age) | Age function Equation (31) |
| f (PPFD) | Photosynthetic photon flux density function Equation (31) |
| f(SWC) | Soil water content function Equation (31) |
| f(T1) | Temperature function Equation (3) |
| f(Ta) | Air temperature function Equation (31) |
| f(Ts) | Soil temperature function Equation (31) |
| f(VPD) | vapor pressure deficit function Equation (31) |
| F(z) | Cumulative density leaf area between the surface of the canopy and height z Equation (27) |
| f1(T) | Absolute Maximum mathematical function due to leaf’s temperature Equation (5) |
| f2 (a) | Function that describes the pattern for the age of the leaf Equation (5) |
| FAO3 | Response relationship treatment to control photosynthesis Equation (23) |
| FC02 | Relative dimensionless factor that estimates the effect on the growth of the concentration of CO2 in the air Equation (34) |
| Fi,d | Photosynthesis gross Equation (33) |
| F’m | Fluorescence flux Equation (37) |
| gb | Conductance of the boundary layer of the leaf Equation (23) |
| gmin | Minimum stomatal conductance Equation (12) |
| gn | Stomatal conductance Equation (12) |
| gs | Stomatal conductance Equation (23) |
| H | Heigh Equation (29) |
| i | Serial number Equation (26) |
| I | Light intensity or irradiance Equations (2), (15), (16), (17), (30), (36) and (39) |
| I* | CO2 photosynthetic compensation point Equation (18) |
| i,d | Time, day Equation (33) |
| IAF | Leaf’s area index Equation (34) |
| Iav | Irradiance Equation (35) |
| Ic | Irradiance concentration Equation (15) |
| Il | Irradiance Equation (21) |
| Io | Light flux density in foliage Equation (20); Light intensity Equation (27); Quanta absorbed by the light Equation (11) |
| Ip | PAR incidence Equation (9) |
| IPAR | Radiation Equation (34) |
| IPPF | Photosynthetic photon flux Equation (25) |
| I’R | PAR intensity of half-saturation Equation (28) |
| J | Potential rate of full-chain electron transport Equations (10), (12) and (22) |
| J (CO2, I) | Maximum photosynthesis rate with Cbs saturation at a given irradiance Equation (9) |
| Jm | Electron transport potential rate Equation (9) |
| k | Represents the fractional rate of photosynthesis vs. irradiance Equations (12) and (13) |
| K | Constant rate of photoclimatory changes in cellular chlorophyll Equations (24) and (26) |
| K (I) | Light extinction coefficient Equations (6), (20) and (27) |
| K (O) | Function of enzymatic kinetics Equation (12) |
| K1 [RuBP] | Carbon fixed by photosynthesis Equation (7) |
| K2 [RuBP]2 | Carbon used by photorespiration Equation (7) |
| K5 [TP] | Carbon used by dark respiration Equation (7) |
| Ka | PPFD extinction on chlorophyll Equation (11) |
| Kc | Michaelis constant for carboxylation Equations (1), (9), (10), (18) and (22) |
| KmC | Rubisco constant for CO2 Equation (19) |
| KmO | Rubisco constant for O2 Equation (19) |
| Ko | Michaelis constant for oxygenation Equations (1), (9), (10), (18) and (22) |
| KO3 | Leaf resistance ratio for O3 Equation (23) |
| kp, ks | Constan Equations (14) and (24) |
| L | Foliar area index Equation (20) |
| LAI | Foliar area index Equation (6) |
| LFmax | Maximum leaf photosynthesis rate Equation (6) |
| LMA | Leaf’s mass per area Equations (26) and (38) |
| m | Leaf light transmission coefficient Equations (6), (20) and (27) |
| msat | Rate of light saturated carboxylation Equation (11) |
| Np | Leaf nitrogen content Equation (11) |
| nsat | Leaf nitrogen content Equation (11) |
| O | Oxygen from the atmosphere Equations (9) and (18), Oxygen in the leaf Equations (1), (10), (12), (13), (17), (19), (22) and (24) |
| O(O) | O2 partial pressures Equation (10) |
| O3 | Oxygen in the leaf Equation (23) |
| Obs | O2 concentration in the bundle-sheath cell Equations (9) and (19) |
| P | Photosynthetic rate Equations (8), (13), (14), (15), (17), (24) and (37) |
| P* | Photosynthesis rate without photoinhibition Equation (4) |
| P(T) | Photosynthesis reduction factor Equation (20) |
| PAR | Radiation Equations (26), (34) and (38) |
| PARt | PAR incident on top of crown Equation (26) |
| PARvp | PAR intensity reaching the epidermis of the leaf Equation (28) |
| PCARB | Potential biomass accumulation per plant Equation (34) |
| Pg | Total gross photosynthesis Equations (6), (20) and (27) |
| Pgmax | Maximum gross photosynthesis Equation (25) |
| PGRED(T) | Function to modify Pg, below suboptimal temperatures Equation (6) |
| pH | Degree of acidity or alkalinity of the plant Equations (35), (36) and (39) |
| Pij | Photosynthesis Equation (28) |
| P l,g | Photosynthetic rate Equation (21) |
| PLTPOP | Plant density per unit area Equation (34) |
| Pm | Maximum photosynthesis rate Equation (14) |
| Pm (O) | Oxygen release rate Equation (24) |
| Pm (T, a) | Maximum net photosynthesis rate per unit of leaf area over all PPFD levels, temperature and ages Equation (5) |
| Pm (T, C, N) | Photosynthetic value with saturation of Il Equation (21) |
| Pmax | Maximum photosynthetic rate Equations (2) and (3); Specific velocity coefficient for photosynthesis calculated by a saturated speed-dependent hyperbolic function of light and is a function of temperature Equations (17) and (28) |
| Pmm | Maximum endogenous photosynthesis capacity Equation (25) |
| Pn | Net photosynthetic rate Equations (2), (7), (16), (30) and (31) |
| PnCO2 | Net photosynthesis rate limited by CO2 Equation (25) |
| Pnl | Photosynthesis rate Equation (25) |
| Pn1max | Maximum net photosynthesis Equation (25) |
| Pnmax | Maximum net photosynthesis Equation (31) |
| PO2 | Net photosynthesis rate Equations (36) and (39) |
| PO2 (DO2) | Net photosynthesis rate in function of dissolved oxygen concentration Equations (36) and (39) |
| PO2 (I) | Net photosynthesis rate in function of irradiance Equations (36) and (39) |
| PO2 (pH) | Net photosynthesis rate in function of pH Equations (36) and (39) |
| PO2(T) | Net photosynthesis rate in function of temperature Equations (36) and (39) |
| Rate of gross photosynthesis Equation (32) | |
| PPFD | Flux density of photosynthetic photon Equations (6) and (31) |
| Ppot | Photosynthetic potential rate Equation (3) |
| Qe | Leaf’s quantum efficiency Equation (6) |
| qp | Photochemical cooling, a measure of the fraction of open reaction centers under ambient photon precipitation rates. Equation (32) |
| R | Leaf’s breathing Equation (8) |
| r*m | Disappearance of PSU activated Equations (14) and (24) |
| Ra | Radiation Equation (33) |
| RCLAI | Relative area of the accumulated leaf Equation (26) |
| Rd | Dark respiration Equations (1), (2), (9), (25), (26) and (31) |
| Rd (C) | Respiratory CO2 released Equation (18) |
| Rd (I) | Leaf respiration in light or day Equations (12) and (22) |
| RDINC | Relative depth in the crown Equations (26) and (38) |
| Rlight | Nonphotorespiratory mitochondrial CO2 release in light Equation (16) |
| RO2 | Photosynthesis rate Equation (35) |
| RO2(DO2) | Photosynthesis rate in function of dissolved oxygen concentration Equation (35) |
| RO2(I) | Leaf respiration in light or day Equation (39) |
| RO2(Iav) | Photosynthesis rate in function of irradiance Equation (35) |
| RO2(pH) | Photosynthesis rate in function of pH Equation (35) |
| RO2(T) | Photosynthesis rate in function of temperature Equation (35) |
| RuBP | Ribulose bisphosphate Equation (7) |
| Sc/o | Relative CO2/O2 specificity factor for Rubisco Equation (19) |
| SWC | Soil water content Equation (31) |
| t | Time Equations (4) and (24), Temperature Equation (28) |
| T | Leaf’s temperature Equations (5), (10), (20) and (22), Ambient temperature Equations (6), (15), (18), (21), (23), (30), (33), (34), (35), (36) and (39) |
| T1 | Ambient temperature Equation (3) |
| Ta | Ambient temperature Equation (31) |
| Tair | Ambient temperature Equation (38) |
| TS | soil temperature Equation (31) |
| TP | Triose phosphate Equation (7) |
| v | Stoichiometric factor Equation (37) |
| Vcm | Maximum saturated CO2 carboxylation rate Equation (9) |
| Vcmax | Maximum carboxylation rate Equations (1), (10), (18), (19) and (22) |
| Vm | Maximum carboxylation rate Equation (12) |
| VPD | Pressure steam Equation (26); Vapor pressure deficit Equations (30), (31) and (38) |
| Wc | Minimum carboxylation rates limited by Rubisco activity Equation (9) |
| WC | Assimilation rate limited by activation and kinetic properties of the Rubisco enzyme Equation (10) |
| Wj | RuBP regeneration Equation (9) |
| WJ | Rate limited by the regeneration capacity of Ribulose bisphosphate (RuBP) Equation (10) |
| Wp | Capacity of utilization of the products of photosynthesis Equation (10) |
| x | Maximum per crop Equation (33) |
| x* | Functional activation fraction PSUs Equation (24) |
| Xe* | PSU steady state fraction functions activated under continuous illumination Equation (14) |
| z | Height Equation (27) |
| α | Quantum efficiency Equations (2), (9), (11) and (21) |
| α (Cl) | Specific absorption of Chlorophyll Equation (13) |
| α (I) | Light utilization efficiency Equations (8), (20) and (27) |
| Photochemical efficacy of the leaf in the absence of oxygen Equation (25) | |
| Γ | CO2 compensation point in the absence of dark breathing Equations (10), (12) and (22) |
| * | Compensation point in the absence of dark Equations (1) and (19); CO2 partial pressure for the compensation of oxygenation and carboxylation reactions Equation (11) |
| ε | Radiation use efficiency Equation (33) |
| nPS2 | Relationship between PS2 reaction centers and chlorophyll Equation (32) |
| θ | Curvature factor dimensionless 0 > θ < 1 Equations (8), (21) and (37) |
| θ(C) | Relationship between physical and total resistance to CO2 diffusion Equation (2) |
| θ(C, I) | Degree of curvature of the CO2 response of net light-saturated photosynthesis Equation (25) |
| σ | Model Parameter Equation (37) |
| σPS2 | Functional absorption of PS2 Equation (32) |
| τ | Leaf conductance to CO2 transfer Equations (8), (9) and (20) |
| ϕ | Quantum performance of oxygen evolution Equation (13) |
| ϕC | Irradiance-dependent parameters Equation (32) |
| Φp | Light intensity Equation (37) |
| ΦRC | Quantum performance on the PS2 Equation (32) |
| ψ | Solar elevation angle Equation (26) |
| Ψ | Light inhibition function Equation (4) |
References
- Marín, M.A. Control Climático y Ciclo de Cultivo. Horticultura 2001, 152, 28–34. [Google Scholar]
- Stirbet, A.; Lazár, D.; Guo, Y.; Govindjee, G. Photosynthesis: Basics, History and Modelling. Ann. Bot. 2020, 126, 511–537. [Google Scholar] [CrossRef]
- Daniell, H.; Lin, C.-S.; Yu, M.; Chang, W.-J. Chloroplast Genomes: Diversity, Evolution, and Applications in Genetic Engineering. Genome Biol. 2016, 17, 134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blankenship, R.E. Molecular Mechanisms of Photosynthesis; John Wiley & Sons: New York, NY, USA, 2021. [Google Scholar]
- Maxwell, K.; Johnson, G.N. Chlorophyll Fluorescence—A Practical Guide. J. Exp. Bot. 2000, 51, 659–668. [Google Scholar] [CrossRef] [PubMed]
- Tillery, B.; Enger, E.; Ross, F. Integratsssed Science; Higher Education; McGraw-Hill: New York, NY, USA, 2012. [Google Scholar]
- Shipman, J.; Wilson, J.D.; Higgins, C.A.; Lou, B. An Introduction to Physical Science; Cengage Learning: Boston, MA, USA, 2020. [Google Scholar]
- Azcon-Bieto, J.; Talón, M. Fundamentos de Fisiología Vegetal; McGraw-Hill: Interamericana de España, Barcelona, Spain, 2000. [Google Scholar]
- Cheng, Y.; He, D. A Photosynthesis Continuous Monitoring System for CAM Plants. Int. J. Agric. Biol. Eng. 2019, 12, 141–146. [Google Scholar] [CrossRef] [Green Version]
- West-Eberhard, M.J.; Smith, J.A.C.; Winter, K. Photosynthesis, Reorganized. Science 2011, 332, 311–312. [Google Scholar] [CrossRef]
- Fereres Castiel, E.; Martín, V.; Francisco, J. Fitotecnia: Principios de Agronomía para Una Agricultura Sostenible; Mundi-Prensa Libros: Madrid, Spain, 2017. [Google Scholar]
- Taiz, L.; Zeiger, E.; Møller, I.M.; Murphy, A. Plant Physiology and Development; Sinauer Associates Incorporated: Sunderland, MA, USA, 2015. [Google Scholar]
- Climático, C.; Biodiversidad, Y. Documento Técnico V Del IPCC; IPCC: Geneva, Switzerland, 2002. [Google Scholar]
- Bidwell, R.G.S. Fisiología Vegetal; AGT: Mexico City, Mexico, 1993. [Google Scholar]
- Murchie, E.H.; Pinto, M.; Horton, P. Agriculture and the New Challenges for Photosynthesis Research. New Phytol. 2009, 181, 532–552. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Xiao, X.; Wu, X.; Zhou, S.; Zhang, G.; Qin, Y.; Dong, J. A Global Moderate Resolution Dataset of Gross Primary Production of Vegetation for 2000–2016. Sci. Data 2017, 4, 170165. [Google Scholar] [CrossRef] [Green Version]
- Ryu, Y.; Berry, J.A.; Baldocchi, D.D. What Is Global Photosynthesis? History, Uncertainties and Opportunities. Remote Sens. Environ. 2019, 223, 95–114. [Google Scholar] [CrossRef]
- Wu, A.; Hammer, G.L.; Doherty, A.; von Caemmerer, S.; Farquhar, G.D. Quantifying Impacts of Enhancing Photosynthesis on Crop Yield. Nat. Plants 2019, 5, 380–388. [Google Scholar] [CrossRef]
- Morales, F.; Ancín, M.; Fakhet, D.; González-Torralba, J.; Gámez, A.L.; Seminario, A.; Soba, D.; Ben Mariem, S.; Garriga, M.; Aranjuelo, I. Photosynthetic Metabolism under Stressful Growth Conditions as a Bases for Crop Breeding and Yield Improvement. Plants 2020, 9, 88. [Google Scholar] [CrossRef] [Green Version]
- Sinclair, T.R.; Rufty, T.W.; Lewis, R.S. Increasing Photosynthesis: Unlikely Solution for World Food Problem. Trends Plant Sci. 2019, 24, 1032–1039. [Google Scholar] [CrossRef]
- Shevela, D.; Bjorn, L.O. Photosynthesis: Solar Energy for Life; World Scientific Publishing: Singapore, 2018. [Google Scholar]
- Tenhunen, J.D.; Weber, J.A.; Yocum, C.S.; Gates, D.M. Development of a Photosynthesis Model with an Emphasis on Ecological Applications. Oecologia 1976, 26, 101–119. [Google Scholar] [CrossRef]
- Field, C.B.; Ball, J.T.; Berry, J.A. Photosynthesis: Principles and Field Techniques. In Plant Physiological Ecology; Springer: New York, NY, USA, 2000; pp. 209–253. [Google Scholar]
- Wolfbeis, O.S. Materials for Fluorescence-Based Optical Chemical Sensors. J. Mater. Chem. 2005, 15, 2657–2669. [Google Scholar] [CrossRef]
- Millan-Almaraz, J.R.; Guevara-Gonzalez, R.G.; Troncoso, R.R.-; Osornio-Rios, R.A.; Torres-Pacheco, I. Advantages and Disadvantages on Photosynthesis Measurement Techniques: A Review. Afr. J. Biotechnol. 2009, 8, 7340–7349. [Google Scholar] [CrossRef]
- Busch, F.A. Photosynthetic Gas Exchange in Land Plants at the Leaf Level. In Photosynthesis; Springer: New York, NY, USA, 2018; pp. 25–44. [Google Scholar]
- Walker, B.J.; Busch, F.A.; Driever, S.M.; Kromdijk, J.; Lawson, T. Survey of Tools for Measuring In Vivo Photosynthesis. Photosynthesis 2018, 1770, 3–24. [Google Scholar]
- Bassow, S.L.; Bazzaz, F.A. How Environmental Conditions Affect Canopy Leaf-Level Photosynthesis in Four Deciduous Tree Species. Ecology 1998, 79, 2660–2675. [Google Scholar] [CrossRef]
- Schulze, E.-D. A New Type of Climatized Gas Exchange Chamber for Net Photosynthesis and Transpiration Measurements in the Field. Oecologia 1972, 10, 243–251. [Google Scholar] [CrossRef]
- Hunt, S. Measurements of Photosynthesis and Respiration in Plants. Physiol. Plant. 2003, 117, 314–325. [Google Scholar] [CrossRef]
- Takahashi, M.; Ishiji, T.; Kawashima, N. Handmade Oxygen and Carbon Dioxide Sensors for Monitoring the Photosynthesis Process as Instruction Material for Science Education. Sens. Actuators B Chem. 2001, 77, 237–243. [Google Scholar] [CrossRef]
- Biosciences LI-6400XT System. Photosyntesis, Fluorescence, Respiration. 2014. Available online: https://www.licor.com/env/products/photosynthesis/LI-6400XT/ (accessed on 28 March 2022).
- Kawachi, N.; Sakamoto, K.; Ishii, S.; Fujimaki, S.; Suzui, N.; Ishioka, N.S.; Matsuhashi, S. Kinetic Analysis of Carbon-11-Labeled Carbon Dioxide for Studying Photosynthesis in a Leaf Using Positron Emitting Tracer Imaging System. IEEE Trans. Nucl. Sci. 2006, 53, 2991–2997. [Google Scholar] [CrossRef]
- Taiz, L. Plant Physiology, 5th ed.; Sinauer Associates Inc.: Sunderland, MA, USA, 2006. [Google Scholar]
- Hermand, J.-P. Photosynthesis of Seagrasses Observed in Situ from Acoustic Measurements. In Proceedings of the Oceans’ 04 MTS/IEEE Techno-Ocean’04 (IEEE Cat. No. 04CH37600), Kobe, Japan, 9–12 November 2004; Volume 1, pp. 433–437. [Google Scholar]
- Velten, K. Mathematical Modeling and Simulation: Introduction for Scientists and Engineers; John Wiley & Sons: New York, NY, USA, 2009. [Google Scholar]
- Hesketh, J.D. Predicting Photosynthesis for Ecosystem Models: Volume II; CRC Press: Boca Raton, FL, USA, 2017; ISBN 978-1-351-07595-4. [Google Scholar]
- Zufferey, V.; Murisier, F.; Schultz, H.R. A Model Analysis of the Photosynthetic Response of Vitis vinifera L. Cvs Riesling and Chasselas Leaves in the Field: I. Interaction of Age, Light and Temperature. VITIS 2000, 39, 19–26. [Google Scholar]
- Polerecky, L.; Lott, C.; Weber, M. In Situ Measurement of Gross Photosynthesis Using a Microsensor-Based Light-Shade Shift Method. Limnol. Oceanogr. Methods 2008, 6, 373–383. [Google Scholar] [CrossRef]
- Torres-Pacheco, I.; Padilla-Medina, J.A.; Osornio-Rios, R.A.; de Jesus Romero-Troncoso, R.; Villasentilde, C.; Rico-Garcia, E.; Guevara-Gonzalez, R.G. Description of Photosynthesis Measurement Methods in Green Plants Involving Optical Techniques, Advantages and Limitations. Afr. J. Agric. Res. 2011, 6, 2638–2647. [Google Scholar]
- Magney, T.S.; Frankenberg, C.; Fisher, J.B.; Sun, Y.; North, G.B.; Davis, T.S.; Kornfeld, A.; Siebke, K. Connecting Active to Passive Fluorescence with Photosynthesis: A Method for Evaluating Remote Sensing Measurements of Chl Fluorescence. New Phytol. 2017, 215, 1594–1608. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aziz, H.A.; Dianursanti, D. Production of Microalgae Nannochloropsis oculata Biomass in a Bubble Column Photobioreactor with Integrated Lighting through Adjustment of Light Intensity and Air Flow Rate. AIP Conf. Proc. 2018, 2024, 020040. [Google Scholar] [CrossRef]
- Kitić, G.; Tagarakis, A.; Cselyuszka, N.; Panić, M.; Birgermajer, S.; Sakulski, D.; Matović, J. A New Low-Cost Portable Multispectral Optical Device for Precise Plant Status Assessment. Comput. Electron. Agric. 2019, 162, 300–308. [Google Scholar] [CrossRef]
- Thornley, J.H.M.; France, J. Mathematical Models in Agriculture: Quantitative Methods for the Plant, Animal and Ecological Sciences; CABI: Wallingford, UK, 2007; ISBN 978-0-85199-010-1. [Google Scholar]
- Ljung, L. Black-Box Models from Input-Output Measurements. In Proceedings of the IMTC 2001. In Proceedings of the 18th IEEE Instrumentation and Measurement Technology Conference. Rediscovering Measurement in the Age of Informatics (Cat. No.01CH 37188), Budapest, Hungary, 21–23 May 2001; Volume 1, pp. 138–146. [Google Scholar]
- Hu, P.; Sun, Y.; Zhang, Y.; Dong, J.; Zhang, X. Application of WDNN for Photosynthetic Rate Prediction in Greenhouse. In Proceedings of the IEEE 2nd International Conference on Big Data, Artificial Intelligence and Internet of Things Engineering (ICBAIE), Nanchang, China, 26–28 March 2021; pp. 331–336. [Google Scholar]
- Guidotti, R.; Monreale, A.; Ruggieri, S.; Turini, F.; Giannotti, F.; Pedreschi, D. A Survey of Methods for Explaining Black Box Models. ACM Comput. Surv. 2018, 51, 93. [Google Scholar] [CrossRef] [Green Version]
- Ljung, L.; Glad, T. Modeling of Dynamic Systems; PTR Ptrnyice-Hall: Hoboken, NJ, USA, 1994. [Google Scholar]
- Von Caemmerer, S. Steady-State Models of Photosynthesis. Plant Cell Environ. 2013, 36, 1617–1630. [Google Scholar] [CrossRef]
- Wohlfahrt, G.; Gu, L. The Many Meanings of Gross Photosynthesis and Their Implication for Photosynthesis Research from Leaf to Globe. Plant Cell Environ. 2015, 38, 2500–2507. [Google Scholar] [CrossRef] [Green Version]
- Gest, H. History of the Word Photosynthesis and Evolution of Its Definition. In Discoveries in Photosynthesis; Govindjee, Beatty, J.T., Gest, H., Allen, J.F., Eds.; Advances in Photosynthesis and Respiration; Springer: Dordrecht, The Netherlands, 2005; pp. 39–42. ISBN 978-1-4020-3324-7. [Google Scholar]
- Carril, E.P.-U. Fotosíntesis: Aspectos Básicos. REDUCA 2011, 2, 39. [Google Scholar]
- Ruiz-Torres, N.; Rincon, F.; Martinez, J.; Reyna, J.; Esquivel, M. Sistemas de producción y asimilación de CO2 EN la variedad jaguan de maíz criollo mejorado. In Proceedings of the XII Simposio Internacional y VII Congreso Nacional de Agricultura Sostenible, Puebla, Puebla, 11–16 November 2013; pp. 1872–1886, ISBN 978-607-715-178-4. [Google Scholar]
- Carvajal, M. Investigación Sobre la Absorción de CO2 por los Cultivos más Representativos; CSIC: Madrid, Spain, 2022. [Google Scholar]
- Barrales, L.V.; Peña, I.R.; de la Reguera, P.B.F. Nota Científica—Validación de Modelos: Un Enfoque Aplicado (Model Validation: An Applied Approach); Instituto de Investigaciones Agropecuarias: Santiago, Chile, 2004. [Google Scholar]
- Medina-Peralta, S.; Vargas-Villamil, L.; Navarro-Alberto, J.; Canul-Pech, C.; Peraza-Romero, S. Comparación de medidas de desviación para validar modelos sin sesgo, sesgo constante o proporcional. Univ. Cienc. 2010, 26, 255–263. [Google Scholar]
- Mayer, D.G.; Butler, D.G. Statistical Validation. Ecol. Model. 1993, 68, 21–32. [Google Scholar] [CrossRef]
- Mitchell, P.L. Misuse of Regression for Empirical Validation of Models. Agric. Syst. 1997, 54, 313–326. [Google Scholar] [CrossRef]
- Paez, T.L. Introduction to Model Validation. In Introduction to Model Validation; Sandia National Lab: Orlando, FL, USA, 2009; p. 11. [Google Scholar]
- Zhu, X.-G.; Wang, Y.; Ort, D.R.; Long, S.P. E-Photosynthesis: A Comprehensive Dynamic Mechanistic Model of C3 Photosynthesis: From Light Capture to Sucrose Synthesis. Plant Cell Environ. 2013, 36, 1711–1727. [Google Scholar] [CrossRef] [Green Version]
- Zhu, X.-G.; Song, Q.; Ort, D.R. Elements of a Dynamic Systems Model of Canopy Photosynthesis. Curr. Opin. Plant Biol. 2012, 15, 237–244. [Google Scholar] [CrossRef]
- Bernard, O.; Rémond, B. Validation of a Simple Model Accounting for Light and Temperature Effect on Microalgal Growth. Bioresour. Technol. 2012, 123, 520–527. [Google Scholar] [CrossRef]
- Bitaubé Pérez, E.; Caro Pina, I.; Pérez Rodríguez, L. Kinetic Model for Growth of Phaeodactylum Tricornutum in Intensive Culture Photobioreactor. Biochem. Eng. J. 2008, 40, 520–525. [Google Scholar] [CrossRef]
- Moisan, J.R.; Moisan, T.A.; Abbott, M.R. Modelling the Effect of Temperature on the Maximum Growth Rates of Phytoplankton Populations. Ecol. Model. 2002, 153, 197–215. [Google Scholar] [CrossRef]
- Rojano-Aguilar, A.; Arias, A.; Lopez-Cruz, I. Modelos Matemáticos de Hortalizas En Invernadero: Trascendiendo La Contemplación de La Dinámica de Cultivos. Rev. Chapingo Ser. Hortic. 2005, 11, 257–267. [Google Scholar] [CrossRef]
- Romero Cuéllar, J.; Torrente Trujillo, A.; Obregón Neira, N.; Torres, A. Acople de un modelo de conductancia estomática, fotosíntesis y transpiración para el cultivo de maracuyá (Passiflora edulis Sims). Acta Agronómica 2013, 62, 42–51. [Google Scholar]
- Walker, A.P.; Johnson, A.L.; Rogers, A.; Anderson, J.; Bridges, R.A.; Fisher, R.A.; Lu, D.; Ricciuto, D.M.; Serbin, S.P.; Ye, M. Multi-Hypothesis Comparison of Farquhar and Collatz Photosynthesis Models Reveals the Unexpected Influence of Empirical Assumptions at Leaf and Global Scales. Glob. Chang. Biol. 2021, 27, 804–822. [Google Scholar] [CrossRef] [PubMed]
- Mengoli, G.; Agustí-Panareda, A.; Boussetta, S.; Harrison, S.P.; Trotta, C.; Prentice, I.C. Ecosystem Photosynthesis in Land-Surface Models: A First-Principles Approach Incorporating Acclimation. J. Adv. Model. Earth Syst. 2022, 14, e2021MS002767. [Google Scholar] [CrossRef]
- Fee, E.J. A Numerical Model for the Estimation of Photosynthetic Production, Integrated Over Time and Depth, in Natural Waters1. Limnol. Oceanogr. 1969, 14, 906–911. [Google Scholar] [CrossRef]
- Thornley, J.H.M. Acclimation of Photosynthesis to Light and Canopy Nitrogen Distribution: An Interpretation. Ann. Bot. 2004, 93, 473–475. [Google Scholar] [CrossRef] [Green Version]
- Pettersson, G.; Ryde-Pettersson, U. A Mathematical Model of the Calvin Photosynthesis Cycle. Eur. J. Biochem. 1988, 175, 661–672. [Google Scholar] [CrossRef] [Green Version]
- Obeid, H.; Rendall, A.D. The Minimal Model of Hahn for the Calvin Cycle. arXiv 2018, arXiv:1812.00620. [Google Scholar] [CrossRef]
- Kim, D.; Kang, W.H.; Hwang, I.; Kim, J.; Kim, J.H.; Park, K.S.; Son, J.E. Use of Structurally-Accurate 3D Plant Models for Estimating Light Interception and Photosynthesis of Sweet Pepper (Capsicum annuum) Plants. Comput. Electron. Agric. 2020, 177, 105689. [Google Scholar] [CrossRef]
- Hinojo-Hinojo, C.; Castellanos, A.E.; Llano-Sotelo, J.; Peñuelas, J.; Vargas, R.; Romo-Leon, J.R. High Vcmax, Jmax and Photosynthetic Rates of Sonoran Desert Species: Using Nitrogen and Specific Leaf Area Traits as Predictors in Biochemical Models. J. Arid Environ. 2018, 156, 1–8. [Google Scholar] [CrossRef]
- Tsujimoto, K.; Hikosaka, K. Estimating Leaf Photosynthesis of C3 Plants Grown under Different Environments from Pigment Index, Photochemical Reflectance Index, and Chlorophyll Fluorescence. Photosynth. Res. 2021, 148, 33–46. [Google Scholar] [CrossRef]
- Bernacchi, C.J.; Pimentel, C.; Long, S.P. In Vivo Temperature Response Functions of Parameters Required to Model RuBP-Limited Photosynthesis. Plant Cell Environ. 2003, 26, 1419–1430. [Google Scholar] [CrossRef]
- Center, B.; Verma, B.P. A Fuzzy Photosynthesis Model for Tomato. Trans. ASAE 1997, 40, 815–821. [Google Scholar] [CrossRef]
- Valenzuela, I.C.; Baldovino, R.G.; Bandala, A.A.; Dadios, E.P. Optimization of Photosynthetic Rate Parameters Using Adaptive Neuro-Fuzzy Inference System (ANFIS). In Proceedings of the 2017 International Conference on Computer and Applications (ICCA), Doha, United Arab Emirates, 6–7 September 2017; pp. 129–134. [Google Scholar]
- Farquhar, G.D.; von Caemmerer, S.; Berry, J.A. A Biochemical Model of Photosynthetic CO2 Assimilation in Leaves of C3 Species. Planta 1980, 149, 78–90. [Google Scholar] [CrossRef] [Green Version]
- Farquhar, G.D.; von Caemmerer, S.; Berry, J.A. Models of Photosynthesis. Plant Physiol. 2001, 125, 42–45. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Chun, J.A.; Fleisher, D.; Reddy, V.; Timlin, D.; Resop, J. Parameter Estimation of the Farquhar—Von Caemmerer—Berry Biochemical Model from Photosynthetic Carbon Dioxide Response Curves. Sustainability 2017, 9, 1288. [Google Scholar] [CrossRef] [Green Version]
- Marshall, B.; Biscoe, P.V. A Model for C3 Leaves Describing the Dependence of Net Photosynthesis on Irradiance. J. Exp. Bot. 1980, 31, 29–39. [Google Scholar] [CrossRef]
- Kano, A.; Gavel, C.H.M.V. Design and Test of a Simulation Model of Tomato Growth and Yield in a Greenhouse. J. Jpn. Soc. Hortic. Sci. 1988, 56, 408–416. [Google Scholar] [CrossRef] [Green Version]
- Pahl-Wostl, C.; Imboden, D.M. DYPHORA—A Dynamic Model for the Rate of Photosynthesis of Algae. J. Plankton Res. 1990, 12, 1207–1221. [Google Scholar] [CrossRef]
- Lieth, J.H.; Pasian, C.C. A Model for Net Photosynthesis of Rose Leaves as a Function of Photosynthetically Active Radiation, Leaf Temperature, and Leaf Age. J. Am. Soc. Hortic. Sci. 1990, 115, 486–491. [Google Scholar] [CrossRef] [Green Version]
- Jones, J.W.; Dayan, E.; Allen, L.H.; van Keulen, H.; Challa, H. A Dynamic Tomato Growth and Yield Model (TOMGRO). Trans. ASAE 1991, 34, 663–672. [Google Scholar] [CrossRef]
- Hahn, B.D. Photosynthesis and Photorespiration: Modelling the Essentials. J. Theor. Biol. 1991, 151, 123–139. [Google Scholar] [CrossRef]
- Pachepsky, L.B.; Haskett, J.D.; Acock, B. An Adequate Model of Photosynthesis—I Parameterization, Validation and Comparison of Models. Agric. Syst. 1996, 50, 209–225. [Google Scholar] [CrossRef]
- Chen, D.-X.; Coughenour, M.B.; Knapp, A.K.; Owensby, C.E. Mathematical Simulation of C4 Grass Photosynthesis in Ambient and Elevated CO2. Ecol. Model. 1994, 73, 63–80. [Google Scholar] [CrossRef]
- Nikolov, N.T.; Massman, W.J.; Schoettle, A.W. Coupling Biochemical and Biophysical Processes at the Leaf Level: An Equilibrium Photosynthesis Model for Leaves of C3 Plants. Ecol. Model. 1995, 80, 205–235. [Google Scholar] [CrossRef]
- Kull, O.; Kruijt, B. Leaf Photosynthetic Light Response: A Mechanistic Model for Scaling Photosynthesis to Leaves and Canopies. Funct. Ecol. 1998, 12, 767–777. [Google Scholar] [CrossRef]
- Chen, J.M.; Liu, J.; Cihlar, J.; Goulden, M.L. Daily Canopy Photosynthesis Model through Temporal and Spatial Scaling for Remote Sensing Applications. Ecol. Model. 1999, 124, 99–119. [Google Scholar] [CrossRef] [Green Version]
- Han, B.-P. Photosynthesis–Irradiance Response at Physiological Level: A Mechanistic Model. J. Theor. Biol. 2001, 213, 121–127. [Google Scholar] [CrossRef]
- Rubio, F.C.; Camacho, F.G.; Sevilla, J.M.F.; Chisti, Y.; Grima, E.M. A Mechanistic Model of Photosynthesis in Microalgae. Biotechnol. Bioeng. 2003, 81, 459–473. [Google Scholar] [CrossRef] [Green Version]
- Ye, Z.-P. A New Model for Relationship between Irradiance and the Rate of Photosynthesis in Oryza Sativa. Photosynth. 2007, 45, 637–640. [Google Scholar] [CrossRef]
- Ye, Z.-P.; Suggett, D.J.; Robakowski, P.; Kang, H.-J. A Mechanistic Model for the Photosynthesis–Light Response Based on the Photosynthetic Electron Transport of Photosystem II in C3 and C4 Species. New Phytol. 2013, 199, 110–120. [Google Scholar] [CrossRef]
- Bernacchi, C.J.; Rosenthal, D.M.; Pimentel, C.; Long, S.P.; Farquhar, G.D. Modeling the Temperature Dependence of C3 Photosynthesis. In Photosynthesis in Silico: Understanding Complexity from Molecules to Ecosystems; Laisk, A., Nedbal, L., Govindjee, Eds.; Advances in Photosynthesis and Respiration; Springer: Dordrecht, The Netherlands, 2009; pp. 231–246. ISBN 978-1-4020-9237-4. [Google Scholar]
- Yin, X.; Struik, P.C. C3 and C4 Photosynthesis Models: An Overview from the Perspective of Crop Modelling. NJAS—Wagening. J. Life Sci. 2009, 57, 27–38. [Google Scholar] [CrossRef] [Green Version]
- Torres, J.C.; Ríos, E.; Peña, R.; Martínez, D.; Sánchez, P.; Reyes, F.; Romero, G. Evaluación del Modelo de Jones para Desarrollar un Sistema por Computadora que Permita Determinar el Crecimiento del Frijol en Ambiente Controlado. In Proceedings of the Congreso Anual 2010 de la Asociación de México de Control Automático, Puerto Vallarta, Jalisco, México, 6–8 October 2010; p. 7. [Google Scholar]
- Johnson, I.R.; Thornley, J.H.M.; Frantz, J.M.; Bugbee, B. A Model of Canopy Photosynthesis Incorporating Protein Distribution through the Canopy and Its Acclimation to Light, Temperature and CO2. Ann. Bot. 2010, 106, 735–749. [Google Scholar] [CrossRef] [Green Version]
- Egea, G.; Verhoef, A.; Vidale, P.L. Towards an Improved and More Flexible Representation of Water Stress in Coupled Photosynthesis–Stomatal Conductance Models. Agric. For. Meteorol. 2011, 151, 1370–1384. [Google Scholar] [CrossRef]
- Lombardozzi, D.; Sparks, J.P.; Bonan, G.; Levis, S. Ozone Exposure Causes a Decoupling of Conductance and Photosynthesis: Implications for the Ball-Berry Stomatal Conductance Model. Oecologia 2012, 169, 651–659. [Google Scholar] [CrossRef]
- García-Camacho, F.; Sánchez-Mirón, A.; Molina-Grima, E.; Camacho-Rubio, F.; Merchuck, J.C. A Mechanistic Model of Photosynthesis in Microalgae Including Photoacclimation Dynamics. J. Theor. Biol. 2012, 304, 1–15. [Google Scholar] [CrossRef]
- Janka, E.; Körner, O.; Rosenqvist, E.; Ottosen, C.-O. A Coupled Model of Leaf Photosynthesis, Stomatal Conductance, and Leaf Energy Balance for Chrysanthemum (Dendranthema grandiflora). Comput. Electron. Agric. 2016, 123, 264–274. [Google Scholar] [CrossRef]
- Liu, Q.; Xie, L.; Li, F. Dynamic Simulation of the Crown Net Photosynthetic Rate for Young Larix Olgensis Henry Trees. Forests 2019, 10, 321. [Google Scholar] [CrossRef] [Green Version]
- Hozumi, K.; Kirita, H. Estimation of the Rate of Total Photosynthesis in Forest Canopies. Shokubutsugaku Zasshi 1970, 83, 144–151. [Google Scholar] [CrossRef]
- Wetzel, R.L.; Neckles, H.A. A Model of Zostera Marina L. Photosynthesis and Growth: Simulated Effects of Selected Physical-Chemical Variables and Biological Interactions. Aquat. Bot. 1986, 26, 307–323. [Google Scholar] [CrossRef]
- De la Torre, D.d.P.; Cabrera, J.D.; Calderón, Ó.A.; Hernández, F.J.; Pérez, J.J. Estimación de biomasa y contenido de carbono de Pinus cooperi Blanco, en Pueblo Nuevo, Durango. Madera Bosques 2007, 13, 35–46. [Google Scholar] [CrossRef]
- Romdhonah, Y.; Fujiuchi, N.; Takahashi, N.; Nishina, H.; Takayama, K. Empirical Model for the Estimation of Whole-Plant Photosynthetic Rate of Cherry Tomato Grown in a Commercial Greenhouse. Environ. Control Biol. 2021, 59, 117–124. [Google Scholar] [CrossRef]
- Shimada, R.; Takahashi, K. Diurnal and Seasonal Variations in Photosynthetic Rates of Dwarf Pine Pinus Pumila at the Treeline in Central Japan. Arct. Antarct. Alp. Res. 2022, 54, 1–12. [Google Scholar] [CrossRef]
- Kolber, Z.; Falkowski, P.G. Use of Active Fluorescence to Estimate Phytoplankton Photosynthesis in Situ. Limnol. Oceanogr. 1993, 38, 1646–1665. [Google Scholar] [CrossRef]
- Jansen, D.M. Uso de integración según Gauss para calcular la fotosíntesis en modelos de Simulación de Crecimiento de Cultivos. Rev. Matemática Teoría Apl. 1995, 2, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Sau, F.; Cedrón, F.X.L.; Confalones, A.E.; Lizaso, J.I. Modelos de Simulación del Cultivo de Maíz: Fundamentos y Aplicaciones en España. Pastos 2012, 40, 117–138. [Google Scholar]
- Costache, T.A.; Acién Fernández, F.G.; Morales, M.M.; Fernández-Sevilla, J.M.; Stamatin, I.; Molina, E. Comprehensive Model of Microalgae Photosynthesis Rate as a Function of Culture Conditions in Photobioreactors. Appl. Microbiol. Biotechnol. 2013, 97, 7627–7637. [Google Scholar] [CrossRef]
- Ippoliti, D.; Gómez, C.; del Mar Morales-Amaral, M.; Pistocchi, R.; Fernández-Sevilla, J.M.; Acién, F.G. Modeling of Photosynthesis and Respiration Rate for Isochrysis Galbana (T-Iso) and Its Influence on the Production of This Strain. Bioresour. Technol. 2016, 203, 71–79. [Google Scholar] [CrossRef] [PubMed]
- Perin, G.; Bernardi, A.; Bellan, A.; Bezzo, F.; Morosinotto, T. A Mathematical Model to Guide Genetic Engineering of Photosynthetic Metabolism. Metab. Eng. 2017, 44, 337–347. [Google Scholar] [CrossRef]
- Liu, Q.; Xie, L.; Dong, L.; Li, F. Dynamic Simulation of the Multilayer Crown Net Photosynthetic Rate and Determination of the Functional Crown for Larch (Larix Olgensis) Trees. New For. 2021, 52, 1011–1035. [Google Scholar] [CrossRef]
- Sánchez Zurano, A.; Gómez Serrano, C.; Acién-Fernández, F.G.; Fernández-Sevilla, J.M.; Molina-Grima, E. Modeling of Photosynthesis and Respiration Rate for Microalgae–Bacteria Consortia. Biotechnol. Bioeng. 2021, 118, 952–962. [Google Scholar] [CrossRef]
- Jordan, D.B.; Ogren, W.L. The CO2/O2 Specificity of Ribulose 1,5-Bisphosphate Carboxylase/Oxygenase. Planta 1984, 161, 308–313. [Google Scholar] [CrossRef]
- Smith, N.G.; Dukes, J.S. Plant Respiration and Photosynthesis in Global-Scale Models: Incorporating Acclimation to Temperature and CO2. Glob. Change Biol. 2013, 19, 45–63. [Google Scholar] [CrossRef]
- Aguirre, E.A.; Pira, D.D.B.; Rodríguez, J.S.S.; Mesa, H.C.B.; Castro, D.A.V. Red de sensores inalámbricos para el monitoreo de variables agroecológicas en cultivos bajo invernadero. Publ. Investig. 2019, 13, 53–60. [Google Scholar] [CrossRef]
- Manrique, J.T.; Martínez y Ojeda, J.; Méndez, G.M.; Gil, A.M.; Martínez, A.R. LA Agroecología como Propuesta de Modelo de Producción Aplicado al Cultivo de Chile Habanero en Peto, Yucatán. Rev. Mex. Agronegocios 2014, 35, 969–978. [Google Scholar]
- Hawkesford, M.J.; Lorence, A.; Hawkesford, M.J.; Lorence, A. Plant Phenotyping: Increasing Throughput and Precision at Multiple Scales. Funct. Plant Biol. 2016, 44, v–vii. [Google Scholar] [CrossRef] [Green Version]
- Osuna, C.d.G. Sistema Basado en Brquitectura Híbrida FPGA-Uc Para la Medición Remota de las Variables Fisiológicas en Plantas. Master’s Thesis, Universidad Autonomia de Queretaro, Santiago, Mexico, 2013. [Google Scholar]
- Murchie, E.H.; Kefauver, S.; Araus, J.L.; Muller, O.; Rascher, U.; Flood, P.J.; Lawson, T. Measuring the Dynamic Photosynthome. Ann. Bot. 2018, 122, 207–220. [Google Scholar] [CrossRef] [Green Version]
- Athanasiou, K.; Dyson, B.C.; Webster, R.E.; Johnson, G.N. Dynamic Acclimation of Photosynthesis Increases Plant Fitness in Changing Environments. Plant Physiol. 2010, 152, 366–373. [Google Scholar] [CrossRef] [Green Version]
- Kaiser, E.; Morales, A.; Harbinson, J.; Kromdijk, J.; Heuvelink, E.; Marcelis, L.F.M. Dynamic Photosynthesis in Different Environmental Conditions. J. Exp. Bot. 2015, 66, 2415–2426. [Google Scholar] [CrossRef] [Green Version]
- Matthews, J.S.A.; Vialet-Chabrand, S.; Lawson, T. Acclimation to Fluctuating Light Impacts the Rapidity of Response and Diurnal Rhythm of Stomatal Conductance. Plant Physiol. 2018, 176, 1939–1951. [Google Scholar] [CrossRef] [Green Version]
- Poorter, H.; Fiorani, F.; Pieruschka, R.; Wojciechowski, T.; van der Putten, W.H.; Kleyer, M.; Schurr, U.; Postma, J. Pampered inside, Pestered Outside? Differences and Similarities between Plants Growing in Controlled Conditions and in the Field. New Phytol. 2016, 212, 838–855. [Google Scholar] [CrossRef]
- Vialet-Chabrand, S.; Matthews, J.S.A.; Simkin, A.J.; Raines, C.A.; Lawson, T. Importance of Fluctuations in Light on Plant Photosynthetic Acclimation. Plant Physiol. 2017, 173, 2163–2179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, V.; Sharma, A.; Soni, J.K.; Pawar, N. Physiological Response of C3, C4 and CAM Plants in Changeable Climate. Pharma Innov. J. 2017, 6, 70–79. [Google Scholar]
- Iversen, L.L.; Winkel, A.; Baastrup-Spohr, L.; Hinke, A.B.; Alahuhta, J.; Baattrup-Pedersen, A.; Birk, S.; Brodersen, P.; Chambers, P.A.; Ecke, F.; et al. Catchment Properties and the Photosynthetic Trait Composition of Freshwater Plant Communities. Science 2019, 366, 878–881. [Google Scholar] [CrossRef] [PubMed]
- Raven, J.A. Photosynthesis in Aquatic Plants. In Ecophysiology of Photosynthesis; Schulze, E.-D., Caldwell, M.M., Eds.; Springer Study Edition; Springer: Berlin/Heidelberg, 1995; pp. 299–318. ISBN 978-3-642-79354-7. [Google Scholar]
- Godin, C. Representing and Encoding Plant Architecture: A Review. Ann. For. Sci. 2000, 57, 413–438. [Google Scholar] [CrossRef]
- Johnson, G.N.; Lawson, T.; Murchie, E.H.; Raines, C. Photosynthesis in Variable Environments. J. Exp. Bot. 2015, 66, 2371–2372. [Google Scholar] [CrossRef] [Green Version]
- Nithia, S.; Shanthi, N. Enhance Solar UVB (280–320 nm) Radiation. Adv. Biores. 2018, 9, 232–239. [Google Scholar] [CrossRef]
- Toro, G.; Flexas, J.; Escalona, J.M. Contrasting Leaf Porometer and Infra-Red Gas Analyser Methodologies: An Old Paradigm about the Stomatal Conductance Measurement. Theor. Exp. Plant Physiol. 2019, 31, 483–492. [Google Scholar] [CrossRef]
- Gomez, L.; Montoya, E.C.; López, Y.; Riaño, N.M. Estimación de La Fotosíntesis En Plantas Completas y Validación de Un Modelo Matemático Para El Cafeto Coffea arabica L. In Proceedings of the Simposio Internacional Medición y Monitoreo de la Captura de Carbono en Ecosistemas Forestales, Valdivia, Chile, 18–20 October 2001. [Google Scholar]
- Bourgault, M.; James, A.T.; Dreccer, M.F.; Bourgault, M.; James, A.T.; Dreccer, M.F. Pot Size Matters Revisited: Does Container Size Affect the Response to Elevated CO2 and Our Ability to Detect Genotypic Variability in This Response in Wheat? Funct. Plant Biol. 2016, 44, 52–61. [Google Scholar] [CrossRef]
- Dambreville, A.; Griolet, M.; Rolland, G.; Dauzat, M.; Bédiée, A.; Balsera, C.; Muller, B.; Vile, D.; Granier, C.; Dambreville, A.; et al. Phenotyping Oilseed Rape Growth-Related Traits and Their Responses to Water Deficit: The Disturbing Pot Size Effect. Funct. Plant Biol. 2016, 44, 35–45. [Google Scholar] [CrossRef]
- Dornhoff, G.M.; Shibles, R.M. Varietal Differences in Net Photosynthesis of Soybean Leaves1. Crop Sci. 1970, 10, 42–45. [Google Scholar] [CrossRef]
- Anderson, J. Simulation: Methodology and Application in Agricultural Economics. Rev. Mark. Agric. Econ. 1974, 42, 9675. [Google Scholar]
- Gass, S.I. Feature Article—Decision-Aiding Models: Validation, Assessment, and Related Issues for Policy Analysis. Oper. Res. 1983, 31, 603–631. [Google Scholar] [CrossRef] [Green Version]
- Himmelblau, D.M. Process Analysis by Statistical Methods; John Wiley & Sons, Inc.: New York, NY, USA, 1970. [Google Scholar]
- Shannon, R.E. Systems Simulation; the Art and Science. In Proceedings of the 1998 Winter Simulation Conference, Washington, DC, USA, 13–16 December 1975. [Google Scholar]
- Gutiérrez Pulido, H.; de la Vara Salazar, R. Analysis and Design of Experiments; McGraw Hill: New York, NY, USA, 2004. [Google Scholar]
- Ubierna, N.; Cernusak, L.A. Preface: Advances in Modelling Photosynthetic Processes in Terrestrial Plants. Photosynth. Res. 2019, 141, 1–3. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Methods Used for Photosynthesis Estimation | Description |
|---|---|
| Destructive | Involves cutting a whole plant or only a portion of it to estimate the photosynthetic activity based on the accumulation of dry matter in the plant, from the point of germination until it is cut [25]. |
| Manometric | Consists of direct measurement of oxygen (O2) pressure or carbon dioxide (CO2) in an isolated chamber with photosynthetic organisms [30]. |
| Electrochemical | Uses electrochemical electrodes to measure O2, CO2, or pH in aqueous solutions of the sample to detect variations that depend on photosynthetic activity [30]. |
| Gas exchange | Consists of isolating the sample for analysis in a closed chamber to quantify the CO2 concentration [29,31]. Concentrated CO2 gas is detected by an infrared gas sensor (called IRGA for Infra-Red Gas Analysis sensors) [32] |
| Carbon isotopes | Uses carbon isotopes such as 11C, 12C, and 14C to produce incorporated CO2 with radioactivity. This methodology is applied to analyze samples in isolated and illuminated chambers to produce a maximum fixation of radioactive CO2 during photosynthesis [33,34]. The main disadvantage is that it is destructive as it fixes a radioactive compound onto the sample; and its precision depends on lighting conditions. |
| Acoustic waves | Based on the principle of sound wave distortion in the medium in which waves propagate. The technique involves placing an acoustic transmitter on the seabed area where you want to monitor photosynthetic activity. The disadvantage is that it dependent on water conditions and sensitive to environmental disturbances [35]. |
| Fluorescence | Way in which a certain amount of light energy absorbed by chlorophylls is dissipated. The fluorescence emission can be analyzed and quantified, which provides information on the electron transport rate, the quantum yield and the existence of photoinhibition of photosynthesis. Indeed, fluorescence is used in various ways, and it has different applications. The interested reader is referred to reference [5,8,24]. |
| Mathematical modeling | Equation or a set of equations that represent a system’s behavior. There is a correspondence between the model variables and the observable quantities [36]. |
| Equation | Author | Mechanistic Mathematical Models |
|---|---|---|
| (1) | Farquhar [79] | |
| (2) | Marshall [82] | |
| (3) | Kano [83] | |
| (4) | C.Pahl-Wostl [84] | |
| (5) | Lieth [85] | |
| (6) | Jones [86] | |
| (7) | Hahn [87] | |
| (8) | Pachepsky [88] | |
| (9) | Chen [89] | |
| (10) | Nikolov [90] | |
| (11) | Kull [91] | |
| (12) | Chen [92] | where Aj: where Ac: for both: |
| (13) | Han [93] | |
| (14) | Rubio [94] | |
| (15) | Ye [95] | |
| (16) | Ye [96] | |
| (17) | Polerecky [39] | |
| (18) | Bernacchi [97] | |
| (19) | Yin [98] | C3 plants for the limited Rubisco part: X1: Vcmax, X2: KmC (1 + O/KmO): for the limited transport part C4 plants : 0.5/Sc/o, X2: KmC/KmO, X3 = KmC |
| (20) | Torres [99] | |
| (21) | Johnson [100] | |
| (22) | Egea [101] | |
| (23) | Lombardozzi [102] | |
| (24) | García-Camacho [103] | |
| (25) | Janka [104] | |
| (26) | Liu [105] | |
| Empirical mathematical models | ||
| (27) | Hozumi & Kirita [106] | |
| (28) | Wetzel [107] | |
| (29) | Pimienta de la Torre [108] | |
| (30) | Romdhonah [109] | (Linear model (G-model)) (Linear interaction model (I-model)) (Linear squared model (S-model)) (Linear interaction-squared model (IS-model)) |
| (31) | Shimada [110] | |
| Gray box mathematical models | ||
| (32) | Kolber [111] | |
| (33) | Jansen [112] | Functions f1 and f2 differ by crop, and by crop history |
| (34) | Sau [113] | PCARB = EUR · IPAR/PLTPOP · FC02 IPAR = PAR · [1 − exp(-EXT · IAF)] |
| (35) | Costache [114] | |
| (36) | Ippoliti [115] | |
| (37) | Perin [116] | |
| (38) | Liu [117] | |
| (39) | Sánchez [118] | |
| Equation [Ref] | Input Variables | Estimated Variable | No. of Samples | Validation Method | R2 |
|---|---|---|---|---|---|
| Mechanistic mathematical models | |||||
| (1) [79] | Ambient temperature [°C] CO2 concentration in the leaf [mbar] Light intensity Oxygen concentration in the leaf [mbar] | CO2 assimilation | - | Gas exchange | - |
| (2) [82] | CO2 in the atmosphere at [bar] Irradiance | Net photosynthetic rate | - | Rectangular hyperbolic model | - |
| (3) [83] | Ambient temperature [°C] Concentration in the leaf Photosynthetically active irradiation | Photosynthetic potential rate | 60 | Simulation | - |
| (4) [84] | Light intensity Time [min] | Photosynthesis rate [Relative Units (bel)] | - | Gas exchange system LI-COR 6000 (First generation) | - |
| (5) [85] | Photosynthetically active radiation PAR Leaf’s age [days] Leaf’s temperature [°C] | Maximum net photosynthesis rate | 15 | Gas exchange system LI-COR 6000(First generation) | 0.95 |
| (6) [86] | Ambient temperature [°C] Foliar area index CO2 in the atmosphere [Ppm] Light (photosynthetic photon flux density, PPFD) [PAR] | Photosynthetic rate | 264 | Previous research and simulation | - |
| (7) [87] | CO2 concentration in the leaf [No information] Ribulose bisphosphate [No information] Triose phosphate [No information] | Net photosynthetic rate | - | Simulation | - |
| (8) [88] | Leaf’s Breathing Light CO2 in the atmostphere Ambient temperature constant [°C] | Photosynthetic rate | - | Analysis of the models by F-test | - |
| (9) [89] | Ambient temperature [°C] Oxygen in the atmosphere [mbar] CO2 in the leaf [Ppm] Irradience Rubisco | Net photosynthesis | - | Photosynthesis measurement model experiment by Collatz | 0.95 |
| (10) [90] | CO2 in the leaf Oxygen in the leaf [Pa] Leaf’s temperature [°C] | Photosynthetic rate | - | Gas exchange system Li-COR 6200 (Second generation) | 0.64 |
| (11) [91] | CO2 in the leaf [Ppm] Light Nitrogen Chlorophyll | Net photosynthesis | - | Gas exchange system Li-COR 6200 | - |
| (12) [92] | CO2 in the atmosphere Oxygen in the leaf [Pa] Ambient temperature [°C] Stomatal conductance Light | Net foliar photosynthesis rate | - | FvCB model | 0.72 |
| (13) [93] | Chlorophyll Oxygen in the leaf Irradiance | Photosynthetic rate | - | Empirical function based on the Poisson distribution | - |
| (14) [94] | Irradiance Energy Composition Concentration of power supplies activated | Photosynthesis | - | J. Neal Phillips, Jr. and Jack Myers Growth Rate Model | - |
| (15) [95] | PAR irradiance Ambient temperature [°C] | Net photosynthesis | - | Non-rectangular and Rectangular hyperbolic model Binomial regression method | - |
| (16) [96] | Nonphotorespiratory mitochondrial CO2 release in light Light intensity | Net photosynthetic rate | - | Simulation | 0.99 |
| (17) [39] | Light intensity O2 on the leaf [mM] | Gross rate of photosynthesis | 2 of microbial 1 of corals | Comparison between the same simulated and implemented model | 0.95 |
| (18) [97] | CO2 in the atmosphere Oxygen in the atmosphere Ambient temperature [°C] | Photosynthesis | - | FvCB model | - |
| (19) [98] | CO2 in the leaf Oxygen in the leaf [µ bar] Stomatal conductance | Net photosynthesis rate | - | FvCB model | - |
| (20) [99] | CO2 in the atmosphere Leaf’s temperature [°C] Light Foliar Area Index CO2 conductance in the leaf Light flux density in foilage Plant population density [m2] | Rate of gross photosynthesis | - | Simulation | - |
| (21) [100] | Irradiance Ambient temperature [°C] CO2 in the atmosphere Nitrogen | Photosynthetic rate | - | Photosynthesis measurement model by Johnson | - |
| (22) [101] | CO2 in the leaf CO2 in the atmosphere Oxigen in the leaf [Pa] Leaf’s temperature [°C] Light | Photosynthetic rate | - | FvCB model | - |
| (23) [102] | CO2 concentration in the atmosphere CO2 concentration on the leaf surface Internal CO2 concentration in the leaf Stomatal conductance Light Ambient temperature [°C] Oxygen in the leaf | Carbon assimilation | 5 | Gas exchange system LI-COR 6400 (Third generation) | - |
| (24) [103] | Oxygen in the leaf Disappearance of PSU activated Total concentration of PSUs [PSU Cell−1] Time [s] | Photosynthesis rate | - | Mathematical modeling previously developed by García-Camacho | - |
| (25) [104] | Leaf’s temperature [°C] Light CO2 concentration in the leaf | Photosynthesis rate | 80 | Biochemical model of photosynthesis | - |
| (26) [105] | Radiation [PAR] CO2 in the leaf Leaf’s temperature [°C] Pressure steam [KPa] Leaf’s mass per area [gm−2] | Photosynthetic net rate | 5 | Gas exchange system LI-COR 6400 (Third generation) andJackknife technique | 0.85 |
| Empirical mathematical models | |||||
| (27) [106] | Light intensity [Klux] Height [m] | Total gross photosynthesis | 5 species Plot (Q 15, 20 m × 20 m) | Monsi Saeki’s mathematical model | 0.60 |
| (28) [107] | Ambient temperature [°C] Irradiation | Photosynthesis | - | Euler’s simple digital integration method, rectangle hyperbolic method | - |
| (29) [108] | Stem diameter [cm] °H: Heigh [m] | Biomass [Kg] | 663 | Comparison between the same models | 0.99 |
| (30) [109] | Photosynthetically active radiation Air temperature [°C] CO2 concentration Vapor pressure déficit [KPa] | Net photosynthetic rate | - | Test data | 0.82 (G-model) 0.91 (I-model) 0.90 (S-model) 0.92 (IS-model) |
| (31) [110] | Photosynthetic photon flux density Air temperature [°C] Soil temperature [°C] Vapor pressure deficit [Kpa] Soil water content [unitless] Age [Year] | Photosynthetic rate | - | Simulation | 0.86 |
| Gray box mathematical models | |||||
| (32) [111] | Irradiance Fluorescence | Rate of gross photosysthesis | 61 | “Pump and probe technique” Forometry | 0.86 |
| (33) [112] | Ambient temperature [°C] Radiation Time [ hours = i, day = d] | Photosynthesis gross | - | Integration model according to Euler | 0.96 |
| (34) [113] | Radiation CO2 in the atmosphere Plants surface [plant m−2] Ambient temperature [°C] | Potential biomass accumulation per plant [g planta−1 d−1] | - | Simulation model | No information |
| (35) [114] | Irradiance Temperature [°C] pH [unitless] Dissolved oxygen concentration | Photosynthesis rate | - | Data obtained by simultaneous modifications of variables | 0.74 |
| (36) [115] | Irradiance Temperature [°C] pH [unitless] Dissolved oxygen concentration | Net photosynthesis rate | - | Experimental data assembly from an industrial-scale outdoor tubular photobioreactor (T-PBRs) | 0.89 |
| (37) [116] | Light Fluorescence [chl] | Photosynthesis | - | Hyperbolic model | 0.85 |
| (38) [117] | Leaf mass per area Photosynthetically active radiation Air temperature [°C] Vapor pressure defcit [KPa] | Net photosynthetic rate | 5 trees | Gas exchange systemLI-6400XT | 0.87 |
| (39) [118] | Irradiance Temperature [°C] pH [unitless] Dissolved oxygen | Net photosynthesis rate | - | Data obtained by simultaneous modifications of variables | 0.60 |
| (33) [112] | Ambient temperature [°C] Radiation Time [ hours = i, day = d] | Photosynthesis gross | - | Integration model according to Euler | 0.96 |
| (34) [113] | Radiation CO2 in the atmosphere Plants surface [plant m−2] Ambient temperature [°C] | Potential biomass accumulation per plant [g planta−1 d−1] | - | Simulation model | No information |
| (35) [114] | Irradiance Temperature [°C] pH [unitless] Dissolved oxygen concentration | Photosynthesis rate | - | Data obtained by simultaneous modifications of variables | 0.74 |
| (36) [115] | Irradiance Temperature [°C] pH [unitless] Dissolved oxygen concentration | Net photosynthesis rate | - | Experimental data assembly from an industrial-scale outdoor tubular photobioreactor (T-PBRs) | 0.89 |
| (37) [116] | Light Fluorescence [chl] | Photosynthesis | - | Hyperbolic model | 0.85 |
| (38) [117] | Leaf mass per area Photosynthetically active radiation Air temperature [°C] Vapor pressure defcit [KPa] | Net photosynthetic rate | 5 trees | Gas exchange system LI-6400XT | 0.87 |
| (39) [118] | Irradiance Temperature [°C] pH [unitless] Dissolved oxygen | Net photosynthesis rate | - | Data obtained by simultaneous modifications of variables | 0.60 |
| Mathematical Models | ||||
|---|---|---|---|---|
| Number of Input Variables | White Box Models | Black Box Models | Gray Box Models | Percentage Equivalent to the Number of Input Variables Used by Models |
| 7 | 8% | - | - | 5% |
| 5 | 15% | 20% | - | 13% |
| 4 | 23% | 20% | 50% | 28% |
| 3 | 31% | - | 13% | 23% |
| 2 | 23% | 60% | 38% | 31% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
García-Rodríguez, L.d.C.; Prado-Olivarez, J.; Guzmán-Cruz, R.; Rodríguez-Licea, M.A.; Barranco-Gutiérrez, A.I.; Perez-Pinal, F.J.; Espinosa-Calderon, A. Mathematical Modeling to Estimate Photosynthesis: A State of the Art. Appl. Sci. 2022, 12, 5537. https://doi.org/10.3390/app12115537
García-Rodríguez LdC, Prado-Olivarez J, Guzmán-Cruz R, Rodríguez-Licea MA, Barranco-Gutiérrez AI, Perez-Pinal FJ, Espinosa-Calderon A. Mathematical Modeling to Estimate Photosynthesis: A State of the Art. Applied Sciences. 2022; 12(11):5537. https://doi.org/10.3390/app12115537
Chicago/Turabian StyleGarcía-Rodríguez, Luz del Carmen, Juan Prado-Olivarez, Rosario Guzmán-Cruz, Martín Antonio Rodríguez-Licea, Alejandro Israel Barranco-Gutiérrez, Francisco Javier Perez-Pinal, and Alejandro Espinosa-Calderon. 2022. "Mathematical Modeling to Estimate Photosynthesis: A State of the Art" Applied Sciences 12, no. 11: 5537. https://doi.org/10.3390/app12115537