
Inactivation of SARS-CoV-2 on Surfaces by Cold-Plasma-Generated Reactive Species
 , , and
, , and
Abstract
:1. Introduction
2. Experimental Methods
2.1. Plasma Apparatus Design
2.2. Virus Construction and Infectivity Experimental Methods
2.3. SARS-CoV-2 Pseudotyped Lentivirus Production
2.4. SARS-CoV-2 Pseudotyped Lentivirus Infectivity Assay
2.5. Preparation of SARS-CoV-2 Pseudotyped Lentivirus for CAP Treatment
3. Results
3.1. Spectroscopy Characterization of Cold Plasma-Generated Reactive Species
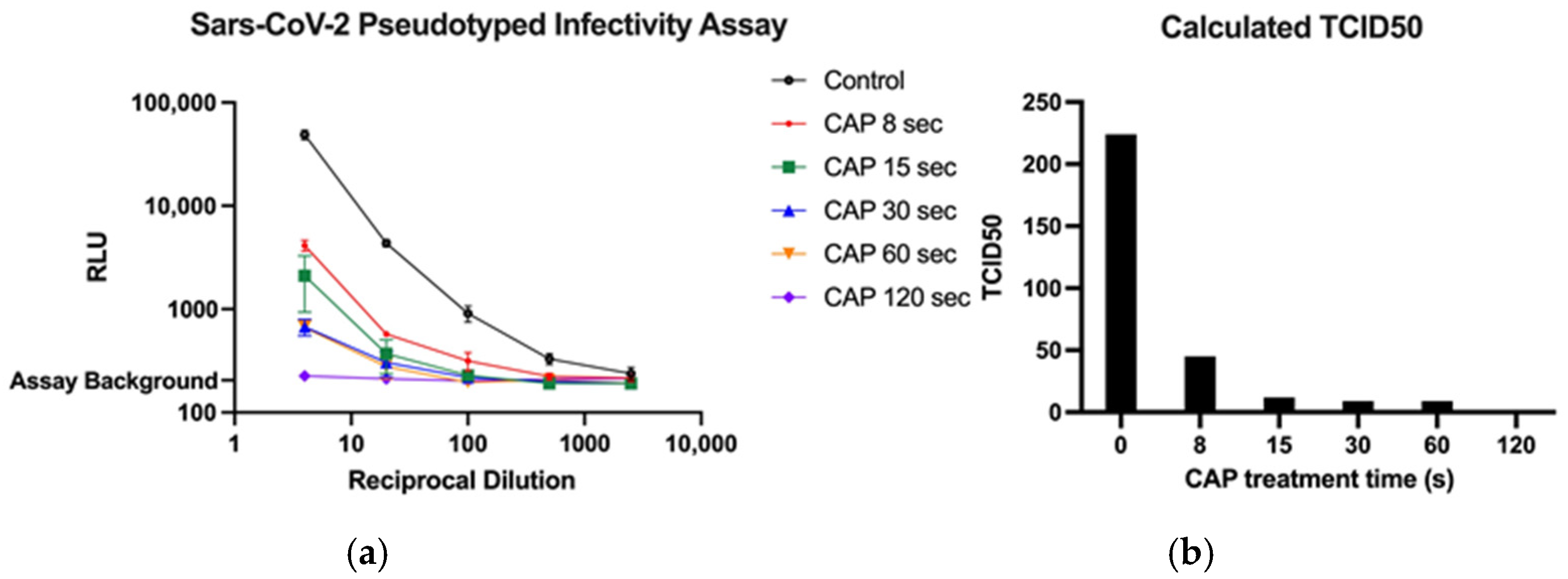
3.2. SARS-CoV-2 Pseudotyped Viral Infectivity Assays
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Chu, P.K.; Lu, X. (Eds.) Low Temperature Plasma Technology: Methods and Applications; CRC Press: Boca Raton, FL, USA, 2013. [Google Scholar]
- Piel, A.; Brown, M. Plasma physics: An introduction to laboratory, space, and fusion plasmas. Phys. Today 2011, 64, 55. [Google Scholar]
- Chen, F.F. Introduction to Plasma Physics and Controlled Fusion; Plenum Press: New York, NY, USA, 1984; Volume 1, pp. 19–51. [Google Scholar]
- Shi, D.; Lian, J.; He, P.; Wang, L.M.; Xiao, F.; Yang, L.; Schulz, M.J.; Mast, D.B. Plasma coating of carbon nanofibers for enhanced dispersion and interfacial bonding in polymer composites. Appl. Phys. Lett. 2003, 83, 5301–5303. [Google Scholar] [CrossRef] [Green Version]
- Zhao, J.; Schaefer, D.W.; Shi, D.; Lian, J.; Brown, J.; Beaucage, G.; Wang, L.; Ewing, R.C. How does surface modification aid in the dispersion of carbon nanofibers? J. Phys. Chem. B 2005, 109, 23351–23357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, D.; He, P.; Wang, S.X.; Van Ooij, W.J.; Wang, L.M.; Zhao, J.; Yu, Z. Interfacial particle bonding via an ultrathin polymer film on Al2O3 nanoparticles by plasma polymerization. J. Mater. Res. 2002, 17, 981–990. [Google Scholar] [CrossRef] [Green Version]
- Shi, D.; Lian, J.; He, P.; Wang, L.M.; van Ooij, W.J.; Schulz, M.; Liu, Y.; Mast, D.B. Plasma deposition of ultrathin polymer films on carbon nanotubes. Appl. Phys. Lett. 2002, 81, 5216–5218. [Google Scholar] [CrossRef] [Green Version]
- Shi, D.; Wang, S.X.; van Ooij, W.J.; Wang, L.M.; Zhao, J.; Yu, Z. Uniform deposition of ultrathin polymer films on the surfaces of Al 2 O 3 nanoparticles by a plasma treatment. Appl. Phys. Lett. 2001, 78, 1243–1245. [Google Scholar] [CrossRef]
- Shi, D.; Lian, J.; Peng, H.; Wang, W.; Dong, Z.; Wang, L.; Ewing, R.C. Luminescent Carbon Nanotubes. Adv. Mater. 2006, 18, 189–193. [Google Scholar] [CrossRef]
- Guo, Y.; Cho, H.; Shi, D.; Lian, J.; Song, Y.; Abot, J.; Poudel, B.; Ren, Z.; Wang, L.; Ewing, R.C. Effects of plasma surface modification on interfacial behaviors and mechanical properties of carbon nanotube-Al2O3 nanocomposites. Appl. Phys. Lett. 2007, 91, 261903. [Google Scholar] [CrossRef]
- Cho, H.; Shi, D.; Guo, Y.; Lian, J.; Ren, Z.; Poudel, B.; Song, Y.; Abot, J.L.; Singh, D.; Routbort, J.; et al. Enhanced thermal stability of carbon nanotubes by plasma surface modification in Al2O3 composites. J. Appl. Phys. 2008, 104, 074302. [Google Scholar] [CrossRef]
- Chaudhary, K.; Imam, A.M.; Rizvi, S.Z.H.; Ali, J. Plasma Kinetic Theory. In Kinetic Theory; InTech: Rijeka, Croatia, 2018; p. 107. ISBN 978-953-51-3801-3. [Google Scholar]
- Sakudo, A.; Yagyu, Y.; Onodera, T. Disinfection and sterilization using plasma technology: Fundamentals and future perspectives for biological applications. Int. J. Mol. Sci. 2019, 20, 5216. [Google Scholar] [CrossRef] [Green Version]
- Annunziata, M.; Canullo, L.; Donnarumma, G.; Caputo, P.; Nastri, L.; Guida, L. Bacterial inactivation/sterilization by argon plasma treatment on contaminated titanium implant surfaces: In vitro study. Med. Oral Patol. Oral Cirugía Bucal 2016, 21, e118. [Google Scholar] [CrossRef] [PubMed]
- Misra, N.N.; Yadav, B.; Roopesh, M.S.; Jo, C. Cold plasma for effective fungal and mycotoxin control in foods: Mechanisms, inactivation effects, and applications. Compr. Rev. Food Sci. Food Saf. 2019, 18, 106–120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gururani, P.; Bhatnagar, P.; Bisht, B.; Kumar, V.; Joshi, N.C.; Tomar, M.S.; Pathak, B. Cold plasma technology: Advanced and sustainable approach for wastewater treatment. Environ. Sci. Pollut. Res. 2021, 28, 65062–65082. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Garcia, G., Jr.; Arumugaswami, V.; Wirz, R.E. Cold atmospheric plasma for SARS-CoV-2 inactivation. Phys. Fluids 2020, 32, 111702. [Google Scholar] [CrossRef]
- Yadav, D.K.; Adhikari, M.; Kumar, S.; Ghimire, B.; Han, I.; Kim, M.H.; Choi, E.H. Cold atmospheric plasma generated reactive species aided inhibitory effects on human melanoma cells: An in vitro and in silico study. Sci. Rep. 2020, 10, 3396. [Google Scholar] [CrossRef] [Green Version]
- Fukuda, N.; Tsuchiya, A.; Toita, R.; Tsuru, K.; Mori, Y.; Ishikawa, K. Surface plasma treatment and phosphorylation enhance the biological performance of poly (ether ether ketone). Colloids Surf. B Biointerfaces 2019, 173, 36–42. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Bai, F.; Jonas, S.J.; Wirz, R.E. Cold atmospheric plasma for addressing the COVID-19 pandemic. Plasma Process. Polym. 2022, 19, 2200012. [Google Scholar] [CrossRef]
- Zhang, X.; Li, X.; Li, Y.; Ma, Y.; Su, X.; Xue, Y.; Liu, D. Inactivation of SARS-CoV-2 by Cold At-mospheric. Plasma. Matter. 2021, 4, 1752–1765. [Google Scholar]
- Zhang, H.; Chen, M.; Huang, L.; Guo, L.; Xu, S.; Zhang, J.; Xi, W.; Wang, Z.; Liu, D.; Kong, M.G.; et al. Using cold atmospheric plasma treated-air for COVID-19 disinfection in cold-chain environment. J. Phys. D Appl. Phys. 2021, 54, 40LT01. [Google Scholar] [CrossRef]
- Cortázar, O.D.; Megía-Macías, A.; Moreno, S.; Brun, A.; Gómez-Casado, E. Vulnerability of SARS-CoV-2 and PR8 H1N1 virus to cold atmospheric plasma activated media. Sci. Rep. 2022, 12, 263. [Google Scholar] [CrossRef]
- Assadi, I.; Guesmi, A.; Baaloudj, O.; Zeghioud, H.; Elfalleh, W.; Benhammadi, N.; Khezami, L.; Assadi, A.A. Review on inactivation of airborne viruses using non-thermal plasma technologies: From MS2 to coronavirus. Environ. Sci. Pollut. Res. 2022, 29, 4880–4892. [Google Scholar] [CrossRef] [PubMed]
- Yan, D.; Malyavko, A.; Wang, Q.; Ostrikov, K.; Sherman, J.H.; Keidar, M. Multi-modal biological destruction by cold atmospheric plasma: Capability and mechanism. Biomedicines 2021, 9, 1259. [Google Scholar] [CrossRef] [PubMed]
- Johnson, M.C.; Lyddon, T.D.; Suarez, R.; Salcedo, B.; Lepique, M.; Graham, M.; Ricana, C.; Robinson, C.; Ritter, D.G. Optimized pseudotyping conditions for the SARS-CoV-2 spike glycoprotein. J. Virol. 2020, 94, e01062-20. [Google Scholar] [CrossRef] [PubMed]
- Sarzotti-Kelsoe, M.; Bailer, R.T.; Turk, E.; Lin, C.L.; Bilska, M.; Greene, K.M.; Gao, H.; Todd, C.A.; Ozaki, D.A.; Seaman, M.S.; et al. Optimization and validation of the TZM-bl assay for standardized assessments of neutralizing antibodies against HIV. J. Immunol. Methods 2014, 409, 131–146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hammonds, J.; Chen, X.; Fouts, T.; DeVico, A.; Montefiori, D.; Spearman, P. Induction of neutralizing antibodies against human immunodeficiency virus type 1 primary isolates by Gag-Env pseudovirion immunization. J. Virol. 2005, 79, 14804–14814. [Google Scholar] [CrossRef] [Green Version]
- Xing, L.; Wang, S.; Hu, Q.; Li, J.; Zeng, Y. Comparison of three quantification methods for the TZM-bl pseudovirus assay for screening of anti-HIV-1 agents. J. Virol. Methods 2016, 233, 56–61. [Google Scholar] [CrossRef]
- Loizou, C.; Kniazeva, V.; Apostolou, T.; Kornev, A.; Kostevitch, S.; Roslyakov, E.; Constantinou, C.; Hadjihannas, L. Effect of Cold Atmospheric Plasma on SARS-CoV-2 Inactivation: A Pilot Study in the Hospital Environment. COVID 2022, 2, 1396–1404. [Google Scholar] [CrossRef]
- Bisag, P.A.; Isabelli, G.; Laghi, R.; Laurita, G.; Dirani, F.; Taddei, C.; Bucci, F.; Capelli, M.; Gherardi, A.; Paglianti, V.; et al. Cold atmospheric plasma decontamination of SARS-CoV-2 bioaerosols. Plasma Process. Polym. 2022, 19, e2100133. [Google Scholar] [CrossRef]
- Mohamed, H.; Nayak, G.; Rendine, N.; Wigdahl, B.; FC, K.; Bruggeman, P.J.; Miller, V. Non-Thermal Plasma as a Novel Strategy for Treating or Preventing Viral Infection and Associated Disease. Front. Phys. 2021, 9, 683118. [Google Scholar] [CrossRef]
- Kaushik, N.; Mitra, S.; Baek, E.J.; Nguyen, L.N.; Bhartiya, P.; Kim, J.H.; Choi, E.H.; Kaushik, N.K. The inactivation and destruction of viruses by reactive oxygen species generated through physical and cold atmospheric plasma techniques: Current status and perspectives. J. Adv. Res. 2022, 38, 143–155. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Zhou, R.; Zhou, R.; Li, W.; Weerasinghe, J.; Chen, S.; Rehm, B.H.A.; Zhao, L.; Frentiu, F.D.; Zhang, Z.; et al. Cold atmospheric plasma for preventing infection of viruses that use ACE2 for entry. Theranostics 2022, 12, 2811. [Google Scholar] [CrossRef] [PubMed]
- Yan, D.; Sherman, J.H.; Keidar, M.; Laroussi, M. Plasma for virus inactivation: Historical perspectives and modern developments. Plasma Sources Sci. Technol. 2021, 30, 033001. [Google Scholar]
- Nathan, C.; Cunningham-Bussel, A. Beyond oxidative stress: An immunologist’s guide to reactive oxygen species. Nat. Rev. Immunol. 2013, 13, 349–361. [Google Scholar] [CrossRef] [Green Version]
- Rahman, I.; MacNee, W. Oxidative stress and regulation of glutathione in lung inflammation. Eur. Respir. J. 2000, 16, 534–554. [Google Scholar] [CrossRef]
- Circu, M.L.; Aw, T.Y. Reactive oxygen species, cellular redox systems, and apoptosis. Free. Radic. Biol. Med. 2010, 48, 749–762. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
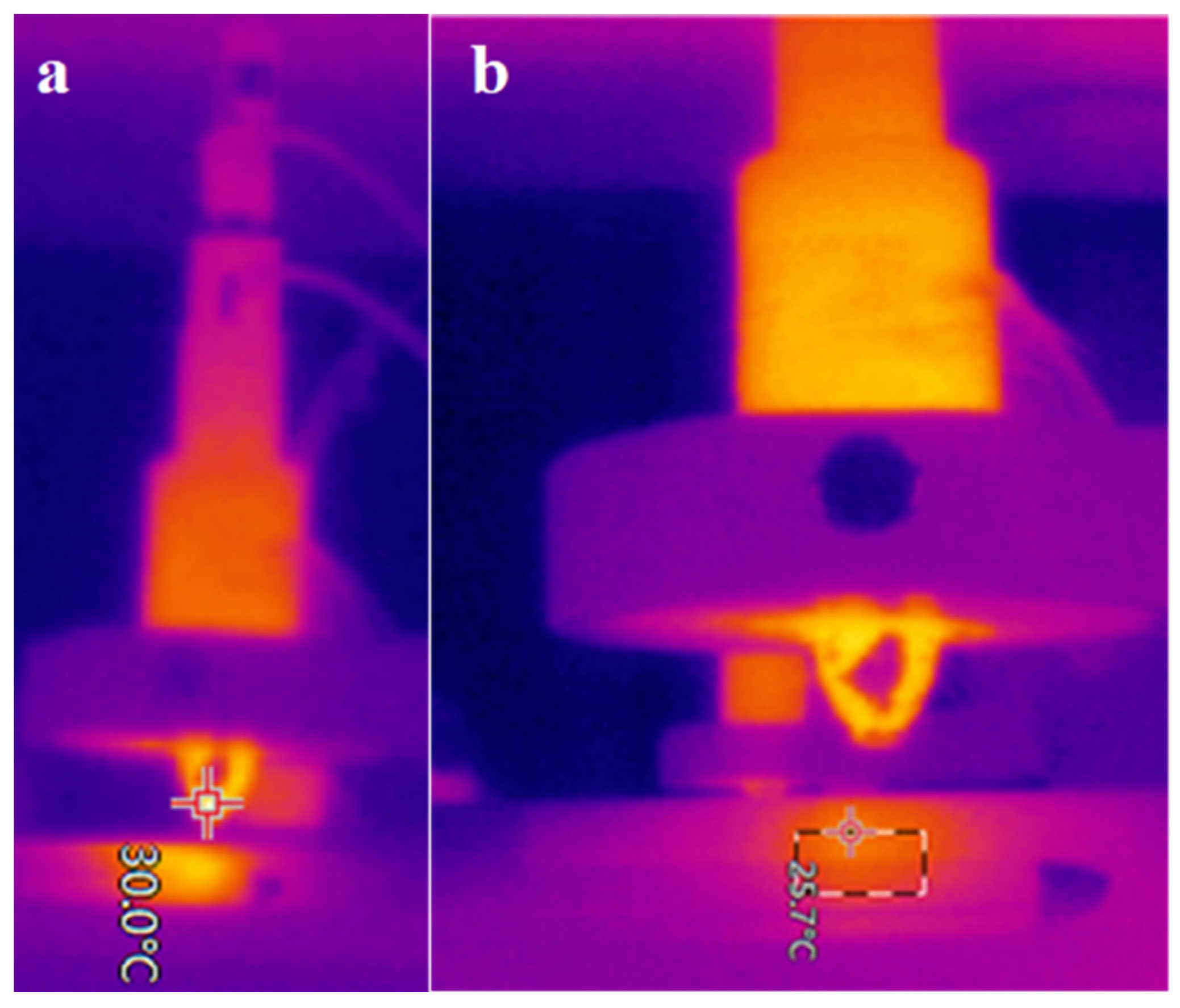{kind=link}
{kind=link}
{kind=link}
| Wave-Length (nm) | Intensity [(µW/cm2)/nm] | Element * | Line Identified (nm) | I (Rel.) | Energy (eV) Lower–Upper | Transition Lower–Upper | Quantum Number Lower–Upper |
|---|---|---|---|---|---|---|---|
| 580.241 | 1121.53 | N I | 580.2836 | 4 | 11.84–13.98 | 3p 4P–5d 4F | 2½–2½ |
| 580.241 | 1121.53 | N I | 580.3043 | 3 | 12.13–14.26 | 3p 2P–8s 2P | 1½–1½ |
| 631.294 | 820.914 | N II | 631.3809 | 10 | 25.65–27.62 | 4p ¹S–6s 3P° | 0–1 |
| 670.831 | 1775.77 | N I | 670.8759 | 13 | 11.84–13.69 | 3p 4P°–4d 4D | 1½–1½ |
| 670.831 | 1775.77 | O III | 670.736 | 2 | 47.03–48.87 | 4d 3D°–3s 3D | 2–3 |
| 670.831 | 1775.77 | O IV | 670.866 | 2 | 71.54–73.39 | 3s 4P–4d 4D° | 1½–½ |
| 670.831 | 1775.77 | O IV | 670.866 | 12 | 71.54–73.39 | 3s 4P–4d 4D° | 1½–1½ |
| 729.647 | 1512.35 | O II | 729.6294 | 0 | 28.86–30.56 | 3d 4D–4p 4P° | 3½–2½ |
| 735.934 | 895.199 | N I | 736.0126 | 1 | 12.01–13.69 | 3p 2D°–4d 2D | 2½–1½ |
| 749.633 | 1197.4 | N I | 749.7122 | 0 | 13.24–14.90 | 4p 4D–3d’ 2G | 2½–3½ |
| 829.625 | 1541.9 | N I | 829.6007 | 3 | 12.13–13.62 | 3p 2P–5s 4P | 1½–1½ |
| 829.625 | 1541.9 | N II | 829.621 | 24 | 23.57–25.07 | 3d ¹P°–4p ¹P | 1–1 |
| 838.084 | 1246.14 | O II | 838.177 | 194 | 30.47–31.95 | 4p 4D°–5s 4P | ½–½ |
| 838.084 | 1246.14 | O III | 838.036 | 1 | 45.44–46.92 | 4p 3D–3p 3P° | 1–1 |
| 858.346 | 1166.76 | O III | 858.361 | 84 | 54.37–55.82 | 4p’ 5D°–4d’ 5P | 4–3 |
| 898.415 | 1465.49 | N II | 898.3277 | 13 | 26.03–27.41 | 4d ¹D°–5f 2[2½] | 2–3 |
| 932.692 | 1845.99 | O II | 932.7453 | 205 | 31.63–32.96 | 4d 4P–5f 4D° | 1½–½ |
| 945.743 | 2754.02 | O II | 945.789 | 34 | 28.51–29.82 | 3p 2D°–4s 2P | 2½–1½ |
| 945.743 | 2754.02 | O II | 945.8429 | 17 | 28.51–29.82 | 3p’ 2D°–4s 2P | 2½–1½ |
| 981.981 | 1702.36 | O II | 982.055 | 63 | 30.80–32.06 | 4p 2P°–5s 2P | ½–1½ |
| 981.981 | 1702.36 | O III | 982.047 | 54 | 45.94–47.20 | 4p 3P–4d 3P° | 2–2 |
| 981.981 | 1702.36 | O IV | 981.895 | 1 | 73.96–75.22 | 4f 4D–3p 4P° | 1½–2½ |
| 1001.81 | 1674.33 | N I | 1001.783 | 34 | 11.76–12.99 | 3p 4D°–3d 2F | 2½–2½ |
| 1001.81 | 1674.33 | N II | 1001.903 | 1 | 26.21–27.45 | 4f’ 2[4½]–5g’ 2[2½]° | 4–3 |
| 1020.2 | 1266.48 | N I | 1020.112 | 1 | 13.68–14.90 | 4f 2D [3]–3d’ 2G | 3½–4½ |
| 1020.2 | 1266.48 | N I | 1020.113 | 2 | 13.68–14.90 | 4f 2D[3]–3d’ 2G | 2½–3½ |
| 1020.2 | 1266.48 | N I | 1020.122 | 0 | 13.68–14.90 | 4f 2D[3]–3d’ 2G | 3½–3½ |
| 1020.2 | 1266.48 | O II | 1020.23 | 1 | 29.59–30.80 | 4s 4P–4p 2P° | ½–½ |
| 1022.12 | 1643.94 | N I | 1022.089 | 3 | 13.20–14.41 | 4p 2S–10d 4P | ½–½ |
| 1022.12 | 1643.94 | N I | 1022.089 | 1 | 13.20–14.41 | 4p 2S–10d 4P | ½–1½ |
| 1032.25 | 884.052 | O II | 1032.284 | 1 | 30.75–31.95 | 4p 2D°–5s 4P | 1½–½ |
| 1032.25 | 884.052 | O III | 1032.177 | 454 | 44.28–45.48 | 4s 3P°–4p 3D | 2–3 |
| 1036.63 | 1342.98 | N I | 1036.569 | 6 | 11.84–13.03 | 3p 4P–3d 2D | ½–1½ |
| 1036.63 | 1342.98 | N I | 1036.574 | 2 | 13.26–14.46 | 4p 4P–12d 2D | ½–1½ |
| 1036.63 | 1342.98 | O III | 1036.687 | 134 | 44.24–45.44 | 4s 3P°–4p 3D | 1–1 |
| 1051.37 | 1579.21 | N I | 1051.341 | 300 | 11.84–13.02 | 3p 4P°–3d 4D | ½–½ |
| 1058.73 | 1288.58 | N I | 1058.655 | 4 | 13.25–14.42 | 4p 4D–12s 4P | 3½–2½ |
| 1058.73 | 1288.58 | N I | 1058.784 | 39 | 13.29–14.46 | 4p 2D–12d 2D | 1½–1½ |
| 1058.73 | 1288.58 | N III | 1058.81 | 2 | 46.72–47.89 | 4p 4D–4d 4P° | 1½–2½ |
| 1058.73 | 1288.58 | O II | 1058.83 | 10 | 31.76–32.93 | 4f 4F°–5d 2F | 4½–3½ |
| 1069.35 | 1383.91 | N I | 1069.278 | 1 | 13.24–14.40 | 4p 4D–10d 4D | 2½–1½ |
| 1069.35 | 1383.91 | N I | 1069.278 | 4 | 13.24–14.40 | 4p 4D–10d 4D | 2½–2½ |
| 1069.35 | 1383.91 | N I | 1069.278 | 2 | 13.24–14.40 | 4p 4D–10d 4D | 2½–3½ |
| 1069.35 | 1383.91 | N I | 1069.317 | 56 | 11.84–13.00 | 3p 4P–3d 2F | 2½–3½ |
| 1069.35 | 1383.91 | N I | 1069.424 | 0 | 13.24–14.40 | 4p 4D–11s 2P | ½–1½ |
| 1069.35 | 1383.91 | N I | 1069.424 | 2 | 13.24–14.40 | 4p 4D–11s 2P | ½–½ |
| 1091.32 | 1964.13 | N I | 1091.364 | 1 | 11.84–12.98 | 3p 4P–3d 2P | 1½–½ |
| 1092.95 | 2458.16 | N I | 1092.991 | 2 | 13.20–14.34 | 4p 2S–8d 4D | ½–½ |
| Wave-Length (nm) | Intensity [(µW/cm2)/nm] | Element * | Line Identified (nm) | I (Rel.) | Energy (eV) Lower-Upper | Transition Lower-Upper | Quantum Number Lower-Upper |
|---|---|---|---|---|---|---|---|
| 670.831 | 1345.65 | N I | 670.8759 | 13 | 11.84–13.69 | 3p 4P°–4d 4D | 1½–1½ |
| 670.831 | 1345.65 | O III | 670.736 | 2 | 47.03–48.87 | 4d 3D°–3s 3D | 2–3 |
| 670.831 | 1345.65 | O IV | 670.866 | 2 | 71.54–73.39 | 3s 4P–4d 4D° | 1½–½ |
| 670.831 | 1345.65 | O IV | 670.866 | 12 | 71.54–73.39 | 3s 4P–4d 4D° | 1½–1½ |
| 735.934 | 1339.33 | N I | 736.0126 | 1 | 12.01–13.69 | 3p 2D°–4d 2D | 2½–1½ |
| 749.633 | 1323.49 | N I | 749.7122 | 0 | 13.24–14.90 | 4p 4D–3d’ 2G | 2½–3½ |
| 759.892 | 1057 | O II | 759.8285 | 15 | 28.86–30.49 | 3d 4D–4p 4D° | 1½–2½ |
| 759.892 | 1057 | O II | 759.85 | 64 | 28.86–30.49 | 3d 4D–4p 4D° | 1½–2½ |
| 759.892 | 1057 | O II | 759.9198 | 24 | 28.86–30.49 | 3d 4D–4p 4D° | 2½–2½ |
| 759.892 | 1057 | O II | 759.9384 | 110 | 28.86–30.49 | 3d 4D–4p 4D° | 2½–2½ |
| 759.892 | 1057 | O II | 759.9407 | 0 | 31.32–32.95 | 3d’ 2G–5f 2G° | 4½–3½ |
| 768.715 | 888.608 | N III | 768.7057 | 3982 | 42.40–44.01 | 5d 2D–6f 2F° | 2½–3½ |
| 803.055 | 851.363 | N I | 802.9967 | 1 | 12.12–13.67 | 3p 2P–4d 2P | ½–1½ |
| 829.625 | 981.434 | N I | 829.6007 | 3 | 12.13–13.62 | 3p 2P–5s 4P | 1½–1½ |
| 829.625 | 981.434 | N II | 829.621 | 24 | 23.57–25.07 | 3d ¹P°–4p ¹P | 1–1 |
| 838.648 | 745.261 | N III | 838.639 | 85 | 36.86–38.33 | 3s 2P°–3p 2P | 1½–1½ |
| 844.563 | 1276.25 | N III | 844.5766 | 4 | 42.54–44.01 | 5g 2G–6f 2F° | 3½–3½ |
| 844.563 | 1276.25 | N III | 844.5766 | 108 | 42.54–44.01 | 5g 2G–6f 2F° | 4½–3½ |
| 844.563 | 1276.25 | O I | 844.6247 | 115 | 9.52–10.99 | 3s 3S°–3p 3P | 1–0 |
| 844.563 | 1276.25 | O I | 844.6359 | 574 | 9.52–10.99 | 3s 3S°–3p 3P | 1–2 |
| 932.692 | 1637.01 | O II | 932.7453 | 205 | 31.63–32.96 | 4d 4P–5f 4D° | 1½–½ |
| 973.976 | 747.988 | N I | 974.0385 | 0 | 10.33–11.60 | 3s 4P–3p 2S | 1½–½ |
| 981.429 | 1722.65 | N I | 981.4021 | 13 | 11.76–13.02 | 3p 4D°–3d 4D | 2½–3½ |
| 981.429 | 1722.65 | O III | 981.369 | 20 | 45.99–47.25 | 4p ¹D–4d ¹P° | 2–1 |
| 1022.94 | 1080.7 | N II | 1022.867 | 2 | 26.21–27.42 | 4f’ 2[4½]–5g 2[3½]° | 5–4 |
| 1022.94 | 1080.7 | O II | 1022.999 | 2 | 31.75–32.96 | 4f 4F°–5d 2D | 1½–2½ |
| 1032.8 | 1059.48 | O II | 1032.772 | 0 | 31.76–32.96 | 4fF 2[4]°–5g 2[5] | 4½–5½ |
| 1071.52 | 2159.25 | N I | 1071.526 | 5 | 13.27–14.43 | 4p 4P–11d 4D | 2½–3½ |
| 1071.52 | 2159.25 | N I | 1071.526 | 0 | 13.27–14.43 | 4p 4P–11d 4D | 2½–1½ |
| 1071.52 | 2159.25 | N I | 1071.526 | 1 | 13.27–14.43 | 4p 4P–11d 4D | 2½–2½ |
| 1083.19 | 776.701 | O II | 1083.244 | 399 | 32.94–34.09 | 5f 4G°–4d’ 2D | 3½–2½ |
| 1089.43 | 1450.25 | N I | 1089.399 | 1 | 13.24–14.38 | 4p 4D–10s 4P | 1½–2½ |
| 1089.43 | 1450.25 | N I | 1089.437 | 2 | 13.24–14.38 | 4p 4D–9d 2D | 2½–2½ |
| 1089.43 | 1450.25 | N I | 1089.476 | 2 | 13.24–14.38 | 4p 2D–11d 2F | 1½–2½ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Thomas, S.V.; Dienger-Stambaugh, K.; Jordan, M.; Wang, Y.; Hammonds, J.; Spearman, P.; Shi, D. Inactivation of SARS-CoV-2 on Surfaces by Cold-Plasma-Generated Reactive Species. Bioengineering 2023, 10, 280. https://doi.org/10.3390/bioengineering10030280
Thomas SV, Dienger-Stambaugh K, Jordan M, Wang Y, Hammonds J, Spearman P, Shi D. Inactivation of SARS-CoV-2 on Surfaces by Cold-Plasma-Generated Reactive Species. Bioengineering. 2023; 10(3):280. https://doi.org/10.3390/bioengineering10030280
Chicago/Turabian StyleThomas, Som V., Krista Dienger-Stambaugh, Michael Jordan, Yuxin Wang, Jason Hammonds, Paul Spearman, and Donglu Shi. 2023. "Inactivation of SARS-CoV-2 on Surfaces by Cold-Plasma-Generated Reactive Species" Bioengineering 10, no. 3: 280. https://doi.org/10.3390/bioengineering10030280