
Efficiency of a Seedling Phenotyping Strategy to Support European Wheat Breeding Focusing on Leaf Rust Resistance

 , , and
, , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Evaluating Leaf Rust Resistance of Adult Plants in Field Trials
2.3. Evaluating Leaf Rust Resistance in Greenhouse Experiments
2.4. Analyses of Data from Field Trials
2.5. Analyses of Data from Greenhouse Experiments
3. Results
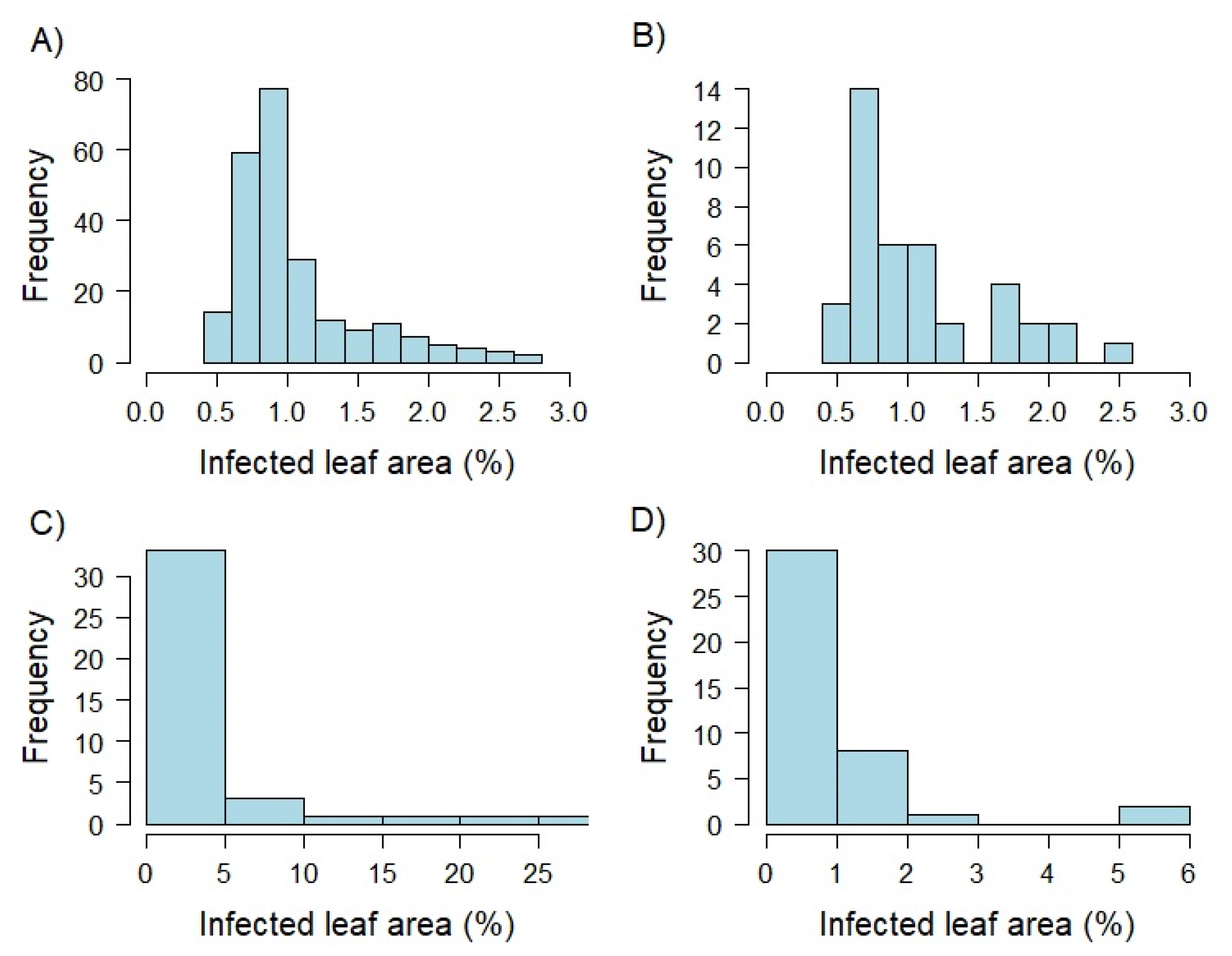
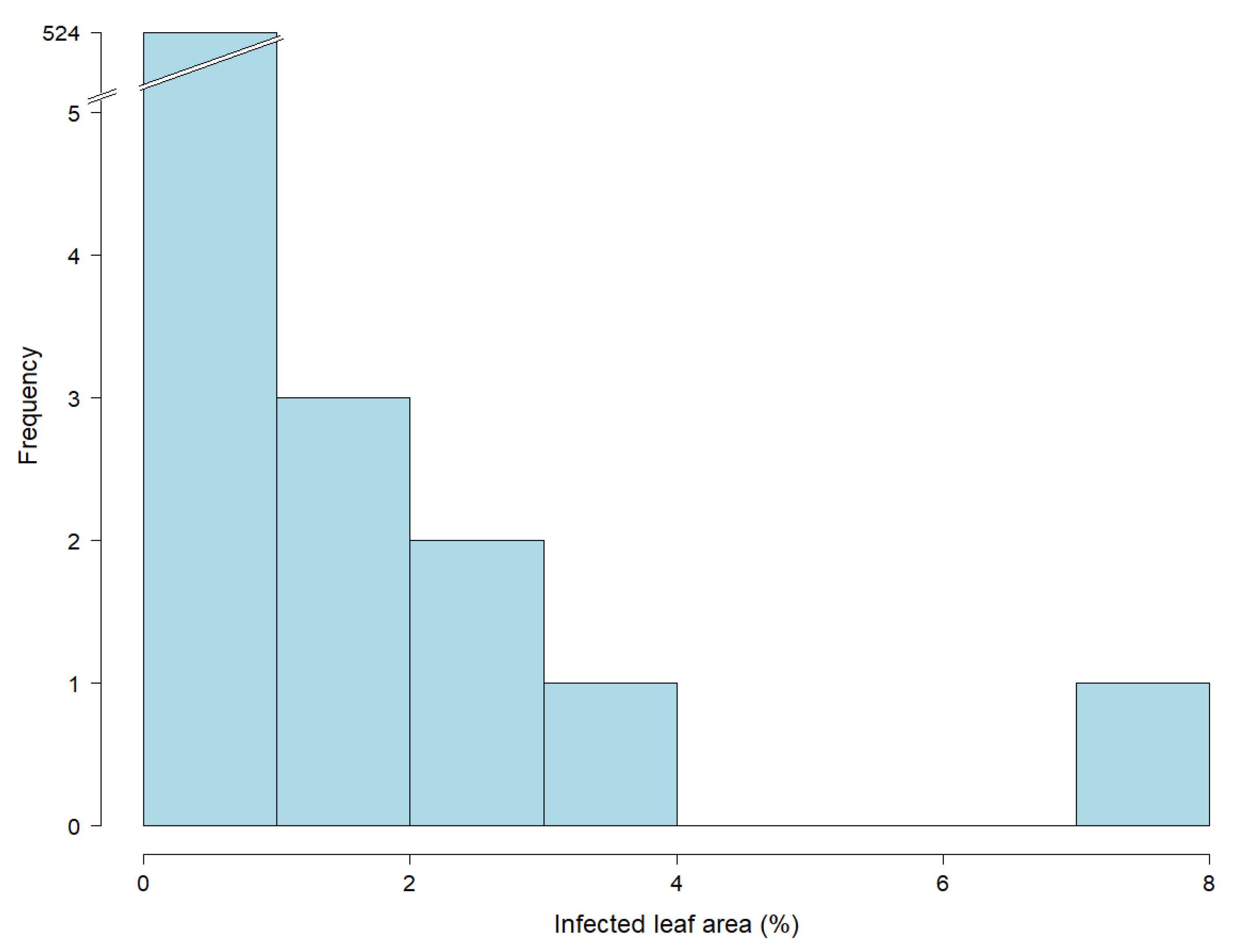
3.1. Extensive Field Trials Resulted in Precise Estimates of Adult Plant Resistance against Leaf Rust
3.2. Ensuring Stable Pathogen Pressure in Greenhouse Experiments Is Challenging
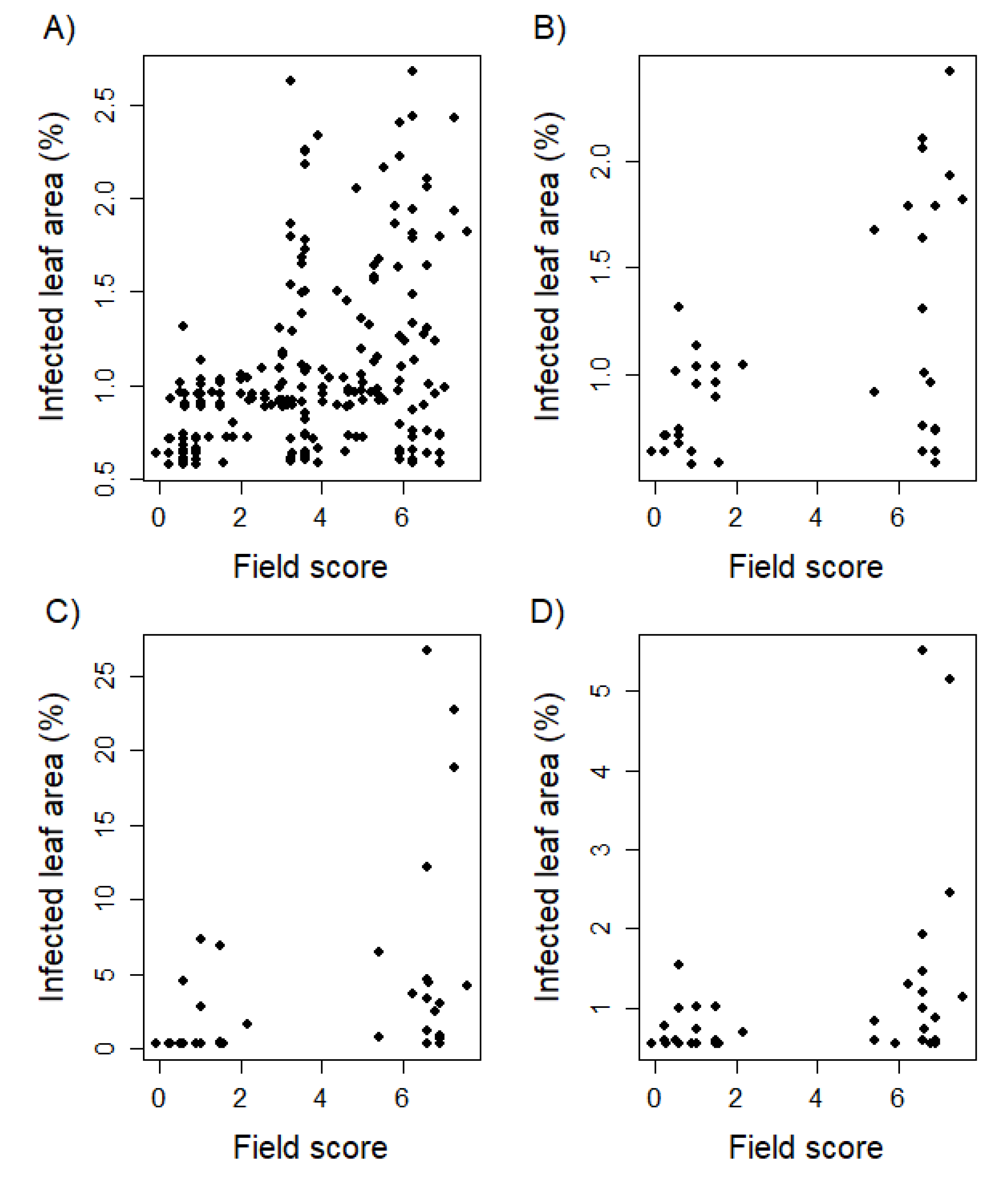
3.3. Seedling Resistance Showed a Significant Correlation to Adult Plant Resistance
4. Discussion
4.1. Divergent Conditions Increase the Quality of Resistance Phenotyping within Controlled Environments
4.2. Examining Seedling Resistance Could Support Leaf Rust Resistance Breeding within European Wheat
4.3. Automated Phenotyping of Detached Juvenile Leaves Is Beneficial for Resistance Breeding
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Huerta-Espino, J.; Singh, R.P.; German, S.; McCallum, B.D.; Park, R.F.; Chen, W.Q.; Bhardwaj, S.C.; Goyeau, H. Global status of wheat leaf rust caused by Puccinia triticina. Euphytica 2011, 179, 143–160. [Google Scholar] [CrossRef]
- Prescott, J.; Burnett, P.; Saari, E.; Ransom, J.; Bowman, J.; De Milliano, W.; Singh, R.P.; Bekele, G. Wheat Diseases and Pests: A Guide for Field Identification; CIMMYT: Mexico City, Mexico, 1986; pp. 7–9. [Google Scholar]
- Catalogue of Gene Symbols for Wheat 2017 Supplement. Available online: https://shigen.nig.ac.jp/wheat/komugi/genes/macgene/supplement2017.pdf (accessed on 6 March 2020).
- Park, R.F.; Goyeau, H.; Felsenstein, F.G.; Bartos, P.; Zeller, F.J. Regional phenotypic diversity of Puccinia triticina and wheat host resistance in western Europe, 1995. Euphytica 2001, 122, 113–127. [Google Scholar] [CrossRef]
- Pathan, A.K.; Park, R.F. Evaluation of seedling and adult plant resistance to leaf rust in European wheat cultivars. Euphytica 2006, 149, 327–342. [Google Scholar] [CrossRef]
- Serfling, A.; Krämer, I.; Lind, V.; Schliephake, E.; Ordon, F. Diagnostic value of molecular markers for Lr genes and characterization of leaf rust resistance of German winter wheat cultivars with regard to the stability of vertical resistance. Eur. J. Plant Pathol. 2011, 130, 559–575. [Google Scholar] [CrossRef]
- Bolton, M.D.; Kolmer, J.A.; Garvin, D.F. Wheat leaf rust caused by Puccinia triticina. Mol. Plant Pathol. 2008, 9, 563–575. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.P.; Huerta-Espino, J.; William, H.M. Genetics and breeding for durable resistance to leaf and stripe rusts in wheat. Turk. J. Agric. For. 2005, 29, 121–127. [Google Scholar]
- Aktar-Uz-Zaman, M.; Tuhina-Khatun, M.; Hanafi, M.M.; Sahebi, M. Genetic analysis of rust resistance genes in global wheat cultivars: An overview. Biotechnol. Biotechnol. Equip. 2017, 31, 431–445. [Google Scholar] [CrossRef] [Green Version]
- Flor, H.H. The complementary genic systems in flax and flax rust. Adv. Genet. 1956, 8, 29–54. [Google Scholar] [CrossRef]
- Flor, H.H. Current status of the gene-for-gene concept. Annu. Rev. Phytopathol. 1971, 9, 275–296. [Google Scholar] [CrossRef]
- Schwessinger, B. Fundamental wheat stripe rust research in the 21st century. New Phytol. 2017, 213, 1625–1631. [Google Scholar] [CrossRef]
- Serfling, A.; Krämer, N.; Perovic, D.; Ordon, F. Broadening the genetic base of leaf rust (Puccinia triticina f. sp. tritici) resistance in wheat (Triticum aestivum). J. Kult. 2013, 65, 262–272. [Google Scholar] [CrossRef]
- Krattinger, S.; Lagudah, E.; Spielmeyer, W.; Singh, R.; Huerta-Espino, J.; McFadden, H.; Bossolini, E.; Selter, L.; Keller, B. A putative ABC transporter confers durable resistance to multiple fungal pathogens in wheat. Science 2009, 323, 1360–1363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lillemo, M.; Asalf, B.; Singh, R.; Huerta-Espino, J.; Chen, X.; He, Z.; Bjornstad, A. The adult plant rust resistance loci Lr34/Yr18 and Lr46/Yr29 are important determinants of partial resistance to powdery mildew in bread wheat line Saar. Theor. Appl. Genet. 2008, 116, 1155–1166. [Google Scholar] [CrossRef]
- Buerstmayr, M.; Matiasch, L.; Mascher, F.; Vida, G.; Ittu, M.; Robert, O.; Holdgate, S.; Flath, K.; Neumayer, A.; Buerstmayr, H. Mapping of quantitative adult plant field resistance to leaf rust and stripe rust in two European winter wheat populations reveals co-location of three QTL conferring resistance to both rust pathogens. Theor. Appl. Genet. 2014, 127, 2011–2028. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krattinger, S.G.; Keller, B. Molecular genetics and evolution of disease resistance in cereals. New Phytol. 2016, 212, 320–332. [Google Scholar] [CrossRef] [Green Version]
- Adult Plant Resistance—Fact Sheet. Available online: https://grdc.com.au/__data/assets/pdf_file/0025/126457/adult-plant-resistance-fact-sheet.pdf.pdf (accessed on 12 March 2021).
- Zhang, H.; Wang, C.; Cheng, Y.; Chen, X.; Han, Q.; Huang, L.; Wei, G.; Kang, Z. Histological and cytological characterization of adult plant resistance to wheat stripe rust. Plant Cell Rep. 2012, 31, 2121–2137. [Google Scholar] [CrossRef]
- Hickey, L.T.; German, S.E.; Pereyra, S.A.; Diaz, J.E.; Ziems, L.A.; Fowler, R.A.; Platz, G.J.; Franckowiak, J.D.; Dieters, M.J. Speed breeding for multiple disease resistance in barley. Euphytica 2017, 213, 64. [Google Scholar] [CrossRef]
- Bernardo, R. Molecular markers and selection for complex traits in plants: Learning from the last 20 years. Crop Sci. 2008, 48, 1649–1664. [Google Scholar] [CrossRef] [Green Version]
- Meuwissen, T.H.E.; Hayes, B.J.; Goddard, M.E. Prediction of total genetic value using genome-wide dense marker maps. Genetics 2001, 157, 1819–1829. [Google Scholar] [CrossRef]
- Lueck, S.; Strickert, M.; Lorbeer, M.; Melchert, F.; Backhaus, A.; Kilias, D.; Seiffert, U.; Douchkov, D. “Macrobot”: An automated segmentation-based system for powdery mildew disease quantification. Plant Phenomics 2020, 5839856. [Google Scholar] [CrossRef]
- Riaz, A.; Periyannan, S.; Aitken, E.; Hickey, L. A rapid phenotypic method for adult plant resistance to leaf rust in wheat. Plant Methods 2016, 12, 17. [Google Scholar] [CrossRef] [Green Version]
- Irwin, J.A.; Musial, J.M.; Mackie, J.M.; Basford, K.E. Utility of cotyledon and detached leaf assays for assessing root reactions of lucerne to Phytophthora root rot caused by Phytoththora medicaginis. Australas. Plant Pathol. 2003, 32, 263–268. [Google Scholar] [CrossRef]
- Bundessortenamt. Richtlinien für die Durchführung von landwirtschaftlichen Wertprüfungen und Sortenversuchen; Landbuch: Hannover, Germany, 2000; pp. 73–74. [Google Scholar]
- Zetzsche, H.; Serfling, A.; Ordon, F. Breeding progress in seedling resistance against various races of stripe and leaf rust in European bread wheat. Crop. Breed. Genet. Genom. 2019, 1, e190021. [Google Scholar] [CrossRef] [Green Version]
- Lueck, S.; Beukert, U.; Douchkov, D. BluVision Macro—A software for automated powdery mildew and rust disease quantification on detached leaves. J. Open Source Softw. 2020, 5, 2259. [Google Scholar] [CrossRef]
- He, S.; Schulthess, A.W.; Mirdita, V.; Zhao, Y.; Korzun, V.; Bothe, R.; Ebmeyer, E.; Reif, J.C.; Jiang, Y. Genomic selection in a commercial winter wheat population. Theor. Appl. Genet. 2016, 129, 641–651. [Google Scholar] [CrossRef] [PubMed]
- Utz, H.F.; Melchinger, A.E.; Schön, C.C. Bias and sampling error of the estimated proportion of genotypic variance explained by quantitative trait loci determined from experimental data in maize using cross validation and validation with independent samples. Genetics 2000, 154, 1839–1849. [Google Scholar] [CrossRef] [PubMed]
- Hinterberger, V.; Douchkov, D.; Lueck, S.; Kale, S.; Mascher, M.; Stein, N.; Reif, J.C.; Schulthess, A.W. Mining new potential sources of resistance against powdery mildew Blumeria graminis f. sp. Tritici in genetic resources of winter wheat Triticum aestivum L. 2021; under review. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019; Available online: http://www.R-project.org/ (accessed on 22 April 2020).
- Gilmour, A.R.; Gogel, B.; Cullis, B.; Thompson, R. ASReml User Guide Release 3.0; VSN International Ltd.: Hemel Hempstead, UK, 2009. [Google Scholar]
- Gao, L.; Turner, M.K.; Chao, S.; Kolmer, J.; Anderson, J.A. Genome wide association study of seedling and adult plant leaf rust resistance in elite spring wheat breeding lines. PLoS ONE 2016, 11, e0148671. [Google Scholar] [CrossRef] [Green Version]
- Rollar, S.; Serfling, A.; Geyer, M.; Hartl, L.; Mohler, V.; Ordon, F. QTL mapping of adult plant and seedling resistance to leaf rust (Puccinia triticina Eriks.) in a multiparent advanced generation intercross (MAGIC) wheat population. Theor. Appl. Genet. 2021, 134, 37–51. [Google Scholar] [CrossRef]
- Hovmøller, M.S. Sources of seedling and adult plant resistance to Puccinia striiformis f.sp. tritici in European wheats. Plant Breed. 2007, 126, 225–233. [Google Scholar] [CrossRef]
- Aregbesola, E.; Ortega-Beltran, A.; Falade, T.; Jonathan, G.; Hearne, S.; Bandyopadhyay, R. A detached leaf assay to rapidly screen for resistance of maize to Bipolaris maydis, the causal agent of southern corn leaf blight. Eur. J. Plant Pathol. 2020, 156, 133–145. [Google Scholar] [CrossRef] [Green Version]
- Boydom, A.; Dawit, W.; Getaneh, W. Evaluation of detached leaf assay for assessing leaf rust (Puccinia triticina Eriks) resistance in wheat. J. Plant Pathol. Microbiol. 2013, 4, 176. [Google Scholar] [CrossRef] [Green Version]
- Zhao, L.; Zhang, H.; Zhang, B.; Bai, X.; Zhou, C. Physiological and molecular changes of detached wheat leaves in responding to various treatments. J. Integr. Plant Biol. 2012, 54, 567–576. [Google Scholar] [CrossRef]
- Douchkov, D. (Department of Breeding Research, Leibniz Institute of Plant Genetics and Crop Plant Research (IPK), Gatersleben, Germany). Personal communication. 2020. [Google Scholar]
- Gouache, D.; Roche, R.; Pieri, P.; Bancal, M.O. Evolution of some pathosystems on wheat and vines. In Climate Change, Agriculture and Forests in France: Simulations of the Impacts on the Main Species. The Green Book of the CLIMATOR Project (2007–2010); Brisson, N., Levrault, F., Eds.; ADEME: Angers, France, 2011; pp. 113–126. Available online: https://www.researchgate.net/publication/262233128 (accessed on 5 May 2021).
- Boyd, L.A.; Smith, P.H.; Hart, N. Mutants in wheat showing multipathogen resistance to biotrophic fungal pathogens. Plant Pathol. 2006, 55, 475–484. [Google Scholar] [CrossRef]
- Bariana, H.S.; McIntosh, R.A. Genetics of adult plant stripe rust resistance in four Australian wheats and the French cultivar ‘Hybride de Bersee’. Plant Breed. 1995, 114, 485–491. [Google Scholar] [CrossRef]
- Watson, A.; Ghosh, S.; Williams, M.J.; Cuddy, W.S.; Simmonds, J.; Rey, M.D.; Asyraf Md Hatta, M.; Hinchliffe, A.; Steed, A.; Reynolds, D.; et al. Speed breeding is a powerful tool to accelerate crop research and breeding. Nat. Plants 2018, 4, 23–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Nasenberg | Nörvenich | Seligenstadt1 | Seligenstadt | Söhnke- Nissenkoog | Wetze1 | Wetze2 | Wetze | Wohlde | |
|---|---|---|---|---|---|---|---|---|---|
| Elevation above sea level (m) | 148 | 106 | 283 | 283 | 1 | 136 | 136 | 136 | 73 |
| Average annual temperature (°C) | 10.69 | 11.25 | 11.02 | 11.02 | 9.77 | 10.13 | 10.13 | 10.13 | 9.94 |
| Average annual precipitation (mm) | 548.54 | 548.66 | 551.36 | 551.36 | 902.50 | 584.92 | 584.92 | 584.92 | 740.14 |
| Year (s) | 2017, 2018 | 2018 | 2017, 2018 | 2017 | 2017 | 2017, 2018 | 2018 | 2017, 2018 | 2017 |
| Plot size (m2) | 17.25 | 7.4 | 0.5 | 14.85, 6.6 | 0.5 | 0.5 | 0.5 | 14.25, 6.25 | 0.5 |
| Field design | Alpha-lattice | Alpha-lattice | Randomized complete block | Alpha-lattice, Moving grids | Randomized complete block | Randomized complete block | Randomized complete block | Alpha-lattice, Moving grids | Randomized complete block |
| No. of genotypes | 116, 119 | 119 | 780, 1138 | 64 | 56 | 780, 1135 | 1138 | 93, 64 | 780 |
| No. of trials | 2 | 2 | 10 | 1 | 1 | 10, 11 | 11 | 2, 11 | 10 |
| Replications | 1 | 1 | 2 | 1 | 2 | 2, 1 | 2 | 1, 2 | 2 |
| Genotypes per trial | 64 | 64 | 42–140 | 64 | 56 | 42–140 | 64–140 | 64, 64–140 | 42–140 |
| Genotypes of selected set | 27, 26 | 26 | 126, 128 | 19 | 5 | 126, 127 | 128 | 14, 17 | 126 |
| Seligenstadt1 2017 | Söhnke-Nissenkoog 2017 | Wetze1 2017 | Wohlde 2017 | Seligenstadt1 2018 | Wetze2 2018 | |
|---|---|---|---|---|---|---|
| Trial1 | 0.82 | - | 0.70 | 0.59 | 0.93 | 0.76 |
| Trial2 | 0.88 | - | 0.75 | 0.54 | 0.89 | 0.77 |
| Trial3 | 0.93 | 0.56 | 0.51 | 0.63 | - | - |
| Trial4 | 0.89 | - | 0.71 | 0.14 | 0.94 | 0.77 |
| Trial5 | 0.89 | - | 0.45 | 0.50 | 0.96 | 0.82 |
| Trial6 | 0.92 | - | 0.48 | 0.62 | - | - |
| Trial7 | 0.77 | - | 0.59 | 0.25 | 0.91 | 0.76 |
| Trial8 | 0.67 | - | 0.53 | 0.12 | 0.93 | 0.65 |
| Trial9 | 0.79 | - | 0.60 | 0.22 | 0.91 | 0.78 |
| Trial10 | 0.91 | - | 0.60 | 0.51 | - | 0.59 |
| Trial11 | - | - | - | - | 0.90 | 0.57 |
| Trial12 | - | - | - | - | 0.84 | 0.68 |
| Trial13 | - | - | - | - | 0.93 | 0.76 |
| Seligenstadt1 2017 | Seligenstadt 2017 | Söhnke- Nissenkoog 2017 | Wetze1 2017 | Wetze 2017 | Wohlde 2017 | Nasenberg 2018 | Nörvenich 2018 | Seligenstadt1 2018 | Wetze1 2018 | Wetze2 2018 | Wetze 2018 | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Nasenberg 2017 | 0.60 *** (116) | 0.56 *** (64) | −0.30 (17) | 0.56 *** (116) | 0.46 *** (93) | 0.42 *** (116) | 0.14 (44) | 0.06 (44) | 0.39 ** (46) | 0.26 (46) | 0.14 (46) | 0.61 * (12) |
| Seligenstadt1 2017 | - | 0.65 *** (64) | 0.28 * (56) | 0.64 *** (780) | 0.64 *** (93) | 0.53 *** (352) | 0.45 *** (84) | 0.34 ** (84) | 0.69 *** (92) | 0.53 *** (92) | 0.56 *** (92) | 0.48 *** (46) |
| Seligenstadt 2017 | - | −0.31 (9) | 0.63 *** (64) | 0.46 *** (51) | 0.48 *** (64) | 0.21 (16) | 0.72 ** (16) | 0.31 (17) | 0.50 * (17) | 0.41 (17) | 0.70* (11) | |
| Söhnke- Nissenkoog 2017 | - | 0.21 (56) | 0.39 (15) | 0.18 (56) | −0.04 (17) | 0.20 (17) | 0.19 (21) | 0.43 (21) | 0.29 (21) | 0 (8) | ||
| Wetze1 2017 | - | 0.54 *** (93) | 0.53 *** (352) | 0.48 *** (84) | 0.04 (84) | 0.73 *** (92) | 0.48 *** (92) | 0.53 *** (92) | 0.47 *** (46) | |||
| Wetze 2017 | - | 0.40 *** (93) | 0.33 * (37) | −0.12 (37) | 0.74 *** (39) | 0.68 *** (39) | 0.57 *** (39) | 0.58 (10) | ||||
| Wohlde 2017 | - | 0.53 *** (59) | 0.17 (59) | 0.40 ** (65) | 0.51 *** (65) | 0.33 ** (65) | 0.41 (21) | |||||
| Nasenberg 2018 | - | 0.14 (119) | 0.67 *** (119) | 0.52 *** (119) | 0.54 *** (119) | 0.50 *** (64) | ||||||
| Nörvenich 2018 | - | 0.04 (119) | 0.16 (119) | 0.06 (119) | 0.43 *** (64) | |||||||
| Seligenstadt1 2018 | - | 0.64 *** (1005) | 0.73 *** (1008) | 0.52 *** (64) | ||||||||
| Wetze1 2018 | - | 0.59 *** (1135) | 0.47 *** (64) | |||||||||
| Wetze2 2018 | - | 0.39 *** (64) |
| Analysis with Whole Data Set | All Genotypes of Plant Stage T1 | T1 | T2 | T3 | |
|---|---|---|---|---|---|
| No. of genotypes | 240 | 240 | 40 | 40 | 41 |
| No. inoculation groups | 18 | 12 | 12 | 2 | 2 |
| Range of infected leaf area (%) | 0.3–26.7 | 0.6–2.6 | 0.6–2.4 | 0.3–26.7 | 0.5–5.5 |
| Mean of infected leaf area (%) | 1.4 | 1.1 | 1.1 | 3.6 | 1.0 |
| 22.2% | 11.9% | 13.7% | 82.8% | 31.0% | |
| 23.8% | - | - | - | - | |
| 12.9% | 21.5% | 28.0% | 3.0% | 8.9% | |
| 14.2% | 46.5% | 39.9% | 0.0% | 14.4% | |
| 26.9% | 20.2% | 18.4% | 14.2% | 45.7% | |
| 0.64 | 0.54 | 0.60 | 0.92 | 0.58 |
| All Genotypes at Plant Stage T1 | T1 | T2 | T3 | |
|---|---|---|---|---|
| Pearson correlation to field data | 0.40 *** | 0.48 ** | 0.39 * | 0.34 * |
| p-value | 2.6 × 10−10 | 0.0018 | 0.013 | 0.032 |
| No. of genotypes | 232 | 40 | 40 | 41 |
| Correlation To Series 2017 | Correlation To Series 2018 | |
|---|---|---|
| Nasenberg | 0.63 *** (128) | 0.63 *** (128) |
| Nörvenich | - | 0.13 (128) |
| Seligenstadt | 0.69 *** (64) | - |
| Seligenstadt1 | 0.64 *** (780) | 0.74 *** (1008) |
| Söhnke-Nissenkoog | 0.23 (56) | - |
| Wetze | 0.58 *** (104) | - |
| Wetze1 | 0.62 *** (780) | 0.62 *** (64) |
| Wetze2 | - | 0.72 *** (1008) |
| Wohlde | 0.58 *** (352) | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Beukert, U.; Pfeiffer, N.; Ebmeyer, E.; Hinterberger, V.; Lueck, S.; Serfling, A.; Ordon, F.; Schulthess, A.W.; Reif, J.C. Efficiency of a Seedling Phenotyping Strategy to Support European Wheat Breeding Focusing on Leaf Rust Resistance. Biology 2021, 10, 628. https://doi.org/10.3390/biology10070628
Beukert U, Pfeiffer N, Ebmeyer E, Hinterberger V, Lueck S, Serfling A, Ordon F, Schulthess AW, Reif JC. Efficiency of a Seedling Phenotyping Strategy to Support European Wheat Breeding Focusing on Leaf Rust Resistance. Biology. 2021; 10(7):628. https://doi.org/10.3390/biology10070628
Chicago/Turabian StyleBeukert, Ulrike, Nina Pfeiffer, Erhard Ebmeyer, Valentin Hinterberger, Stefanie Lueck, Albrecht Serfling, Frank Ordon, Albert Wilhelm Schulthess, and Jochen Christoph Reif. 2021. "Efficiency of a Seedling Phenotyping Strategy to Support European Wheat Breeding Focusing on Leaf Rust Resistance" Biology 10, no. 7: 628. https://doi.org/10.3390/biology10070628