Consequences of Haemorrhagic Smolt Syndrome (HSS) for the Immune Status of Atlantic salmon (Salmo salar L.) (Case Study)

 ,
,
Abstract
:1. Introduction
2. Material and Methods
2.1. Fish
2.2. Microarray
2.3. Ig-seq
2.4. Direct ELISA for Total IgM and Blood Cell Count
2.5. Bead Coupling and Multiplex Immunoassay
2.6. Ethical Statement
3. Results
3.1. Microarray
3.2. Ig-seq, Blood Cell Count, and Serological Immunoassays
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Rodger, H.D.; Richards, R.H. Haemorrhagic smolt syndrome: A severe anaemic condition in farmed salmon in Scotland. Vet. Rec. 1998, 142, 538–541. [Google Scholar] [CrossRef] [PubMed]
- Nylund, A.; Plarre, H.; Hodneland, K.; Devold, M.; Aspehaug, V.; Aarseth, M.; Koren, C.; Watanabe, K. Haemorrhagic smolt syndrome (HSS) in Norway: Pathology and associated virus-like particles. Dis. Aquat. Organ. 2003, 54, 15–27. [Google Scholar] [CrossRef] [PubMed]
- Krasnov, A.; Timmerhaus, G.; Afanasyev, S.; Jørgensen, S.M. Development and assessment of oligonucleotide microarrays for Atlantic salmon (Salmo salar L.). Comp. Biochem. Physiol. Part D Genom. Proteom. 2011, 6, 31–38. [Google Scholar] [CrossRef] [PubMed]
- Krasnov, A.; Jørgensen, S.M.; Afanasyev, S. Ig-seq: Deep sequencing of the variable region of Atlantic salmon IgM heavy chain transcripts. Mol. Immunol. 2017, 88, 99–105. [Google Scholar] [CrossRef] [PubMed]
- Brochet, X.; Lefranc, M.P.; Giudicelli, V. IMGT/V-QUEST: The highly customized and integrated system for IG and TR standardized V-J and V-D-J sequence analysis. Nucleic Acids Res. 2008, 36, W503–W508. [Google Scholar] [CrossRef] [Green Version]
- Magnadottir, B. Comparison of immunoglobulin (IgM) from four fish species. Icel. Agric. Sci. 1998, 12, 47–59. [Google Scholar]
- Strømsheim, A.; Eide, D.M.; Hofgaard, P.O.; Larsen, H.J.S.; Refstie, T.; Røed, K.H. Genetic variation in the humoral immune response against Vibrio salmonicida and in antibody titre against Vibrio anguillarum and total IgM in Atlantic salmon (Salmo salar). Vet. Immunol. Immunopathol. 1994, 44, 85–95. [Google Scholar] [CrossRef]
- Lund, H.; Bakke, A.F.; Sommerset, I.; Afanasyev, S.; Schriwer, G.; Thorisdottir, A.; Boysen, P.; Krasnov, A. A time-course study of gene expression and antibody repertoire at early time post vaccination of Atlantic salmon. Mol. Immunol. 2019, 106, 99–107. [Google Scholar] [CrossRef]
- He, H.; Li, J.; Weng, S.; Li, M.; Yu, Y. S100A11: Diverse function and pathology corresponding to different target proteins. Cell Biochem. Biophys. 2009, 55, 117–126. [Google Scholar] [CrossRef]
- Cerezo, L.A.; Šumová, B.; Prajzlerová, K.; Veigl, D.; Damgaard, D.; Nielsen, C.H.; Pavelka, K.; Vencovský, J.; Šenolt, L. Calgizzarin (S100A11): A novel inflammatory mediator associated with disease activity of rheumatoid arthritis. Arthritis Res. Ther. 2017, 19, 79. [Google Scholar] [CrossRef] [Green Version]
- Lee, C.G.; Da Silva, C.A.; Dela Cruz, C.S.; Ahangari, F.; Ma, B.; Kang, M.J.; He, C.H.; Takyar, S.; Elias, J.A. Role of chitin and chitinase/chitinase-like proteins in inflammation, tissue remodeling, and injury. Annu. Rev. Physiol. 2011, 73, 479–501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aratani, Y. Myeloperoxidase: Its role for host defense, inflammation, and neutrophil function. Arch. Biochem. Biophys. 2018, 640, 47–52. [Google Scholar] [CrossRef] [PubMed]
- Mu, Y.; Wang, K.; Ao, J.; Chen, X. Molecular characterization and biological effects of a CXCL8 homologue in large yellow croaker (Larimichthys crocea). Fish Shellfish Immunol. 2015, 44, 462–470. [Google Scholar] [CrossRef] [PubMed]
- Manilay, J.O.; Zouali, M. Tight relationships between B lymphocytes and the skeletal system. Trends Mol. Med. 2014, 20, 405–412. [Google Scholar] [CrossRef]
- Yun, T.J.; Tallquist, M.D.; Aicher, A.; Rafferty, K.L.; Marshall, A.J.; Moon, J.J.; Ewings, M.K.; Mohaupt, M.; Herring, S.W.; Clark, E.A. Osteoprotegerin, a crucial regulator of bone metabolism, also regulates B cell development and function. J. Immunol. 2001, 166, 1482–1491. [Google Scholar] [CrossRef] [Green Version]
- Chi, Y.; Huang, Z.; Chen, Q.; Xiong, X.; Chen, K.; Xu, J.; Zhang, Y.; Zhang, W. Loss of runx1 function results in B cell immunodeficiency but not T cell in adult zebrafish. Open Biol. 2018, 8, 180043. [Google Scholar] [CrossRef] [Green Version]
- Shulga-Morskaya, S.; Dobles, M.; Walsh, M.E.; Ng, L.G.; MacKay, F.; Rao, S.P.; Kalled, S.L.; Scott, M.L. B cell-activating factor belonging to the TNF family acts through separate receptors to support B cell survival and T cell-independent antibody formation. J. Immunol. 2004, 173, 2331–2341. [Google Scholar] [CrossRef] [Green Version]
- Kim, C.H. Control of Innate and Adaptive Lymphocytes by the RAR-Retinoic Acid Axis. Immune Netw. 2018, 18, e1. [Google Scholar] [CrossRef] [Green Version]
- Krasnov, A.; Timmerhaus, G.; Schiøtz, B.L.; Torgersen, J.; Afanasyev, S.; Iliev, D.; Jørgensen, J.; Takle, H.; Jørgensen, S.M. Genomic survey of early responses to viruses in Atlantic salmon, Salmo salar L. Mol. Immunol. 2011, 49, 163–174. [Google Scholar] [CrossRef]
- Sobhkhez, M.; Krasnov, A.; Chang, C.J.; Robertsen, B. Transcriptome analysis of plasmid-induced genes sheds light on the role of type I IFN as adjuvant in DNA vaccine against infectious salmon anemia virus. PLoS ONE 2017, 12, e0188456. [Google Scholar] [CrossRef] [Green Version]
- Arias-Jayo, N.; Abecia, L.; Alonso-Sáez, L.; Ramirez-Garcia, A.; Rodriguez, A.; Pardo, M.A. High-Fat Diet Consumption Induces Microbiota Dysbiosis and Intestinal Inflammation in Zebrafish. Microb. Ecol. 2018, 76, 1089–1101. [Google Scholar] [CrossRef] [PubMed]
- Coll, J.M. Herpesvirus Infection Induces both Specific and Heterologous Antiviral Antibodies in Carp. Front. Immunol. 2018, 9, 39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Magnadóttir, B.; Gudmundsdóttir, S.; Gudmundsdóttir, B.K. Study of the humoral response of Atlantic salmon (Salmo salar L.), naturally infected with Aeromonas salmonicida ssp. achromogenes. Vet. Immunol. Immunopathol. 1995, 49, 127–142. [Google Scholar]
- Magnadottir, B.; Gudmundsdottir, S.; Gudmundsdottir, B.K.; Helgason, S. Natural antibodies of cod (Gadus morhua L.): Specificity, activity and affinity. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2009, 154, 309–316. [Google Scholar] [CrossRef] [PubMed]
- Lund, M.; Røsæg, M.V.; Krasnov, A.; Timmerhaus, G.; Nyman, I.B.; Aspehaug, V.; Rimstad, E.; Dahle, M.K. Experimental Piscine orthoreovirus infection mediates protection against pancreas disease in Atlantic salmon (Salmo salar). Vet. Res. 2016, 47, 107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Røsæg, M.V.; Lund, M.; Nyman, I.B.; Markussen, T.; Aspehaug, V.; Sindre, H.; Dahle, M.K.; Rimstad, E. Immunological interactions between Piscine orthoreovirus and Salmonid alphavirus infections in Atlantic salmon. Fish Shellfish Immunol. 2017, 64, 308–319. [Google Scholar] [CrossRef] [PubMed]
- Teige, L.H.; Subramaniam, K.; Johansen, G.M.; Wessel, Ø.; Vendramin, N.; Lund, M.; Rimstad, E.; Boysen, P.; Dahle, M.K. Detection of Salmonid IgM Specific to the Piscine Orthoreovirus Outer Capsid Spike Protein Sigma 1 Using Lipid-Modified Antigens in a Bead-Based Antibody Detection Assay. Front. Immunol. 2019, 10, 2119. [Google Scholar] [CrossRef] [Green Version]
- Sadrzadeh, S.M.; Graf, E.; Panter, S.S.; Hallaway, P.E.; Eaton, J.W. Hemoglobin. A biologic fenton reagent. J. Biol. Chem. 1984, 259, 14354–14356. [Google Scholar]
- Nilsen, T.O.; Ebbesson, L.O.; Madsen, S.S.; McCormick, S.D.; Andersson, E.; Björnsson, B.T.; Prunet, P.; Stefansson, S.O. Differential expression of gill Na+,K+-ATPase alpha- and beta-subunits, Na+,K+,2Cl- cotransporter and CFTR anion channel in juvenile anadromous and landlocked Atlantic salmon Salmo salar. J. Exp. Biol. 2007, 210 Pt 16, 2885–2896. [Google Scholar] [CrossRef] [Green Version]
- McCormick, S.D.; Regish, A.M.; Christensen, A.K.; Björnsson, B.T. Differential regulation of sodium-potassium pump isoforms during smolt development and seawater exposure of Atlantic salmon. J. Exp. Biol. 2013, 216 Pt 7, 1142–1151. [Google Scholar] [CrossRef] [Green Version]
- Johansson, L.H.; Timmerhaus, G.; Afanasyev, S.; Jørgensen, S.M.; Krasnov, A. Smoltification and seawater transfer of Atlantic salmon (Salmo salar L.) is associated with systemic repression of the immune transcriptome. Fish Shellfish Immunol. 2016, 58, 33–41. [Google Scholar] [CrossRef] [PubMed]




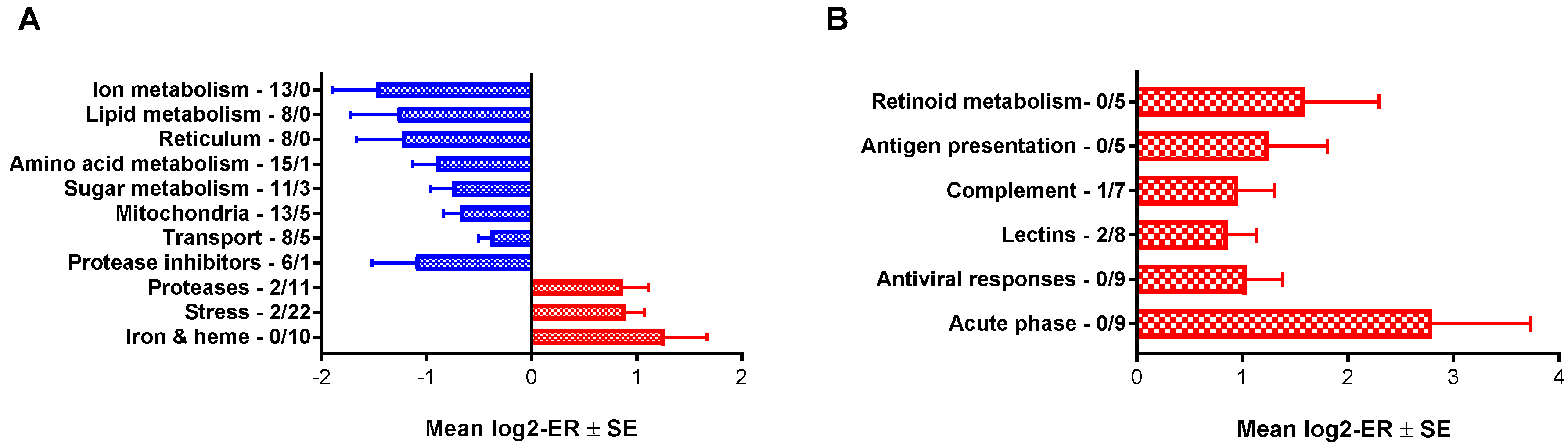{kind=link}
{kind=link}
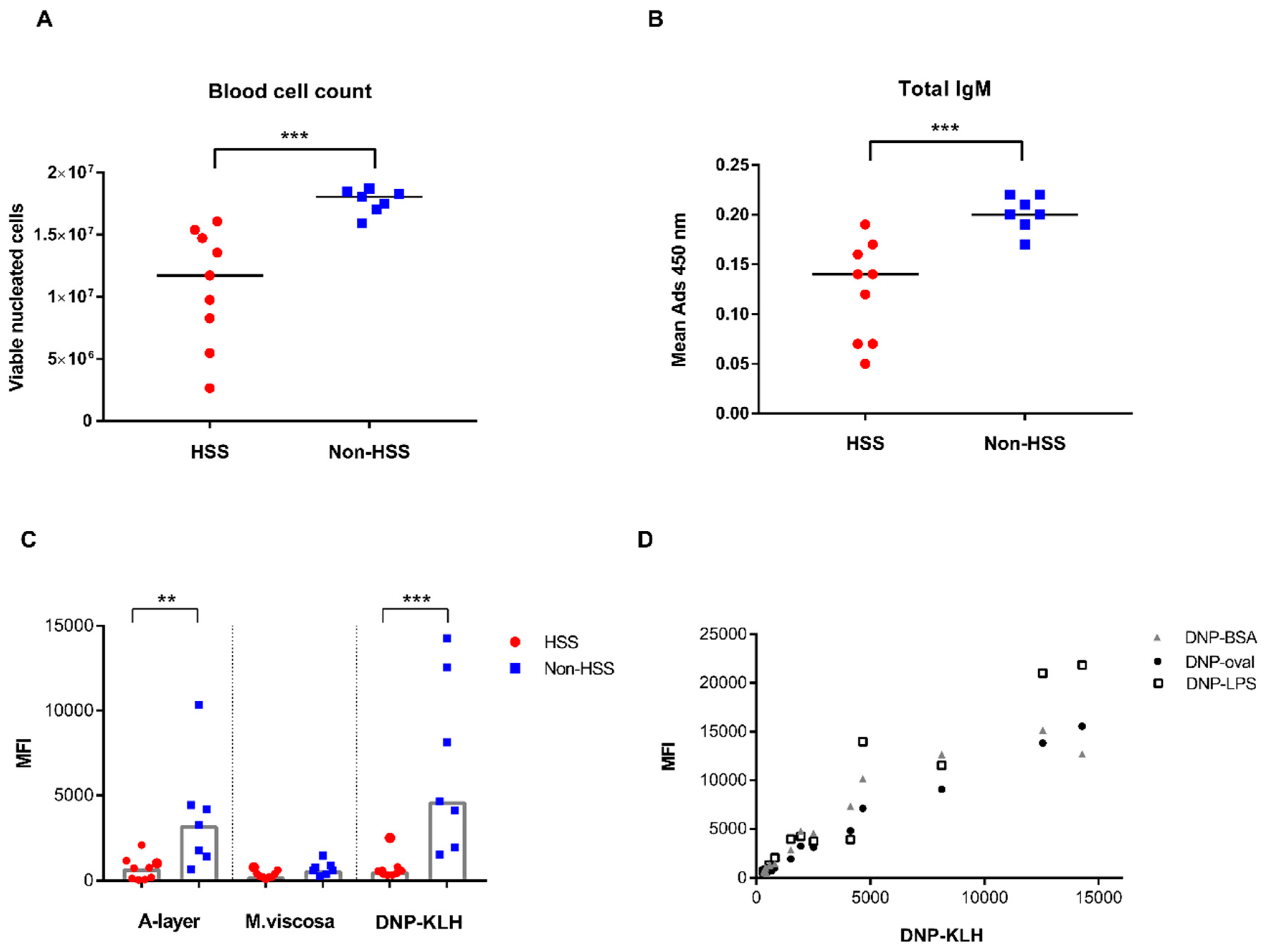{kind=link}
| Fold Difference HSS/non-HSS | |
|---|---|
| Stress and iron metabolism | |
| Ferritin, heavy polypeptide 1b (fth1b, 4 genes) | 1.8–2.1 |
| Haptoglobin (hp, 3 genes) | 6.3–6.7 |
| Heme oxygenase (ho, 4 genes) | 1.8–4.7 |
| Hepcidin-1 (hepc1) | 3.5 |
| Glucose-regulated protein 78 kDa (grp78) | 2.2 |
| Glucose-regulated protein 94 kDa (grp94) | 2.0 |
| Heat shock cognate 70 (hsc70, 7 genes) | 2.0–2.5 |
| Heat shock protein 90, alpha (hsp90a) | 1.8 |
| C/EBP beta (2 genes) | 2.1–2.2 |
| C/EBP delta | 2.6 |
| GADD45 beta (2 genes) | 1.9–2.1 |
| Hypoxia-inducible factor 1a (hif1a, 2 genes) | 2.2–2.4 |
| Jun B-1 | 1.9 |
| Cathepsin L1 (catl1, 2 genes) | 2.6–2.8 |
| Cathepsin S (cats) | 2.3 |
| Cathepsin Z (catz) | 1.8 |
| Ion metabolism and transport | |
| S100-A11 (2 genes) | 7.6–14.4 |
| Na+/K+ ATPase, b1a (at1b1, 2 genes) | −(1.9–2.7) |
| Na+/K+ ATPase a2 (at1a2, 3 genes) | 1.8–2.3 |
| Aquaporin 8b (aqp8b, OrthoDB) | −2.9 |
| Multidrug resistance-associated protein (mrp) | −2.9 |
| Solute carrier family 5 member 8a (slc5a8) | −2.0 |
| Solute carrier family 13 member 3 (slc13a3) | −2.7 |
| Solute carrier family 22 member 4 (slc22a4) | −2.6 |
| Solute carrier family 22 member 6-A (slc22a6) | −1.8 |
| Solute carrier family 22 member 7 (slc22a7) | −2.9 |
| Solute carrier family 34 (slc34) | −2.0 |
| Immune Genes | Fold Difference HSS/non-HSS |
|---|---|
| C-C motif chemokine 13 | 3.6 |
| C-C motif chemokine 19-4 | 5.2 |
| Serum amyloid A5 (saa5) | 6.2 |
| C type lectin receptor A | 3.1 |
| Cathelicidin (camp) | 6.1 |
| C-type lectin 4E (clec4e) | 6.1 |
| Macrophage receptor MARCO (mrc1) | 2.5 |
| Mannose receptor | 2.6 |
| C1q components b, c (3 genes) | 1.9–3.1 |
| C1q-like (2 genes) | 15.7–17.4 |
| C1q-like 2 (c1ql2) | 5.0 |
| C6 | 2.1 |
| H-2 class II HC antigen, a chain (hla-dpa) | 2.8 |
| H-2 class II HC antigen, g chain (hla-dpg, 2 genes) | 1.9–2.2 |
| CD274 | 2.4 |
| CD28 | 3.0 |
| CD4-like | 3.4 |
| TNF receptor superfamily member 11B (tnfrsf11b) | 19.6 |
| Retinol-binding protein 1a, cellular (ret1a) | 3.0 |
| Retinol-binding protein 4 (ret4, 2 features) | 3.7–4.0 |
| Chitinase, acidic.3 (chia) | −7.5 |
| Myeloperoxidase (mpo) | −2.0 |
| IL8 | −2.7 |
| Mitogen-activated protein kinase 12b (mapk12b) | −3.1 |
| Mitogen-activated protein kinase-activated protein kinase 3 (mapkapk3) | 2.8 |
| TNF receptor superfamily member 13b (tnfrsf13b) | −2.2 |
| Early B-cell factor 1a (ebf1a) | −4.0 |
| TNF receptor superfamily member 14 (tnfrsf14) | −2.3 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Krasnov, A.; Sommerset, I.; Søfteland, T.; Afanasyev, S.; Boysen, P.; Lund, H. Consequences of Haemorrhagic Smolt Syndrome (HSS) for the Immune Status of Atlantic salmon (Salmo salar L.) (Case Study). Biology 2020, 9, 1. https://doi.org/10.3390/biology9010001
Krasnov A, Sommerset I, Søfteland T, Afanasyev S, Boysen P, Lund H. Consequences of Haemorrhagic Smolt Syndrome (HSS) for the Immune Status of Atlantic salmon (Salmo salar L.) (Case Study). Biology. 2020; 9(1):1. https://doi.org/10.3390/biology9010001
Chicago/Turabian StyleKrasnov, Aleksei, Ingunn Sommerset, Tina Søfteland, Sergey Afanasyev, Preben Boysen, and Hege Lund. 2020. "Consequences of Haemorrhagic Smolt Syndrome (HSS) for the Immune Status of Atlantic salmon (Salmo salar L.) (Case Study)" Biology 9, no. 1: 1. https://doi.org/10.3390/biology9010001