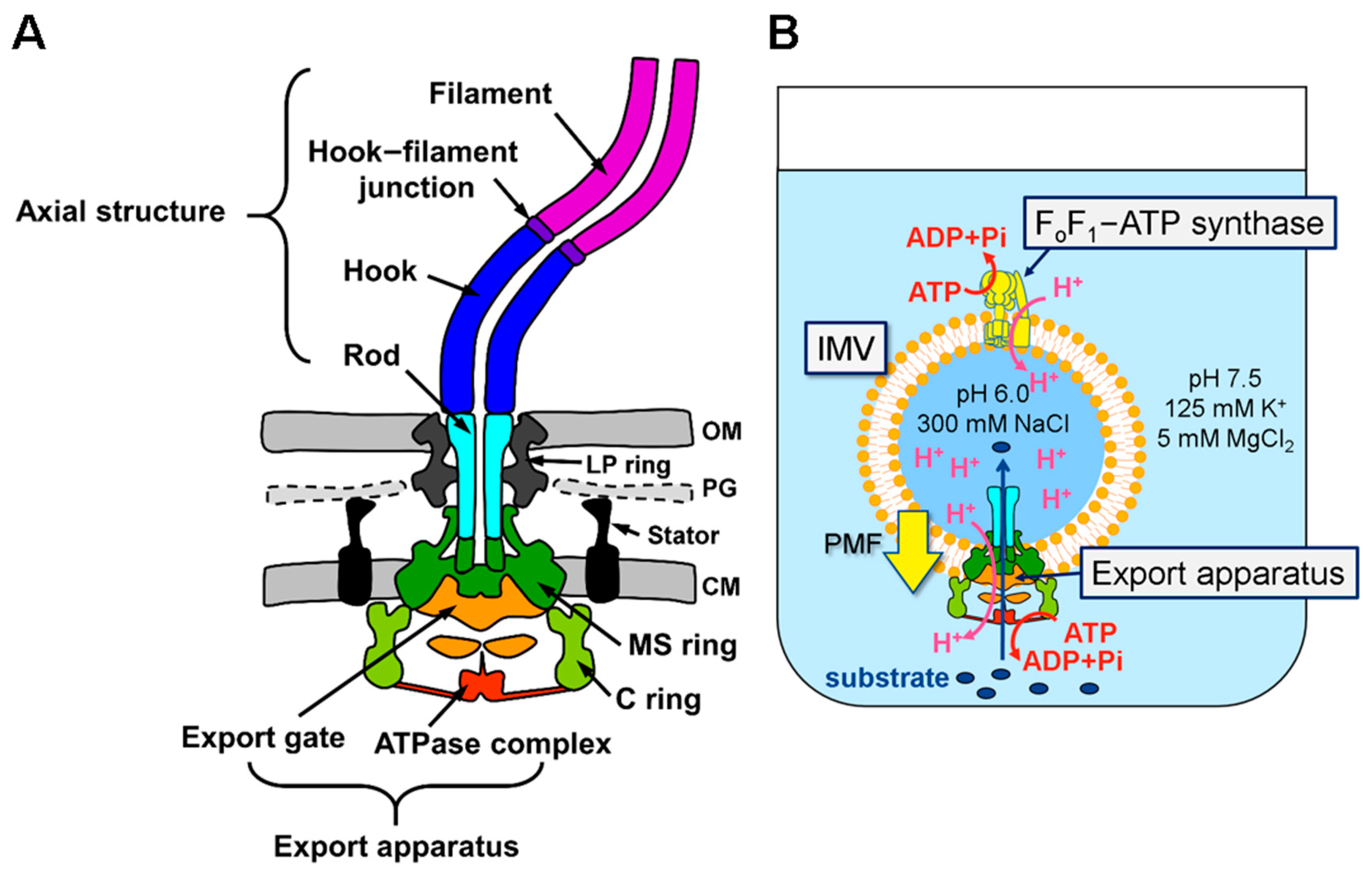
In Vitro Autonomous Construction of the Flagellar Axial Structure in Inverted Membrane Vesicles

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacteria Strains and Plasmids
2.2. Preparation of Inverted Membrane Vesicles
2.3. Protein Purification
2.4. Transport Assay
2.5. Purification of the Hook–Basal Body from IMV
2.6. Dynamic Light Scattering
2.7. Negative-Staining Electron Microscopy
3. Results
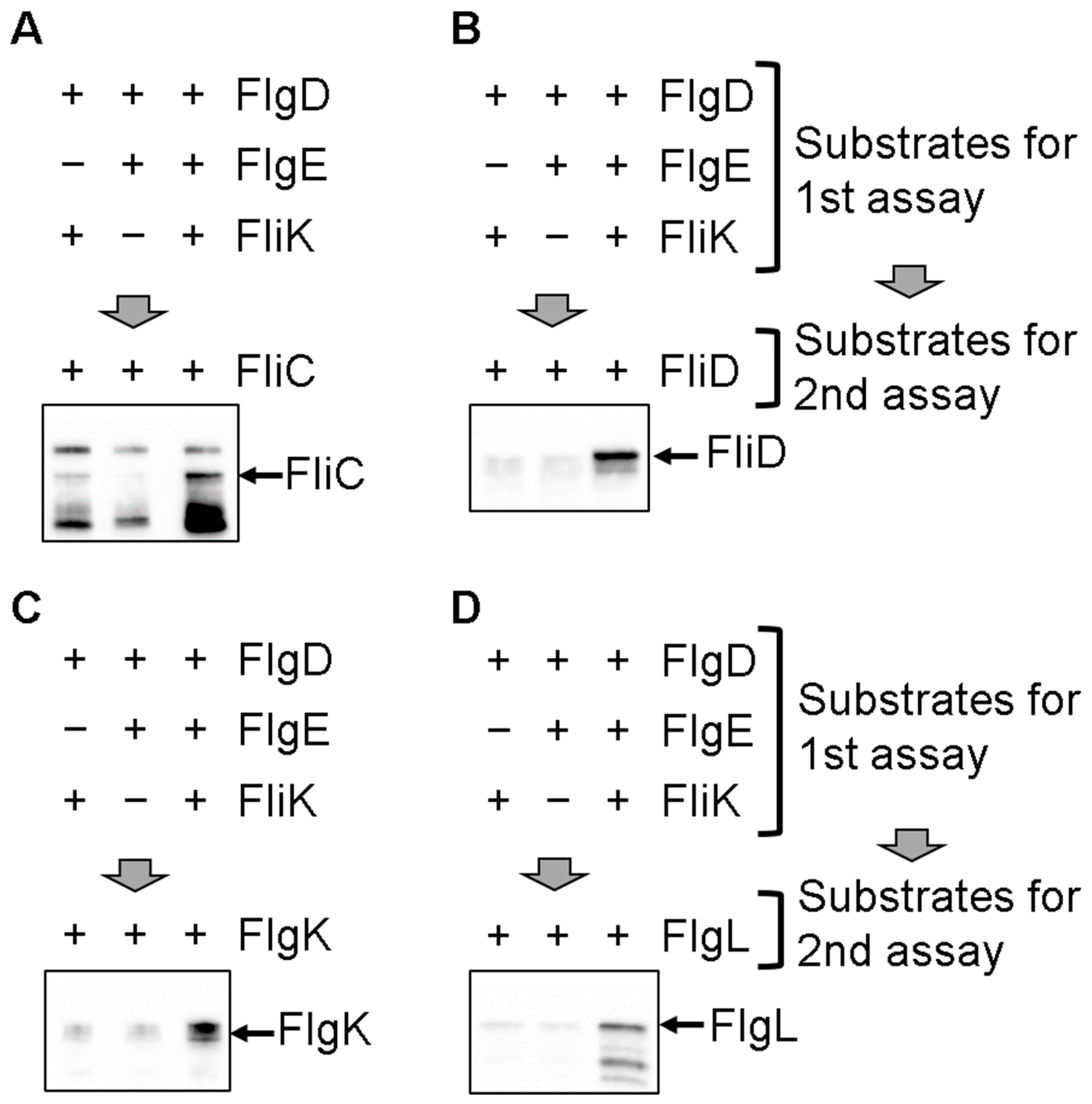
3.1. In Vitro Protein Transport of the Filament-Type Proteins into the IMVs
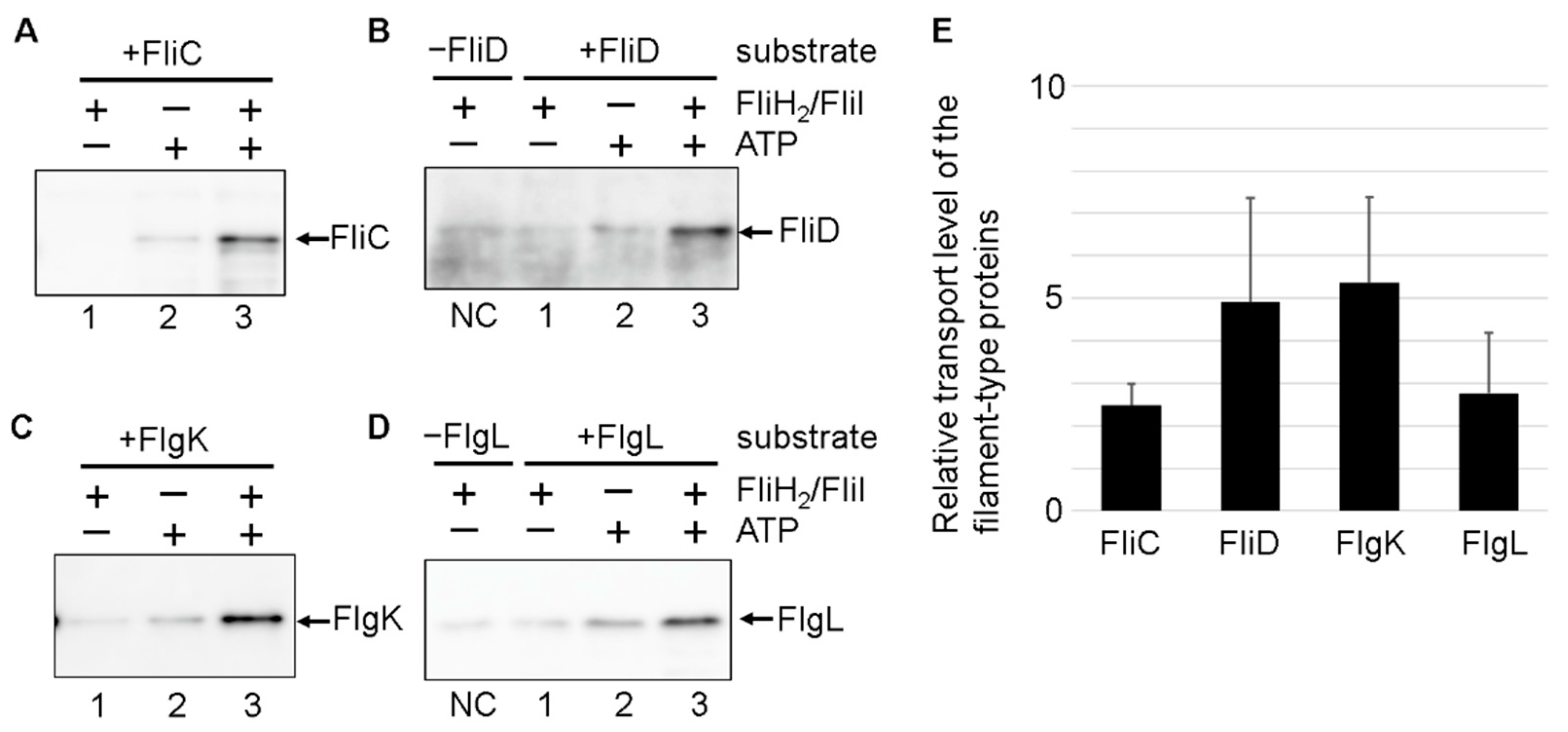
3.2. Effect of the FliH2/FliI Complex on Filament-Type Protein Export
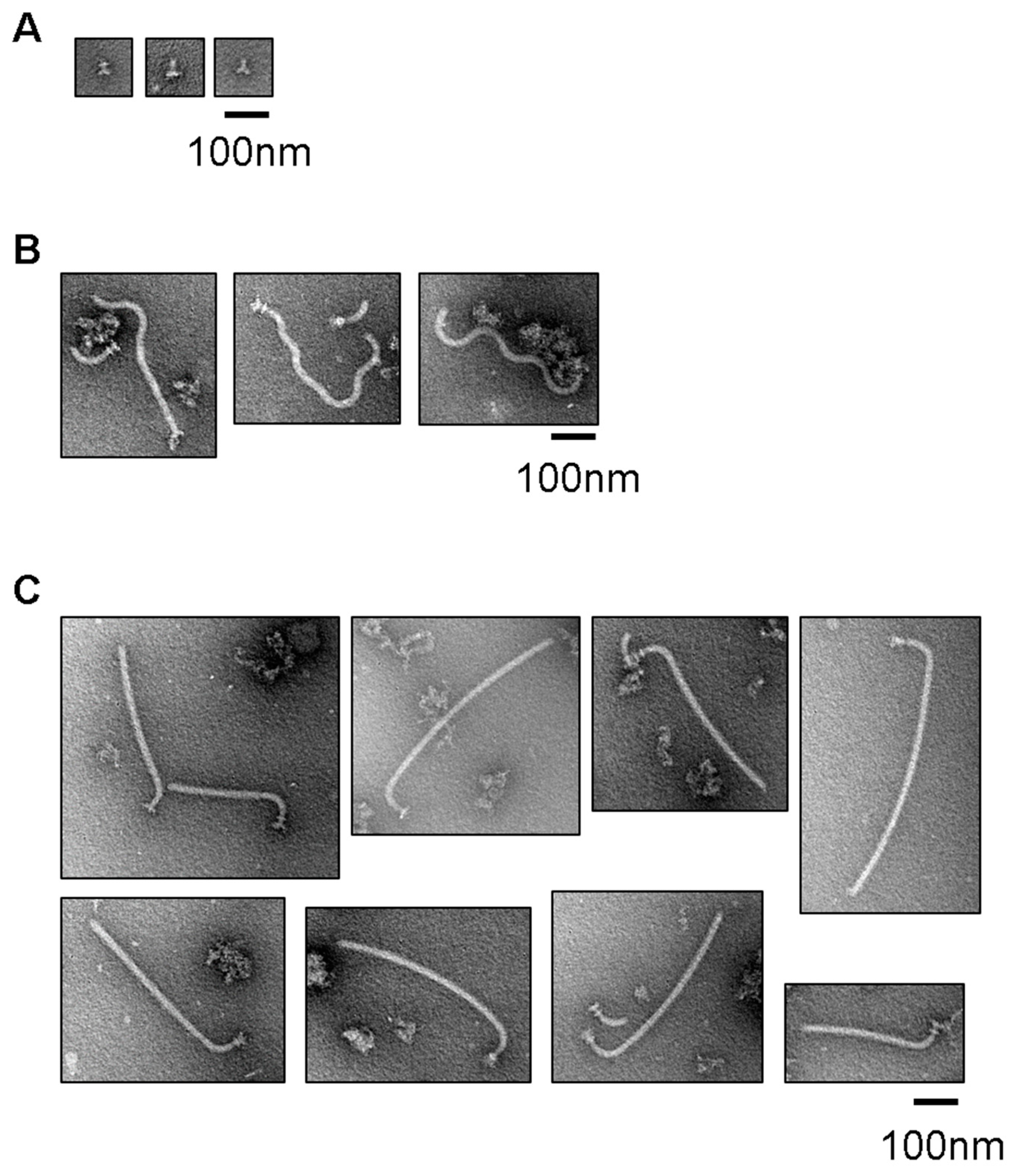
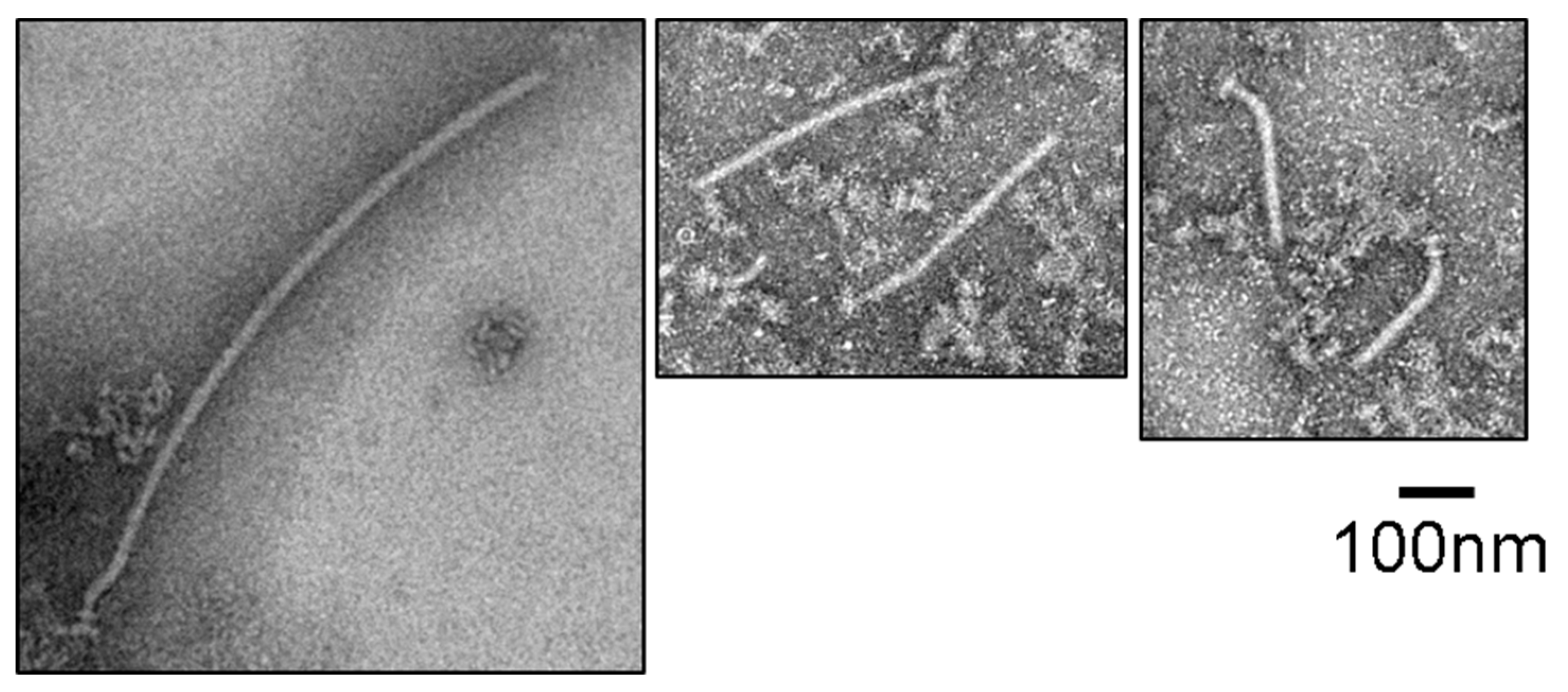
3.3. Filament Structure Formation on the Hook in the IMVs
3.4. Effect of Uncoupling of Flagellar Expression with Assembly on the Entire Assembly Process of the Hook–Filament Complex in the IMVs
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Macnab, R.M. How bacteria assemble flagella. Annu. Rev. Microbiol. 2003, 57, 77–100. [Google Scholar] [CrossRef] [PubMed]
- Minamino, T. Protein export through the bacterial flagellar type III export pathway. Biochim. Biophys. Acta 2014, 1843, 1642–1648. [Google Scholar] [CrossRef] [Green Version]
- Terashima, H.; Kojima, S.; Homma, M. Flagellar motility in bacteria: Structure and function of flagellar motor. Int. Rev. Cell. Mol. Biol. 2008, 270, 39–85. [Google Scholar] [PubMed]
- Iino, T. Assembly of Salmonella flagellin in vitro and in vivo. J. Supramol. Struct. 1974, 2, 372–384. [Google Scholar] [PubMed]
- Zhao, Z.; Zhao, Y.; Zhuang, X.Y.; Lo, W.C.; Baker, M.A.B.; Lo, C.J.; Bai, F. Frequent pauses in Escherichia coli flagella elongation revealed by single cell real-time fluorescence imaging. Nat. Commun. 2018, 9, 1885. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, S.; Beeby, M.; Murphy, G.E.; Leadbetter, J.R.; Hendrixson, D.R.; Briegel, A.; Li, Z.; Shi, J.; Tocheva, E.I.; Müller, A.; et al. Structural diversity of bacterial flagellar motors. EMBO J. 2011, 30, 2972–2981. [Google Scholar] [CrossRef] [Green Version]
- Kawamoto, A.; Morimoto, Y.V.; Miyata, T.; Minamino, T.; Hughes, K.T.; Kato, T.; Namba, K. Common and distinct structural features of Salmonella injectisome and flagellar basal body. Sci. Rep. 2013, 3, 3369. [Google Scholar] [CrossRef] [Green Version]
- Minamino, T.; Doi, H.; Kutsukake, K. Substrate specificity switching of the flagellum-specific export apparatus during flagellar morphogenesis in Salmonella typhimurium. Biosci. Biotechnol. Biochem. 1999, 63, 1301–1303. [Google Scholar] [CrossRef] [Green Version]
- Hirano, T.; Minamino, T.; Namba, K.; Macnab, R.M. Substrate specificity classes and the recognition signal for Salmonella type III flagellar export. J. Bacteriol. 2003, 185, 2485–2492. [Google Scholar] [CrossRef] [Green Version]
- Hirano, T.; Yamaguchi, S.; Oosawa, K.; Aizawa, S. Roles of FliK and FlhB in determination of flagellar hook length in Salmonella typhimurium. J. Bacteriol. 1994, 176, 5439–5449. [Google Scholar] [CrossRef] [Green Version]
- Moriya, N.; Minamino, T.; Hughes, K.T.; Macnab, R.M.; Namba, K. The type III flagellar export specificity switch is dependent on FliK ruler and a molecular clock. J. Mol. Biol. 2006, 359, 466–477. [Google Scholar] [CrossRef] [PubMed]
- Shibata, S.; Takahashi, N.; Chevance, F.F.; Karlinsey, J.E.; Hughes, K.T.; Aizawa, S. FliK regulates flagellar hook length as an internal ruler. Mol. Microbiol. 2007, 64, 1404–1415. [Google Scholar] [CrossRef] [PubMed]
- Kutsukake, K.; Minamino, T.; Yokoseki, T. Isolation and characterization of FliK-independent flagellation mutants from Salmonella typhimurium. J. Bacteriol. 1994, 176, 7625–7629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, A.W.; Yamaguchi, S.; Togashi, F.; Aizawa, S.I.; Kawagishi, I.; Macnab, R.M. Mutations in fliK and flhB affecting flagellar hook and filament assembly in Salmonella typhimurium. J. Bacteriol. 1996, 178, 2960–2970. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minamino, T.; González-Pedrajo, B.; Yamaguchi, K.; Aizawa, S.I.; Macnab, R.M. FliK, the protein responsible for flagellar hook length control in Salmonella, is exported during hook assembly. Mol. Microbiol. 1999, 34, 295–304. [Google Scholar] [CrossRef]
- Erhardt, M.; Singer, H.M.; Wee, D.H.; Keener, J.P.; Hughes, K.T. An infrequent molecular ruler controls flagellar hook length in Salmonella enterica. EMBO J. 2011, 30, 2948–2961. [Google Scholar] [CrossRef] [Green Version]
- Hirano, T.; Shibata, S.; Ohnishi, K.; Tani, T.; Aizawa, S. N-terminal signal region of FliK is dispensable for length control of the flagellar hook. Mol. Microbiol. 2005, 56, 346–360. [Google Scholar] [CrossRef]
- Minamino, T.; Ferris, H.U.; Moriya, N.; Kihara, M.; Namba, K. Two parts of the T3S4 domain of the hook-length control protein FliK are essential for the substrate specificity switching of the flagellar type III export apparatus. J. Mol. Biol. 2006, 362, 1148–1158. [Google Scholar] [CrossRef]
- Kinoshita, M.; Aizawa, S.-I.; Inoue, Y.; Namba, K.; Minamino, T. The role of intrinsically disordered C-terminal region of FliK in substrate specificity switching of the bacterial flagellar type III export apparatus. Mol. Microbiol. 2017, 105, 572–588. [Google Scholar] [CrossRef]
- Terahara, N.; Inoue, Y.; Kodera, N.; Morimoto, Y.V.; Uchihashi, T.; Imada, K.; Ando, T.; Namba, K.; Minamino, T. Insight into structural remodeling of the FlhA ring responsible for bacterial flagellar type III protein export. Sci. Adv. 2018, 4, eaao7054. [Google Scholar] [CrossRef] [Green Version]
- Inoue, Y.; Ogawa, Y.; Kinoshita, M.; Terahara, N.; Shimada, M.; Kodera, N.; Ando, T.; Namba, K.; Kitao, A.; Imada, K.; et al. Structural insight into the substrate specificity switch mechanism of the type III protein export apparatus. Structure 2018, 27, 965–976. [Google Scholar] [CrossRef] [PubMed]
- Minamino, T.; Inoue, Y.; Kinoshita, M.; Namba, K. FliK-driven conformational rearrangements of FlhA and FlhB are rquired for export switching of the flagellar potein export apparatus. J. Bacteriol. 2020, 202. [Google Scholar] [CrossRef]
- Fraser, G.M.; Bennett, J.C.; Hughes, C. Substrate-specific binding of hook-associated proteins by FlgN and FliT, putative chaperones for flagellum assembly. Mol. Microbiol. 1999, 32, 569–580. [Google Scholar] [CrossRef] [PubMed]
- Auvray, F.; Thomas, J.; Fraser, G.M.; Hughes, C. Flagellin polymerisation control by a cytosolic export chaperone. J. Mol. Biol. 2001, 308, 221–229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aldridge, P.; Karlinsey, J.; Hughes, K.T. The type III secretion chaperone FlgN regulates flagellar assembly via a negative feedback loop containing its chaperone substrates FlgK and FlgL. Mol. Microbiol. 2003, 49, 1333–1345. [Google Scholar] [CrossRef] [PubMed]
- Yokoseki, T.; Iino, T.; Kutsukake, K. Negative regulation by fliD, fliS, and fliT of the export of the flagellum-specific anti-sigma factor, FlgM, in Salmonella typhimurium. J. Bacteriol. 1996, 178, 899–901. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karlinsey, J.E.; Lonner, J.; Brown, K.L.; Hughes, K.T. Translation/secretion coupling by type III secretion systems. Cell 2000, 102, 487–497. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, S.; Kutsukake, K. FliT acts as an anti-FlhD2C2 factor in the transcriptional control of the flagellar regulon in Salmonella enterica serovar typhimurium. J. Bacteriol. 2006, 188, 6703–6708. [Google Scholar] [CrossRef] [Green Version]
- Terashima, H.; Kawamoto, A.; Tatsumi, C.; Namba, K.; Minamino, T.; Imada, K. In vitro reconstitution of functional type III protein export and insights into flagellar assembly. MBio 2018, 9. [Google Scholar] [CrossRef] [Green Version]
- Terashima, H.; Imada, K. Novel insight into an energy transduction mechanism of the bacterial flagellar type III protein export. Biophys. Phys. 2018, 15, 173–178. [Google Scholar] [CrossRef] [Green Version]
- Minamino, T.; Namba, K. Distinct roles of the FliI ATPase and proton motive force in bacterial flagellar protein export. Nature 2008, 451, 485–488. [Google Scholar] [CrossRef] [PubMed]
- Paul, K.; Erhardt, M.; Hirano, T.; Blair, D.F.; Hughes, K.T. Energy source of flagellar type III secretion. Nature 2008, 451, 489–492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minamino, T.; Morimoto, Y.V.; Hara, N.; Namba, K. An energy transduction mechanism used in bacterial flagellar type III protein export. Nat. Commun. 2011, 2, 475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minamino, T.; Morimoto, Y.V.; Kinoshita, M.; Aldridge, P.D.; Namba, K. The bacterial flagellar protein export apparatus processively transports flagellar proteins even with extremely infrequent ATP hydrolysis. Sci. Rep. 2014, 4, 7579. [Google Scholar] [CrossRef] [Green Version]
- Ibuki, T.; Imada, K.; Minamino, T.; Kato, T.; Miyata, T.; Namba, K. Common architecture of the flagellar type III protein export apparatus and F- and V-type ATPases. Nat. Struct. Mol. Biol. 2011, 18, 277–282. [Google Scholar] [CrossRef]
- Imada, K.; Minamino, T.; Uchida, Y.; Kinoshita, M.; Namba, K. Insight into the flagella type III export revealed by the complex structure of the type III ATPase and its regulator. Proc. Natl. Acad. Sci. USA 2016, 113, 3633–3638. [Google Scholar] [CrossRef] [Green Version]
- Kinoshita, M.; Nakanishi, Y.; Furukawa, Y.; Namba, K.; Imada, K.; Minamino, T. Rearrangements of α-helical structures of FlgN chaperone control the binding affinity for its cognate substrates during flagellar type III export. Mol. Microbiol. 2016, 101, 656–670. [Google Scholar] [CrossRef]
- Ikeda, T.; Asakura, S.; Kamiya, R. Total reconstitution of Salmonella flagellar filaments from hook and purified flagellin and hook-associated proteins in vitro. J. Mol. Biol. 1989, 209, 109–114. [Google Scholar] [CrossRef]
- Minamino, T. Hierarchical protein export mechanism of the bacterial flagellar type III protein export apparatus. FEMS Microbiol. Lett. 2018, 365, 117. [Google Scholar] [CrossRef]
- Evans, L.D.; Stafford, G.P.; Ahmed, S.; Fraser, G.M.; Hughes, C. An escort mechanism for cycling of export chaperones during flagellum assembly. Proc. Natl. Acad. Sci. USA 2006, 103, 17474–17479. [Google Scholar] [CrossRef] [Green Version]
- Minamino, T.; Kinoshita, M.; Hara, N.; Takeuchi, S.; Hida, A.; Koya, S.; Glenwright, H.; Imada, K.; Aldridge, P.D.; Namba, K. Interaction of a bacterial flagellar chaperone FlgN with FlhA is required for efficient export of its cognate substrates. Mol. Microbiol. 2012, 83, 775–788. [Google Scholar] [CrossRef] [PubMed]
- Kinoshita, M.; Hara, N.; Imada, K.; Namba, K.; Minamino, T. Interactions of bacterial flagellar chaperone-substrate complexes with FlhA contribute to co-ordinating assembly of the flagellar filament. Mol. Microbiol. 2013, 90, 1249–1261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bange, G.; Kümmerer, N.; Engel, C.; Bozkurt, G.; Wild, K.; Sinning, I. FlhA provides the adaptor for coordinated delivery of late flagella building blocks to the type III secretion system. Proc. Natl. Acad. Sci. USA 2010, 107, 11295–11300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomas, J.; Stafford, G.P.; Hughes, C. Docking of cytosolic chaperone-substrate complexes at the membrane ATPase during flagellar type III protein export. Proc. Natl. Acad. Sci. USA 2004, 101, 3945–3950. [Google Scholar] [CrossRef] [Green Version]
- Imada, K.; Minamino, T.; Kinoshita, M.; Furukawa, Y.; Namba, K. Structural insight into the regulatory mechanisms of interactions of the flagellar type III chaperone FliT with its binding partners. Proc. Natl. Acad. Sci. USA 2010, 107, 8812–8817. [Google Scholar] [CrossRef] [Green Version]
- Minamino, T.; Kinoshita, M.; Imada, K.; Namba, K. Interaction between FliI ATPase and a flagellar chaperone FliT during bacterial flagellar protein export. Mol. Microbiol. 2012, 83, 168–178. [Google Scholar] [CrossRef]
- Sajó, R.; Liliom, K.; Muskotál, A.; Klein, A.; Závodszky, P.; Vonderviszt, F.; Dobó, J. Soluble components of the flagellar export apparatus, FliI, FliJ, and FliH, do not deliver flagellin, the major filament protein, from the cytosol to the export gate. Biochim. Biophys. Acta 2014, 1843, 2414–2423. [Google Scholar] [CrossRef] [Green Version]
- Inoue, Y.; Morimoto, Y.V.; Namba, K.; Minamino, T. Novel insights into the mechanism of well-ordered assembly of bacterial flagellar proteins in Salmonella. Sci. Rep. 2018, 8, 1787. [Google Scholar] [CrossRef]
- Végh, B.M.; Gál, P.; Dobó, J.; Závodszky, P.; Vonderviszt, F. Localization of the flagellum-specific secretion signal in Salmonella flagellin. Biochem. Biophys. Res. Commun. 2006, 345, 93–98. [Google Scholar] [CrossRef]
- Singer, H.M.; Erhardt, M.; Hughes, K.T. Comparative analysis of the secretion capability of early and late flagellar type III secretion substrates. Mol. Microbiol. 2014, 93, 505–520. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain or Plasmid | Genotype or Description | Reference |
|---|---|---|
| E. coli strains | ||
| DH5α | F−, mcrA, Δ(mrr-hsdRMS-mcrBC), Φ80dlacZΔM15, ΔlacX74, deoR, recA1, araD139, Δ(ara leu)7697, galU, galK, λ−, rpsL, endA1,nupG | |
| BL21(DE3) | F− ompT hsdSB (rB− mB−) gal dcm (DE3) | Novagen |
| Salmonella strains | ||
| STH001 | ∆flhB ∆flgD ∆fliT | [29] |
| Plasmids | ||
| pBAD33SD | Cmr, pBAD33-based vector substituted NheI and EcoRI sites (GCTAGCGAATTC) into SD sequence (GCAGGAGGATTC) | [29] |
| pTrc99a | Ampr, Ptrc expression vector | |
| pTrc99aNde | pTrc99a-based vector substituted NcoI sites (cagACCATGgaa) into NdeI sequence (cagCATATGgaa) | This study |
| pET3c | Ampr, T7 expression vector | Novagen |
| pET15b | Ampr, T7 expression vector | Novagen |
| pITH103 | pBAD33SD-flhB + flhDC (FlhB, FlhD/FlhC) | [29] |
| pITH105 | pET15b-flgD (His-FlgD) | [29] |
| pITH106 | pET15b-flgE (His-FlgE) | [29] |
| pMMIJ001 | pET15b-fliJ (His-FliJ) | [35] |
| pMKM1702iH | pTrc99a-his-fliI + fliH (FliH/His-FliI), his-fliI was derived from the pET19b-based plasmid, pMM1901. | [36] |
| pITH107 | pET15b-fliK (His-FliK) | [29] |
| pITH108 | pET3c-fliS (FliS), the fliS gene amplified by PCR was cloned into the NdeI and BamHI sites of the pET3c vector | This study |
| pITH109 | pET15b-fliC (His-FliC), the fliC gene amplified by PCR was inserted into the NdeI and BamHI sites of pET15b | This study |
| pITH110 | pTrc99aNde-fliS + his-fliC (His-FliC/FliS), fliS derived from pITH108 and his-fliC derived from pITH109 were inserted into the NdeI and BamHI sites, and the XbaI and HindIII sites of pTrc99aNde, respectively | This study |
| pITH111 | pET3c-fliT (FliT), the fliT gene amplified by PCR was cloned into the NdeI and BamHI sites of pET3c | This study |
| pITH112 | pET15b-fliD (His-FliD), the fliD gene amplified by PCR was inserted into the NdeI and BamHI sites of pET15b | This study |
| pITH113 | pTrc99aNde-fliT + his-fliD (His-FliD/FliT), fliT derived from pITH111 and his-fliD derived from pITH112 were subcloned into the NdeI and BamHI sites and the XbaI and HindIII sites of pTrc99aNde, respectively | This study |
| pMMGN110 | pET22b-flgN (FlgN), The flgN gene amplified by PCR was cloned into the NdeI and BamHI sites of pET22b | [37] |
| pMMGN300 | pET19b-flgN (His-FlgN), flgN derived from pMMGN110 was subcloned into the NdeI and BamHI sites of pET19b | This study |
| pMMGK130 | pET15b-flgK (His-FlgK), the flgK gene amplified by PCR was inserted into the NdeI and BamHI sites of pET15b | [37] |
| pITH116 | pET15b-flgL (His-FlgL), the flgL amplified by PCR was inserted into the NdeI and BamHI sites of pET15b | This study |
| pITH117 | pTrc99aNde-flgN + his-flgK (His-FlgK/FlgN), flgN derived from pMMGN300 and his-flgK derived from pMMGK130 were subcloned into the NdeI and BamHI sites and the XbaI and HindIII sites of pTrc99aNde, respectively | This study |
| pITH118 | pTrc99aNde-flgN + his-flgL (His-FlgL/FlgN), flgN derived from pMMGN300 and his-flgL derived from pITH116 were subcloned into the NdeI and BamHI sites and XbaI and HindIII sites of pTrc99aNde, respectively | This study |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Terashima, H.; Tatsumi, C.; Kawamoto, A.; Namba, K.; Minamino, T.; Imada, K. In Vitro Autonomous Construction of the Flagellar Axial Structure in Inverted Membrane Vesicles. Biomolecules 2020, 10, 126. https://doi.org/10.3390/biom10010126
Terashima H, Tatsumi C, Kawamoto A, Namba K, Minamino T, Imada K. In Vitro Autonomous Construction of the Flagellar Axial Structure in Inverted Membrane Vesicles. Biomolecules. 2020; 10(1):126. https://doi.org/10.3390/biom10010126
Chicago/Turabian StyleTerashima, Hiroyuki, Chinatsu Tatsumi, Akihiro Kawamoto, Keiichi Namba, Tohru Minamino, and Katsumi Imada. 2020. "In Vitro Autonomous Construction of the Flagellar Axial Structure in Inverted Membrane Vesicles" Biomolecules 10, no. 1: 126. https://doi.org/10.3390/biom10010126