Automatic Quantification of Cardiomyocyte Dimensions and Connexin 43 Lateralization in Fluorescence Images

 ,
,  , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
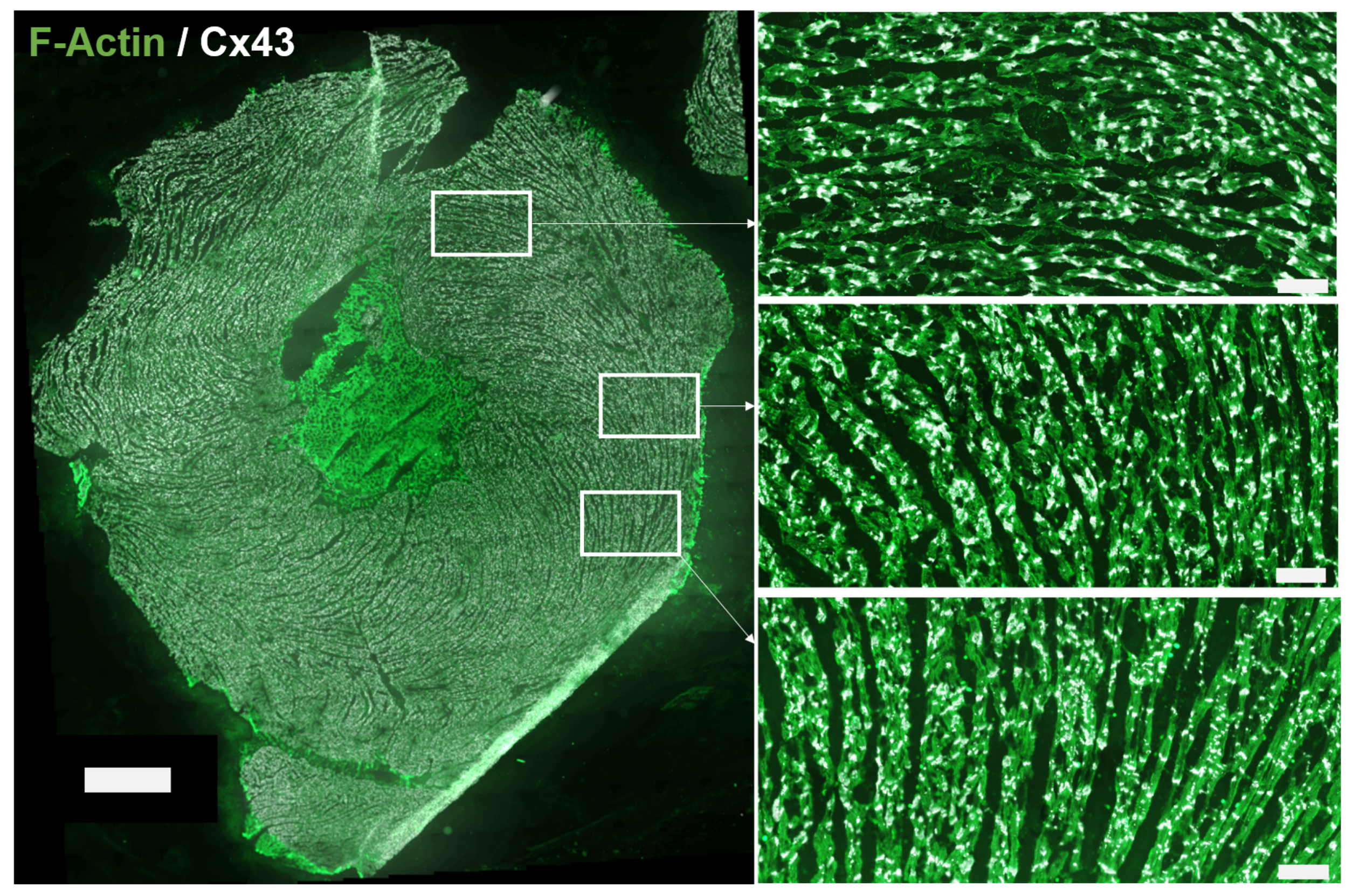
2.1. Sample Collection and Fluorescent Immunohistochemistry
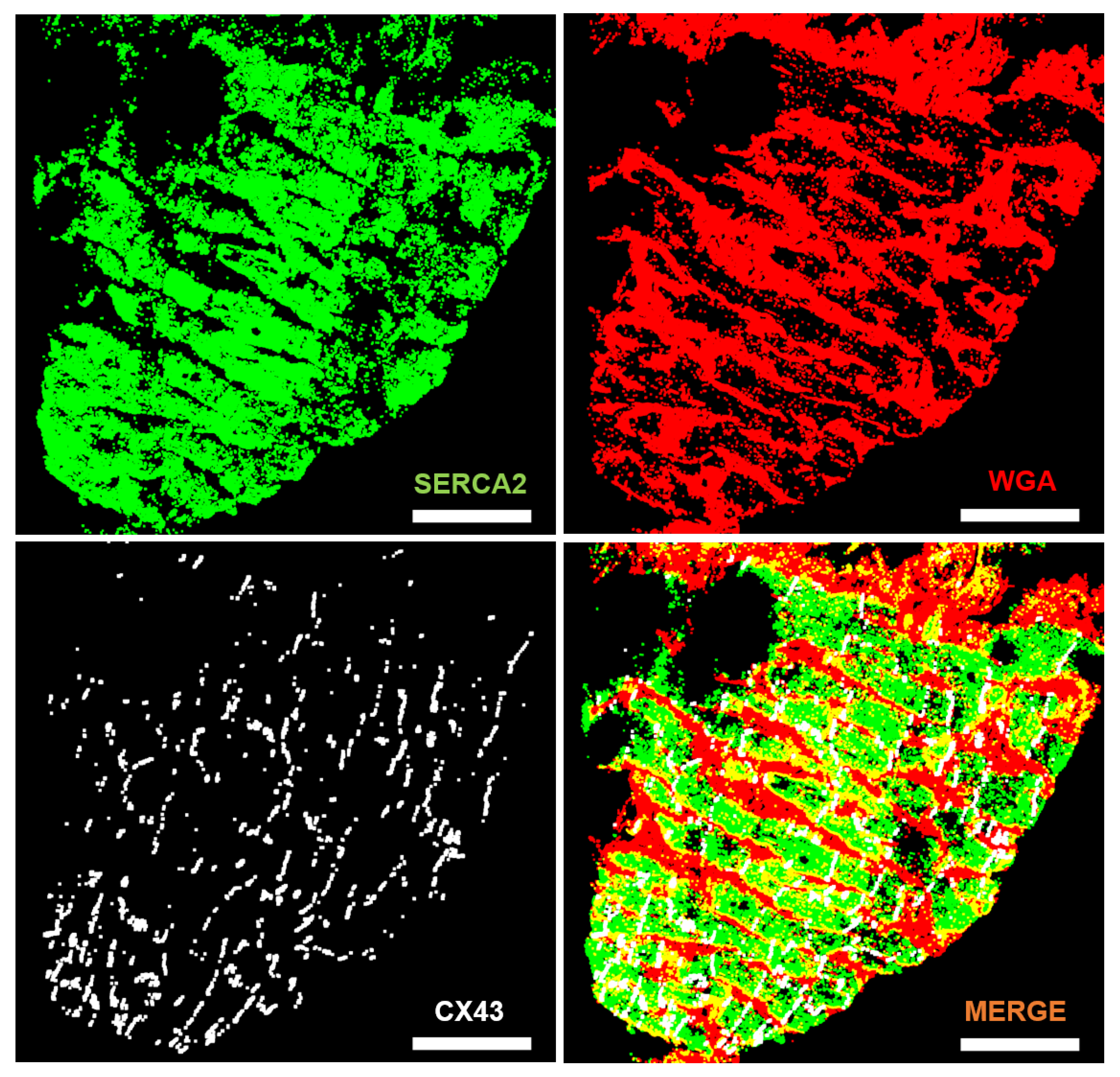
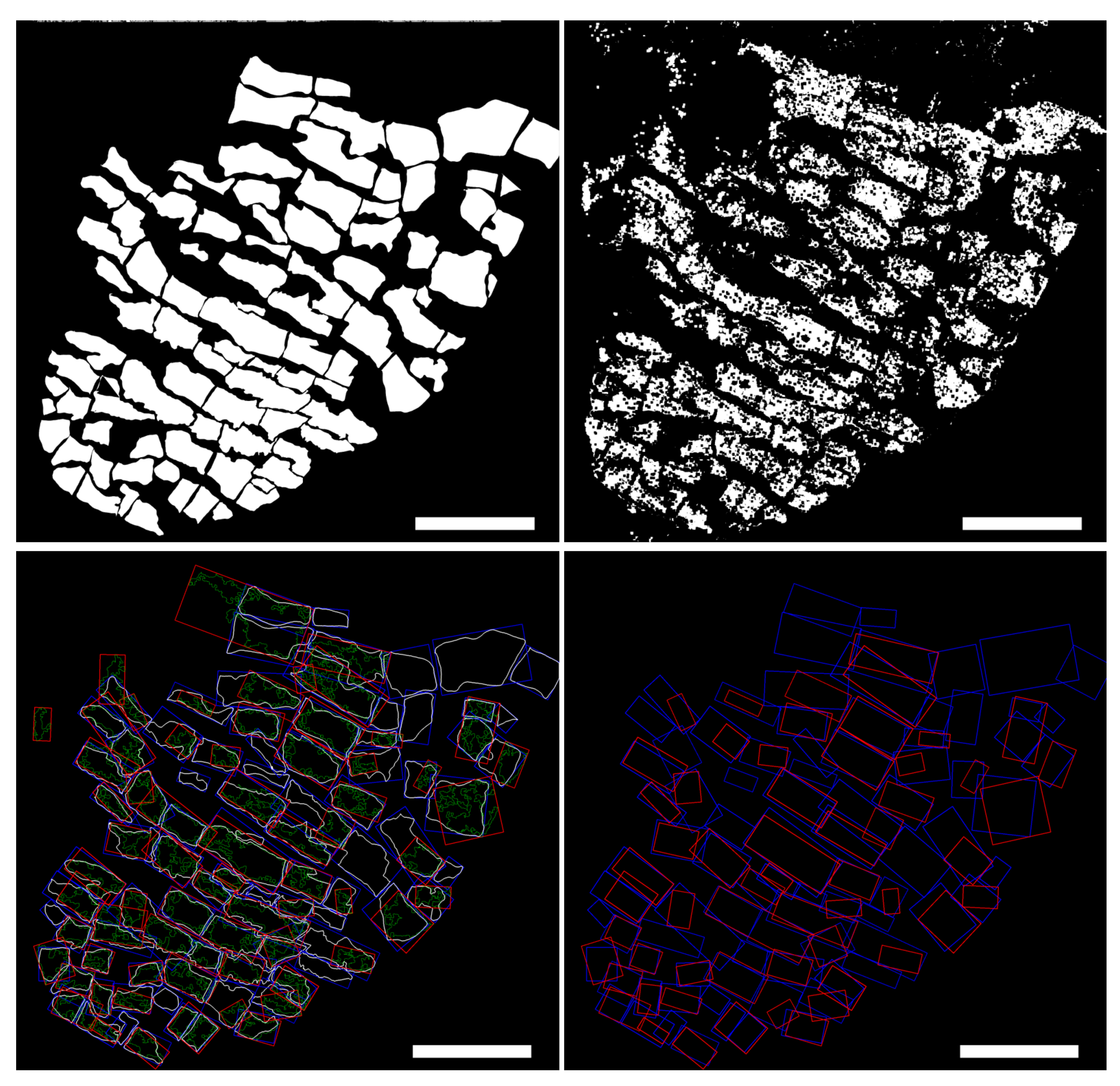
2.2. From Fluorescence Images to Cardiomyocyte Binary Masks
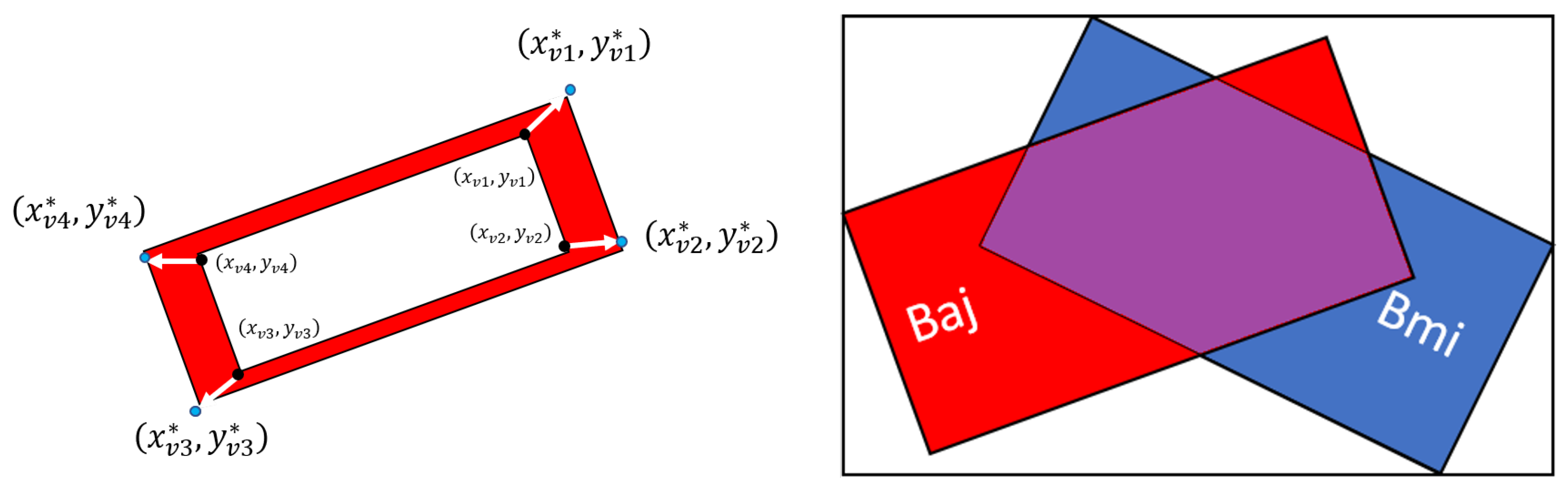
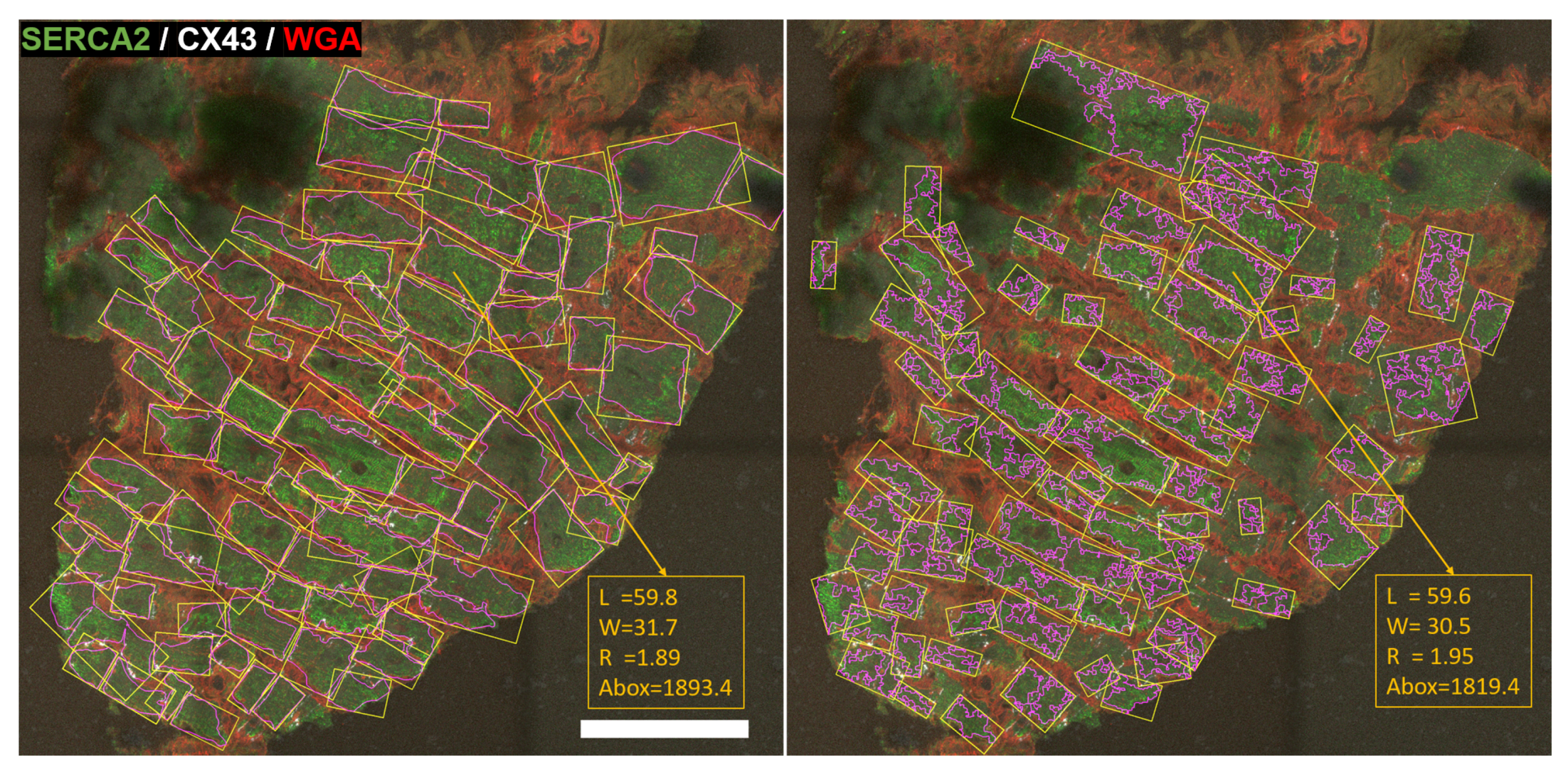
2.3. Cardiomyocyte Detection and Morphological Characterization
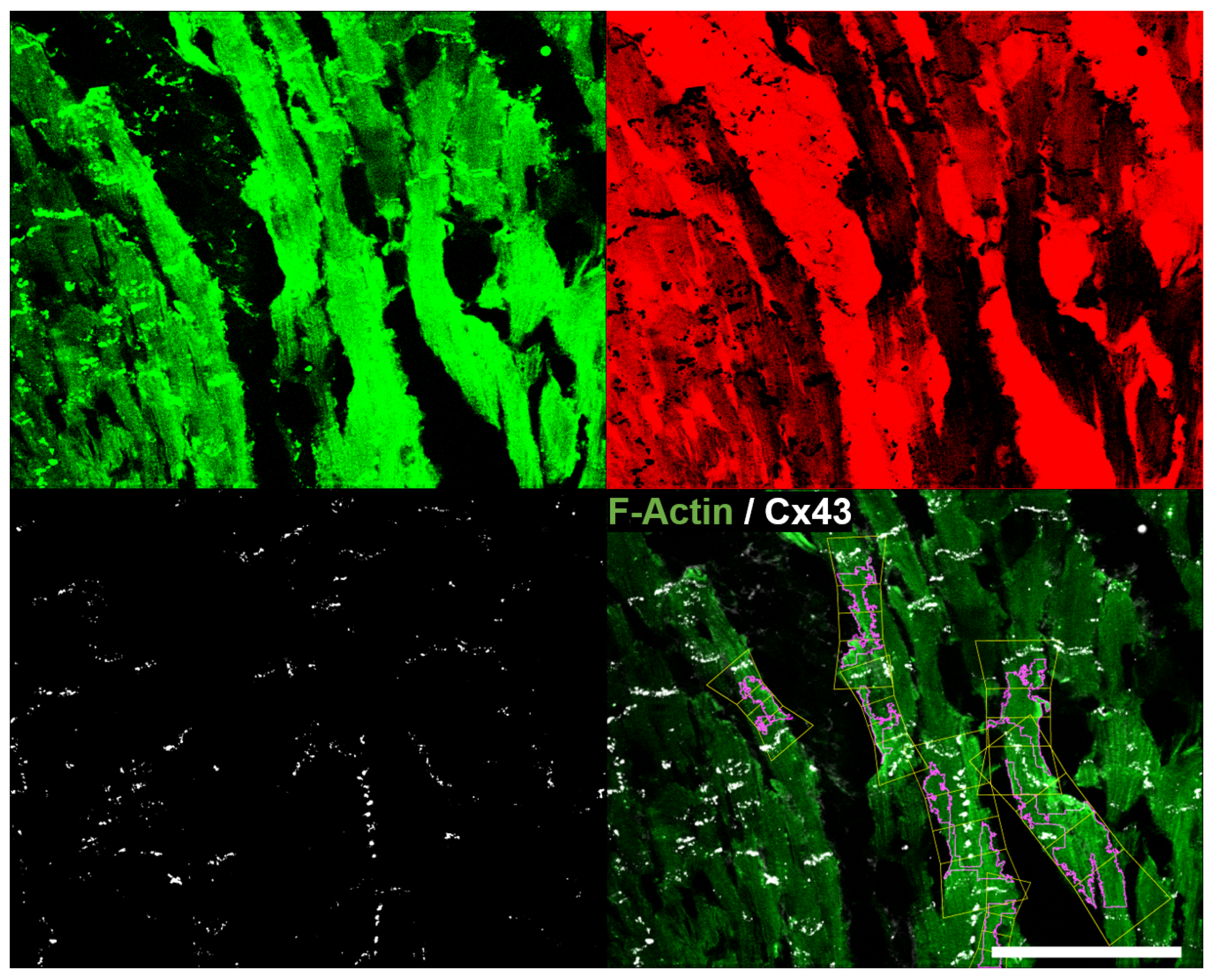
2.4. Quantification of CX43 Expression
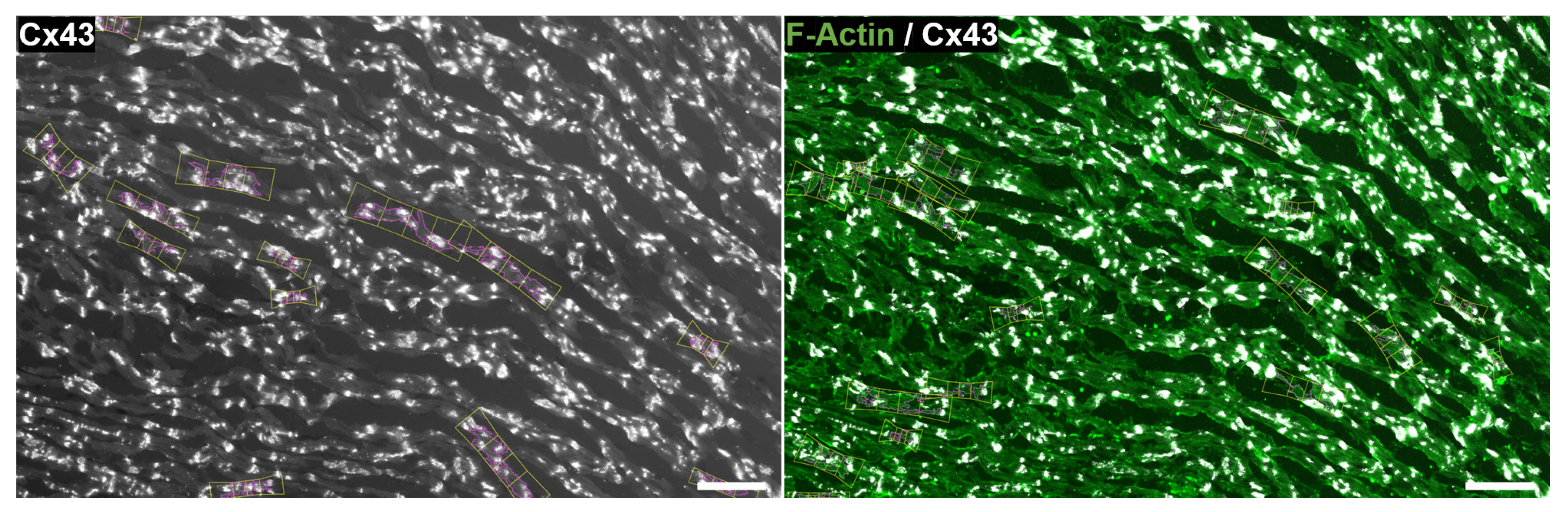
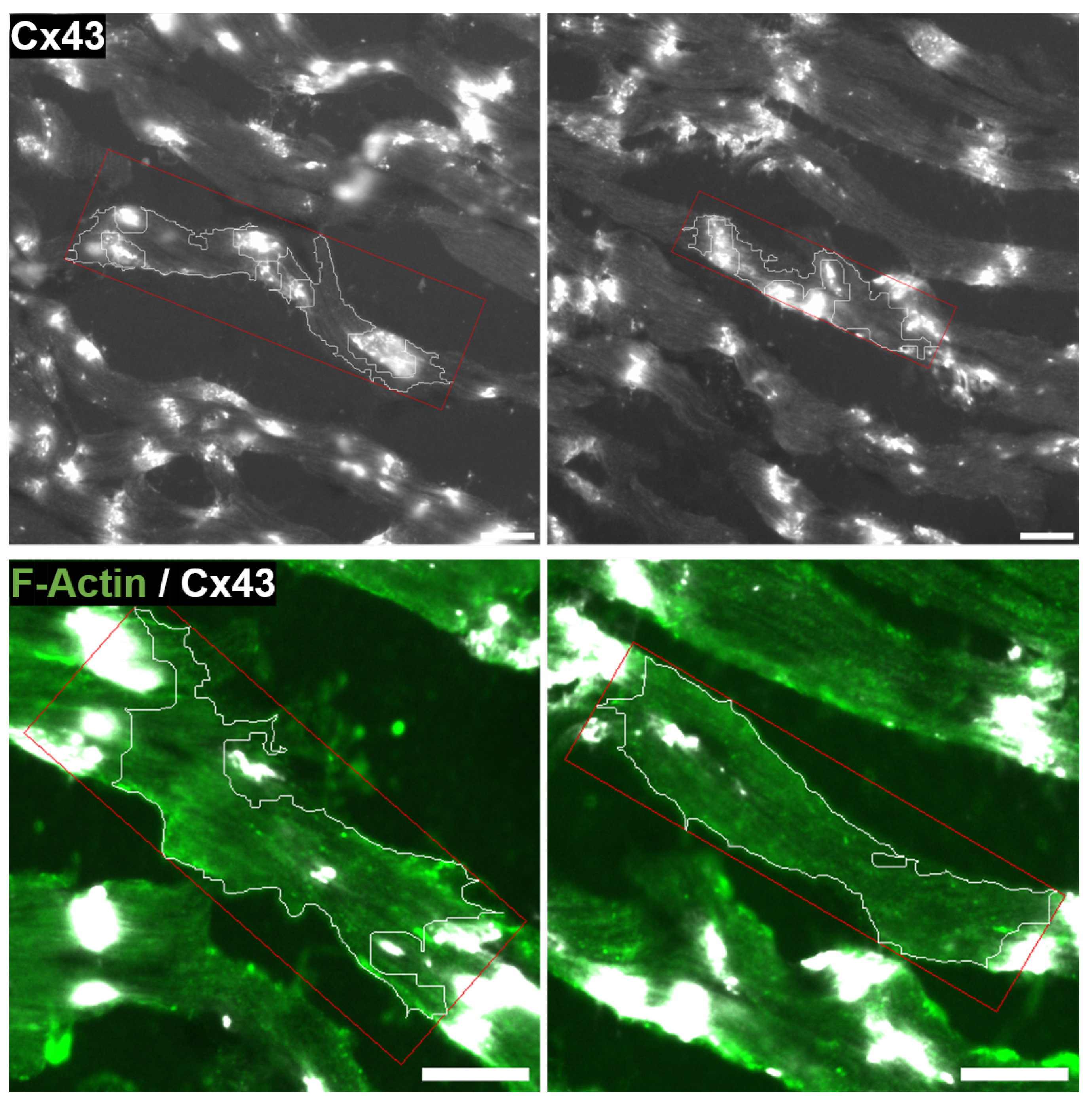
2.5. Quantification of CX43 Distribution
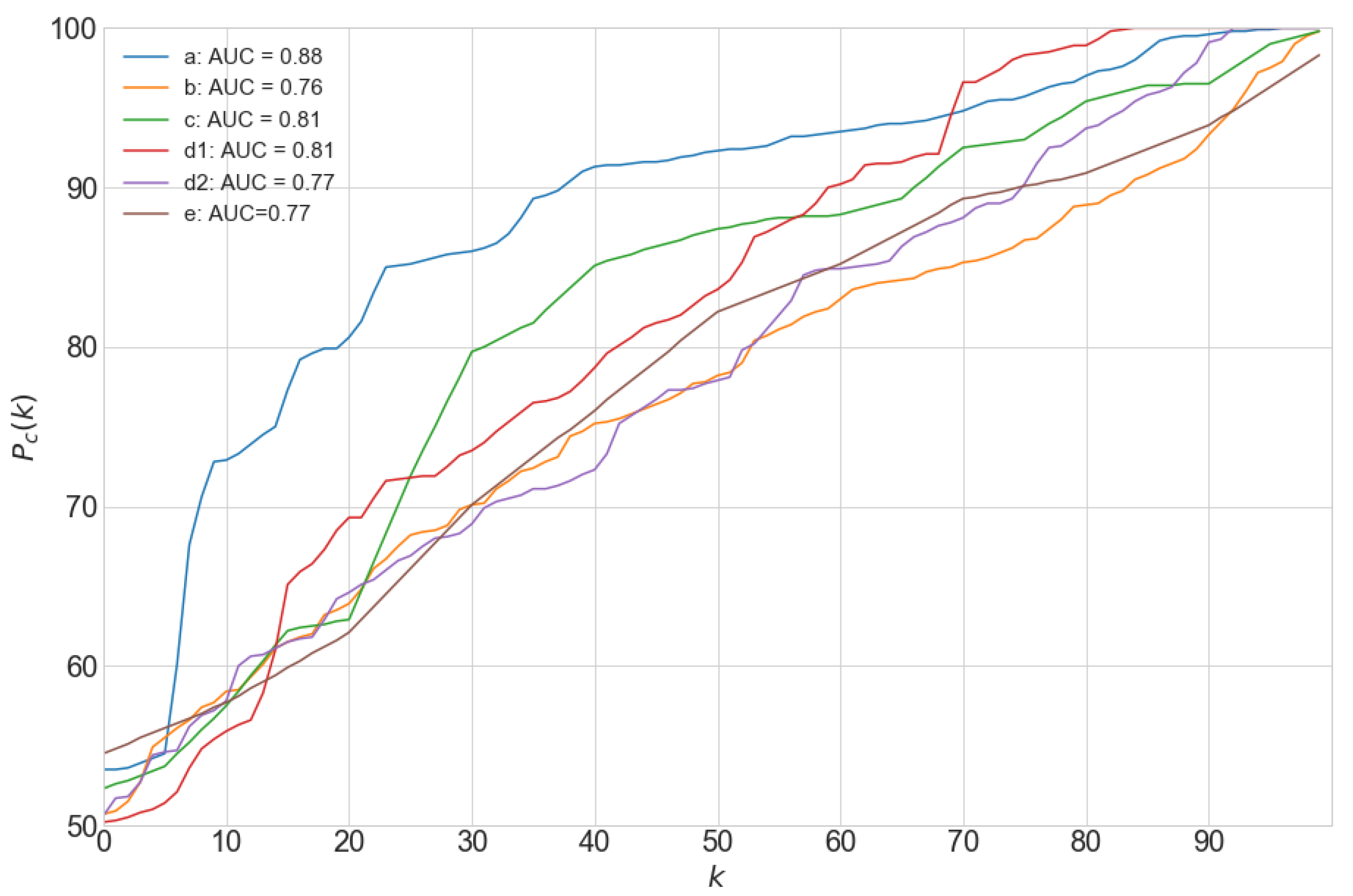
2.6. Performance Evaluation
3. Results
3.1. Automated Image Analysis
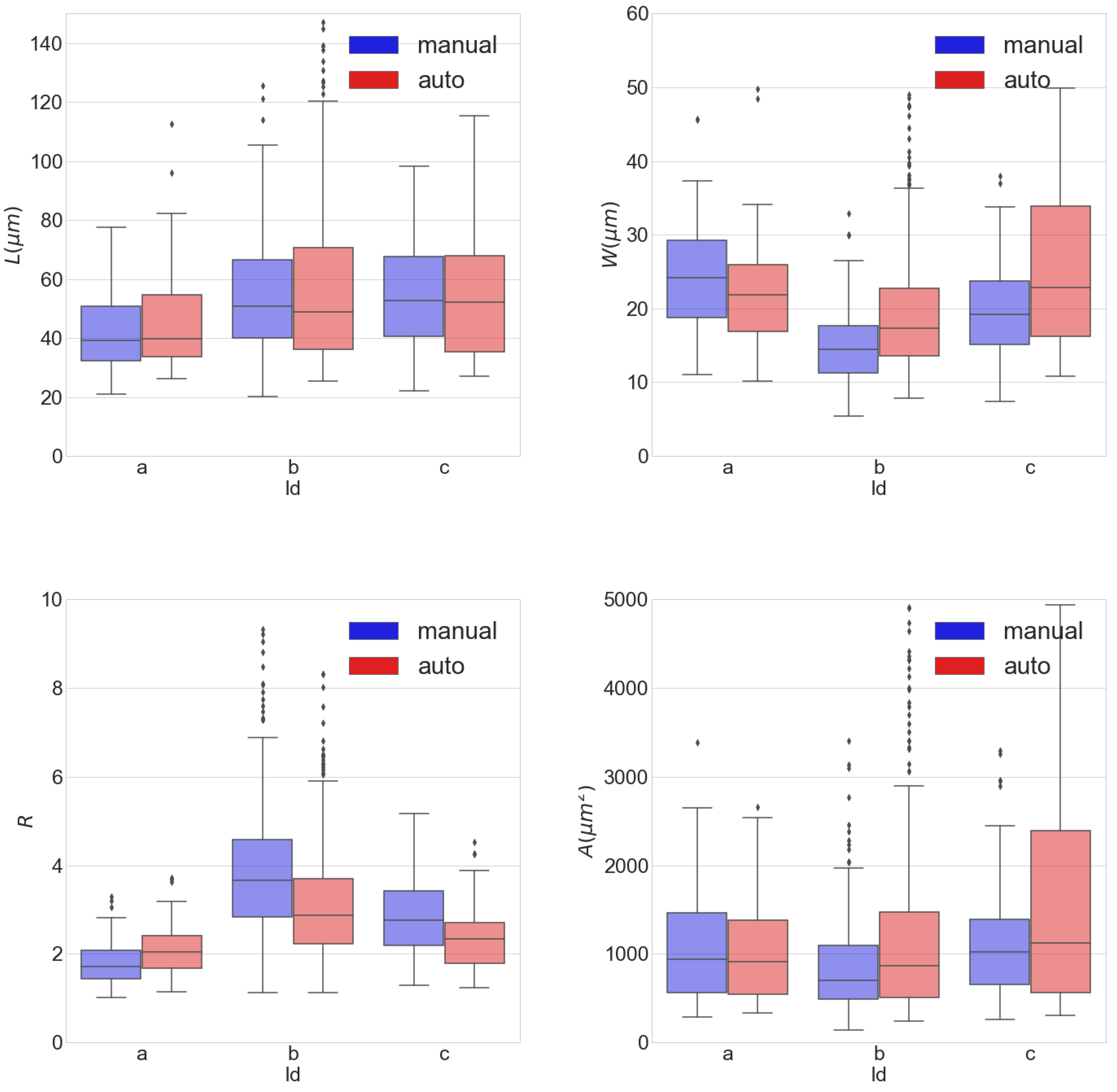
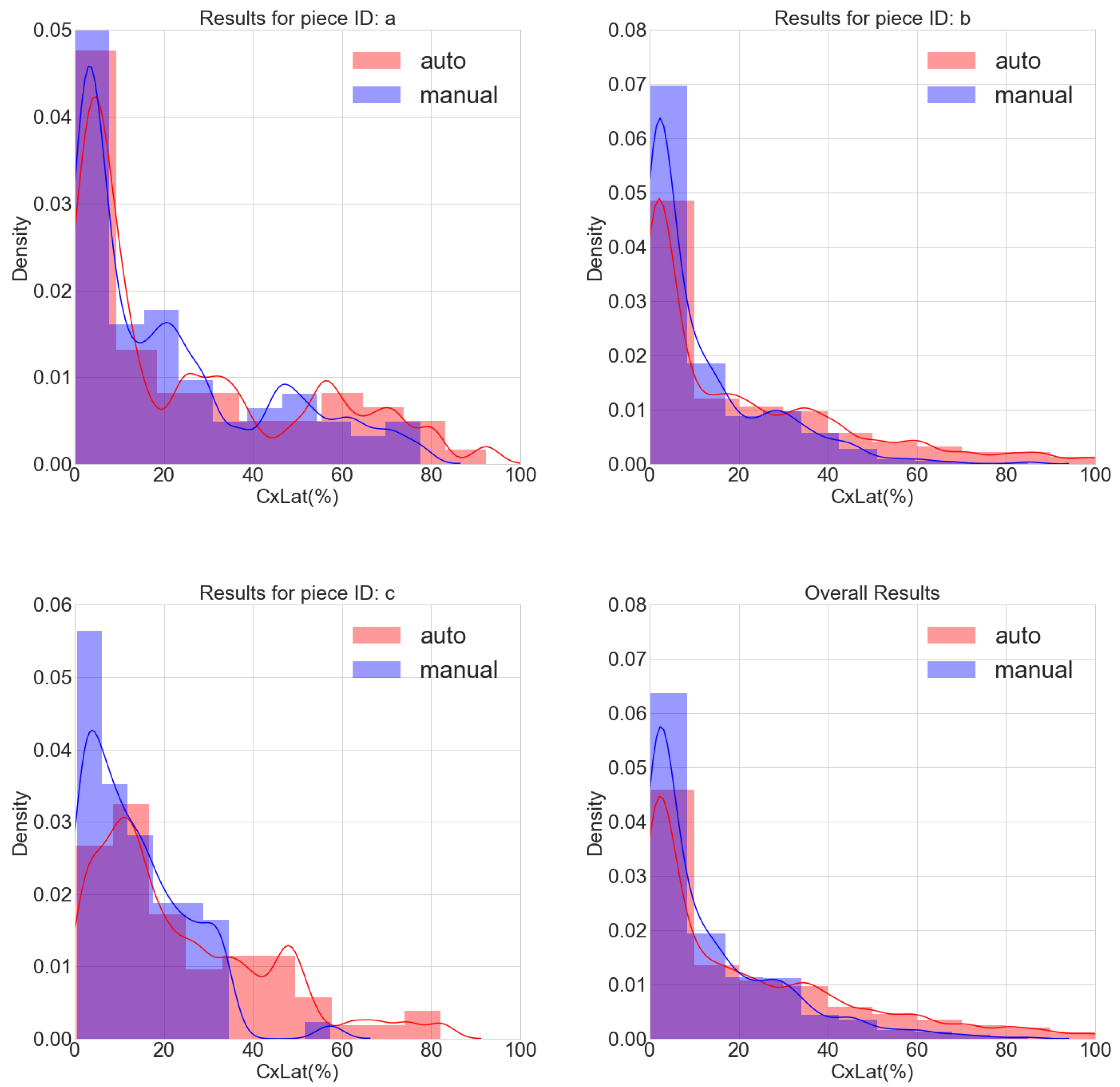
3.2. Agreement between Automatic and Manual Cell Delineation
3.3. Cardiomyocytes’ Morphological Measurements
3.4. Quantification of CX43 Expression
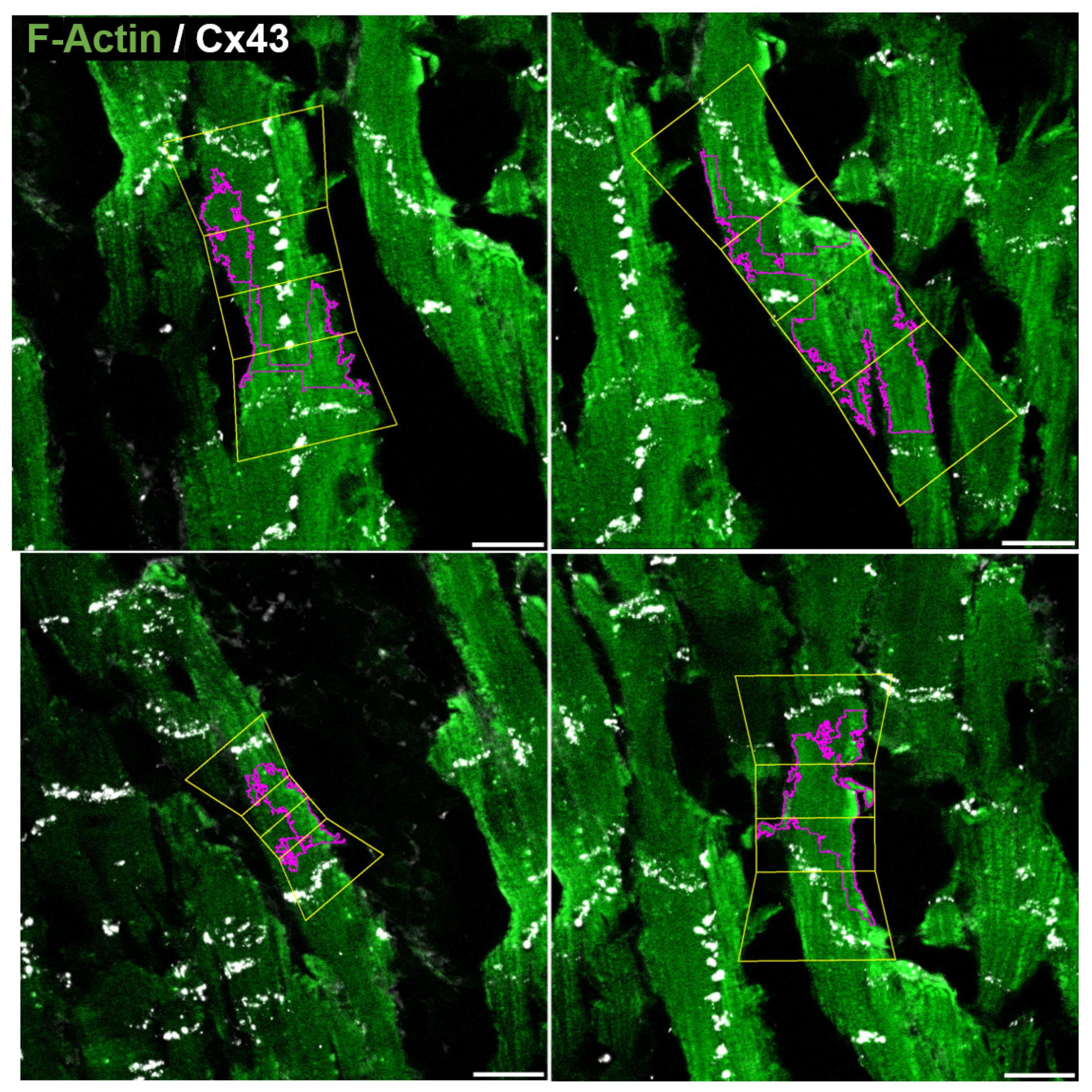
3.5. Determination of CX43 Distribution
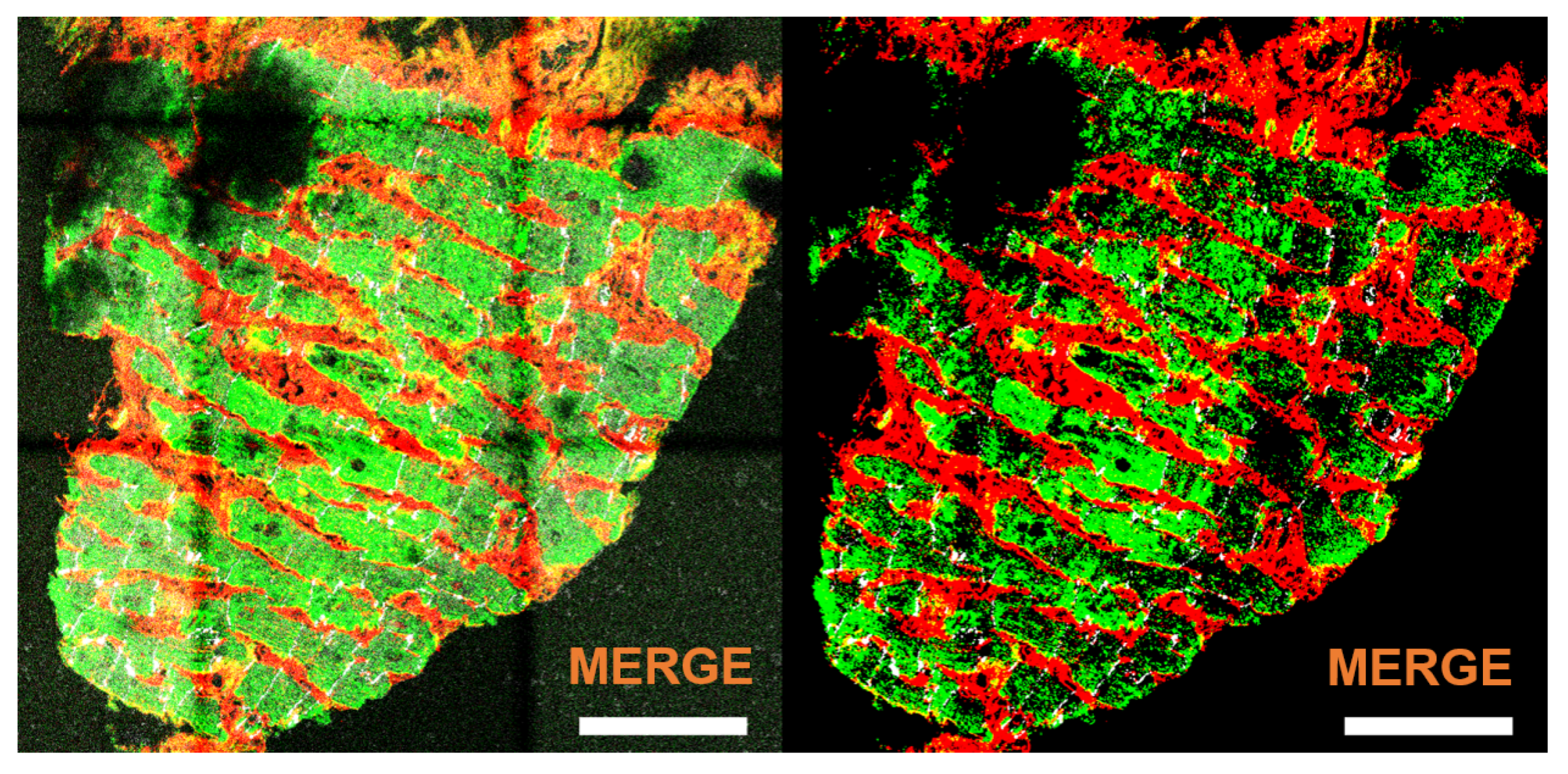
3.6. Application to Images with One or Two Channels
3.7. Processing Time
4. Discussion
4.1. Cardiomyocytes’ Morphological Measurements
4.2. CX43 Expression and Distribution
4.3. Additional Features of the Proposed Software
4.4. Study Limitations and Future Extensions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| CX43 | Connexin43 |
| CM | Cardiomyocite |
| WGA | Wheat Germ Agglutinin |
| LV | Left Ventricle |
| SL | Sarcomere Length |
| SERCA | Sarco/endoplasmic reticulum ATPase |
| DsRed | Red Fluorescent Protein |
| AUC | Area Under The Curve |
| FITC | Fluorescein isothiocyanate |
References
- Hubbard, M.L.; Ying, W.; Henriquez, C.S. Effect of gap junction distribution on impulse propagation in a monolayer of myocytes: A model study. Europace 2007, 9 (Suppl. S6), 20–28. [Google Scholar] [CrossRef] [PubMed]
- Stinstra, J.G.; Hopenfeld, B.; MacLeod, R.S. On the passive cardiac conductivity. Ann. Biomed. Eng. 2005, 33, 1743–1751. [Google Scholar] [CrossRef] [PubMed]
- Krzyzak, A.; Fevens, T.; Habibzadeh, M.; Jeleń, Ł. Application of pattern recognition techniques for the analysis of histopathological images. Adv. Intel. Soft Comput. 2011, 95, 623–644. [Google Scholar] [CrossRef]
- Spach, M.S.; Heidlage, J.F.; Dolber, P.C.; Barr, R.C. Electrophysiological effects of remodeling cardiac gap junctions and cell size. Experimental and model studies of normal cardiac growth. Circ. Res. 2000, 86, 302–311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dhein, S.; Seidel, T.; Salameh, A.; Jozwiak, J.; Hagen, A.; Kostelka, M.; Hindricks, G.; Mohr, F.W. Remodeling of cardiac passive electrical properties and susceptibility to ventricular and atrial arrhythmias. Front. Physiol. 2014, 5, 1–13. [Google Scholar] [CrossRef]
- Benova, T.E.; Bacova, B.S.; Viczenczova, C.; Diez, E.; Barancik, M.; Tribulova, N. Protection of Cardiac Cell-to-Cell Coupling Attenuate Myocardial Remodeling and Proarrhythmia Induced by Hypertension. Physiol. Res. 2016, 65, S29–S42. [Google Scholar] [CrossRef]
- Prado, N.J.; Egan Beňová, T.; Diez, E.R.; Knezl, V.; Lipták, B.; Ponce Zumino, A.Z.; Llamedo-Soria, M.; Szeiffová Bačová, B.; Miatello, R.M.; Tribulová, N. Melatonin receptor activation protects against low potassium-induced ventricular fibrillation by preserving action potentials and connexin-43 topology in isolated rat hearts. J. Pineal Res. 2019, 67. [Google Scholar] [CrossRef]
- Jansen, J.A.; Noorman, M.; Stein, M.; De Jong, S.; Van Der Nagel, R.; Hund, T.J.; Mohler, P.J.; Vos, M.A.; Van Veen, T.A.; De Bakker, J.M.; et al. Reduced heterogeneous expression of cx43 combined with decreased nav1.5 expression account for arrhythmia vulnerability in conditional cx43 knockout mice. Abnormal conduction in the diseased heart 2011, 88–104. [Google Scholar]
- Van Der Velden, H.M.W.; Jongsma, H.J. Cardiac gap junctions and connexins: Their role in atrial fibrillation and potential as therapeutic targets. Cardiovas. Res. 2002, 54, 270–279. [Google Scholar] [CrossRef]
- Boulaksil, M.; Bierhuizen, M.F.A.; Engelen, M.A.; Stein, M.; Kok, B.J.M.; van Amersfoorth, S.C.M.; Vos, M.A.; van Rijen, H.V.M.; de Bakker, J.M.T.; van Veen, T.A.B. Spatial Heterogeneity of Cx43 is an Arrhythmogenic Substrate of Polymorphic Ventricular Tachycardias during Compensated Cardiac Hypertrophy in Rats. Front. Cardiovasc. Med. 2016, 3, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Hesketh, G.G.; Shah, M.H.; Halperin, V.L.; Cooke, C.A.; Akar, F.G.; Yen, T.E.; Kass, D.A.; MacHamer, C.E.; Van Eyk, J.E.; Tomaselli, G.F. Ultrastructure and regulation of lateralized connexin43 in the failing heart. Circ. Res. 2010, 106, 1153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fontes, M.S.; Van Veen, T.A.; De Bakker, J.M.; Van Rijen, H.V. Functional consequences of abnormal Cx43 expression in the heart. Biochim. Biophys. Acta Biomembr. 2012, 1818, 2020–2029. [Google Scholar] [CrossRef] [Green Version]
- Seidel, T.; Salameh, A.; Dhein, S. A simulation study of cellular hypertrophy and connexin lateralization in cardiac tissue. Biophys. J. 2010, 99, 2821–2830. [Google Scholar] [CrossRef] [Green Version]
- Kohl, P.; Camelliti, P.; Burton, F.L.; Smith, G.L. Electrical coupling of fibroblasts and myocytes: Relevance for cardiac propagation. J. Electrocardiol. 2005, 38, 45–50. [Google Scholar] [CrossRef] [PubMed]
- Ongstad, E.; Kohl, P. Fibroblast–myocyte coupling in the heart: Potential relevance for therapeutic interventions. J. Mol. Cell. Cardiol. 2016, 91, 238–246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahoney, V.M.; Mezzano, V.; Mirams, G.R.; Maass, K.; Li, Z.; Cerrone, M.; Vasquez, C.; Bapat, A.; Delmar, M.; Morley, G.E. Connexin43 contributes to electrotonic conduction across scar tissue in the intact heart. Sci. Rep. 2016, 6. [Google Scholar] [CrossRef]
- Mannai, M.M.; Karâa, W.B.A. Biomedical image processing overview. Med. Imaging Concepts Methodol. Tools Appl. 2016, 59–71. [Google Scholar] [CrossRef]
- Vasuki, P.; Kanimozhi, J.; Devi, M.B. A survey on image preprocessing techniques for diverse fields of medical imagery. In Proceedings of the 2017 IEEE International Conference on Electrical, Instrumentation and Communication Engineering, ICEICE 2017, Karur, India, 27–28 April 2017; pp. 1–6. [Google Scholar] [CrossRef]
- Guirado, R.; Carceller, H.; Castillo-Gómez, E.; Castrén, E.; Nacher, J. Automated analysis of images for molecular quantification in immunohistochemistry. Heliyon 2018, 4. [Google Scholar] [CrossRef]
- Habibzadeh, M.; Krzyzak, A.; Fevens, T. White blood cell differential counts using convolutional neural networks for low resolution images. In International Conference on Artificial Intelligence and Soft Computing; Springer: Berlin/Heidelberg, Germany, 2013; pp. 263–274. [Google Scholar] [CrossRef]
- Hofgaard, J.P.; Mollerup, S.; Holstein-Rathlou, N.H.; Nielsen, M.S. Quantification of gap junctional intercellular communication based on digital image analysis. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2009, 297, 243–247. [Google Scholar] [CrossRef] [Green Version]
- Lackey, D.P.; Carruth, E.D.; Lasher, R.A.; Boenisch, J.; Sachse, F.B.; Hitchcock, R.W. Three-dimensional modeling and quantitative analysis of gap junction distributions in cardiac tissue. Ann. Biomed. Eng. 2011, 39, 2683. [Google Scholar] [CrossRef]
- Yan, J.; Thomson, J.K.; Wu, X.; Zhao, W.; Pollard, A.E.; Ai, X. Novel methods of automated quantification of gap junction distribution and interstitial collagen quantity from animal and human atrial tissue sections. PLoS ONE 2014, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suzuki, S.; Be, K.A. Topological structural analysis of digitized binary images by border following. Comput. Vis. Graph. Image Process. 1985, 30, 32–46. [Google Scholar] [CrossRef]
- Olivetti, G.; Cigola, E.; Maestri, R.; Corradi, D.; Lagrasta, C.; Gambert, S.R.; Anversa, P. Aging, cardiac hypertrophy and ischemic cardiomyopathy do not affect the proportion of mononucleated and multinucleated myocytes in the human heart. J. Mol. Cell. Cardiol. 1996, 28, 1463–1477. [Google Scholar] [CrossRef]
- Tracy, R.E.; Sander, G.E. Histologically measured cardiomyocyte hypertrophy correlates with body height as strongly as with body mass index. Cardiol. Res. Pract. 2011, 1. [Google Scholar] [CrossRef] [Green Version]
- Grajek, S.; Lesiak, M.; Pyda, M.; Zajac, M.; Paradowski, S.T.; Kaczmarek, E. Hypertrophy or hyperplasia in cardiac muscle. Post-mortem human morphometric study. Eur. Heart J. 1993, 14, 40–47. [Google Scholar] [CrossRef]
- Vliegen, H.W.; van der Laarse, A.; Huysman, J.A.; Wijnvoord, E.C.; Mentar, M.; Cornelisse, C.J.; Eulderink, F. Morphometric quantification of myocyte dimensions validated in normal growing rat hearts and applied to hypertrophic human hearts. Cardiovasc. Res. 1987, 21, 352–357. [Google Scholar] [CrossRef]
- Du, Y.; Plante, E.; Janicki, J.S.; Brower, G.L. Temporal evaluation of cardiac myocyte hypertrophy and hyperplasia in male rats secondary to chronic volume overload. Am. J. Pathol. 2010, 177, 1155–1163. [Google Scholar] [CrossRef]
- Bass, G.T.; Ryall, K.A.; Katikapalli, A.; Taylor, B.E.; Dang, S.T.; Acton, S.T.; Saucerman, J.J. Automated image analysis identifies signaling pathways regulating distinct signatures of cardiac myocyte hypertrophy. J. Mol. Cell. Cardiol. 2012, 52, 923–930. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mühlfeld, C.; Rajces, A.; Manninger, M.; Alogna, A.; Wierich, M.C.; Scherr, D.; Post, H.; Schipke, J. A transmural gradient of myocardial remodeling in early-stage heart failure with preserved ejection fraction in the pig. J. Anat. 2020, 236, 531–539. [Google Scholar] [CrossRef] [Green Version]
- Petersen, F.; Rodrigo, R.; Richter, M.; Kostin, S. The effects of polyunsaturated fatty acids and antioxidant vitamins on atrial oxidative stress, nitrotyrosine residues, and connexins following extracorporeal circulation in patients undergoing cardiac surgery. Mol. Cell. Biochem. 2017, 433, 27–40. [Google Scholar] [CrossRef]
- Aguet, F.; Van De Ville, D.; Unser, M. Model-based 2.5-D deconvolution for extended depth of field in brightfield microscopy. IEEE Trans. Image Process. 2008, 17.7, 1144–1153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Plataniotis, K.N. A Complete Color Normalization Approach to Histopathology Images Using Color Cues Computed From Saturation-Weighted Statistics. IEEE Trans. Biomed. Eng. 2015, 62, 1862–1873. [Google Scholar] [CrossRef]
- Zuiderveld, K. Contrast Limited Adaptive Histogram Equalization. Graph. Gems 1994, 474–485. [Google Scholar] [CrossRef]
- Otsu, N. Threshold selection method from gray-level histograms. IEEE Trans. Syst. Man Cybern. 1979, 9, 62–66. [Google Scholar] [CrossRef] [Green Version]
- Sauvola, J.; Pietikäinen, M. Adaptive document image binarization. Pattern Recognit. 2000, 33, 225–236. [Google Scholar] [CrossRef] [Green Version]
- Niblack, W. An Introduction to Digital Image Processing; Strandberg Publishing Company: Birkeroed, Denmark, 1986. [Google Scholar] [CrossRef]
- Saxena, L.P. Niblack’s binarization method and its modifications to real-time applications: A review. Artif. Intell. Rev. 2019, 51, 673–705. [Google Scholar] [CrossRef]
- Cirillo, D.; Valencia, A. Big data analytics for personalized medicine. Curr. Opin. Biotechnol. 2019, 58, 161–167. [Google Scholar] [CrossRef]

















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ID | a | b | c | e | ||
|---|---|---|---|---|---|---|
| ♯ inputs | 3 | 3 | 3 | 1 | 1 | 1 |
| ♯ channels | 3 | 3 | 3 | 1 | 2 | 2 |
| equalized (y/n) | n | n | n | y | y | y |
| supervised (y/n) | n | n | n | y | y | y |
| scale | 0.21 | 0.21 | 0.21 | 0.227 | 0.227 | 0.114 |
| 8 | 8 | 8 | 128 | 100 | 70 | |
| 15 | 15 | 15 | 254 | 254 | 254 | |
| 2 | 2 | 2 | 128 | 100 | 70 | |
| 254 | 254 | 254 | 254 | 254 | 254 | |
| 3 | 3 | 3 | 1 | 1 | 1 | |
| 3 | 3 | 3 | 3 | 3 | 2 | |
| 3 | 3 | 3 | 3 | 3 | 1 | |
| 3 | 3 | 3 | 1 | 1 | 1 | |
| 3 | 3 | 3 | 4 | 4 | 5 | |
| 5 | 5 | 5 | 8 | 8 | 20 | |
| 3 | 3 | 3 | 1 | 1 | 1 | |
| 3 | 3 | 3 | 3 | 3 | 2 | |
| 3 | 3 | 3 | 3 | 3 | 1 | |
| 66 | 453 | 64 | 111 (28) | 170 (44) | 15 (7) | |
| 84 | 371 | 82 | 344 | 344 | 46 |
| (%) | (%) | (%) | (%) | (%) | (%) | |||
|---|---|---|---|---|---|---|---|---|
| 252 | 1.41 | 0.111 | 1.53 | 0.063 | 2.63 | 0.353 | 0.54 | 0.58 |
| 253 | 0.82 | 0.039 | 0.97 | 0.031 | 1.60 | 0.169 | 0.51 | 0.61 |
| 254 | 0.27 | 0.007 | 0.31 | 0.007 | 0.58 | 0.039 | 0.47 | 0.53 |
| CM | L (m) | W (m) | R | A (m) | (%) |
|---|---|---|---|---|---|
| 166.9 | 44.5 | 3.7 | 7428.6 | 48.8 | |
| 120.3 | 25.2 | 4.8 | 3031.7 | 60.2 | |
| 107.7 | 35.1 | 3.1 | 3777.3 | 5.4 | |
| 108.2 | 25.2 | 4.3 | 2730.6 | 0.0 | |
| 81.6 | 35.6 | 2.3 | 2903.8 | 44.1 | |
| 111.1 | 33.3 | 3.3 | 3695.3 | 41.6 | |
| 42.7 | 17.9 | 2.4 | 762.9 | 0.0 | |
| 71.1 | 33.8 | 2.1 | 2399.0 | 11.9 |
| ID | (s) | Ratio | (s) | S (Mb) | |||
|---|---|---|---|---|---|---|---|
| a | 0.11 | 1.0 | 66.0 | 600 | 22 | 66 | 52.0 |
| b | 0.11 | 15.4 | 29.4 | 267 | 1036 | 453 | 363.0 |
| c | 0.11 | 3.4 | 18.8 | 171 | 54 | 64 | 92.5 |
| d1 | 0.11 | 4.6 | 24.1 (6.1) | 219 (55) | 201 (522) | 111 (28) | 18.3 |
| d2 | 0.11 | 4.6 | 36.9 (9.6) | 335 (87) | 300 (683) | 170 (44) | 43.5 |
| e | 0.11 | 2.1 | 7.1 (3.3) | 64 (30) | 26 (71) | 15 (7) | 18.7 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oliver-Gelabert, A.; García-Mendívil, L.; Vallejo-Gil, J.M.; Fresneda-Roldán, P.C.; Andelová, K.; Fañanás-Mastral, J.; Vázquez-Sancho, M.; Matamala-Adell, M.; Sorribas-Berjón, F.; Ballester-Cuenca, C.; et al. Automatic Quantification of Cardiomyocyte Dimensions and Connexin 43 Lateralization in Fluorescence Images. Biomolecules 2020, 10, 1334. https://doi.org/10.3390/biom10091334
Oliver-Gelabert A, García-Mendívil L, Vallejo-Gil JM, Fresneda-Roldán PC, Andelová K, Fañanás-Mastral J, Vázquez-Sancho M, Matamala-Adell M, Sorribas-Berjón F, Ballester-Cuenca C, et al. Automatic Quantification of Cardiomyocyte Dimensions and Connexin 43 Lateralization in Fluorescence Images. Biomolecules. 2020; 10(9):1334. https://doi.org/10.3390/biom10091334
Chicago/Turabian StyleOliver-Gelabert, Antoni, Laura García-Mendívil, José María Vallejo-Gil, Pedro Carlos Fresneda-Roldán, Katarína Andelová, Javier Fañanás-Mastral, Manuel Vázquez-Sancho, Marta Matamala-Adell, Fernando Sorribas-Berjón, Carlos Ballester-Cuenca, and et al. 2020. "Automatic Quantification of Cardiomyocyte Dimensions and Connexin 43 Lateralization in Fluorescence Images" Biomolecules 10, no. 9: 1334. https://doi.org/10.3390/biom10091334