
Pancreatic Progenitor Commitment Is Marked by an Increase in Ink4a/Arf Expression

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
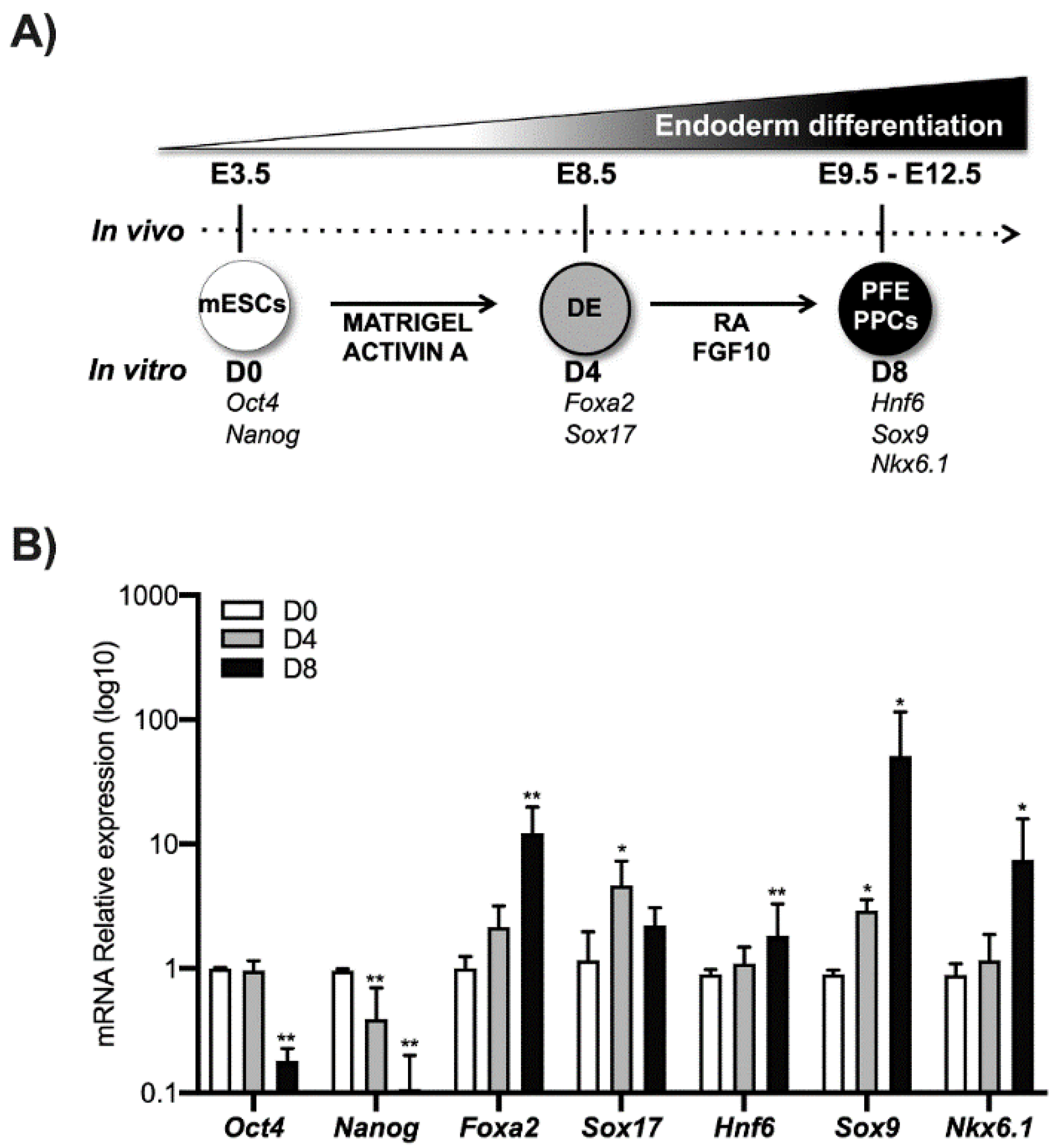
2.1. ESCs Culture and Differentiation
2.2. qPCR Analysis
2.3. Western Blot Analysis
2.4. Immunofluorescence
2.5. Statistics
3. Results
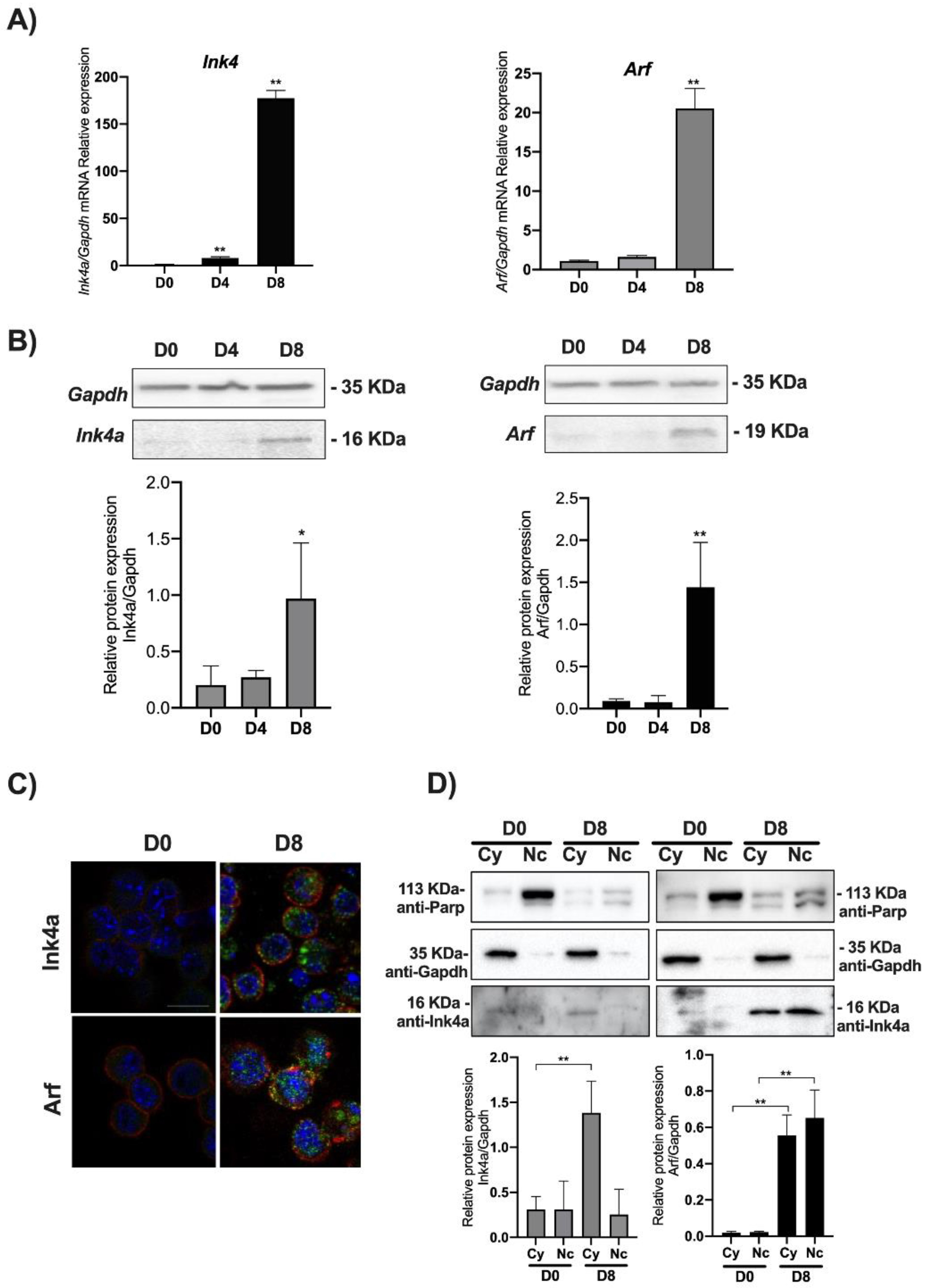
3.1. Ink4a/Arf Gene Expression Increases in mESCs Induced to Differentiate towards Pancreatic Lineage
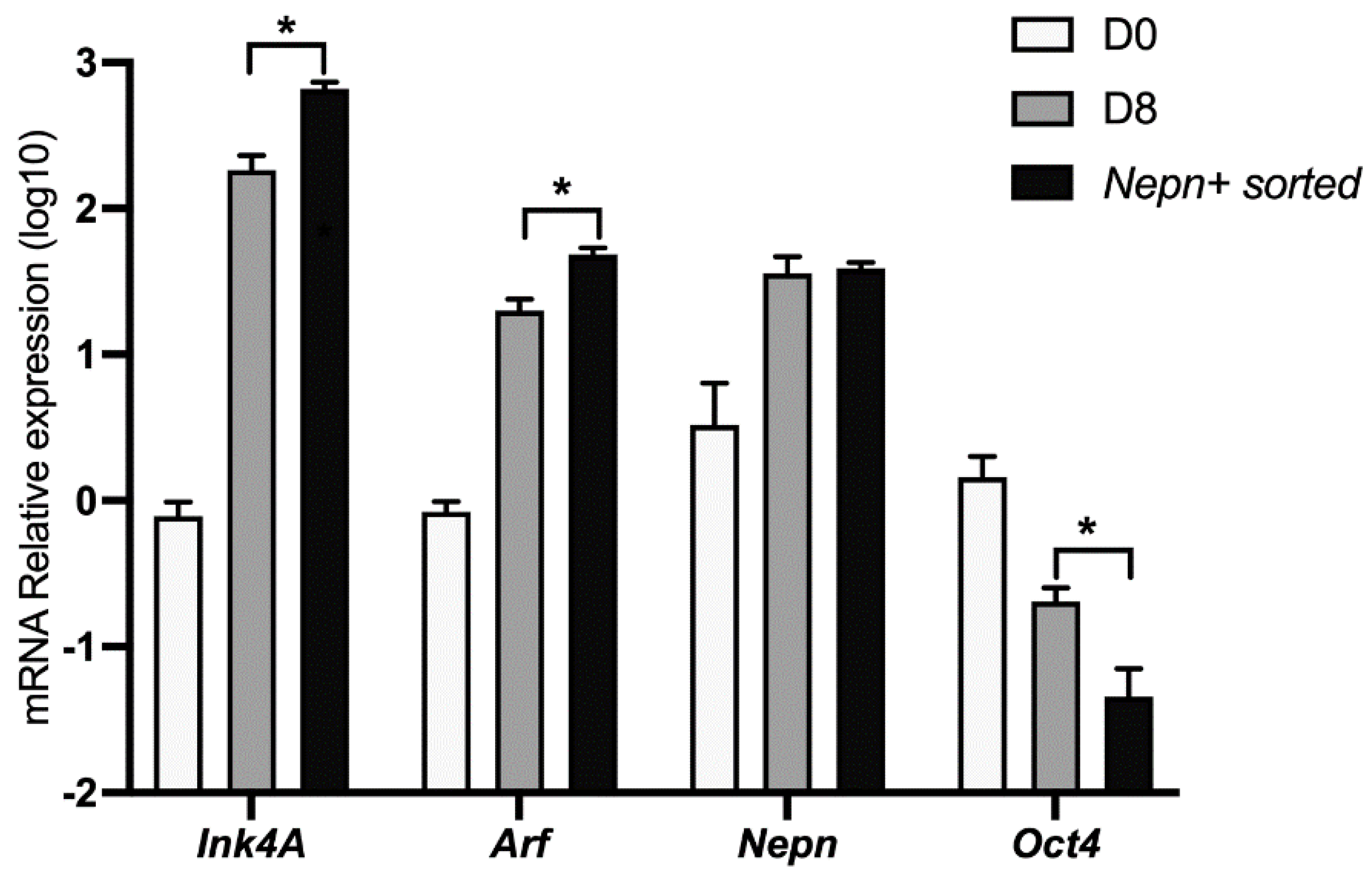
3.2. Ink4a/Arf Gene Expression Correlates with Endoderm-Nepn Cells
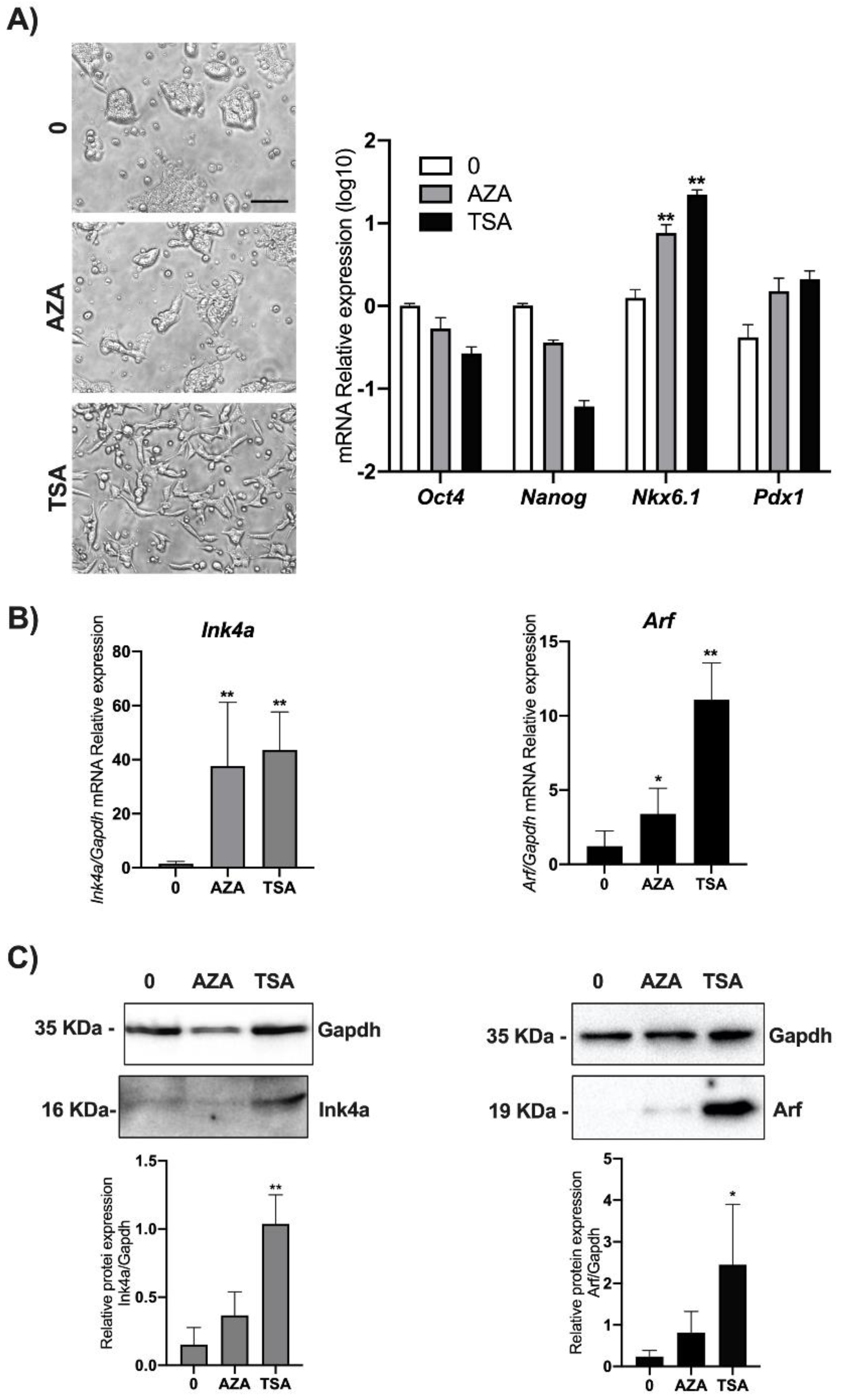
3.3. DNA Methylation and Chromatin Acetylation Affect Ink4a and Arf Gene Expression in ESCs
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Tanase, C.P.; Neagu, A.I.; Necula, L.G.; Mambet, C.; Enciu, A.-M.; Calenic, B.; Cruceru, M.L.; Albulescu, R. Cancer stem cells: Involvement in pancreatic cancer pathogenesis and perspectives on cancer therapeutics. World J. Gastroenterol. 2014, 20, 10790–10801. [Google Scholar] [CrossRef]
- Wang, S.; Huang, S.; Sun, Y.L. Epithelial-Mesenchymal Transition in Pancreatic Cancer: A Review. BioMed Res. Int. 2017, 2017, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Sperka, T.; Wang, J.; Rudolph, K.L. DNA damage checkpoints in stem cells, ageing and cancer. Nat. Rev. Mol. Cell Biol. 2012, 13, 579–590. [Google Scholar] [CrossRef] [PubMed]
- Fontana, R.; Ranieri, M.; La Mantia, G.; Vivo, M. Dual Role of the Alternative Reading Frame ARF Protein in Cancer. Biomolecules 2019, 9, 87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, H.; Gu, X.; Su, I.-H.; Bottino, R.; Contreras, J.L.; Tarakhovsky, A.; Kim, S.K. Polycomb protein Ezh2 regulates pancreatic β-cell Ink4a/Arf expression and regeneration in diabetes mellitus. Genes Dev. 2009, 23, 975–985. [Google Scholar] [CrossRef] [Green Version]
- Sherr, C.J. Ink4-Arf locus in cancer and aging. Wiley Interdiscip. Rev. Dev. Biol. 2012, 1, 731–741. [Google Scholar] [CrossRef] [Green Version]
- Lagopati, N.; Belogiannis, K.; Angelopoulou, A.; Papaspyropoulos, A.; Gorgoulis, V. Non-Canonical Functions of the ARF Tumor Suppressor in Development and Tumorigenesis. Biomolecules 2021, 11, 86. [Google Scholar] [CrossRef]
- Ming, Z.; Lim, S.Y.; Rizos, H. Genetic Alterations in the INK4a/ARF Locus: Effects on Melanoma Development and Progression. Biomolecules 2020, 10, 1447. [Google Scholar] [CrossRef]
- Pal, A.; Potjer, T.P.; Thomsen, S.K.; Ng, H.J.; Barrett, A.; Scharfmann, R.; James, T.J.; Bishop, T.; Karpe, F.; Godsland, I.F.; et al. Loss-of-Function Mutations in the Cell-Cycle Control GeneCDKN2AImpact on Glucose Homeostasis in Humans. Diabetes 2015, 65, 527–533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lal, G.; Liu, L.; Hogg, D.; Lassam, N.J.; Redston, M.S.; Gallinger, S. Patients with both pancreatic adenocarcinoma and melanoma may harbor germline CDKN2A mutations. Genes Chromosomes Cancer 2000, 27, 358–361. [Google Scholar] [CrossRef]
- Gerdes, B.; Bartsch, D.K.; Ramaswamy, A.; Kersting, M.; Wild, A.; Schuermann, M.; Frey, M.; Rothmund, M. Multiple Primary Tumors as an Indicator for p16INK4a Germline Mutations in Pancreatic Cancer Patients? Pancreas 2000, 21, 369–375. [Google Scholar] [CrossRef]
- Bartsch, D.K.; Sina-Frey, M.; Lang, S.; Wild, A.; Gerdes, B.; Barth, P.; Kress, R.; Grützmann, R.; Colombo-Benkmann, M.; Ziegler, A.; et al. CDKN2A Germline Mutations in Familial Pancreatic Cancer. Ann. Surg. 2002, 236, 730–737. [Google Scholar] [CrossRef] [PubMed]
- Bao, X.Y.; Xie, C.; Yang, M.S. Association between Type 2 Diabetes and CDKN2A/B: A meta-analysis study. Mol. Biol. Rep. 2011, 39, 1609–1616. [Google Scholar] [CrossRef]
- Addeo, M.; Buonaiuto, S.; Guerriero, I.; Amendola, E.; Visconte, F.; Marino, A.; De Angelis, M.T.; Russo, F.; Roberto, L.; Marotta, P.; et al. Insight into Nephrocan Function in Mouse Endoderm Patterning. Int. J. Mol. Sci. 2019, 21, 8. [Google Scholar] [CrossRef] [Green Version]
- De Angelis, M.T.; Russo, F.; D’Angelo, F.; Federico, A.; Gemei, M.; Del Vecchio, L.; Ceccarelli, M.; De Felice, M.; Falco, G. Novel Pancreas Organogenesis Markers Refine the Pancreatic Differentiation Roadmap of Embryonic Stem cells. Stem Cell Rev. Rep. 2014, 10, 269–279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stryke, D.; Kawamoto, M.; Huang, C.C.; Johns, S.J.; King, L.A.; Harper, C.A.; Meng, E.C.; Lee, R.E.; Yee, A.; L’Italien, L.; et al. BayGenomics: A resource of insertional mutations in mouse embryonic stem cells. Nucleic Acids Res. 2003, 31, 278–281. [Google Scholar] [CrossRef] [PubMed]
- Poot, M.; Arttamangkul, S. Verapamil inhibition of enzymatic product efflux leads to improved detection of beta-galactosidase activity in lacZ-transfected cells. Cytometry 1997, 28, 36–41. [Google Scholar] [CrossRef]
- Vivo, M.; Fontana, R.; Ranieri, M.; Capasso, G.; Angrisano, T.; Pollice, A.; Calabrò, V.; La Mantia, G. p14ARF interacts with the focal adhesion kinase and protects cells from anoikis. Oncogene 2017, 36, 4913–4928. [Google Scholar] [CrossRef] [Green Version]
- Bracken, A.P.; Kleine-Kohlbrecher, D.; Dietrich, N.; Pasini, D.; Gargiulo, G.; Beekman, C.; Theilgaard-Mönch, K.; Minucci, S.; Porse, B.; Marine, J.-C.; et al. The Polycomb group proteins bind throughout the INK4A-ARF locus and are disassociated in senescent cells. Genes Dev. 2007, 21, 525–530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.; Collado, M.; Villasante, A.; Strati, K.; Ortega, S.; Cañamero, M.; Blasco, M.A.; Serrano, M. The Ink4/Arf locus is a barrier for iPS cell reprogramming. Nat. Cell Biol. 2009, 460, 1136–1139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farooq, U.; Saravanan, B.; Islam, Z.; Walavalkar, K.; Singh, A.K.; Jayani, R.S.; Meel, S.; Swaminathan, S.; Notani, D. An interdependent network of functional enhancers regulates transcription and EZH2 loading at the INK4a/ARF locus. Cell Rep. 2021, 34, 108898. [Google Scholar] [CrossRef]
- Wang, W.; Zhu, Y.; Huang, K.; Shan, Y.; Du, J.; Dong, X.; Ma, P.; Wu, P.; Zhang, J.; Huang, W.; et al. Suppressing p16 Ink4a and P14 ARF pathways overcomes apoptosis in individualized human embryonic stem cells. FASEB J. 2016, 31, 1130–1140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hattori, N.; Nishino, K.; Ko, Y.-G.; Hattori, N.; Ohgane, J.; Tanaka, S.; Shiota, K. Epigenetic Control of Mouse Oct-4 Gene Expression in Embryonic Stem Cells and Trophoblast Stem Cells. J. Biol. Chem. 2004, 279, 17063–17069. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huan, Y.; Wang, H.; Wu, Z.; Zhang, J.; Zhu, J.; Liu, Z.; He, H. Epigenetic Modification of Cloned Embryos ImprovesNanogReprogramming in Pigs. Cell. Reprogram. 2015, 17, 191–198. [Google Scholar] [CrossRef]
- Sambathkumar, R.; Kalo, E.; Van Rossom, R.; Faas, M.M.; de Vos, P.; Verfaillie, C.M. Epigenetic Induction of Definitive and Pancreatic Endoderm Cell Fate in Human Fibroblasts. Stem Cells Int. 2016, 2016, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liao, X.; Xue, H.; Wang, Y.-C.; Nazor, K.L.; Guo, S.; Trivedi, N.; Peterson, S.E.; Liu, Y.; Loring, J.F.; Laurent, L.C. Matched miRNA and mRNA signatures from a hESC-based in vitro model of pancreatic differentiation reveal novel regulatory interactions. J. Cell Sci. 2013, 126, 3848–3861. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gromley, A.; Churchman, M.L.; Zindy, F.; Sherr, C.J. Transient expression of the Arf tumor suppressor during male germ cell and eye development in Arf-Cre reporter mice. Proc. Natl. Acad. Sci. USA 2009, 106, 6285–6290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vivo, M.; Di Costanzo, A.; Fortugno, P.; Pollice, A.; Calabrò, V.; La Mantia, G. Downregulation of ΔNp63α in keratinocytes by p14ARF-mediated SUMO-conjugation and degradation. Cell Cycle 2009, 8, 3545–3551. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Finkelstein, D.; Sherr, C.J. Arf tumor suppressor and miR-205 regulate cell adhesion and formation of extraembryonic endoderm from pluripotent stem cells. Proc. Natl. Acad. Sci. USA 2013, 110, E1112–E1121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lowe, S.W.; Sherr, C.J. Tumor suppression by Ink4a–Arf: Progress and puzzles. Curr. Opin. Genet. Dev. 2003, 13, 77–83. [Google Scholar] [CrossRef]
- Muniz, V.P.; Barnes, J.M.; Paliwal, S.; Zhang, X.; Tang, X.; Chen, S.; Zamba, K.D.; Cullen, J.; Meyerholz, D.; Meyers, S.; et al. The ARF Tumor Suppressor Inhibits Tumor Cell Colonization Independent of p53 in a Novel Mouse Model of Pancreatic Ductal Adenocarcinoma Metastasis. Mol. Cancer Res. 2011, 9, 867–877. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dhawan, S.; Tschen, S.-I.; Bhushan, A. Bmi-1 regulates the Ink4a/Arf locus to control pancreatic β-cell proliferation. Genes Dev. 2009, 23, 906–911. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mastracci, T.L.; Anderson, K.R.; Papizan, J.B.; Sussel, L. Regulation of Neurod1 Contributes to the Lineage Potential of Neurogenin3+ Endocrine Precursor Cells in the Pancreas. PLoS Genet. 2013, 9, e1003278. [Google Scholar] [CrossRef] [Green Version]
- Hatzistergos, K.E.; Williams, A.R.; Dykxhoorn, D.M.; Bellio, M.A.; Yu, W.; Hare, J.M. Tumor Suppressors RB1 and CDKN2a Cooperatively Regulate Cell-Cycle Progression and Differentiation during Cardiomyocyte Development and Repair. Circ. Res. 2019, 124, 1184–1197. [Google Scholar] [CrossRef] [PubMed]
- Carbone, C.; Piro, G.; Gaianigo, N.; Ligorio, F.; Santoro, R.; Merz, V.; Simionato, F.; Zecchetto, C.; Falco, G.; Conti, G.; et al. Adipocytes sustain pancreatic cancer progression through a non-canonical WNT paracrine network inducing ROR2 nuclear shuttling. Int. J. Obes. 2017, 42, 334–343. [Google Scholar] [CrossRef] [Green Version]
- Russi, S.; Calice, G.; Ruggieri, V.; Laurino, S.; La Rocca, F.; Amendola, E.; Lapadula, C.; Compare, D.; Nardone, G.; Musto, P.; et al. Gastric Normal Adjacent Mucosa Versus Healthy and Cancer Tissues: Distinctive Transcriptomic Profiles and Biological Features. Cancers 2019, 11, 1248. [Google Scholar] [CrossRef] [Green Version]
- Kong, Y.; Sharma, R.B.; Ly, S.; Stamateris, R.; Jesdale, W.; Alonso, L.C. CDKN2A/BT2D Genome-Wide Association Study Risk SNPs Impact Locus Gene Expression and Proliferation in Human Islets. Diabetes 2018, 67, 872–884. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruggieri, V.; Russi, S.; Zoppoli, P.; La Rocca, F.; Angrisano, T.; Falco, G.; Calice, G.; Laurino, S. The Role of MicroRNAs in the Regulation of Gastric Cancer Stem Cells: A Meta-Analysis of the Current Status. J. Clin. Med. 2019, 8, 639. [Google Scholar] [CrossRef] [Green Version]
- Voight, B.F.; The MAGIC Investigators; Scott, L.J.; Steinthorsdottir, V.; Morris, A.P.; Dina, C.; Welch, R.P.; Zeggini, E.; Huth, C.; Aulchenko, Y.; et al. Twelve type 2 diabetes susceptibility loci identified through large-scale association analysis. Nat. Genet. 2010, 42, 579–589. [Google Scholar] [CrossRef] [PubMed]
- Natale, F.; Vivo, M.; Falco, G.; Angrisano, T. Deciphering DNA methylation signatures of pancreatic cancer and pancreatitis. Clin. Epigenetics 2019, 11, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brancaccio, M.; Natale, F.; Falco, G.; Angrisano, T. Cell-Free DNA Methylation: The New Frontiers of Pancreatic Cancer Biomarkers’ Discovery. Genes 2019, 11, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, C.; Gan, Y.; Gu, L.; Wilson, J.; Liu, Z.; Zhang, B.; Deng, D. P16-specific DNA methylation by engineered zinc finger methyltransferase inactivates gene transcription and promotes cancer metastasis. Genome Biol. 2015, 16, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Name | Primer fw 5′–3′ | Primer rv 5′–3′ |
|---|---|---|---|
| Foxa2 | Forkhead Box A2 | CTGGGAGCCGTGAAGATGGAAG | TCCAGCGCCCACATAGGATG |
| Gapdh | Glyceraldehyde 3-phosphate dehydrogenase | AATGGTGAAGGTCGGTGTG | GAAGATGGTGATGGGCTTCC |
| Hnf6 | Hepatocyte nuclear factor 6 | CAAAGAGGTGGCGCAGCGTATC | GCTCTTTCCGTTTGCAGGCTG |
| Ink4a | p16Ink4a | CCCAACGCCCCGAACT | GCAGAAGAGCTGCTACGTGAA |
| Nanog | Nanog homeobox | AACCAGTGGTTGAAGACTAGCAATGGTC | TTCCAGATGCGTTCACCAGATAGC |
| Nepn | Nephrocan | AACCTCTGTGTTGGACAATGC | TCAGAGTTTTGAAGGTGTCATTTT |
| Nkx6.1 | Nk6 homeobox1 | ACTTGGCAGGACCAGAGAGA | AGAGTTCGGGTCCAGAGGTT |
| Oct4 | Octamer-binding transcription factor4 | CCGTGTGAGGTGGAGTCTGGAGAC | CGCCGGTTACAGAACCATACTCG |
| Pdx1 | Pancreatic and duodenal homeobox 1 | GCTCACCTCCACCGGACCTTC | GGGTCCTCTTGTTTTCCTCGGG |
| Arf | p19Arf | TGAGGCTAGAGAGGATCTT | CGTGAACGTTGCCCATCAT |
| Sox9 | Sry-Box transcription factor 9 | GGTCTGCCTGGACTGTATGTGGATG | CTGTCCGATGTCTCTCTGCAGGAG |
| Sox17 | Sry-Box transcription factor 17 | GCCGATGAACGCCTTTATGGTG | CATGCGCTTCACCTGCTTGC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Montano, E.; Pollice, A.; Lucci, V.; Falco, G.; Affinito, O.; La Mantia, G.; Vivo, M.; Angrisano, T. Pancreatic Progenitor Commitment Is Marked by an Increase in Ink4a/Arf Expression. Biomolecules 2021, 11, 1124. https://doi.org/10.3390/biom11081124
Montano E, Pollice A, Lucci V, Falco G, Affinito O, La Mantia G, Vivo M, Angrisano T. Pancreatic Progenitor Commitment Is Marked by an Increase in Ink4a/Arf Expression. Biomolecules. 2021; 11(8):1124. https://doi.org/10.3390/biom11081124
Chicago/Turabian StyleMontano, Elena, Alessandra Pollice, Valeria Lucci, Geppino Falco, Ornella Affinito, Girolama La Mantia, Maria Vivo, and Tiziana Angrisano. 2021. "Pancreatic Progenitor Commitment Is Marked by an Increase in Ink4a/Arf Expression" Biomolecules 11, no. 8: 1124. https://doi.org/10.3390/biom11081124