
Cyclic Dipeptides: The Biological and Structural Landscape with Special Focus on the Anti-Cancer Proline-Based Scaffold

 , , ,
, , ,  , ,
, ,  ,
,  and
and
Abstract
:1. Introduction
2. Occurence and Biosynthesis of DKPs
3. DKP Scaffold
4. Bio-Landscape and Structural Profile of Proline-Based DKPs
4.1. Anticancer Activity
4.1.1. Bicyclic Proline-Based DKP with Simple Side Chains
4.1.2. Bicyclic Proline-Based DKP Modified with Indole-Based Side Chains
4.1.3. Tetracyclic Proline-Based DKP
4.1.4. Penta- and Hexacyclic Proline-Based DKP
4.1.5. Hepta-, Polycyclic and Dimeric Proline-Based DKP
4.1.6. Sulfur-Containing Proline-Based DKPs
4.2. Other Bio-Activities
4.2.1. Neuroprotection
TRH-Related DKP
Unsaturated DKPs
Other DKPs
Mode of Actions
The Blood-Brain Barrier Transporters
Antioxidant Activity
DKP and Gut Microbiota
Protein–Protein Interaction Mechanism
4.2.2. Metalloproteinase Inhibitors
4.2.3. Antibacterial Activity
Quorum-Sensing Agonists and Antagonists
4.2.4. Antifungal Activity
4.2.5. Antiviral Activity
Nucleoside Natural Products
4.2.6. Anthelmintic Activity
4.2.7. Antitoxin Activity
4.2.8. Antagonists of G Protein–Coupled Receptors
Inhibitors of GABAergic Receptors
Antagonists of Serotoninergic 5-HT1A Receptors
Antagonists of Oxytocin Receptor
4.2.9. PDE-5 Inhibitors
4.2.10. PAF Inhibitors: Anti-Inflammatory Activity and Beyond
4.2.11. Cardio-Metabolic Disorders
4.2.12. Hemo-Regulation
4.2.13. FSH Receptor Antagonists
4.2.14. Immunosupressants
4.2.15. Heat Shock Response
4.2.16. Photoprotective Agents
4.2.17. Activators of Dioxygen in Oxidative Processes
4.2.18. Biological Herbicides and Plant Growth Regulation
4.2.19. Biotechnological Applications
Bone Tissue Engineering
Vaccinology
DNA Delivery
Fluorescent DKPs
Inhalable Powder Formulations
Pheromonal Scaffold
5. Supramolecular Structuring of DKPs
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sammes, P.G. Naturally occurring 2,5-dioxopiperazines and related compounds. Fortschr. Chem. Org. Naturst. 1975, 32, 51–118. [Google Scholar]
- De Rosa, S.; Mitova, M.; Tommonaro, G. Marine bacteria associated with sponge as source of cyclic peptides. Biomol. Eng. 2003, 20, 311–316. [Google Scholar] [CrossRef]
- Bugni, T.S.; Ireland, C.M. Marine-derived fungi: A chemically and biologically diverse group of microorganisms. Nat. Prod. Rep. 2004, 21, 143–163. [Google Scholar] [CrossRef] [PubMed]
- Huang, R.; Zhou, X.; Xu, T.; Yang, X.; Liu, Y. Diketopiperazines from marine organisms. Chem. Biodivers. 2010, 7, 2809–2829. [Google Scholar] [CrossRef]
- Ying, J.; Lin, R.; Xu, P.; Wu, Y.; Liu, Y.; Zhao, Y. Prebiotic Formation of Cyclic dipeptides Under Potentially Early Earth Conditions. Sci. Rep. 2018, 8, 936–944. [Google Scholar] [CrossRef] [Green Version]
- Bojarska, J.; Wolf, W.M. Ultra-short cyclo-peptides as bio-inspired therapeutics: Proline-based 2,5-diketopiperazines (DKP). Proceedings 2021, 79, 10. [Google Scholar] [CrossRef]
- Fischer, E. Untersuchungen über aminosäuren, polypeptide und proteine. Ber. Dtsch. Chem. Ges. 1906, 39, 530–610. [Google Scholar] [CrossRef]
- O’Neill, J.C.; Blackwell, H.E. Solid-phase and microwave-assisted syntheses of 2,5-DKPs: Small molecules with great potential. Comb. Chem. High Throughput Screen. 2007, 10, 857–876. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borgman, P.; Lopez, R.D.; Lane, A.L. The expanding spectrum of DKP natural product biosynthetic pathways containing cyclodipeptide synthases. Org. Biomol. Chem. 2019, 17, 2305–2314. [Google Scholar] [CrossRef] [PubMed]
- Canu, N.; Moutiez, M.; Belin, P.; Gondry, M. Cyclodipeptide synthases: A promising biotechnological tool for the synthesis of diverse 2,5-DKPs. Nat. Prod. Rep. 2020, 37, 312–321. [Google Scholar] [CrossRef]
- Apostolopoulos, V.; Bojarska, J.; Chai, T.T.; Elnagdy, S.; Kaczmarek, K.; Matsoukas, J.; New, R.; Parang, K.; Lopez, O.P.; Parhiz, H.; et al. A Global Review on Short Peptides: Frontiers and Perspectives. Molecules 2021, 26, 430. [Google Scholar] [CrossRef]
- Bojarska, J.; Remko, M.; Breza, M.; Madura, I.; Fruziński, A.; Wolf, W.M. A Proline-Based Tectons and Supramolecular Synthons for Drug Design 2.0: A Case Study of ACEI. Pharmaceuticals 2020, 13, 338. [Google Scholar] [CrossRef]
- Otsuka, Y.; Arita, H.; Sakaji, M.; Yamamoto, K.; Kashiwagi, T.; Shimamura, T.; Ukeda, H. Investigation of the formation mechanism of proline-containing cyclic dipeptide from the linear peptide. Biosci. Biotechnol. Biochem. 2019, 83, 2355–2363. [Google Scholar] [CrossRef] [PubMed]
- Song, Z.; Hou, Y.; Yang, Q.; Li, X.; Wu, S. Structures and Biological Activities of Diketopiperazines from Marine Organisms: A Review. Mar. Drugs 2021, 19, 403. [Google Scholar] [CrossRef] [PubMed]
- Abderhalden, E.; Komm, E. The formation of diketopiperazines from polypeptides under various conditions. Z. Physiol. Chem. 1924, 139, 147–152. [Google Scholar] [CrossRef]
- Falorni, M.; Giacomelli, G.; Porcheddu, A.; Taddei, M. Solution-Phase Synthesis of Mixed Amide Libraries by Simultaneous Addition of Functionalities (SPSAF) to a Diketopiperazine Tetracarboxylic Acid Scaffold Monitored by GC Analysis of Isobutyl Alcohol. Eur. J. Org. Chem. 2000, 2000, 1669–1675. [Google Scholar] [CrossRef]
- Martins, M.B.; Carvalho, I. Diketopiperazines: Biological activity and synthesis. Tetrahedron 2007, 63, 9923–9932. [Google Scholar] [CrossRef]
- Harizani, M.; Katsini, E.; Georgantea, P.; Roussis, V.; Ioannou, E. New chlorinated 2,5DKPs from marine-derived bacteria isolated from sediments of the eastern Mediterranean sea. Molecules 2020, 25, 1509. [Google Scholar] [CrossRef] [Green Version]
- Cornacchia, C.; Cacciatore, I.; Baldassarre, L.; Mollica, A.; Feliciani, F.; Pinnen, F. 2,5 Diketopiperazines as neuroprotective agents. Mini Rev. Med. Chem. 2012, 12, 2–12. [Google Scholar] [CrossRef]
- Prasad, C. Bioactive cyclic dipeptides. Peptides 1995, 16, 151–164. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, P.; Ma, H.; Zhu, W. Developments around the bioactive diketopiperazines: A patent review. Expert Opin. Ther. Pat. 2013, 23, 1415–1433. [Google Scholar] [CrossRef] [PubMed]
- Mishra, A.K.; Choi, J.; Choi, S.J.; Baek, K.H. Cyclodipeptides: An overview of their biosynthesis and biological activity. Molecules 2017, 22, 1796. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borthwick, A.D.; Davies, D.E.; Exall, A.M.; Livermore, D.G.; Sollis, S.L.; Nerozzi, F.; Allen, M.J.; Perren, M.; Shabbir, S.S.; Woollard, P.M.; et al. 2,5-DKPs as potent, selective, and orally bioavailable oxytocin antagonists. Synthesis, chirality, and pharmacokinetics. J. Med. Chem. 2005, 48, 6956–6969. [Google Scholar] [CrossRef] [PubMed]
- Sollis, S.L. Short and Novel Stereospecific Synthesis of Trisubstituted 2,5-Diketopiperazines. J. Org. Chem. 2005, 70, 4735–4740. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Y.; Li, Q.; Hanzlik, R.P.; Aube, J. Synthesis of a Small Library of Diketopiperazines as Potential Inhibitors of Calpain. Bioorgan. Med. Chem. Lett. 2005, 15, 3034–3038. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, Y.; Orikasa, S.; Tanaka, K.; Kanoh, K.; Kiso, Y. Total synthesis of anti-microtubule DKP derivatives: Phenylahistin and aurantiamine. J. Org. Chem. 2000, 65, 8402–8405. [Google Scholar] [CrossRef]
- Sun, S.J.; Liu, Y.C.; Weng, C.H.; Sun, S.W.; Li, F.; Li, H.; Zhu, H. Cyclic dipeptides mediating quorum sensing and their biological effects in Hypsizygus Marmoreus. Biomolecules 2020, 10, 298. [Google Scholar] [CrossRef] [Green Version]
- Park, A.R.; Jeong, S.I.; Jeon, H.W.; Kim, J.; Kim, N.; Ha, M.T.; Mannaa, M.; Kim, J.; Lee, C.W.; Min, B.S.; et al. A Diketopiperazine, Cyclo(L-Pro-L-Ile), Derived from Bacillus thuringiensis JCK-1233 Controls Pine Wilt Disease by Elicitation of Moderate Hypersensitive Reaction. Front. Plant Sci. 2020, 11, 1023–1037. [Google Scholar] [CrossRef]
- Kogut, E.F.; Thoen, J.C.; Lipton, M.A. Examination and enhancement of enantioselective autoinduction in cyanohydrin formation by cyclo[(R)-His-(R)-Phe]. J. Org. Chem. 1998, 63, 4604–4610. [Google Scholar] [CrossRef]
- Oku, J.I.; Inoue, S. Asymmetric cyanohydrin synthesis catalysed by a synthetic cyclic dipeptide. J. Chem. Soc. Chem. Commun. 1981, 5, 229–230. [Google Scholar] [CrossRef]
- Huang, R.M.; Yi, X.X.; Zhou, Y.; Su, X.; Pengs, Y.; Gao, C.H. An Update on 2,5-Diketopiperazines from Marine Organisms. Mar. Drugs 2014, 12, 6213–6235. [Google Scholar] [CrossRef] [PubMed]
- Borthwick, A.D. 2,5-Diketopiperazines: Synthesis, reactions, medicinal chemistry, and bioactive natural products. Chem. Rev. 2012, 112, 3641–3716. [Google Scholar] [CrossRef]
- Ressurreição, A.S.M.; Delatouche, R.; Gennari, C.; Piarulli, U. Bifunctional 2,5-diketopiperazines as rigid three-dimensional scaffolds in receptors and peptidomimetics. Eur. J. Org. Chem. 2011, 2, 217–228. [Google Scholar] [CrossRef]
- Chanda, K.; Chou, C.-T.; Lai, J.J.; Lin, S.F.; Yellol, G.S.; Sun, C.M. Traceless synthesis of diketopiperazine fused tetrahydro-β-carbolines on soluble polymer support. Mol. Divers. 2011, 15, 569–581. [Google Scholar] [CrossRef]
- Ressurreicao, A.S.M.; Bordessa, A.; Civera, M.; Belvisi, L.; Gennari, C.; Piarulli, U.J. Synthesis and Conformational Studies of Peptidomimetics Containing a New Bifunctional Diketopiperazine Scaffold Acting as a β-Hairpin Inducer. Org. Chem. 2008, 73, 652–660. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Brahimi, F.; Saragovi, H.U.; Burgess, K.J. Bivalent Diketopiperazine-Based Tropomysin Receptor Kinase C (TrkC) Antagonists. Med. Chem. 2010, 53, 5044–5048. [Google Scholar] [CrossRef] [Green Version]
- Ferlay, J.; Ervik, M.; Lam, F.; Colombet, M.; Mery, L.; Piñeros, M.; Znaor, A.; Soerjomataram, I.; Bray, F. Global Cancer Observatory: Cancer Today; International Agency for Research on Cancer: Lyon, France, 2020. [Google Scholar]
- Elkahoui, S.; Abdel Rahim, H.; Tabbene, O.; Shaaban, M.; Limam, F.; Laatsch, H. Cyclo(His,Leu): A new microbial diketopiperazine from a terrestrial Bacillus subtilis strain B38. Nat. Prod. Res. 2013, 27, 108–116. [Google Scholar] [CrossRef]
- Holden, M.T.; Ram Chhabra, S.; de Nys, R.; Stead, P.; Bainton, N.J.; Hill, P.J.; Manefield, M.; Kumar, N.; Labatte, M.; England, D.; et al. Quorum-sensing cross talk: Isolation and chemical characterization of cyclic dipeptides from Pseudomonas aeruginosa and other Gram-negative bacteria. Mol. Microbiol. 1999, 33, 1254–1266. [Google Scholar] [CrossRef] [PubMed]
- Ström, K.; Sjögren, M.; Broberg, A.; Schnürer, J. Lactobacillus plantarum MiLAB 393 produces the antifungal cyclic dipeptides cyclo(l-Phe-l-Pro) and cyclo(l-Phe-trans-4-OH-l-Pro) and 3-phenyllactic acid. Appl. Environ. Microbiol. 2002, 68, 4322–4327. [Google Scholar] [CrossRef] [Green Version]
- Zhao, P.; Xue, Y.; Li, J.; Li, X.; Zu, X.; Zhao, Z.; Quan, C.; Gao, W.; Feng, S. Non-lipopeptide fungi-derived peptide antibiotics developed since 2000. Biotechnol. Lett. 2019, 41, 651–673. [Google Scholar] [CrossRef] [PubMed]
- Lin, A.; Fang, Y.; Zhu, T.; Gu, Q.; Zhu, W. A new diketopiperazine alkaloid isolated from an algicolous Aspergillus flavus strain. Pharmazie 2008, 63, 323–325. [Google Scholar]
- Stierle, A.C.; Cardellina, J.H.; Strobel, G.A. Maculosin, a host-specific phytotoxin for spotted knapweed from Alternaria alternate. Proc. Natl. Acad. Sci. USA 1988, 85, 8008–8011. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kazlauskas, R.; Murphy, P.T.; Wells, P.J. A diketopiperazine derived from trichloroleucine from the sponge Dysidea herbacea. Tetrahedron Lett. 1978, 49, 4945–4948. [Google Scholar] [CrossRef]
- Prasad, C. Cyclo(His-Pro): Its distribution, origin and function in the human. Neurosci. Biobehav. Rev. 1988, 12, 19–22. [Google Scholar] [CrossRef]
- Ginz, M.; Engelhardt, U.H. Identification of proline-based diketopiperazines in roasted coffee. J. Agr. Food Chem. 2000, 48, 3528–3532. [Google Scholar] [CrossRef]
- Chen, M.Z.; Dewis, M.L.; Kraut, K.; Merritt, D.; Reiber, L.; Trinnaman, L.; Da Costa, N.C. 2,5-Diketopiperazines (cyclic dipeptides) in beef: Identification, synthesis, and sensory evaluation. J. Food Sci. 2009, 74, C100–C105. [Google Scholar] [CrossRef] [PubMed]
- Gautschi, M.; Schmid, J.P.; Peppard, T.L.; Ryan, T.P.; Tuorto, R.M.; Yang, X. Chemical characterization of diketopiperazines in beer. J. Agr. Food Chem. 1997, 45, 3183–3189. [Google Scholar] [CrossRef]
- Li, Y.; Li, F.; Zhu, Y.; Li, X.; Zhou, Z.; Liu, C.; Zhang, W.; Tang, M. DFT study onreaction mechanisms of cyclic dipeptide generation. Struct. Chem. 2016, 27, 1165–1173. [Google Scholar] [CrossRef]
- Crowley, S.; Mahony, J.; van Sinderen, D. Current perspectives on antifungal lactic acid bacteria as natural bio-preservatives. Trends Food Sci. Technol. 2013, 33, 93–109. [Google Scholar] [CrossRef]
- Cui, C.; Kakeya, H.; Osada, H. Novel mammalian cell cycle inhibitors, cyclotropstatins A-D, produced by Aspergillus fumigatus, which inhibit mammalian cell cycle at G2/M phase. Tetrahedron 1997, 53, 59–72. [Google Scholar] [CrossRef]
- Bojarska, J.; Maniukiewicz, W.; Sieron, L.; Remko, M. An orthorhombic polymorph of a cyclization product of perindopril. Acta Crystallogr. C 2013, 69, 630–633. [Google Scholar] [CrossRef] [PubMed]
- Bojarska, J.; Maniukiewicz, W.; Główka, M.L.; Sieron, L.; Remko, M. Crystal structure of perindopril cyclization product. J. Chil. Chem. Soc. 2013, 58, 1530–1532. [Google Scholar] [CrossRef] [Green Version]
- Fischer, P.M. Diketopiperazines in peptide and combinatorial chemistry. J. Peptide Sci. 2003, 9, 9–35. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Conant, C.R.; El-Baba, T.J.; Raab, S.A.; Fuller, D.R.; Hales, D.A.; Clemmer, D.E. Diketopiperazine formation from FPGnK (n = 1-9) peptides: Rates of structural rearrangements and mechanisms. J. Phys. Chem. B 2021, 29, 8107–8116. [Google Scholar] [CrossRef]
- Tareq, F.S.; Kim, J.H.; Lee, M.A.; Lee, H.S.; Lee, J.S.; Lee, Y.J.; Shin, H.J. Antimicrobial gageomacrolactins characterized from the fermentation of the marine-derived bacterium Bacillus subtilis under optimum growth conditions. J. Agric. Food. Chem. 2013, 61, 3428–3434. [Google Scholar] [CrossRef] [PubMed]
- Jia, B.; Ma, Y.; Liu, B.; Chen, P.; Hu, Y.; Zhang, R. Synthesis, Antimicrobial Activity, Structure-Activity Relationship, and Molecular Docking Studies of Indole Diketopiperazine Alkaloids. Front. Chem. 2019, 7, 837–850. [Google Scholar] [CrossRef]
- Nielsen, M.R.; Wollenberg, R.D.; Westphal, K.R.; Sondergaard, T.E.; Wimmer, R.; Gardiner, D.M.; Sørensen, J.L. Heterologous expression of intact biosynthetic gene clusters in Fusarium graminearum. Fungal Genet. Biol. 2019, 132, 103248. [Google Scholar] [CrossRef]
- Giessen, T.W.; Marahiel, M.A. Rational and combinatorial tailoring of bioactive cyclic dipeptides. Front. Microbiol. 2015, 6, 785–796. [Google Scholar] [CrossRef] [PubMed]
- Giessen, T.W.; Von Tesmar, A.M.; Marahiel, M.A. Insights into the generation of structural diversity in a tRNA-dependent pathway for highly modified bioactive cyclic dipeptides. Chem. Biol. 2013, 20, 828–838. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schofield, M.M.; Sherman, D.H. Meta-omic characterization of prokaryotic gene clusters for natural product biosynthesis. Curr. Opin. Biotechnol. 2013, 24, 1151–1158. [Google Scholar] [CrossRef] [Green Version]
- Fischbach, M.A.; Walsh, C.T.; Clardy, J. The evolution of gene collectives: How natural selection drives chemical innovation. Proc. Natl. Acad. Sci. USA 2008, 105, 4601–4608. [Google Scholar] [CrossRef] [Green Version]
- Dasgupta, A.; Chowdhury, N.; De, R.K. Metabolic pathway engineering: Perspectives and applications. Comput. Methods Programs Biomed. 2020, 192, 105436. [Google Scholar] [CrossRef] [PubMed]
- Boettcher, M.; McManus, M.T. Choosing the right tool for the job: RNAi, TALEN, or CRISPR. Mol. Cell. 2015, 58, 575–585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leitão, A.L.; Costa, M.C.; Enguita, F.J. Applications of genome editing by programmable nucleases to the metabolic engineering of secondary metabolites. J. Biotechnol. 2017, 241, 50–60. [Google Scholar] [CrossRef]
- Shi, T.Q.; Liu, G.N.; Ji, R.Y.; Shi, K.; Song, P.; Ren, L.J.; Huang, H.; Ji, X.J. CRISPR/Cas9-based genome editing of the filamentous fungi: The state of the art. Appl. Microbiol. Biotechnol. 2017, 101, 7435–7443. [Google Scholar] [CrossRef] [PubMed]
- Jiang, C.; Lv, G.; Tu, Y.; Cheng, X.; Duan, Y.; Zeng, B.; He, B. Applications of CRISPR/Cas9 in the synthesis of secondary metabolites in filamentous fungi. Front. Microbiol. 2021, 12, 638096–638111. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Chen, L.; Jiang, Y.; Zhou, Z.; Zou, G. Efficient genome editing in filamentous fungus Trichoderma reesei using the CRISPR/Cas9 system. Cell Discov. 2015, 1, 15007–15011. [Google Scholar] [CrossRef] [Green Version]
- Nodvig, C.S.; Nielsen, J.B.; Kogle, M.E.; Mortensen, U.H. A CRISPR-Cas9 System for genetic engineering of filamentous fungi. PLoS ONE 2015, 10, e0133085. [Google Scholar] [CrossRef] [Green Version]
- Hu, Y.; Stumpfe, D.; Bajorath, J. Computational exploration of molecular scaffolds in medicinal chemistry. J. Med. Chem. 2016, 59, 4062–4076. [Google Scholar] [CrossRef]
- Bemis, G.W.; Murcko, M.A. The properties of known drugs. Molecular frameworks. J. Med. Chem. 1996, 39, 2887–2893. [Google Scholar] [CrossRef]
- Bajorath, J. Computational scaffold hopping: Cornerstone for the future of drug design. Future Med. Chem. 2017, 9, 629–631. [Google Scholar] [CrossRef] [PubMed]
- Ertl, P. Intuitive ordering of scffolds and scaffold similarity searching using scaffold keys. J. Chem. Inf. Model. 2014, 54, 1617–1622. [Google Scholar] [CrossRef] [PubMed]
- Balachandra, C.; Padhi, D.; Govindaraju, T. Cyclic dipeptide: A privileged molecular scaffold to derive structural diversity and functional utility. ChemMedChem 2021, 16, 1–31. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.M.; Liang, X.A.; Kong, Y.; Jia, B. Structural diversity and biological activities of indole diketopiperazine alkaloids from fungi. J. Agric. Food Chem. 2016, 64, 6659–6671. [Google Scholar] [CrossRef]
- Sano, S.; Nakao, M. Chemistry of 2,5-diketopiperazine and its bis-lactim ether: A brief review. Heterocycles 2015, 91, 1349–1375. [Google Scholar] [CrossRef]
- Corey, R.B. The crystal structure of giketopiperazine. J. Am. Chem. Soc. 1938, 60, 1598–1604. [Google Scholar] [CrossRef]
- Sung, B.J.; Hwang, K.Y.; Jeon, Y.H.; Lee, J.I.; Heo, Y.S.; Kim, J.H.; Moon, J.; Yoon, J.M.; Hyun, Y.L.; Kim, E.; et al. Structure of the catalytic domain of human phosphodiesterase 5 with bound drug molecules. Nature 2003, 425, 98–102. [Google Scholar] [CrossRef]
- Daugan, A.; Grondin, P.; Ruault, C.; Le Monnier de Gouville, A.C.; Coste, H.; Kirilovsky, J.; Hyafil, F.; Labaudiniere, R. The discovery of tadalafil: A novel and highly selective PDE5 inhibitor. 1: 5,6,11,11a-tetrahydro-1H-imidazo[10,50:1,6]pyrido[3,4-b]indole-1,3(2H)-dione analogues. J. Med. Chem. 2003, 46, 4525–4532. [Google Scholar] [CrossRef]
- Daugan, A.; Grondin, P.; Ruault, C.; Le Monnier de Gouville, A.C.; Coste, H.; Linget, J.M.; Kirilovsky, J.; Hyafil, F.; Labaudiniere, R. The discovery of tadalafil: A novel and highly selective PDE5 inhibitor. 2: 2,3,6,7,12,12a-hexahydropyrazino[10,20:1,6]pyrido[3,4-b]indole-1,4-dione analogues. J. Med. Chem. 2003, 46, 4533–4542. [Google Scholar] [CrossRef]
- Liddle, J.; Allen, M.J.; Borthwick, A.D.; Brooks, D.P.; Davies, D.E.; Edwards, R.M.; Exall, A.M.; Hamlett, C.; Irving, W.R.; Mason, A.M.; et al. The discovery of GSK221149A: A potent and selective oxytocin antagonist. Bioorgan. Med. Chem. Lett. 2008, 18, 90–94. [Google Scholar] [CrossRef] [PubMed]
- Maeda, K.; Nakata, H.; Koh, Y.; Miyakawa, T.; Ogata, H.; Takaoka, Y. Spiro diketopiperazine-based CCR5 inhibitor whichpreservesCC- chemokine/CCR5 interactions and exerts potent activitya gainst R5 human immuno deficiency virus type 1 in vitro. J. Virol. 2004, 78, 8654–8662. [Google Scholar] [CrossRef] [Green Version]
- Gomes, N.G.M.; Lefranc, F.; Kijjoa, A.; Kiss, R. Can some marine-derived fungal metabolites become actual anticancer agents? Mar. Drugs 2015, 13, 3950–3991. [Google Scholar] [CrossRef]
- Mohanlal, R.W.; Lloyd, K.; Huang, L. Plinabulin, a novel small molecule clinical stage IO agent with anti-cancer activity, to prevent chemo–induced neutropenia and immune related AEs. J. Clin. Oncol. 2018, 36, 126. [Google Scholar] [CrossRef]
- Kanzaki, H.; Yanagisawa, S.; Kanoh, K.; Nitoda, T. A novel potent cell cycle inhibitor dehydrophenylahisti enzymatic synthesis and inhibitory activity towards eaurchin embryo. J. Antibiot. 2002, 55, 1042–1047. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gu, B.; He, S.; Yan, X.; Zhang, L. Tentative biosynthetic pathways of some microbial diketopiperazines. Appl. Microbiol. Biotechnol. 2013, 97, 8439–8453. [Google Scholar] [CrossRef] [PubMed]
- Sugie, Y.; Hirai, H.; Inagaki, T.; Ishiguro, M.; Kim, Y.-J.; Kojima, Y.; Sakakibara, T.; Sakemi, S.; Sugiura, A.; Suzuki, Y.; et al. A new antibiotic CJ-17,665 from Aspergillus ochraceus. J. Antibiot. 2001, 54, 911–916. [Google Scholar] [CrossRef] [Green Version]
- Pardo, J.; Urban, C.; Galvez, E.M.; Ekert, P.G.; Müller, U.; Kwon-Chung, J.; Lobigs, M.; Müllbacher, A.; Wallich, R.; Borner, C.; et al. The mitochondrial protein Bak is pivotal for gliotoxin-induced apoptosis and a critical host factor of Aspergillus fumigatus virulence in mice. J. Cell Biol. 2006, 174, 509–519. [Google Scholar] [CrossRef]
- Waring, P.; Eichner, R.D.; Müllbacher, A.; Sjaarda, A. Gliotoxin induces apoptosis in macrophages unrelated to its antiphagocytic properties. J. Biol. Chem. 1988, 263, 18493–18499. [Google Scholar] [CrossRef]
- King, R.R.; Calhoun, L.A. The thaxtomin phytotoxins: Sources, synthesis, biosynthesis, biotransformation and biological activity. Phytochemistry 2009, 70, 833–841. [Google Scholar] [CrossRef]
- Borthwick, A.; Da Costa, N.C. 2,5-DKPs in food and beverages: Taste and bioactivity. Crit. Rev. Food Sci. Nutr. 2017, 718–742. [Google Scholar] [CrossRef]
- Milne, P.J.; Kilian, G. The properties, formation, and biological activity of 2,5-DKPs. Comprehensive Natural Products II. Reference Module in Chemistry. Mol. Sci. Chem. Eng. Chem. Biol. 2010, 5, 657–698. [Google Scholar]
- Ono, S.; Yamafuji, T.; Yamamoto, H.; Egawa, H.; Furuta, Y.; Kaga, H. Novel 2,3-DKP Derivative or Salt Thereof. Patent WO1996016947, 24 November 1995. [Google Scholar]
- Goulding, C.E.; Pollard, C.B. Synthesis of 2,3-DKP. J. Am. Chem. Soc. 1948, 70, 1967–1968. [Google Scholar] [CrossRef]
- Deveau, A.M.; Labroli, M.A.; Dieckhaus, C.M.; Barthen, M.T.; Smith, K.S.; Macdonald, T.L. The synthesis of amino-acid functionalized beta-carbolines as topoisomerase II inhibitors. Bioorgan. Med. Chem. Lett. 2001, 11, 1251–1255. [Google Scholar] [CrossRef]
- Dawidowski, M.; Turło, J. Multicomponent synthesis and anticonvulsant activity of monocyclic 2,6-DKP derivatives. Med. Chem. Res. 2014, 23, 2007–2018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fytas, C.; Zoidis, G.; Fytas, G. A facile and effective synthesis of lipophilic 2,6-DKP analogues. Tetrahedron 2008, 64, 6749–6754. [Google Scholar] [CrossRef]
- Fytas, C.; Zoidis, G.; Tzoutzas, N.; Taylor, M.C.; Fytas, G.; Kelly, J.M. Novel lipophilic acetohydroxamic acid derivatives based on conformationally constrained spiro carbocyclic 2,6-DKP scaffolds with potent trypanoicidal activity. J. Med. Chem. 2011, 54, 5250–5254. [Google Scholar] [CrossRef] [PubMed]
- Brady, R.M.; Zhang, M.; Gable, R.; Norton, R.S.; Baell, J.B. De novo design and synthesis of a conotoxin KIIIA peptidomimetic. Bioorgan. Med. Chem. Let. 2013, 23, 4892–4895. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, H.; Murao, S.; Arai, M. New dihydroquinoline toxic to artemia salina produced by Penicillium sp. NTC-47. Chem. Express 1991, 6, 989–992. [Google Scholar]
- Deffieux, G.; Filleau, M.J.; Baute, R. New antibiotics from the fungus Epicoccum nigrum. III. Epicorazine B: Structure elucidation and absolute configuration. J. Antibiot. 1978, 31, 1106–1109. [Google Scholar] [CrossRef]
- Seigle Murandi, F.; Krivobok, S.; Steiman, R.; Marzin, D. Production, mutagenicity, and immunotoxicity of gliotoxin. J. Agric. Food Chem. 1990, 38, 1854–1856. [Google Scholar] [CrossRef]
- Takahashi, C.; Numata, A.; Ito, Y.; Matsumura, E.; Araki, H.; Iwaki, H.; Kushida, K. Leptosins, antitumor metabolites of a fungus isolated from a marine alga. J. Chem. Soc. Perkin Trans. 1994, 1, 1859–1864. [Google Scholar] [CrossRef]
- Cavelier, F.; Marchand, D.; Mbassi, P.; Martinez, J.; Marraud, M. Conformational studies ofproline-, thiaproline-and dimethylsilaproline-containing diketopiperazines. J. Pept. Sci. 2006, 12, 621–625. [Google Scholar] [CrossRef]
- Ye, X.; Chai, W.; Lian, X.Y.; Zhang, Z. Novel propanamide analogue and antiproliferative diketopiperazines from mangrove Streptomyces sp. Q24. Nat. Prod. Res. 2017, 31, 1390–1396. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, E.G.; Torres, M.C.M.; da Silva, A.B.; Colares, L.L.F.; Pires, K.; Lotufo, T.M.C.; Silveira, E.R.; Pessoa, O.D.L.; Costa-Lotufo, L.V.; Jimenez, P.C. Prospecting anticancer compounds in Actinomycetes recovered from the sediments of Saint Peter and Saint Paul’s Archipelago, Brazil. Chem. Biodivers. 2016, 13, 1149–1157. [Google Scholar] [CrossRef] [PubMed]
- Lin, W.X.; Xie, C.L.; Zhou, M.; Xia, M.L.; Zhou, T.T.; Chen, H.F.; Yang, X.W.; Yang, Q. Chemical constituents from the deep sea-derived Streptomyces xiamenensis MCCC 1A01570 and their effects on RXRα transcriptional regulation. Nat. Prod. Res. 2020, 34, 1461–1464. [Google Scholar] [CrossRef] [PubMed]
- Shaala, L.A.; Youssef, D.T.A.; Badr, J.M.; Harakeh, S.M. Bioactive 2(1H)-pyrazinones and diketopiperazine alkaloids from a tunicate-derived Actinomycete Streptomyces sp. Molecules 2016, 21, 1116. [Google Scholar] [CrossRef]
- Ovenden, S.P.B.; Nielson, J.L.; Liptrot, C.H.; Willis, R.H.; Tapiolas, D.M.; Wright, A.D.; Motti, C.A. A new diketopiperazine, cyclo(4-S-hydroxy-R-proline-R-isoleucine), from an australian specimen of the Sponge Stelletta sp. Mar. Drugs 2011, 9, 2469–2478. [Google Scholar] [CrossRef] [PubMed]
- Youssef, D.T.A.; Alahdal, A.M. Cytotoxic and antimicrobial compounds from the marine-derived fungus, Penicillium Spec. Molecules 2018, 23, 394. [Google Scholar] [CrossRef] [Green Version]
- Wang, N.; Cui, C.B.; Li, C.W. A new cyclic dipeptide penicimutide: The activated production of cyclic dipeptides by introduction of neomycin-resistance in the marine-derived fungus Penicillium purpurogenum G59. Arch. Pharm. Res. 2016, 39, 762–770. [Google Scholar] [CrossRef]
- Li, B.; Chen, G.; Bai, J.; Jin, Y.K.; Pei, Y.H. A bisamide and four diketopiperazines from a marine-derived Streptomyces sp. J. Asian Nat. Prod. Res. 2011, 13, 1146–1150. [Google Scholar] [CrossRef]
- Vázquez-Riviera, D.; González, O.; Guzmán-Rodríguez, J.; Díaz-Pérez, A.L.; Ochoa-Zarzosa, A.; López-Bucio, J.; Meza-Carmen, V.; Campos-García, J. Cytotoxicity of cyclodipeptides from Pseudomonas aeruginosa PAO1 leads to apoptosis in human cancer cell lines. BioMed Res. Int. 2015, 2015, 197608. [Google Scholar] [CrossRef] [Green Version]
- Zhao, S.; Smith, K.S.; Deveau, A.M.; Dieckhaus, C.M.; Johnson, M.A.; Macdonald, T.L.; Cook, J.M. Biological activity of the tryprostatins and their diastereomers on human carcinoma cell lines. J. Med. Chem. 2002, 45, 1559–1562. [Google Scholar] [CrossRef]
- Jain, H.D.; Zhang, C.; Zhou, S.; Zhou, H.; Ma, J.; Liu, X.; Liao, X.; Deveau, A.M.; Dieckhaus, C.M.; Johnson, M.A.; et al. Synthesis and structure–activity relationship studies on tryprostatin A, an inhibitor of breast cancer resistance protein. Bioorgan. Med. Chem. Lett. 2008, 16, 4626–4651. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kozlovsky, A.; Vinokurova, N.G.; Adanin, V.M.; Gräfe, U. Piscarinines, new polycyclic diketopiperazine alkaloids from Penicillium piscarium NKM F-691. Nat. Prod. Lett. 2000, 14, 333–340. [Google Scholar] [CrossRef]
- Zhelifonova, V.P.; Maier, A.; Kozlovskii, A.G. Effect of various factors on the biosynthesis of piscarinines, secondary metabolites of the fungus Penicillium piscarium Westling. Appl. Biochem. Microbiol. 2008, 44, 608–612. [Google Scholar] [CrossRef]
- Kato, H.; Yoshida, T.; Tokue, T.; Nojiri, Y.; Hirota, H.; Ohta, T.; Williams, R.M.; Tsukamoto, S. Notoamides A–D: Prenylated indole alkaloids isolated from a marine-derived fungus, Aspergillus sp. Angew. Chem. Int. Ed. 2007, 46, 2254–2256. [Google Scholar] [CrossRef]
- Afiyatullow, S.S.; Zhuravleva, O.I.; Antonov, A.S.; Berdyshev, D.V.; Pivkin, M.V.; Denisenko, V.A.; Popov, R.S.; Gerasimenko, A.V.; von Amsberg, G.; Dyshlovoy, S.A.; et al. Prenylated indole alkaloids from co-culture of marine-derived fungi Aspergillus sulphureus and Isaria felina. J. Antibiot. 2018, 71, 846–853. [Google Scholar] [CrossRef]
- Xianglan, K.; Shengxin, C.; Tianjiao, Z.; Qianqun, G.; Dehai, L.; Yepeng, L. Secondary metabolites of a deep sea derived fungus Aspergillus versicolor CXCTD-06-6a and their bioactivity. J. Ocean Univ. China 2014, 13, 691–695. [Google Scholar]
- Che, Q.; Zhu, T.; Qi, X.; Mándi, A.M.; Kurtán, T.; Mo, X.; Li, J.; Gu, Q.; Li, D. Hybrid isoprenoids from a reeds rhizosphere soil derived actinomycete Streptomyces sp. CHQ-64. Org. Lett. 2012, 14, 3438–3441. [Google Scholar] [CrossRef]
- Che, Q.; Zhu, T.; Keysers, R.A.; Liu, X.; Li, J.; Gu, Q.; Li, D. Polycyclic hybrid isoprenoids from a reed rhizosphere soil derived Streptomyces sp. CHQ-64. J. Nat. Prod. 2013, 76, 759–763. [Google Scholar] [CrossRef]
- Yu, X.; Müller, W.E.G.; Guo, Z.; Lin, W.; Zou, K.; Liu, Z.; Proksch, P. Indole alkaloids from the coprophilous fungus Aphanoascus fulvescens. Fitoterapia 2019, 136, 104168. [Google Scholar] [CrossRef]
- Cai, S.; Sun, S.; Peng, J.; Konh, X.; Zhou, H.; Zhu, T.; Gu, Q.; Li, D. Okaramines S-U, three new indole diketopiperazine alkaloids from Aspergillus taichungensis ZHN-7-07. Tetrahedron 2015, 71, 3715–3719. [Google Scholar] [CrossRef]
- Du, L.; Zhu, T.; Cai, S.; Wang, F.; Xiao, X.; Gu, Q. New alkaloids and diterpenes from a deep ocean sediment derived fungus Penicillium sp. Tetrahedron 2009, 65, 1033–1039. [Google Scholar] [CrossRef]
- Wang, N.; Dong, Y.; Yang, Y.; Xu, R.; Li, C.W.; Cui, C.B. Penicimutanin C, a new alkaloidal compound, isolated from a neomycin-resistant mutant 3-f-31 of Penicillium purpurogenum G59. Chem. Biodivers. 2020, 17, e2000241. [Google Scholar] [CrossRef] [PubMed]
- Zhong, W.M.; Wang, J.F.; Shi, X.F.; Wei, X.Y.; Chen, Y.C.; Zeng, Q.; Xiang, Y.; Chen, X.Y.; Tian, X.P.; Xiao, Z.H.; et al. Eurotiumins A–E, five new alkaloids from the marine-derived fungus Eurotium sp. SCSIO F452. Mar. Drugs 2018, 16, 136. [Google Scholar] [CrossRef] [Green Version]
- Cui, C.-B.; Kakeya, H.; Osada, H. Novel mammalian cell cycle inhibitors, Spirotryprostatins A and B, produced by Aspergillus fumigatus, which inhibit mammalian cell cycle at G2/M phase. Tetrahedron 1996, 52, 12651–12666. [Google Scholar] [CrossRef]
- Cui, C.B.; Kakeya, H.; Osada, H. Spirotryprostatin B, a novel mammalian cell cycle inhibitor produced by Aspergillus fumigatus. J. Antibiot. 1996, 49, 832–835. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sebahar, P.R.; Osada, H.; Usui, T.; Williams, R.M. Asymmetric, stereocontrolled total synthesis of (+) and (−)-spirotryprostatin B via a diastereoselective azomethine ylide [1,3]-dipolar cycloaddition reaction. Tetrahedron 2002, 58, 6311–6322. [Google Scholar] [CrossRef]
- Zhang, Y.H.; Geng, C.; Zhang, X.W.; Zhu, H.J.; Shao, C.L.; Cao, F.; Wang, C.Y. Discovery of bioactive indole-diketopiperazines from the marine-derived fungus Penicillium brasilianum aided by genomic information. Mar. Drugs 2019, 17, 514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, F.; Sun, Y.L.; Liu, K.S.; Zhang, X.Y.; Qian, P.Y.; Wang, Y.F.; Qi, S.H. Indole alkaloids from marine-derived fungus Aspergillus sydowii SCSIO 00305. J. Antibiot. 2012, 65, 109–111. [Google Scholar] [CrossRef] [Green Version]
- Peng, J.; Gao, H.; Li, J.; Ai, J.; Geng, M.; Zhang, G.; Zhu, T.; Gu, Q.; Li, D. Prenylated indole diketopiperazines from the marine-derived fungus Aspergillus versicolor. J. Org. Chem. 2014, 79, 7895–7904. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Xu, D.; Sun, W.; Yang, B.; Li, F.; Liu, M.; Wang, J.; Xue, Y.; Hu, Z.; Zhang, Y. HPLC-DAD-directed isolation of linearly fused prenylated indole alkaloids from a soil-derived Aspergillus versicolor. J. Nat. Prod. 2019, 82, 2181–2188. [Google Scholar] [CrossRef]
- Chang, Y.W.; Yuan, C.M.; Zhang, J.; Liu, S.; Cao, P.; Hua, H.M.; Di, Y.T.; Hao, X.-J. Speramides A–B, two new prenylated indole alkaloids from the freshwater-derived fungus Aspergillus ochraceus KM007. Tetrahedron Lett. 2016, 57, 4952–4955. [Google Scholar] [CrossRef]
- Qian-Cutrone, J.; Huang, S.; Shu, Y.Z.; Vyas, D.; Fairchild, C.; Menendez, A.; Krampitz, K.; Dalterio, R.; Klohr, S.E.; Gao, Q. Stephacidin A and B: Two structurally novel, selective inhibitors of the testosterone-dependent prostate LNCaP cells. J. Am. Chem.Soc. 2002, 124, 14556–14557. [Google Scholar] [CrossRef] [PubMed]
- Che, Q.; Li, J.; Li, D.; Gu, Q.; Zhu, T. Structure and absolute configuration of drimentine I, an alkaloid from Streptomyces sp. CHQ-64. J. Antibiot. 2016, 69, 467–469. [Google Scholar] [CrossRef]
- He, W.; Xu, Y.; Fu, P.; Zuo, M.; Liu, W.; Jiang, Y.; Wang, L.; Zhu, W. Cytotoxic indolyl diketopiperazines from the Aspergillus sp. GZWMJZ-258, endophytic with the medicinal and edible plant Garcinia multiflora. J. Agric. Food Chem. 2019, 67, 10660–10666. [Google Scholar] [CrossRef]
- Wulff, J.E.; Siegrist, R.; Myers, A.G. The natural product Avrainvillamide binds to the oncoprotein nucleophosmin. J. Am. Chem. Soc. 2007, 129, 14444–14451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andresen, V.; Eriksen, B.S.; Mukherjee, H.; Sulen, A.; Popa, M.; Sørnes, S.; Reikvam, H.; Chan, K.P.; Hovland, R.; McCormack, E.; et al. Anti-proliferative activity of the NPM1 interacting natural product avrainvillamide in acute myeloid leukemia. Cell Death Dis. 2016, 7, e2497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, F.; Sarotti, A.M.; Jiang, G.; Huguet-Tapia, C.H.; Zheng, S.L.; Wu, X.; Li, C.; Ding, Y.; Cao, S. Waikikiamides A−C: Complex diketopiperazine dimer and diketopiperazine−polyketide hybrids from a hawaiian marine fungal strain Aspergillus sp. FM242. Org. Lett. 2020, 22, 4408–4412. [Google Scholar] [CrossRef]
- Raju, E.; Piggott, A.M.; Conte, M.; Aalbersberg, W.G.L.; Feussner, K.; Capon, R.J. Naseseazines A and B: A new dimeric diketopiperazine framework from a marine-derived actinomycete, Streptomyces sp. Org. Lett. 2009, 11, 3862–3865. [Google Scholar] [CrossRef] [PubMed]
- Gu, B.B.; Gui, Y.H.; Liu, L.; Su, Z.Y.; Jiao, W.H.; Li, L.; Sun, F.; Wang, S.P.; Yang, F.; Lin, H.-W. A new asymmetric diketopiperazine dimer from the sponge-associated fungus Aspergillus versicolor 16F-11. Magn. Reson. Chem. 2019, 57, 49–54. [Google Scholar] [CrossRef] [Green Version]
- Fan, Z.; Sun, Z.H.; Liu, Z.; Chen, Y.C.; Liu, H.X.; Li, H.H.; Zhang, W.M. Dichotocejpins A–C: New diketopiperazines from a deep-sea-derived fungus Dichotomomyces cejpii FS110. Mar. Drugs 2016, 14, 164. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.; Takada, K.; Takemoto, Y.; Yohida, M.; Nogi, Y.; Okada, S.; Matsunaga, S. Gliotoxin analogues from a marine-derived fungus, Penicillium sp., and their cytotoxic and histone methyltransferase inhibitory activities. J. Nat. Prod. 2012, 75, 111–114. [Google Scholar] [CrossRef]
- Liang, W.L.; Le, X.; Li, H.J.; Yang, X.L.; Chen, J.X.; Xu, J.; Liu, H.L.; Wang, L.Y.; Wang, K.T.; Hu, K.C.; et al. Exploring the chemodiversity and biological activities of the secondary metabolites from the marine fungus Neosartorya pseudofischeri. Mar. Drugs 2014, 12, 5657–5676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, W.Y.; Zhu, T.J.; Han, X.X.; Fan, G.T.; Liu, H.B.; Zhu, W.M.; Gu, Q.Q. A new gliotoxin analogue from a marine-derived fungus Aspergillus fumigatus Fres. Nat. Prod. Lett. 2009, 23, 203–207. [Google Scholar] [CrossRef]
- Wang, Y.; Li, Z.-L.; Bai, J.; Zhang, L.-M.; Wu, X.; Zhang, L.; Pei, Y.-H.; Jing, Y.-K.; Hua, H.-M. 2,5-Diketopiperazines from the marine-derived fungus Aspergillus fumigatus YK-7. Chem. Biodivers. 2012, 9, 385–393. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Q.H.; Yang, Z.D.; Shu, Z.M.; Wang, Y.G.; Wang, M.G. Secondary Metabolites and Biological Activities of Talaromyces sp. LGT-2, an Endophytic Fungus from Tripterygium wilfordii. Iran. J. Pharm. Res. 2016, 15, 453–457. [Google Scholar] [PubMed]
- Park, H.B.; Kwon, H.C.; Lee, C.H.; Yang, H.O. Glionitrin A, an antibiotic-antitumor metabolite derived from competitive interaction between abandoned mine microbes. J. Nat. Prod. 2009, 72, 248–252. [Google Scholar] [CrossRef] [PubMed]
- Park, H.B.; Kim, Y.J.; Park, J.S.; Yang, H.O.; Lee, K.R.; Kwon, H.C. Glionitrin B, a cancer invasion inhibitory diketopiperazine produced by microbial coculture. J. Nat. Prod. 2011, 74, 2309–2312. [Google Scholar] [CrossRef]
- Kong, F.; Wang, Y.; Liu, P.; Dong, T.; Zhu, W. Thiodiketopiperazines from the marine-derived fungus Phoma sp. OUCMDZ-1847. J. Nat. Prod. 2014, 77, 132–137. [Google Scholar] [CrossRef]
- Baumann, M.; Dieskau, A.; Loertscher, B.M.; Walton, M.C.; Nam, S.; Xie, J.; Horne, D.; Overman, L.E. Tricyclic analogues of epidithiodioxopiperazine alkaloids with promising in vitro and in vivo antitumor activity. Chem. Sci. 2015, 6, 4451–4457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, F.Z.; Huang, Z.; Shi, X.F.; Chen, Y.C.; Zhang, W.M.; Tian, X.P.; Li, J.; Zhang, S. Cytotoxic indole diketopiperazines from the deep sea-derived fungus Acrostalagmus luteoalbus SCSIO F457. Bioorgan. Med. Chem. Lett. 2012, 22, 7265–7267. [Google Scholar] [CrossRef]
- Rivera-Chávez, J.; El-Elimat, T.; Gallagher, J.M.; Graf, T.N.; Fourier, J.; Panigrahi, G.K.; Deep, G.; Bunch, R.L.; Raja, H.A.; Oberlies, N.H. Delitpyrones: α-Pyrone derivatives from a freshwater Delitschia sp. Planta Med. 2019, 85, 62–71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meng, L.H.; Li, X.M.; Lv, C.T.; Huang, C.G.; Wang, B.G. Brocazines A−F, cytotoxic bisthiodiketopiperazine derivatives from Penicillium brocae MA-231, an endophytic fungus derived from the marine mangrove plant Avicennia marina. J. Nat. Prod. 2014, 77, 1921–1927. [Google Scholar] [CrossRef] [PubMed]
- Meng, L.H.; Wang, C.Y.; Mándi, A.; Li, X.M.; Hu, X.Y.; Kassack, M.U.; Kurta, T.; Wang, B.G. Three diketopiperazine alkaloids with spirocyclic skeletons and one bisthiodiketopiperazine derivative from the mangrove-derived endophytic fungus Penicillium brocae MA-231. Org. Lett. 2016, 18, 5304–5307. [Google Scholar] [CrossRef] [PubMed]
- Chi, L.P.; Li, X.M.; Li, L.; Li, C.; Wang, B.G. Cytotoxic thiodiketopiperazine derivatives from the deep sea-derived fungus Epicoccum nigrum SD-388. Mar. Drugs 2020, 18, 160. [Google Scholar] [CrossRef] [Green Version]
- Chinworrungsee, M.; Kittakoop, P.; Saenboonrueng, J.; Kongsaeree, P.; Thebtaranonth, Y. Bioactive compounds from the seed fungus Menisporopsis theobromae BCC 3975. J. Nat. Prod. 2006, 69, 1404–1410. [Google Scholar] [CrossRef]
- Yun, K.; Khong, T.T.; Leutou, A.S.; Kim, G.D.; Hong, J.; Lee, c.H.; Son, B.W. Cristazine, a new cytotoxic dioxopiperazine alkaloid from the mudflat-sediment-derived fungus Chaetomium cristatum. Chem. Pharm. Bull. 2016, 64, 59–62. [Google Scholar] [CrossRef] [Green Version]
- Jo, M.J.; Patil, M.P.; Jung, H.I.; Seo, Y.B.; Lim, H.K.; Son, B.W.; Kim, G.D. Cristazine, a novel dioxopiperazine alkaloid, induces apoptosis via the death receptor pathway in A431 cells. Drug Dev. Res. 2019, 80, 504–512. [Google Scholar] [CrossRef]
- Lai, Y.S.; Chen, J.Y.; Tsai, H.J.; Chen, T.Y.; Hung, W.C. The SUV39H1 inhibitor chaetocin induces differentiation and shows synergistic cytotoxicity with other epigenetic drugs in acute myeloid leukemia cells. Blood Cancer J. 2015, 5, e313–e319. [Google Scholar] [CrossRef]
- Figueroa, M.; Graf, T.N.; Ayers, S.; Adcock, A.F.; Kroll, D.J.; Yang, J.; Swanson, S.M.; Munoz-Acuna, U.; Carcache de Blanco, E.J.; Agrawal, R.; et al. Cytotoxic epipolythiodioxopiperazine alkaloids from filamentous fungi of the Bionectriaceae. J. Antibiot. 2012, 65, 559–564. [Google Scholar] [CrossRef] [Green Version]
- Amrine, C.S.M.; Huntsman, A.C.; Doyle, M.G.; Burdette, J.E.; Pearce, C.J.; Fuchs, J.R.; Oberlies, N.H. Semisynthetic derivatives of the Verticillin class of natural products through acylation of the C11 hydroxy group. ACS Med. Chem. Lett. 2021, 12, 625–630. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Li, X.; Tu, K.; Guan, Y.; Fung, K.P.; Liu, F. Verticillin A suppresses HGF-induced migration and invasion via repression of the c-Met/FAK/Src pathway in human gastric and cervical cancer cells. OncoTargets Ther. 2019, 12, 5823–5833. [Google Scholar] [CrossRef] [Green Version]
- Salvi, A.; Amrine, C.S.M.; Austin, J.R.; Kilpatrick, K.; Russo, A.; Lanvit, D.; Calderon-Gierszal, E.; Mattes, Z.; Pearce, C.J.; Grinstaff, M.W.; et al. Verticillin A causes apoptosis and reduces tumor burden in high-grade serous ovarian cancer by inducing DNA damage. Mol. Cancer Ther. 2020, 19, 89–100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Son, B.W.; Jensen, P.R.; Kauffman, C.A.; Fenical, W. New cytotoxic epidithiodioxopiperazhes related to Verticillin A from a marine isolate of the fungus Penicillium. Nat. Prod. Lett. 1999, 13, 213–222. [Google Scholar] [CrossRef]
- Zhang, Y.X.; Chen, Y.; Guo, X.N.; Zhang, X.W.; Zhao, W.M.; Zhong, L.; Zhou, J.; Xia, Y.; Lin, L.P.; Dinga, J. 11,11’-Dideoxy-verticillin: A natural compound possessing growth factor receptor tyrosine kinase-inhibitory effect with anti-tumor activity. Anticancer Drugs 2005, 16, 515–524. [Google Scholar] [CrossRef] [PubMed]
- Isham, C.R.; Tibodeau, J.D.; Jin, W.; Xu, R.; Timm, M.M.; Bible, K.C. Chaetocin: A promising new antimyeloma agent with in vitro and in vivo activity mediated via imposition of oxidative stress. Blood 2007, 109, 2579–2588. [Google Scholar] [CrossRef] [Green Version]
- Isham, C.R.; Tibodeau, J.D.; Bossou, A.R.; Merchan, J.R.; Bible, K.C. The anticancer effects of chaetocin are independent of programmed cell death and hypoxia, and are associated with inhibition of endothelial cell proliferation. Br. J. Cancer 2012, 106, 314–323. [Google Scholar] [CrossRef] [Green Version]
- Yu, L.; Ding, W.; Wang, Q.; Ma, Z.; Xu, X.; Zhao, X.; Chen, Z. Induction of cryptic bioactive 2,5-diketopiperazines in fungus Penicillium sp. DT-F29 by microbial co-culture. Tetrahedron 2017, 73, 907–914. [Google Scholar] [CrossRef]
- Allen, J.D.; van Loevezijn, A.; Lakhai, J.M.; van der Valk, M.; van Tellingen, O.; Reid, G.; Schellens, J.H.M.; Koomen, G.J.; Schinkel, A.H. Potent and specific inhibition of the breast cancer resistance protein multidrug transporter in vitro and in mouse intestine by a novel analogue of Fumitremorgin C. Mol. Cancer Ther. 2002, 1, 417–425. [Google Scholar]
- Raju, R.; Piggott, A.M.; Huang, X.C.; Capon, R.J. Nocardioazines: A novel bridged diketopiperazine scaffold from a marine-derived bacterium inhibits P-glycoprotein. Org. Lett. 2011, 13, 2770–2773. [Google Scholar] [CrossRef]
- Sohtome, Y.; Shimazu, T.; Barjau, J.; Fujishiro, S.; Akakabe, M.; Terayama, N.; Dodo, K.; Ito, A.; Yoshida, M.; Shinkai, Y.; et al. Unveiling epidithiodiketopiperazine as a non-histone arginine methyltransferase inhibitor by chemical protein methylome analyses. ChemComm 2018, 54, 9202–9205. [Google Scholar] [CrossRef] [Green Version]
- Yanagihara, M.; Sasaki-Takahasi, N.; Sugahara, T.; Yamamoto, S.; Shinomi, M.; Yamashita, L.; Hayashida, M.; Yamanoha, B.; Numata, A.; Yamori, T.; et al. Leptosins isolated from marine fungus Leptoshaeria species inhibit DNA topoisomerases I and/or II and induce apoptosis by inactivation of Akt/protein kinase B. Cancer Sci. 2005, 96, 816–824. [Google Scholar] [CrossRef]
- Minelli, A.; Bellezza, I.; Grotelli, S.l.; Galli, F. Focus on cyclo(His-Pro): History and perspectives as antioxidant peptide. Amino Acids 2008, 35, 283–289. [Google Scholar] [CrossRef]
- Peterkofsky, A.; Battaini, F. The biological activities of the neuropeptide histydyl-proline diketopiperaine. Neuropeptides 1980, 1, 105–118. [Google Scholar] [CrossRef]
- Bellezza, I.; Peirce, M.J.; Minelli, A. Cyclic dipeptides: From bugs to brain. Trends Mol. Med. 2014, 20, 551–558. [Google Scholar] [CrossRef]
- Turkez, H.; Cacciatore, I.; Arslan, M.E.; Fornasari, E.; Marinelli, L.; Di Stefano, A.; Mardinoglu, A. Histidyl-proline diketopiperazine isomers as multipotent anti-Alzheimer drug candidates. Biomolecules 2020, 10, 737. [Google Scholar] [CrossRef] [PubMed]
- Marinelli, L.; Fornasari, E.; Di Stefano, A.; Turkez, H.; Arslan, M.E.; Eusepi, P.; Ciulla, M.; Cacciatore, I. (R)-α-Lipoyl-Gly-l-Pro-l-Glu dimethyl ester as dual acting agent for the treatment of Alzheimer’s disease. Neuropeptides 2017, 66, 52–58. [Google Scholar] [CrossRef]
- Bellezza, I.; Peirce, M.J.; Minelli, A. Cyclic Peptides in Neurological Disorders: The Case of Cyclo(His-Pro); Elsevier Inc.: Amsterdam, The Netherlands, 2019. [Google Scholar]
- Guan, J. Insulin-like growth factor-1 and its derivatives: Potential pharmaceutical application for ischemic brain injury. Recent Pat. CNS Drug Discov. 2008, 3, 112–127. [Google Scholar] [CrossRef] [PubMed]
- Melo, I.S.; Candeia-Medeiros, N.; Ferro, J.N.S.; Cavalcante-Arau´jo, P.M.; Oliveira, T.L.; Santos, C.E.A.; Cardoso-Sousa, L.; Aguiar, E.M.G.; Wutke Oliveira, S.; Castro, O.W.; et al. Restoration of Cyclo-Gly-Pro-induced salivary hyposecretion and submandibular composition by naloxone in mice. PLoS ONE 2020, 15, e0229761. [Google Scholar] [CrossRef] [PubMed]
- Cogram, P.; Pilcher, J.; Glass, L.I. Bicyclic Compounds and Methods for Their Use in Treating Pitt Hopkins Syndrome. Patent WO/2021/080646, 24 April 2021. [Google Scholar]
- Fan, D.; Alamri, Y.; Liu, K.; MacAskill, M.; Harris, P.; Brimble, M.; Dalrymple-Alford, J.; Prickett, T.; Menzies, O.; Laurenson, A.; et al. Supplementation of blackcurrant anthocyanins increased cyclic-proline in the cerebrospinal fluid of Parkinson patients: Potential treatment to improve insulin-like growth factor-1 function. Nutrients 2018, 10, 714. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, F.; Liu, K.; Gray, C.; Harris, P.; Reynolds, C.M.; Vickers, M.H. Guan, Cyclic glycine-proline normalizes systolic blood pressure in high-fat diet-induded obese male rats. J. Nutr. Metab. Cardiovasc. Dis. 2020, 30, 339–346. [Google Scholar] [CrossRef]
- Brimble, M.A.; Guan, J. Cyclic G-2allylproline in Treatment of Parkinson’s Disease. U.S. Patent 7,776,876, 17 August 2010. [Google Scholar]
- Majumder, S.; Ghosh, A.; Chakraborty, S.; Saha, S.; Malay Bhattacharya, M. Metabolomics affirms traditional alcoholic beverage raksi as a remdy for high-altitude sickness. J. Ethnic Foods 2021, 8, 17–27. [Google Scholar] [CrossRef]
- Dubois, P.; Correia, I.; Le Chevalier, F.; Dubois, S.; Jacques, I.; Canu, N.; Moutiez, M.; Thai, R.; Gondry, M.; Lequin, O.; et al. Reprogramming Escherichia coli for the production of prenylated indole diketopiperazine alkaloids. Sci. Rep. 2019, 9, 9208–9220. [Google Scholar] [CrossRef] [Green Version]
- Manabe, S.; Machida, H.; Aihara, Y.; Yasanuga, M.; Itoa, Y.; Matsumura, Y. Development of a diketopiperazine-forming dipeptidyl Gly-Pro spacer for preparation of an antibody-drug conjugate. Med. Chem. Commun. 2013, 4, 792–796. [Google Scholar] [CrossRef]
- Hossain, M.A. Novel class of natural product-based opioid antagonists. In Proceedings of the AAAS Annual Meeting, Austin, TX, USA, 17 February 2018. [Google Scholar]
- Kilian, G.; Jamie, H.; Brauns, S.C.A.; Dyason, K.; Milne, P.J. Biological activity of selected tyrosine-containing 2,5-diketopiperazines. Pharmazie 2005, 60, 305–309. [Google Scholar]
- Balboni, G.; Guerrini, R.; Salvadori, S.; Tomatis, R.; Bryant, S.D.; Bianchi, C.; Attila, M.; Lazarus, L.H. Opioid diketopiperaines: Synthesis and activity of a prototypic class of opioid antagonists. Biol. Chem. 1997, 378, 19–29. [Google Scholar] [CrossRef]
- Copmans, D.; Crawford, A.; DeWitte, P.; Esguerra, C.; Kildgaard, S.; Ostenfeld, L. Treatment of Epilepsy with Plinabulin, Halimide or Diketopiperazine Derivatives. Patent W02019043012, 7 March 2019. [Google Scholar]
- Blazquez-Sanchez, M.T.; de Matos, A.M.; Rauter, A.P. Exploring anti-prion glyco-based and aromatic scaffolds: A chemical strategy for the quality of life. Molecules 2017, 22, 864. [Google Scholar] [CrossRef]
- Teixidó, M.; Zurita, E.; Malakoutikhah, M.; Tarragó, T.; Giralt, E. Diketopiperazines as a tool for the study of transport across the blood-brain barrier (BBB) and their potential use as BBB-Shuttles. J. Am. Chem. Soc. 2007, 129, 11802–11813. [Google Scholar] [CrossRef] [PubMed]
- Virgone-Carlotta, A.; Dufour, E.; Bacot, S.; Ahmadi, M.; Cornou, M.; Moni, L.; Garcia, J.; Chierici, S.; Garin, D.; Marti-Batlle, D.; et al. New diketopiperazines as vectors for peptide protection and brain delivery: Synthesis and biological evaluation. J. Label. Compd. Radiopharm. 2016, 59, 517–530. [Google Scholar] [CrossRef] [PubMed]
- Sun, S.; Ma, K.; Tao, Q.; Han, J.; Bao, L.; Liu, L.; Liu, H. Diketopiperazines and 2H-pyran-2-ones with antioxidant activity from the rice fermented with Aspegillus luchuensis. Fitoterapia 2018, 125, 266–272. [Google Scholar] [CrossRef]
- Zhong, W.; Wang, J.; Wei, X.; Fu, T.; Chen, Y.; Zeng, Q.; Huang, Z.; Huang, X.; Zhang, W.; Zhang, S.; et al. Three pairs of new spirocyclic alkaloid enantiomers from the marine-derived fungus Eurotium sp. SCSIO F452. Front. Chem. 2019, 7, 350–360. [Google Scholar] [CrossRef]
- Santoso, B.; Supardjan, A.M. Radical Scavenging Activity of 3,6-bis[(4-hydroxy-3-methoxyphenyl)metiliden]piperazine-2,5-dione and 3,6-bis[(4-hydroxy-3,5-dimethylphenyl)metiliden]piperazine-2,5-dione on 2,2-diphenyl-1-picrylhydrazyl (DPPH). In Proceedings of the Seminar Nasional Eight Star Performance Pharmacist, Faculty of Pharmacy, Gadjah Mada University, Yogyakarta, Indonesia, 27 December 2010. [Google Scholar]
- Manchineella, S.; Voshavar, C.; Govindaraju, T. Scavenging Antioxidant Cyclic Dipeptides and Silk Fibroin Biomaterials. Eur. J. Org. Chem. 2017, 2017, 4363–4369. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.N.; Lankalapalli, R.S.; Kumar, B.S. In vitro antibacterial screening of six proline-based cyclic dipeptides in combination with β-lactam antibiotics against medically important bacteria. Appl. Biochem. Biotechnol. 2014, 173, 116–128. [Google Scholar] [CrossRef]
- Gudasheva, T.A.; Voronina, T.A.; Ostrovstaya, R.U.; Rozantsev, G.G.; Vasilevich, N.I.; Trofimov, S.S.; Kravshenko, E.V.; Skoldinov, A.R.; Seredenin, S.B. Synthesis and antiamnesic activity of a series of N-acylprolyl-containing dipeptides. Eur. J. Med. Chem. 1996, 31, 151–157. [Google Scholar] [CrossRef]
- Romanova, G.A.; Mirzoav, T.K.; Barskov, I.V.; Victorov, I.V.; Gudasheva, T.A.; Ostrovskaya, R.U. Antiamnesic effect of acylprolyl-containing dipeptide (GVS-111) in compression-induced damage to frontal cortex. Bull. Exp. Biol. Med. 2000, 130, 846–848. [Google Scholar] [CrossRef] [PubMed]
- Gu, Q.; Fu, L.; Wang, Y.; Lin, J. Identification and characterization of extracellular cyclic dipeptides as quorum-sensing signal molecules from Shewanella baltica, the specific spoilage organism of Pseudoscianena crocea during 4 °C storage. J. Agric. Food Chem. 2013, 61, 11645–11652. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, S.R.; Banerjee, S. Gut microbiota in neurodegenerative disorders. J. Neuroimmun. 2019, 328, 98–104. [Google Scholar] [CrossRef]
- Bolognesi, M.L.; Ai Tran, H.N.; Staderini, M.; Monaco, A.; López-Cobeñas, A.; Bongarzone, S.; Biarnés, X.; López-Alvarado, P.; Cabezas, N.; Caramelli, M.; et al. Discovery of a class of diketopiperazines as antiprion compounds. ChemMedChem 2010, 5, 1324–1334. [Google Scholar] [CrossRef]
- Minelli, A.; Grottelli, S.; Mierla, A.; Pinnen, F.; Cacciatore, I. Cyclo(His_pro) exerts anti-inflammatory effects by modulating NF-kB and Nrf2 signalling. Intern. J. Biochem. Cell Biol. 2012, 44, 525–535. [Google Scholar] [CrossRef]
- O’Neill, J. Tackling Drug-Resistant infections Globally: Final Report and Recommendations. Rev. Antimicrob. Resist. 2016, 23, 590. [Google Scholar]
- Kamiya, T.; Maeno, S.; Hashimoto, M.; Mine, Y. Bicyclomycin, a new antibiotic. II. Structural elucidation and acyl derivatives. J. Antibiot. 1972, 25, 576–581. [Google Scholar] [CrossRef] [Green Version]
- Williams, R.M.; Durham, C.A. Bicyclomycin: Synthetic, mechanistic, and biological studies. Chem. Rev. 1988, 88, 511–540. [Google Scholar] [CrossRef]
- De Carvalho, M.P.; Abraham, W.-R. Antimicrobial and biofilm inhibiting diketopiperazines. Curr. Med. Chem. 2012, 19, 3564–3577. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Lee, B.C.; Kim, T.S.; Bae, K.S.; Hong, J.; Ho Choi, S.; Bao, B.; Jung, J.H. Bioactive Cyclic Dipeptides from a Marine Sponge-Associated Bacterium, Psychrobacter sp. Biomol. Ther. 2008, 16, 356–363. [Google Scholar] [CrossRef] [Green Version]
- Rhee, K.H. Isolation and characterization of Streptomyces sp. KH-614 producing anti-VRE (vancomycin-resistant enterococci) antibiotics. J. Gen. Appl. Microbiol. 2002, 48, 321–327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rhee, K.H. Cyclic dipeptides exhibit synergistic, broad spectrum antimicrobial effects and have anti-mutagenic properties. Int. J. Antimicrob. Agents 2004, 24, 423–427. [Google Scholar] [CrossRef] [PubMed]
- Ben Ameur Mehdi, R.; Shaaban, K.A.; Rebai, I.K.; Smaoui, S.; Bejar, S.; Mellouli, L. Five naturally bioactive molecules including two rhamnopyranoside derivatives isolated from the Streptomyces sp. Strain TN58. Nat. Prod. Res. 2009, 23, 1095–1107. [Google Scholar] [CrossRef]
- Vergne, C.; Boury-Esnault, N.; Perez, T.; Martin, M.T.; Adeline, M.T.; Tran Huu Dau, E.; Al-Mourabit, A. Verpacamides A-D, a sequence of C11N5 diketopiperazines relating cyclo(Pro-Pro) to cyclo(Pro-Arg), from the marine sponge Axinella vaceleti: Possible biogenetic precursors of pyrrole2-aminoimidazole alkaloids. Org. Lett. 2006, 8, 2421–2424. [Google Scholar] [CrossRef]
- Furtadoa, N.A.J.C.; Pupoa, M.T.; Carvalhoa, I.; Campoa, V.L.; Duarteb, M.C.T.; Bastos, J.K. Diketopiperazines produced by an Aspergillus fumigatus Brazilian strain. J. Braz. Chem. Soc. 2005, 16, 1448–1453. [Google Scholar] [CrossRef]
- Huberman, L.; Gollop, N.; Mumcuoglu, K.Y.; Breuer, E.; Bhusare, S.R.; Shai, Y.; Galun, R. Antibacterial substances of low molecular weight isolated from the blowfly, Lucilia sericata. Med. Vet. Entomol. 2007, 21, 127–131. [Google Scholar] [CrossRef]
- Fdhila, F.; Vazquez, V.; Sanchez, J.L.; Riguera, R. DD-Diketopiperazines: Antibiotics active against Vibrio anguillarum isolated from marine bacteria associated with cultures of Pecten maximus. J. Nat. Prod. 2003, 66, 1299–1301. [Google Scholar] [CrossRef] [PubMed]
- Zhao, K.; Xing, R.; Yan, X. Cyclic dipeptides: Biological activities and self-assembled materials. Peptide Sci. 2021, 113, 24202–24215. [Google Scholar] [CrossRef]
- Shaala, L.A.; Youssef, D.T.A.; Badr, J.M.; Harakeh, S.M.; Genta-Jouve, G. Bioactive DKPs and nucleoside derivatives from a sponge-derived Streptomyces species. Mar. Drugs 2019, 16, 584. [Google Scholar] [CrossRef] [Green Version]
- Carrieri, R.; Borriello, G.; Piccirillo, G.; Lahoz, E.; Sorrentino, R.; Cermola, M.; Bolletti Censi, S.; Grauso, L.; Mangoni, A.; Vinale, F. Antibiotic activity of a Paraphaeosphaeria sporulosa—Produced diketopiperazine against Salmonella enterica. J. Fungi 2020, 6, 83. [Google Scholar] [CrossRef] [PubMed]
- Haoran, M.; Fuqian, W.; Xiaoqi, J.; Jie, J.; Song, H.; Lu, C.; Geng, Z. A new DKP from an endophytic fungus Aspergillus aculeatus FO27. Nat. Prod. Res. 2021, 35, 2370–2375. [Google Scholar]
- Withers, H.; Swift, S.; Williams, P. Quorum sensing as an integral component of gene regulatory networks in Gram-negative bacteria. Curr. Opin. Microbiol. 2001, 4, 186–193. [Google Scholar] [CrossRef]
- Abraham, W.R. Controlling Gram-negative pathogenic bacteria by interfering with their biofilm formation. Drug Des. Rev. 2005, 2, 13–33. [Google Scholar] [CrossRef]
- Yu, X.; Li, L.; Sun, S.; Chang, A.; Dai, X.; Li, H.; Wang, Y.; Zhu, H. A cyclic dipeptide from marine fungus Penicillium chrysogenum DXY-1 exhibits anti-quorum sensing activity. ACS Omega 2021, 6, 7693–7700. [Google Scholar] [CrossRef]
- Verbeke, F.; De Craemer, S.; Debunne, N.; Janssens, Y.; Wynendaele, E.; Van de Wiele, C.; De Spiegeleer, B. Peptides as Quorum Sensing Molecules: Measurement Techniques and Obtained Levels In vitro and In vivo. Front. Neurosci. 2017, 11, 183–201. [Google Scholar] [CrossRef] [Green Version]
- Buroni, S.; Scoffone, V.C.; Fumagalli, M.; Makarov, V.; Cagnone, M.; Trespidi, G.; De Rossi, E.; Forneris, F.; Riccardi, G.; Chiarelli, L.R. Investigating the Mechanism of Action of Diketopiperazines Inhibitors of the Burkholderia cenocepacia Quorum Sensing Synthase CepI: A Site-Directed Mutagenesis Study. Front. Pharmacol. 2018, 9, 836–845. [Google Scholar] [CrossRef] [PubMed]
- Degrassi, G.; Aguilar, C.; Bosco, M.; Zahariev, S.; Pongor, S.; Venturi, V. Plant growth-promoting Pseudomonas putida WCS358 produces and secretes four cyclic dipeptides: Cross-talk with quorum sensing bacterial sensors. Curr. Microbiol. 2002, 45, 250–254. [Google Scholar] [CrossRef] [PubMed]
- Park, D.K.; Lee, K.E.; Baek, C.H.; Kim, I.H.; Kwon, J.H.; Lee, W.K.; Lee, K.H.; Kim, B.S.; Choi, S.H.; Kim, K.S. Cyclo(Phe-Pro) modulates the expression of ompU in Vibrio spp. J. Bacteriol. 2006, 188, 2214–2221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Wang, W.; Xu, S.X.; Magarvey, N.A.; McCormick, J.K. Lactobacillus reuteri—Produced cyclic dipeptides quench agr-mediated expression of toxic shock syndrome toxin-1 in staphylococci. Proc. Natl. Acad. Sci. USA 2011, 108, 3360–3365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ortiz-Castro, R.; Díaz-Pérez, C.; Martínez-Trujillo, M.; del Río, R.E.; Campos-García, J.; López-Bucio, J. Transkingdom signaling based on bacterial cyclodipeptides with auxin activity in plants. Proc. Natl. Acad. Sci. USA 2011, 108, 7253–7258. [Google Scholar] [CrossRef] [Green Version]
- Sun, C.; Luo, Z.; Zhang, W.; Deng, Z.; Mobli, M.; Kobe, B.; Jia, X.; Qu, X. Molecular Basis of Regio- and Stereo-Specificity in Biosynthesis of Bacterial Heterodimeric Diketopiperazines. Nat. Commun. 2020, 11, 6251–6261. [Google Scholar] [CrossRef]
- Houston, D.R.; Synstad, B.; Ejsink, V.G.H.; Stark, M.J.; Eggleston, I.; Van Aalten, D.M.F. Structure-Based Exploration of Cyclic Dipeptide Chitinase Inhibitors. J. Med. Chem. 2004, 47, 5713–5720. [Google Scholar] [CrossRef] [Green Version]
- Kumar, N.; Mohandas, C.; Nambisan, B.; Kumar, D.R.; Lankalapalli, R.S. Isolation of proline-based cyclic dipeptides from Bacillus sp. N strain associated with rhabditid [corrected] entomopathogenic nematode and its antimicrobial properties. World J. Microbiol. Biotechnol. 2013, 29, 355–364. [Google Scholar] [CrossRef]
- Semon, B.A. Dietary cyclic dipeptides, apoptosis and psychiatric disorders: A hypothesis. Med. Hypotheses 2014, 82, 740–743. [Google Scholar] [CrossRef]
- Andersen, O.A.; Dixon, M.J.; Eggleston, I.M.; van Aalten, D.M.F. Natural product family 18 chitinase inhibitors. Nat. Prod. Rep. 2005, 22, 563–579. [Google Scholar] [CrossRef]
- Kumar, S.N.; Nambisan, B. Antifungal activity of DKPs and stilbenes against plant pathogenic fungi in vitro. Appl. Biochem. Biotechnol. 2014, 172, 741–754. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Xu, K.; Yuan, X.L.; Ren, G.W.; Wang, X.Q.; Li, W.; Deng, N.; Wang, X.F.; Zhang, P. Characterization of DKP heterodimers as potential chemical markers for discrimination of two dominant black aspergilli. Phytochemistry 2020, 176, 112399. [Google Scholar] [CrossRef]
- Houston, D.; Eggleston, I.; Synstad, B.; Eijsink, V.; Van Aalten, D. The cyclic dipeptide CI-4 [cyclo(I-Arg-d-Pro)] inhibits family 18 chitinases by sructural mimicry of a reaction intermediate. Biochem. J. 2002, 27, 23–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hawas, U.W.; El-Kassem, L.T.A. Anticancer and antiviral DKP produced by the red sea endophytic fungus Penicillium chrysogenum. Lett. Org. Chem. 2019, 16, 409–414. [Google Scholar] [CrossRef]
- Ding, Y.; Zhu, X.; Hao, L.; Zhao, M.; Hua, Q.; An, F. Bioactive indolyl DKPs from the marine derived endophytic Aspergillus versicolor DY180635. Mar. Drugs 2020, 18, 338. [Google Scholar] [CrossRef] [PubMed]
- Niu, S.; Liu, D.; Shao, Z.; Proksch, P.; Lin, W. Euytypellazines A-M, thiodiketopiperazine-type alkaloids from deep sea derived fungus Eutypella sp. MCCC 3A00281. RSC Adv. 2017, 7, 33580–33590. [Google Scholar] [CrossRef] [Green Version]
- Guo, H.; Sun, B.; Gao, H.; Chen, X.; Liu, S.; Yao, X.; Liu, X.; Che, Y. DKPs from the Cordyceps-colonizing fungus Epicoccum nigrum. J. Nat. Prod. 2009, 72, 2115–2119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwak, M.K.; Liu, R.; Kwon, J.O.; Kim, M.K.; Kim, A.H.; Kang, S.O. Cyclic dipeptides from lactic acid bacteria inhibit proliferation of the influenza A virus. J. Microbiol. 2013, 51, 836–843. [Google Scholar] [CrossRef]
- Ahmed, E.F.; Rateb, M.E.; El-Kassem, L.T.A.; Hawas, U.W. Anti-HCV protease of DKP produced by the Red Sea sponge-associated fungus Aspergillus versicolor. Appl. Biochem. Microbiol. 2017, 53, 101–106. [Google Scholar] [CrossRef] [Green Version]
- Lin, C.K.; Wang, Y.T.; Hung, E.M.; Yang, Y.L.; Lee, J.C.; Sheu, J.H.; Liaw, C.C. Butyrolactones and DKPs from marine microbes: Inhibition effects on Dengue virus type 2 replication. Planta Med. 2017, 83, 158–163. [Google Scholar]
- Gondry, M.; Sauguet, L.; Belin, P.; Thai, R.; Amouroux, R.; Tellier, C.; Tuphile, K.; Jacquet, M.; Braud, S.; Courcon, M.; et al. Cyclodipeptide synthases are a family of tRNA-dependent peptide bond-forming enzymes. Nat. Chem. Biol. 2009, 5, 414–420. [Google Scholar] [CrossRef] [PubMed]
- Mas, V.; Falco, A.; Brocal, I.; Perez, L.; Coll, J.M.; Estepa, A. Identification of selective inhibitors of VHSV from biased combinatorial libraries of N,N′-disubstituted 2,5-piperazinediones. Antiviral Res. 2006, 72, 107–115. [Google Scholar] [CrossRef]
- Ichikawa, S.; Matsuda, A. Chemistry and structure-activity relationship of antibacterial nucleoside natural products. Nucleic Acids Symp. Ser. 2008, 52, 77–78. [Google Scholar] [CrossRef] [Green Version]
- Hirano, S.; Ichikawa, S.; Matsuda, A. Design and synthesis of diketopiperazine and acyclic analogs related to the caprazamycins and liposidomycins as potential antibacterial agents. Bioorgan. Med. Chem. 2008, 16, 428–436. [Google Scholar] [CrossRef]
- Yao, T.; Liu, J.; Jin, E.; Liu, Z.; Li, H.; Che, Q.; Zhu, T.; Li, D.; Li, W. Expanding the structural diversity of drimentines by exploring the promiscuity of two N-methyltransferases. Science 2020, 23, 101323–101346. [Google Scholar] [CrossRef]
- Martínez-Luis, S.; Rodríguez, R.; Acevedo, L.; González, M.C.; Lira-Rocha, A.; Mata, R. Malbrancheamide, anew calmodulin inhibitor from the fungus Malbranchea aurantiaca. Tetrahedron 2006, 62, 1817–1822. [Google Scholar] [CrossRef]
- Walchshofer, N.; Sarciron, M.E.; Garnier, F.; Delatour, P.; Petavy, A.F. Paris. Anthelmintic activity of 3,6-dibenzyl-2,5-dioxopiperazine, cyclo(l-Phe-l-Phe). J. Amino Acids 1997, 12, 41–47. [Google Scholar] [CrossRef]
- Perez-Picaso, L.; Olivo, H.F.; Argotte-Ramos, R.; Rodriguez-Gutierrez, M.; Rios, M.Y. Linear and cyclic dipeptides with antimalarial acitivity. Bioorgan. Med. Chem. Let. 2012, 22, 7048–7051. [Google Scholar] [CrossRef]
- Ishiyama, A.; Hokari, R.; Nonaka, K.; Chiba, T.; Miura, H.; Otoguro, K.; Iwatsuki, M. Diatretol, an alpha-dioxo-diketopiperazine, is a potent in vitro and in vivo antimalarial. J. Antibiot. 2021, 74, 266–268. [Google Scholar] [CrossRef] [PubMed]
- Beghyn, T.B.; Charton, J.; Leroux, F.; Laconde, G.; Bourin, A.; Cos, P.; Maes, L.; Deprez, B. Drug to Genome to Drug: Discovery of New Antiplasmodial Compounds. J. Med. Chem. 2011, 54, 3222–3240. [Google Scholar] [CrossRef] [PubMed]
- Beghyn, T.B.; Charton, J.; Leroux, F.; Henninot, A.; Reboule, I.; Cos, P.; Maes, L.; Deprez, B. Drug-to-genome-to-drug, step 2: Reversing selectivity in a series of antiplasmodial compounds. J. Med. Chem. 2012, 55, 1274–1286. [Google Scholar] [CrossRef] [PubMed]
- Buedenbender, L.; Grkovic, T.; Duffy, S.; Kurtboke, D.I.; Avery, V.M.; Carroll, A.R. Naseseazine C, a new anti-plasmodial dimeric DKP drom a marine sediment derived Streptomyces sp. Tetrahedron Let. 2016, 57, 5893–5895. [Google Scholar] [CrossRef]
- Leesombun, A.; Iijima, M.; Pagmadulam, B.; Orkhon, B.; Doi, H.; Issiki, K.; Sawa, R.; Nihei, C.; Nishikawa, Y. Metacytofilin has potent anti-malarial activity. Parasitol. Intern. 2021, 81, 102267. [Google Scholar] [CrossRef]
- Zin, N.M.; Baba, M.S.; Zainal-Abidin, A.H.; Latip, J.; Mazlan, N.W.; Edrada-Ebel, R. Gancidin W, a potential low-toxicity antimalarial agent isolated from an endophytic Streptomyces SUK10. Drug Des. Dev. Ther. 2017, 11, 351–363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tunes, L.G.; Goncalves, V.N.; Bueno, D.N.; Zani, C.L.; Rosa, L.H.; Cota, B.B. Diketopiperazine alkaloids produced by the endophytic fungus Penicillium citrinum and evaluation of their antileishmanial activity. Afr. J. Microbiol. Res. 2019, 13, 562–567. [Google Scholar]
- Metwaly, A.M.; Ghoneim, M.M.; Musa, A. Two new antileishmanial DKP alkaloids from the endophytic fungus Trichosporum sp. Pharma Chem. 2015, 7, 322–327. [Google Scholar]
- Yan, P.S.; Song, Y.; Sakuno, E.; Nakajima, H.; Nakagawa, H.; Yabe, K. Cyclo (l-leucyl-l-prolyl) produced by Achromobacter xylosoxidans inhibits aflatoxin production by Aspergillus parasiticus. Appl. Environ. Microbiol. 2004, 70, 7466–7473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iimura, K.; Furukawa, T.; Yamamoto, T.; Negishi, L.; Suzuki, M.; Sakuda, S. The mode of action of cyclo(l-Ala-l-Pro) in inhibiting aflatoxin production of Aspergillus flavus. Toxins 2017, 12, 219. [Google Scholar] [CrossRef] [Green Version]
- Brandl, C.J.; Deber, C.M. Hypothesis about the function of membrane-buried proline residues in transport proteins. Proc. Natl. Acad. Sci. USA 1986, 83, 917–921. [Google Scholar] [CrossRef] [Green Version]
- Graz, M.; Hunt, A.; Jamie, H.; Grant, G.; Milne, P. Antimicrobial activity of selected cyclic dipeptides. Pharmazie 1999, 54, 772–775. [Google Scholar]
- Imamura, M.; Prasad, C. Cyclo (His-Pro) potentiates GABA/ethanol-mediated chloride uptake by neurosynaptosomes. Peptides 2003, 24, 445–448. [Google Scholar] [CrossRef]
- Maujean, T.; Girard, N.; Ganesan, A.; Gulea, M.; Bonnet, D. Three cheers for nitrogen: Aza-DKPs, the aza analogues of 2,5-diketopiperazines. RSC Adv. 2020, 10, 43358–43370. [Google Scholar] [CrossRef]
- Kumar, D.; Gupta, S.K.; Ganeshpurkar, A.; Singh, R.; Kumar, D.; Das, N.; Krishnamurthy, S.; Singh, S.K. Biological profiling of piperazinediones for the management of anxiety. Pharmacol. Biochem. Behav. 2019, 176, 63–71. [Google Scholar] [CrossRef] [PubMed]
- Seredenin, S.B.; Gudasheva, T.A.; Boiko, S.S.; Kovalev, G.I.; Voronin, M.|V.; Yarkova, M.A. Endogenous dipeptide cycloprolylglycine shows selective anxiolytic activity in animals with manifest fear reaction. Bull. Exp. Biol. Med. 2002, 133, 360–362. [Google Scholar] [CrossRef]
- Lopez-Rodrıguez, M.L.; Morcillo, M.J.; Fernandez, E.; Porras, E.; Orensanz, L.; Beneytez, M.E.; Manzanares, J.; Fuentes, J.A. Synthesis and structure-activity relationships of a new model of arylpiperazines. 5.1 Study of the physicochemical influence of the pharmacophore on 5-HT1A/r1-adrenergic receptor affinity: Synthesis of a new derivative with mixed 5-HT1A/D2 antagonist properties. J. Med. Chem. 2001, 44, 186–197. [Google Scholar]
- Wyatt, P.G.; Allen, M.J.; Borthwick, A.D.; Davies, D.E.; Exall, A.M.; Hatley, R.J.D.; Irving, W.R.; Livermore, D.G.; Miller, N.D.; Nerozzi, F.; et al. 2,5-Diketopiperazines as potent and selective oxytocin antagonists 1: Identification, stereochemistry and initial SAR. Bioorgan. Med. Chem. Lett. 2005, 15, 2579–2582. [Google Scholar] [CrossRef]
- Liddle, J. Substituted Diketopiperazines and Their Use as Oxytocin Antagonists. PCT Int. Appl. WO2005000840; A1 20050106. European Patent Specification EP164787B1, 6 January 2005. [Google Scholar]
- Brooks, D.P. Substituted Diketopiperazines for the Treatment of Benign Prostatic Hyperplasia. PCT Int. Appl. CODEN PIXXD2; WO 2005000311; A1 20050106, 6 January 2005. [Google Scholar]
- Borthwick, A.D.; Hatley, R.J.; Hickey, D.M.B.; Liddey, J.; Livermore, D.G.H.; Mason, A.M.; Miller, N.D.; Nerozzi, F.; Sollis, S.L.; Szardenings, A.K.; et al. Substituted Diketopiperazines as Oxytocin Antagonists. United States Patent Application Publication 12/ 412, 837, 30 October 2009. [Google Scholar]
- Borthwick, A.D.; Liddle, J. The design of orally bioavailable 2, 5-diketopiperazine oxytocin antagonists: From concept to clinical candidate for premature labor. Med. Res. Rev. 2011, 31, 576–604. [Google Scholar] [CrossRef]
- Waltenspühl, Y.; Schöppe, J.; Ehrenmann, J.; Kummer, L.; Plückthun, A. Crystal structure ofthe human oxytocin receptor. Sci. Adv. 2020, 6, 1–11. [Google Scholar] [CrossRef]
- Yang, Y.H.; Yang, D.S.; Li, X.J.; Mo, M.H.; Zhao, P.J. Antibacterial diketopiperazines from an endophytic fungus Bionectria sp. Y1085. J. Antibiot. 2019, 72, 752–758. [Google Scholar] [CrossRef]
- Borthwick, A.D. Oral oxytocin antagonists. J. Med. Chem. 2010, 53, 6525–6538. [Google Scholar] [CrossRef]
- Raja, U.; Garcia, E.; The, C.; Sun, D.; Collis, D.; Ghilchik, M.; King, P.; Korbonits, M.; Smyth, D. Inhibitory actions of DKPs within the thyroid gland, and their system-wide presence. Endocr. Abstr. 2019. [Google Scholar] [CrossRef]
- Gomes, N.G.M.; Pereira, R.B.; Andrade, P.B.; Valentao, P. Double the chemistry, double the fun: Structural diversity and biological activity of marine-derived diketopiperazine dimers. Mar. Drugs 2019, 17, 551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coward, R.M.; Carson, C.C. Tadalafil in the treatment of erectile dysfunction, Ther. Clin. Risk Manag. 2008, 4, 1315–1330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gong, B.; Ma, M.; Xie, W.; Yang, X.; Huang, Y.; Sun, T.; Luo, Y.; Huang, J. Direct comparison of tadalafil with sildenafil for the treatment of erectile dysfunction: A systematic review and meta-analysis. Int. Urol. Nephrol. 2017, 49, 1731–1740. [Google Scholar] [CrossRef]
- Peng, Z.; Yang, L.; Dong, Q.; Wei, Q.; Liu, L.; Yang, B. Efficacy and safety of Tadalafil once-a-day versus Tadalafil on-demand in patients with erectile dysfunction: A systematic review and meta-analyses. Urol. Int. 2017, 99, 343–352. [Google Scholar] [CrossRef] [PubMed]
- Washington, S.L., 3rd; Shindel, A.W. A once-daily dose of tadalafil for erectile dysfunction: Compliance and efficacy. Drug Des. Dev. 2010, 4, 159–171. [Google Scholar]
- Ventimiglia, E.; Capogrosso, P.; Montorsi, F.; Salonia, A. The safety of phosphodiesterase type 5 inhibitors for erectile dysfunction. Expert Opin. Drug Saf. 2016, 15, 141–152. [Google Scholar] [CrossRef] [PubMed]
- Curran, M.; Keating, G. Tadalafil. Drugs 2003, 63, 2203–2212. [Google Scholar] [CrossRef] [PubMed]
- Mieczkowski, A.; Speina, E.; Trzybinski, D.; Winiewska-Szajewska, M.; Winska, P.; Borsuk, E.M.; Podsiadła-Białoskórska, M.; Przygodzki, T.; Drabikowski, K.; Stanczyk, L.; et al. Diketopiperazine-Based, Flexible Tadalafil Analogues: Synthesis, Crystal Structures and Biological Activity Profile. Molecules 2021, 26, 794. [Google Scholar] [CrossRef] [PubMed]
- Afroz, S.; Ara, A.; Arif, M. Pak. Platelet activating factor antagonists. J. Pharm. Sci. 1998, 11, 5–12. [Google Scholar]
- Bate, C.; Kempster, S.; Williams, A. Platelet-activating factor antagonists protec amyloid-beta damaged neurons from microglia-mediated death. Neuropharmacology 2006, 51, 173–181. [Google Scholar] [CrossRef]
- Randazzo, A.; Bifulco, G.; Giannini, C.; Bucci, M.; Debitus, C.; Cirino, G.; Gomez-Paloma, L. (Halipeptins A and B: Two novel potent anti-inflammatory cyclic depsipeptides from the Vanuatu marine sponge Haliclona species. J. Am. Chem. Soc. 2001, 123, 10870–10876. [Google Scholar] [CrossRef] [PubMed]
- Bar-Or, D.; Curtis, C.G.; Nagaraja, K.R.R.; Thomas, G. Method of Using DKPs and Composition Containing Them. U.S. Patent 2012/0094918 A1, 11 April 2012. [Google Scholar]
- Song, M.K.; Bischoff, D.S.; Song, A.M.; Uyemura, K.; Yamaguchi, D.T. Metabolic relationship between diabetes and Alzheimer’s disease affected by Cyclo(His-Pro) plus zinc treatment. BBA Clin. 2017, 7, 41–54. [Google Scholar] [CrossRef] [PubMed]
- Seiyaku, T. Remedy for Inflammatiory Disease Containing Diketopiperazine Derivative. Patent JP2000327575, 28 November 2000. [Google Scholar]
- Begum Ahil, S.; Hira, K.; Shaik, A.B.; Pal, P.P.; Kulkarni, O.P.; Araya, H.; Fujimoto, Y. l-Proline-based-cyclic dipeptides from Pseudomonas sp. (ABS-36) inhibit pro-inflammatory cytokines and alleviate crystal-induced renal injury in mice. Int. Immunopharmacol. 2019, 73, 395–404. [Google Scholar] [CrossRef]
- Ferro, J.N.S.; de Aquino, F.L.T.; de Brito, R.G.; dos Santos, P.L.; Quintas, J.S.S.; de Souza, L.C.; de Araujo, A.F.; Diaz, B.L.; Lucca-Junior, W.; Quintas-Junior, L.J.; et al. Cyclo(Gly-Pro), a cyclic dipeptide, attenuates nociceptive behavior and inflammatory response in mice. Clin. Exp. Pharmacol. Physiol. 2015, 42, 1287–1295. [Google Scholar] [CrossRef]
- Deepak, K.G.K.; Kumari, S.; Malla, R.R. Marine cyclic dipeptide cyclo(L-Leu-L-Pro) protects normal breast epithelial cells from tBHP-induced oxidative damage by targeting CD151. Acrh. Breast Cancer 2021, 8, 162–173. [Google Scholar]
- Lee, W.; Mi-Ae, K.; InWha, P.; Sam, J.; MinKyun, N.; Jong-Sup, B. Novel direct factor Xa inhibitory compounds from Tenebrio molitor with anti-platelet aggregation activity. Food Chem. Toxicol. 2017, 109, 19–27. [Google Scholar] [CrossRef]
- Zhang, H.H.; Yu, W.Y.; Li, L.; Wu, F.; Chen, Q.; Yang, Y.; Yu, C.H. Protective effects of DKPs from Moslae Herba against influenza A virus –induced pulmonary inflammation via inhibition of viral replication and platelets aggregation. J. Ethnopharmacol. 2018, 215, 156–166. [Google Scholar] [CrossRef]
- Liu, Z.; Chen, Y.; Li, S.; Hu, C.; Liu, H.; Zhang, W. Indole DKP alkaloids from the deep-sea derived fungus Aspergillus sp. FS445. Nat. Prod.Res. 2021, 12, 1–9. [Google Scholar]
- Nuhoglu, Y.; Nuhoglu, C. Aminophylline for treating asthma and chronic obstructive pulmonary disease. Expert Rev. Respir. Med. 2008, 2, 305–313. [Google Scholar] [CrossRef]
- Milne, P.J.; Hunt, A.L.; Rostoll, K.; van der Walt, J.J.; Graz, C.J.M. The biological activity of selected cyclic dipeptides. J. Pharm. Pharmacol. 1998, 50, 1331–1335. [Google Scholar] [CrossRef]
- Cole, R.; Kirksey, J.; Moore, J.; Blankenship, B.; Diener, U.; Davis, N. Tremorgenic toxin from Penicillium veruculosum. Appl. Microbiol. 1972, 24, 248–250. [Google Scholar] [CrossRef] [PubMed]
- McCleland, K.; Milne, P.J.; Lucieto, F.R.; Frost, C.; Brauns, S.C.; Van De Venter, M.; Du Plessis, J.; Dyason, K. An investigation into the biological activity of the selected histidine-containing diketopiperazines cyclo(His-Phe) and cyclo(His-Tyr). J. Pharm. Pharmacol. 2004, 56, 1143–1153. [Google Scholar] [CrossRef]
- Jamie, H.; Kilian, G.; Dyason, K.; Milne, P.J. The effect of the isomers of cyclo(Trp-Pro) on heart and ion-channel activity. J. Pharm. Pharmacol. 2002, 54, 1659–1665. [Google Scholar] [CrossRef]
- Chen, L.; Magliano, D.J.; Zimmet, P.Z. The worldwide epidemiology of type 2 diabetes mellitus-present and future perspectives. Nat. Rev. Endocrinol. 2012, 8, 228–236. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Fact Sheet: Diabetes. 2020. Available online: https://www.who.int/news-room/fact-sheets/detail/diabetes (accessed on 20 April 2020).
- Song, M.K.; Hwang, I.K.; Rosenthal, M.J.; Harris, D.M.; Yamaguchi, D.T.; Yip, I.; Go, V.L.W. Anti-hyperglycemic activity of zinc plus cyclo (His-Pro) in genetically diabetic Goto-Kakizaki and aged rats. Exp. Biol. Med. 2003, 228, 1338–1345. [Google Scholar] [CrossRef] [Green Version]
- Kwon, O.S.; Park, S.H.; Yun, B.S.; Pyun, Y.R.; Kim, C.J. Cyclo(dehydroala-L-Leu), an α-glucosidase inhibitor from Penicillium sp. F7Q6l4. J. Antibiot. 2000, 53, 954–958. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ye, G.; Huang, C.; Li, J.; Chen, T.; Tang, J.; Liu, W.; Long, Y. Isolation, Structural Characterization and Antidiabetic Activity of New Diketopiperazine Alkaloids from Mangrove Endophytic Fungus Aspergillus sp. 16-5c. Mar. Drugs 2021, 19, 402. [Google Scholar] [CrossRef]
- Song, M.M.; Xie, Y.H.; Chen, W.H.; Hu, Y.W.; Zhao, K.; Liu, Y.H.; Huang, X.L.; Liu, Q.C.; Wang, J.F. Diketopiperazine and enterotoxin analogues from the mangrove derived-soil Streptomyces sp. SCSIO 41400 and their biological evaluation. Nat. Prod. Res. 2020, 1–8. [Google Scholar] [CrossRef]
- Wilber, J.F. Histidyl-Proline Diketopiperazine (cyclo(His-Pro)), a CNS-Active Pharmacologic Agent. U.S. Patent 5418218; USOO54,8218A, 23 May 1995. [Google Scholar]
- Smithkline Beecham Corp. Hemoregulatory Compounds. Patent WO9718214, 22 May 1997. C07487/04A61K. [Google Scholar]
- Guo, T.; Adang, A.E.; Dolle, R.E.; Dong, G.; Fitzpatrick, D.; Geng, P.; Ho, K.K.; Kultgen, S.G.; Liu, R.; McDonald, E.; et al. Small molecule biaryl FSH receptor agonists. Part 1: Lead discovery via encoded combinatorial synthesis. Bioorgan. Med. Chem. Lett. 2004, 14, 1713–1716. [Google Scholar] [CrossRef]
- Guo, T.; Adang, A.E.; Dong, G.; Fitzpatrick, D.; Geng, P.; Ho, K.K.; Jibilian, C.H.; Kultgen, S.G.; Liu, R.; McDonald, E.; et al. Small molecule biaryl FSH receptor agonists. Part 2: Lead optimization via parallel synthesis. Bioorgan. Med. Chem. Lett. 2004, 14, 1717–1720. [Google Scholar] [CrossRef] [PubMed]
- Abad, A.; Fernandez-Molina, J.V.; Bikandi, J.; Ramirez, A.; Margareto, J.; Sendino, J.; Hernando, F.L.; Ponton, J.; Garaizar, J.; Rementeria, A. What makes Aspergillus fumigatus a successful pathogen? Genes and molecules involved in invasive aspergillosis. Rev. Iberoam. Micol. 2010, 27, 155–182. [Google Scholar] [CrossRef] [PubMed]
- Inoue, S.; Takanari, J.; Abe, K.; Nagayama, A.; Ikeya, Y.; Kohda, N. Isolation and structure determination of a heat shock protein inducer, asparagus-derived proline-containing 3-alkyldiketopiperazines (Asparaprolines), from a standardized extract of Asparagus officinalis stem. Nat. Prod. Commun. 2020, 15, 1–7. [Google Scholar] [CrossRef]
- Campanini Maciel, O.M.; Napoleao Tavares, R.S.; Engracia Caluz, D.R.; Gaspar, L.R.; Debonsi, H.M. Photoprotective potential of metabolites isolated from algae-associated fungi Annulohypoxylon stygium. J. Photochem. Photobiol. B Biol. 2018, 178, 316–322. [Google Scholar] [CrossRef] [PubMed]
- Petsi, M.; Zografos, A.L. 2,5-DKP catalysts as activators of dioxygen in oxidative processes. ACS Catal. 2020, 10, 7093–7099. [Google Scholar] [CrossRef]
- Wang, Z.F.; Sun, Z.C.; Xiao, L.; Zhou, Y.M.; Du, F.Y. Herbicidal polyketides and diketopiperazine derivatives from Penicillium viridicatum. J. Agricult. Food Chem. 2019, 67, 14102–14109. [Google Scholar] [CrossRef]
- Wang, T.; Zhang, Y.R.; Liu, X.H.; Ge, S.; Zhu, Y.S. Strategy for the biosynthesis of short oligopeptides: Green and sustainable chemistry. Biomolecules 2019, 9, 733. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Wang, M.; Fu, Y.; Huang, P.; Kong, D.; Niu, G. Emgineered biosynthesis of thaxtomin phytotoxins. Crit. Rev. Biotechnol. 2020, 40, 1163–1171. [Google Scholar] [CrossRef]
- Esfandiari, E.; Shekari, F.; Shekari, F.; Esfandiari, M. The Effect of salt stress on antioxidant enzymes’ activity and lipid peroxidation on the wheat seedling. Not. Bot. Hortiagrobot. Cluj-Napoca 2007, 35, 48–56. [Google Scholar]
- Hasan, M.; Ahmad-Hamdani, M.S.; Rosli, A.M.; Hamdan, H. Bioherbicides: An eco-freindly tool for sustainable weed management. Plants 2021, 10, 1212. [Google Scholar] [CrossRef]
- Cecchini, N.M.; Monteoliva, M.I.; Alvarez, M.E. Proline dehydrogenase is a positive regulator of cell death in different kingdoms. Plant Signal. Behav. 2011, 6, 1195–1197. [Google Scholar] [CrossRef] [Green Version]
- Zarse, K.; Schmeisser, S.; Groth, M.; Priebe, S.; Beuster, G.; Kuhlow, D.; Ristow, M. impaired insulin/IGF1 signaling extends life span by promoting mitochondrial l-proline catabolism to induce a transient ROS signal. Cell Metab. 2012, 15, 451–465. [Google Scholar] [CrossRef] [Green Version]
- Liang, X.; Zhang, L.; Natarajan, S.K.; Becker, D.F. Proline mechanisms of stress survival. Antioxid. Redox Signal. 2013, 19, 998–1011. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, J.; Xu, W.; Song, H.; Liu, Y.; Zhang, J.; Wang, Q. Synthesis and antiviral/fungicidal/insecticidal activities study of novel chiral indole DKP derivatives containing acylhydrazone moiety. J. Agric. Food Chem. 2020, 68, 5555–5571. [Google Scholar] [CrossRef]
- De Guzman, F.S.; Glober, J.B. New diketopiperazine metabolites from the sclerotia of Aspergillus ochrace us. J. Nat. Prod. 1992, 55, 931–939. [Google Scholar] [CrossRef] [PubMed]
- Kimura, Y.; Sawada, A.; Kuramata, M.; Kusano, M.; Fujioka, S.; Kawano, T.; Shimada, A. Brevicompanine C, cyclo(d-Ile-l-Trp), and cyclo(d-Leu-l-Trp), plant growth regulators from Penicillium brevi—Compactum. J. Nat. Prod. 2005, 68, 237–239. [Google Scholar] [CrossRef]
- Panjla, A.; Qayoom, I.; Kumar, A.; Verma, S. Anionic diketopiperazine induces osteogenic differentiation and supports osteogenesis in a 3D cryogel microenvironment. Chem. Commun. 2021, 57, 7422–7425. [Google Scholar] [CrossRef] [PubMed]
- Collins, M.N.; Ren, G.; Young, K.S.; Rui, L.; Reis, P.; Oliveira, J.M. Scaffold Fabrication Technologies and Structure/Function Properties in Bone Tissue Engineering. Adv. Funct. Mater. 2021, 31, 2010609–2010631. [Google Scholar] [CrossRef]
- Ohte, S.; Shiokawa, T.; Koyama, N.; Katagiri, T.; Imada, C.; Tomoda, H. A new diketopiperazine-like inhibitor of bone morphogenetic protein-induced osteoblastic differentiation produced by marine-derived Aspergillus sp. BFM-0085. J. Antibiot. 2020, 73, 554–558. [Google Scholar] [CrossRef]
- Pollard, A.J. and Bijker, E.M. A guide to vaccinology: From basic principles to new developments. Nat. Rev. Immunol. 2021, 21, 83–100. [Google Scholar] [CrossRef]
- Purcell, A.W.; McCluskey, J.; Rossjohn, J. More than one reason to rethink the use of peptides in vaccine design. Nat. Rev. Drug Discov. 2007, 6, 404–414. [Google Scholar] [CrossRef] [PubMed]
- Skwarczynski, M.; Toth, I. Peptide-based synthetic vaccines. Chem. Sci. 2016, 7, 842–854. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cresswell, P.; Ackerman, A.L.; Giodini, A.; Peaper, D.R.; Wearsch, P.A. Mechanisms of MHC class I-restricted antigen processing and cross-presentation. Immunol. Rev. 2005, 207, 145–157. [Google Scholar] [CrossRef] [PubMed]
- Pulendran, B.; Arunachalam, P.S.; O’Hagan, D.T. Emerging concepts in the science of vaccine adjuvants. Nat. Rev. Drug Discov. 2021, 20, 454–475. [Google Scholar] [CrossRef] [PubMed]
- Azuar, A.; Li, Z.; Shibu, M.A.; Zhao, L.; Luo, Y.; Shalash, A.O.; Khalil, Z.G.; Capon, R.J.; Hussein, W.M.; Totoh, I.; et al. Poly(hydrophobic amino acid)-Based Self-Adjuvanting Nanoparticles for Group A Streptococcus Vaccine Delivery. J. Med. Chem. 2021, 64, 2648–2658. [Google Scholar] [CrossRef]
- Skwarczynski, M.; Zhao, G.; Boer, J.C.; Ozberk, V.; Azuar, A.; Gonzalez Cruz, J.; Kumar Giddam, A.; Khalil, Z.G.; Pandey, M.; Shibu, M.A.; et al. Poly(amino acids) as a potent self-adjuvanting delivery system for peptide-based nanovaccines. Sci. Adv. 2020, 6, eaax2285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bartlett, S.; Skwarczynski, M.; Toth, I. Lipids as Activators of Innate Immunity in Peptide Vaccine Delivery. Curr. Med. Chem. 2020, 27, 2887–2901. [Google Scholar] [CrossRef] [PubMed]
- Bashiri, S.; Koirala, P.; Toth, I.; Skwarczynski, M. Carbohydrate Immune Adjuvants in Subunit Vaccines. Pharmaceutics 2020, 12, 965. [Google Scholar] [CrossRef]
- Yang, J.R.; Luo, Y.; Shibu, M.A.; Toth, I.; Skwarczynski, M. Cell-Penetrating Peptides: Efficient Vectors for Vaccine Delivery. Curr. Drug Deliv. 2019, 16, 430–443. [Google Scholar] [CrossRef] [PubMed]
- Nevagi, R.J.; Skwarczynski, M.; Toth, I. Polymers for subunit vaccine delivery. Eur. Polym. J. 2019, 114, 397–410. [Google Scholar] [CrossRef]
- Zhao, L.; Skwarczynski, M.; Toth, I. Polyelectrolyte-Based Platforms for the Delivery of Peptides and Proteins. ACS Biomater. Sci. Eng. 2019, 5, 4937–4950. [Google Scholar] [CrossRef]
- Zhao, G.; Chandrudu, S.; Skwarczynski, M.; Toth, I. The application of self-assembled nanostructures in peptide-based subunit vaccine development. Eur. Polym. J. 2017, 93, 670–681. [Google Scholar] [CrossRef]
- Marasini, N.; Ghaffar, K.A.; Skwarczynski, M.; Toth, I. Liposomes as a Vaccine Delivery System. In Micro- and Nanotechnology in Vaccine Development; Skwarczynski, M., Toth, I., Eds.; William Andrew Inc.: Norwich, UK, 2017; pp. 221–239. [Google Scholar]
- Loughlin, W.A.; Marshall, R.L.; Carreiro, A.; Elson, K.E. Solution-phase combinatorial synthesis and evaluation of piperazine-2,5-dione derivatives. Bioorg. Med. Chem. Lett. 2000, 10, 91–94. [Google Scholar] [CrossRef]
- Robinson, J.A. Beta-hairpin peptidomimetics: Design, structures and biological activities. Acc. Chem. Res. 2008, 41, 1278–1288. [Google Scholar] [CrossRef]
- Lyons, B.; Kwan, A.H.; Truscott, R. Spontaneous cyclization of polypeptides with a penultimate Asp, Asn or isoAsp at the N-terminus and implications for cleavage by aminopeptidase. FEBS J. 2014, 281, 2945–2955. [Google Scholar] [CrossRef]
- Haabeth, O.A.W.; Lohmeyer, J.J.K.; Sallets, A.; Blake, T.R.; Sagiv-Barfi, I.; Czerwinski, D.K.; McCarthy, B.; Powell, A.E.; Wender, P.A.; Waymouth, R.M.; et al. An mRNA SARS-CoV-2 Vaccine Employing Charge-Altering Releasable Transporters with a TLR-9 Agonist Induces Neutralizing Antibodies and T Cell Memory. ACS Cent. Sci. 2021, 7, 1191–1204. [Google Scholar] [CrossRef] [PubMed]
- Shimonkevitz, R.; Thomas, G.; Sue Slone, D.; Craun, M.; Mains, C.; Bar-Or, D. A diketopiperazine fragment of human serum albumin modulates T-lymphocyte cytokine production through rap1. J. Trauma 2008, 64, 35–41. [Google Scholar] [CrossRef]
- Cole, B.; McGrath, B.; Salottolo, K. LMWF-5A for the Treatment of Severe Osteoarthritis of the Knee: Integrated Analysis of Safety and Efficacy. Orthopedics 2017, 41, e77–e83. [Google Scholar] [CrossRef] [Green Version]
- Madhu, C.; Voshavar, C.; Rajasekhar, K.; Govindaraju, T. Cyclic dipeptide based cel-penetrating peptidomimetics for effective DNA delivery. Org. Biomol. Chem. 2017, 15, 3170–3174. [Google Scholar] [CrossRef] [PubMed]
- Yamashita, H.; Kato, T.; Oba, M.; Misawa, T.; Hattori, T.; Ohoka, N.; Tanaka, M.; Naito, M.; Kurihara, M.; Demizu, Y. Development of a cell-penetrating peptide that exhibits responsive changes in its secondary structure in the cellular environment. Sci. Rep. 2016, 6, 33003–33012. [Google Scholar] [CrossRef] [Green Version]
- Kato, T.; Yamashita, H.; Misawa, T.; Nishida, K.; Kurihara, M.; Tanaka, M.; Demizu, Y.; Oba, M. Plasmid DNA delivery by arginine-rich cell-penetrating peptides containing unnatural amino acids. Bioorgan. Med. Chem. 2016, 24, 2681–2687. [Google Scholar] [CrossRef] [PubMed]
- Mendive-Tapia, L.; Wang, J.; Vendrell, M. Fluorescent cyclic peptides for cell imaging. Pept. Sci. 2021, 113, e24181. [Google Scholar] [CrossRef]
- Jingee, Y.; Yang, L.; Xiaoai, W.; Dong, W.; Yawei, S.; Jiqian, W.; Hai, X. Self-assembly of cyclic dipeptides and their fluorescent properties. Acta Chim. Sinica 2019, 77, 1279–1286. [Google Scholar]
- Xia, Y.; Su, Y.; Wang, Q.; Yang, C.; Tang, B.; Zhang, Y.; Tu, J.; Shen, J. Preparation, characterization, and pharmacodynamics of insulinloaded fumaryl diketopiperazine microparticle dry powder inhalation. Drug Deliv. 2019, 26, 650–660. [Google Scholar] [CrossRef] [PubMed]
- Frid, A.H.; Hirsch, L.J.; Menchior, A.R.; Morel, D.R.; Strauss, K.W. Worldwide injection technique questionnaire study: Injecting complications and the role of the professional. Mayo Clin. Proc. 2016, 91, 1224–12230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daboussi, F.; Leduc, S.; Maréchal, A.; Dubois, G.; Guyot, V.; Perez-Michaut, C.; Amato, A.; Falciatore, A.; Juillerat, A.; Beurdeley, M.; et al. Genome engineering empowers the diatom Phaeodactylum tricornutum for biotechnology. Nat. Commun. 2014, 5, 3831–3839. [Google Scholar] [CrossRef] [PubMed]
- Bonneure, E.; De Baets, A.; De Decker, S.; Van den Berge, K.; Clement, L.; Vyverman, W.; Mangelinckx, S. Altering the Sex Pheromone Cyclo(l-Pro-l-Pro) of the Diatom Seminavis robusta towards a Chemical Probe. Int. J. Mol. Sci. 2021, 22, 1037. [Google Scholar] [CrossRef] [PubMed]
- Frenkel, J.; Wess, C.; Vyverman, W.; Pohnert, G. Chiral separation of a DKP pheromone from marine diatoms using supercritical fluid chromatography. J. Chromatogr. B 2014, 951, 58–61. [Google Scholar] [CrossRef]
- Sittmann, J.; Bae, M.; Mevers, E.; Li, M.; Quinn, A.; Sriram, G.; Clardy, J.; Liu, Z. Bacterial DKPs stimulate diatom growth and lipid accumulation. Plant Physiol. 2021, 186, 1159–1170. [Google Scholar] [CrossRef] [PubMed]
- Manchineella, S.; Govindaraju, T. Molecular self-assembly of cyclic dipeptide derivatives and their applications. ChemPlusChem 2017, 82, 88–106. [Google Scholar] [CrossRef]
- Chibh, S.; Mishra, J.; Kour, A.; Chauha, V.S.; Panda, J.J. Recent advances in the fabrication and bio-medical applications of self-assembled dipeptide nanostructures. Nanomedicine 2021, 16, 139–163. [Google Scholar] [CrossRef]
- You, Y.; Xing, R.; Zou, Q.; Shi, F.; Yan, X. High-tolerance crystalline hydrogels formed from self-assembling cyclic dipeptide. Beilstein J. Nanotechnol. 2019, 10, 1894–1901. [Google Scholar] [CrossRef] [PubMed]
- Scarel, M.; Marchesan, S. Diketopiperazine gels: New horizons from the self-assembly of cyclic dipeptides. Molecules 2021, 26, 3376. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Tao, K.; Ji, W.; Makam, P.; Rencus-Lazar, S.; Gazit, E. Self-assembly of cyclic dipeptides: Platforms for functional materials. Protein Pept. Lett. 2020, 27, 688–697. [Google Scholar] [CrossRef]
- Petry, N.; Benakki, H.; Clot, E.; Retailleau, P.; Guenoun, F.; Asserar, F.; Sekkat, C.; Metro, T.X.; Martinez, J.; Lamaty, F. A mechanochemical approach to access the proline-proline diketopiperazine framework. Beilstein J. Org. Chem. 2017, 13, 2169–2178. [Google Scholar] [CrossRef]
- Corey, E.J. General Methods for the Construction of Complex Molecules. Pure Appl. Chem. 1967, 14, 19–38. [Google Scholar] [CrossRef]
- Desiraju, G. Supramolecular Synthons in Crystal Engineering—A New Organic Synthesis. Angew. Chem. Int. Ed. Engl. 1995, 34, 2311–2327. [Google Scholar] [CrossRef]
- Spackman, P.R.; Yu, J.; Morton, C.J.; Parker, M.W.; Bond, C.S.; Spackman, M.A.; Jayatilaka, D.; Thomas, S.P. Bridging crystal engineering and drug discovery bu ytilizing intermolceluar interactions and molecular shapes in crystals. Anghewandte 2019, 58, 16780–16784. [Google Scholar]
- Bojarska, J.; New, R.; Borowiecki, P.; Remko, M.; Breza, M.; Madura, I.D.; Fruzinski, A.; Pietrzak, A.; Wolf, W.M. The First Insight Into the Supramolecular System of d,lα-Difluoromethylornithine: A New Antiviral Perspective. Front. Chem. 2021, 9, 679776. [Google Scholar] [CrossRef]
- Bojarska, J. Short Peptides: On the Trail of Future Stem Cell-Based Regenerative Therapies. Int. J. Nutr. Sci. 2021, 6, 1046. [Google Scholar]
- Bojarska, J.; Remko, M.; Breza, M.; Madura, I.D.; Kaczmarek, K.; Zabrocki, J.; Wolf, W.M. A supramolecular approach to structure-based design with a focus on synthon hierarchy in ornithine-derived ligands: Review, synthesis, experimental and in silico studies. Molecules 2020, 25, 1135. [Google Scholar] [CrossRef] [Green Version]
- Bojarska, J.; Remko, M.; Madura, I.D.; Kaczmarek, K.; Zabrocki, J.; Wolf, W.M. Synthesis, Experimental and In Silico Studies of N-Fluorenylmethoxycarbonyl-O-Tert-Butyl-N-Methyltyrosine, Coupled with CSD Data: A Survey of Interactions in the Crystal Structures of Fmoc-Amino Acids. Acta Crystallogr. C 2020, 76, 328–345. [Google Scholar] [CrossRef] [Green Version]
- Bojarska, J.; Remko, M.; Wojciechowski, J.; Madura, I.; Kaczmarek, K.; Zabrocki, J.; Zimecki, M.; Wolf, W.M. Cyclic Tetrapeptides as Promising Scaffold for Innovative Therapeutic Agents: Synthesis, Crystallographic, Biological and In Silico Studies. Z. Kristallogr. 2020, 40, 1. [Google Scholar]
- Bojarska, J. Amino Acids and Short Peptides as Anti-Aging “Superfood”. Int. J. Nutr. Sci. 2020, 5, 1039–1044. [Google Scholar]
- Bojarska, J.; Remko, M.; Madura, I.D.; Wojciechowski, J.M.; Olczak, A.; Kaczmarek, K.; Zabrocki, J.; Wolf, W.M. Supramolecular Synthon Polymorphism in Modified Amino Acids. Structural, Conformational and Energy Landscapes of N-Benzoyl-2′-Hydroxy-3-Methylisovaline. J. Mol. Struct. 2019, 1190, 11–22. [Google Scholar] [CrossRef]
- Bojarska, J.; Kaczmarek, K.; Zabrocki, J.; Wolf, W.M. Amino Acids: Molecules of Life. Int. J. Nutr. Sci. 2019, 4, 1035–1037. [Google Scholar]
- Bojarska, J.; Kaczmarek, K.; Zabrocki, J.; Wolf, W.M. Supramolecular Synthons as Related to Cooperativity in Biocomplexes: Towards Design and Development of Oligopeptide-Based Modern Drugs and Cosmeceuticals. Nov. Approaches Drug Des. Dev. 2019, 129, 1–27. [Google Scholar]
- Bojarska, J.; Fruzinski, A.; Sieron, L.; Maniukiewicz, W. The First Insight into the Supramolecular Structures of Popular Drug Repaglinide: Focus on Intermolecular Interactions in Antidiabetic Agents. J. Mol. Struct. 2019, 1179, 411–420. [Google Scholar] [CrossRef]
- Bojarska, J.; Wolf, W.; Zabrocki, J.; Kaczmarek, K.; Remko, M. New Synthons in Supramolecular Chemistry of Short Biologically Active Peptides. Acta Cryst. Sect A 2019, 75, e588. [Google Scholar] [CrossRef]
- Bojarska, J.; Remko, M.; Fruzinski, A.; Maniukiewicz, W. The Experimental and Theoretical Landscape of a New Antiplatelet Drug Ticagrelor: Insight into Supramolecular Architecture Directed by C-H⋯F, π⋯π and C-H⋯π Interactions. J. Mol. Struct. 2018, 1154, 290–300. [Google Scholar] [CrossRef]
- Bojarska, J.; Kaczmarek, K.; Zabrocki, J.; Wolf, W.M. Supramolecular Chemistry of Modified Amino Acids and Short Peptides. In Advances in Organic Synthesis; Rahman, A., Ed.; Bentham Science Publishers Ltd.: Sharjah, United Arab Emirates, 2018; Volume 11, pp. 43–107. [Google Scholar]
- Bojarska, J.; Fruzinski, A.; Maniukiewicz, W. Quantifying Intermolecular Interactions in Solid State Indapamide and Other Popular Diuretic Drugs: Insights from Hirshfeld Surface Study. J. Mol. Struct. 2016, 1116, 22–29. [Google Scholar] [CrossRef]
- Bojarska, J.; Maniukiewicz, W. Investigation of Intermolecular Interactions in Finasteride Drug Crystals in View of X-ray and Hirshfeld Surface Analysis. J. Mol. Struct. 2015, 1099, 419–426. [Google Scholar] [CrossRef]
- Bojarska, J.; Maniukiewicz, W.; Fruzinski, A.; Sieron, L.; Remko, M. Captopril and its Dimer Captopril Disulfide: Comparative Structural and Conformational Studies. Acta Crystallogr. C 2015, 71, 199–203. [Google Scholar] [CrossRef] [PubMed]
- Bojarska, J.; Maniukiewicz, W.; Fruzinski, A.; Jedrzejczyk, M.; Wojciechowski, J.; Krawczyk, H. Structural and Spectroscopic Characterization and Hirshfeld Surface Analysis of Major Component of Antibiotic Mupirocin—Pseudomonic Acid A. J. Mol. Struct. 2014, 1076, 126–135. [Google Scholar] [CrossRef]
- Bojarska, J.; Maniukiewicz, W.; Sieron, L. Three New Olanzapine Structures: The Acetic Acid Monosolvate, and the Propan-2-Ol and Propan-2-One Hemisolvate Monohydrates. Acta Crystallogr. C 2013, 69, 781–786. [Google Scholar] [CrossRef] [PubMed]
- Bojarska, J.; Maniukiewicz, W.; Sieron, L.; Fruzinski, A.; Kopczacki, P.; Walczynski, K.; Remko, M. Novel Pseudopolymorph of the Active Metabolite of Perindopril. Acta Crystallogr. C 2012, 68, o341–o343. [Google Scholar] [CrossRef] [PubMed]
- Bojarska, J.; Maniukiewicz, W.; Sieron, L.; Kopczacki, P.; Walczynski, K.; Remko, M. Perindoprilat Monohydrate. Acta Crystallogr. C 2012, 68, o443–o446. [Google Scholar] [CrossRef] [PubMed]
- Remko, M.; Bojarska, J.; Jezko, L.; Olczak, A.; Maniukiewicz, W. Molecular structure of antihypertensive drug perindopril, its active metabolite perindoprilat and impurity F. J. Mol. Struct. 2013, 1036, 292–297. [Google Scholar] [CrossRef]
- Remko, M.; Bojarska, J.; Jezko, P.; Sieron, L.; Olczak, A.; Maniukiewicz, W. Crystal and Molecular Structure of Perindopril Erbumine Salt. J. Mol. Struct. 2011, 997, 103–109. [Google Scholar] [CrossRef]
- Remko, M.; Bojarska, J.; Remková, A.; Maniukiewicz, W. (Molecular Structure and Acidity of Captopril, Zofenopril and Their Metabolites Captopril Disulfide and Zofenoprilat. Comput. Theor. Chem. 2015, 1062, 50–55. [Google Scholar] [CrossRef]
- Olczak, A.; Główka, M.L.; Szczesio, M.; Bojarska, J.; Duax, W.L.; Burkhart, B.M.; Wawrzak, Z. Nonstoichiometric Complex of Gramicidin D with KI at 0.80 Å Resolution. Acta Crystallogr. D Biol. Cryst. 2007, 63, 319–327. [Google Scholar] [CrossRef]
- Olczak, A.; Główka, M.L.; Szczesio, M.; Bojarska, J.; Wawrzak, Z.; Duax, W.L. The First Crystal Structure of a Gramicidin Complex with Sodium: High-Resolution Study of a Nonstoichiometric Gramicidin D-NaI Complex. Acta Crystallogr. D Biol. Cryst. 2010, 66, 874–880. [Google Scholar] [CrossRef] [PubMed]
- Główka, M.; Olczak, A.; Bojarska, J.; Szczesio, M. Structural Puzzles of Complexed Gramicidins in Their Crystals. J. Pol. Chem. Soc. 2007, 61, 161–187. [Google Scholar]
- Główka, M.L.; Olczak, A.; Bojarska, J.; Szczesio, M.; Duax, W.L.; Burkhart, B.M.; Pangborn, W.A.; Langs, D.A.; Wawrzak, Z. Structure of Gramicidin D-RbCl Complex at Atomic Resolution from Low-Temperature Synchrotron Data: Interactions of Double-Stranded Gramicidin Channel Contents and Cations with Channel Wall. Acta Crystallogr. D Biol. Crystallogr. 2005, 61, 433–441. [Google Scholar] [CrossRef] [PubMed]
- Główka, M.; Olczak, A.; Bojarska, J.; Szczesio, M.; Duax, W.; Burkhart, B.; Pangborn, W.A.; Langs, D.A.; Li, N.; Wawrzak, Z. Ion Channels in Crystals of Gramicidin D Complex with RbCl. Atomic Resolution Low-Temperature Synchrotron X-ray Data. Acta Crystallogr. Sect. A Found. Crystallogr. 2004, 60, 165. [Google Scholar] [CrossRef]



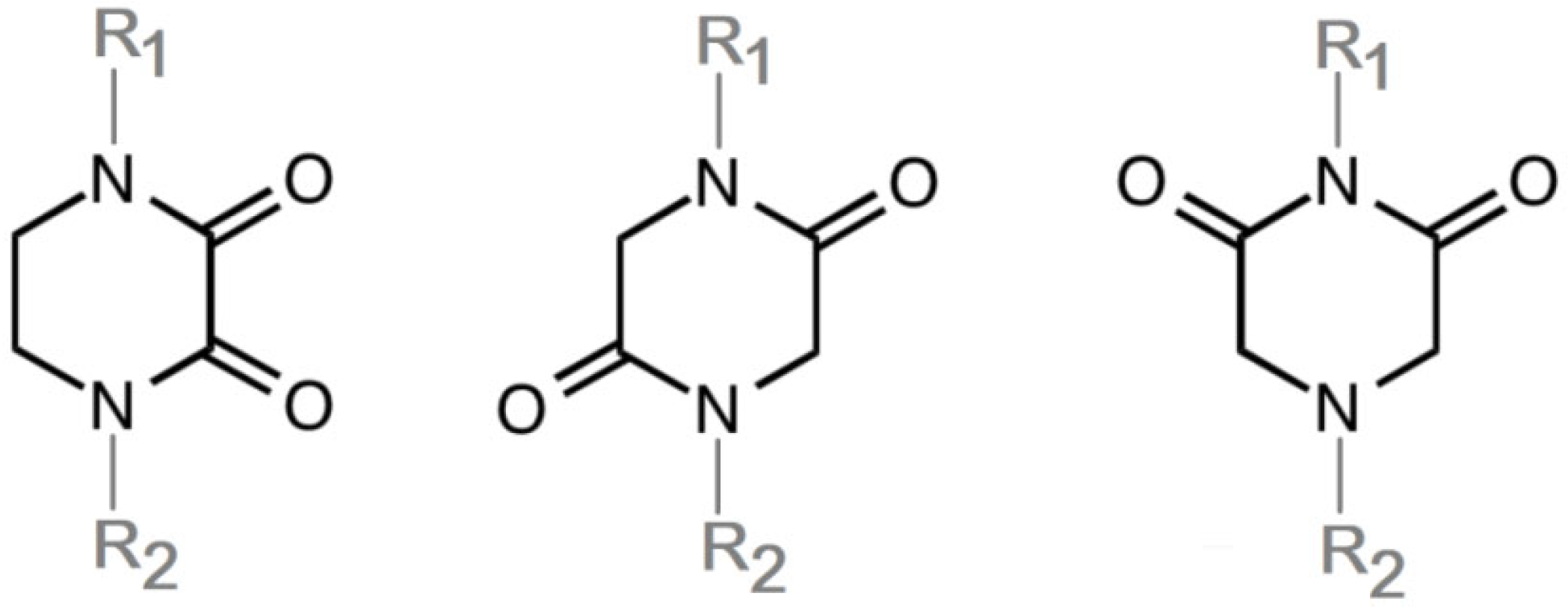{kind=link}
{kind=link}
| Entry | Structure | Name | Cell Line | Activity IC50 [μM] | Reference |
|---|---|---|---|---|---|
| 1. |  1 | cyclo(l-Phe-l-Hyp) | U87-MG | 5.8 ± 1.7 | [105] |
| U251 | 18.6 ± 0.1 | [105] | |||
| HCT-116 | >25 1 | [106] | |||
| OVCAR-8 | >25 1 | [106] | |||
| SF-295 | >25 1 | [106] | |||
| 2. |  2 | cyclo(l-Leu-l-Hyp) | U87-MG | 14.5 ± 1.6 | [105] |
| U251 | 29.4 ± 1.3 | [105] | |||
| 3. |  3 | cyclo(l-Leu-l-Pro) | U87-MG | 1.3 ± 0.1 | [105] |
| U251 | 19.8 ± 0.8 | [105] | |||
| ECA-109 | 55 2 | [107] | |||
| HeLa-S3 | 41 2 | [107] | |||
| PANC-1 | 14 2 | [107] | |||
| HCT-116 | 16 1 | [108] | |||
| HepG2 | ≥50 1 | [108] | |||
| MCF7 | 30 1 | [108] | |||
| 4. |  4 | cyclo(d-Leu-l-Pro) | ECA-109 | 44 2 | [107] |
| HeLa-S3 | 52 2 | [107] | |||
| PANC-1 | 55 2 | [107] | |||
| 5. |  5 | cyclo(l-Ile-l-Pro) | ECA-109 | 50 2 | [107] |
| HeLa-S3 | 45 2 | [107] | |||
| PANC-1 | 56 2 | [107] | |||
| HCT-116 | 22 1 | [108] | |||
| HepG2 | ≥50 1 | [108] | |||
| MCF7 | 27 1 | [108] | |||
| 6. |  6 | cyclo(l-Ile-l-Hyp) | ECA-109 | 54 2 | [107] |
| HeLa-S3 | 47 2 | [107] | |||
| PANC-1 | 42 2 | [107] | |||
| 7. |  7 | cyclo(4-S-hydroxy-d-Pro-d-Ile) | SF-268 | >295 3 | [109] |
| MCF-7 | 204 3 | [109] | |||
| H460 | 234 3 | [109] | |||
| HT-29 | 270 3 | [109] | |||
| CHO-K1 | >295 3 | [109] | |||
| 8. |  8 | cyclo(l-Phe-l-Pro) | HCT-116 | 21.4 1 | [106] |
| OVCAR-8 | 18.3 1 | [106] | |||
| SF-295 | 16.0 1 | [106] | |||
| ECA-109 | 42 2 | [107] | |||
| HeLa-S3 | 36 2 | [107] | |||
| PANC-1 | 50 2 | [107] | |||
| 9. |  9 | cyclo(l-Phe-d-Pro) | HCT-116 | 38.9 | [110] |
| HepG2 | ≥50 | [110] | |||
| MCF-7 | 102.0 | [110] | |||
| 10. |  10 | cyclo(d-Phe-d-Pro) | HCT-116 | 94.0 | [110] |
| HepG2 | ≥50 | [110] | |||
| MCF-7 | 114.0 | [110] | |||
| 11. |  11 | Penicillatide B | HCT-116 | 23.0 | [110] |
| HepG2 | ≥50 | [110] | |||
| MCF-7 | ≥50 | [110] | |||
| 12. |  12 | cyclo(l-Phe-2OH-d-Pro) | HCT-116 | 30 1 | [108] |
| HepG2 | ≥50 1 | [108] | |||
| MCF7 | 30 1 | [108] | |||
| 13. |  13 13 | cyclo(l-Val-l-Pro) | HeLa | 33.3 4 | [111] |
| K562 | n.a. | [111] | |||
| HL-60 | n.a. | [111] | |||
| BGC-823 | n.a. | [111] | |||
| MCF-7 | n.a. | [111] | |||
| 14. |  14 | Bacillusamide B | HCT-116 | 25 1 | [108] |
| HepG2 | ≥50 1 | [108] | |||
| MCF7 | 27 1 | [108] | |||
| 15. |  15 | Brevianamide F, cyclo(l-Trp-l-Pro) | HCT-116 | >25 1 | [106] |
| OVCAR-8 | 11.9 | [106] | |||
| SF-295 | >25 1 | [106] | |||
| 16. |  16 | cyclo(l-Trp-l-Hyp) | HL-60 | 64.34 | [112] |
| 17. |  17 | cyclo(d-Leu-2-OH-Pro) | HL-60 | 98.49 | [112] |
| 18. |  18 | Penicimutide | HeLa | 39.4 5 | [111] |
| K562 | n.a. | [111] | |||
| HL-60 | n.a. | [111] | |||
| BGC-823 | n.a. | [111] | |||
| MCF-7 | n.a. | [111] | |||
| 19. |  19 | cyclo(l-Pro-l-Pro) | ECA-109 | 30 2 | [107] |
| HeLa-S3 | 40 2 | [107] | |||
| PANC-1 | 34 2 | [107] |
| Entry | Structure | Name | Cell Line | Cytotoxic Effect | Reference |
|---|---|---|---|---|---|
| 1 |  21 | Tryprostatin A (Try A) | H520 | 80.1 ± 4.1 1, 79.4 ± 4.2 2 | [114] |
| MCF7 | >100 1, 95.0 ± 4.7 2 | [114] | |||
| PC-3 | 99.2 ± 4.2 1, 95.6 ± 5.0 2 | [114] | |||
| 2 |  22 | Tryprostatin B (Try B) | H520 | 77.6 ± 3.6 1, 60.5 ± 3.5 2 | [114] |
| MCF7 | 88.2 ± 5.8 1, 66.7 ± 5.3 2 | [114] | |||
| PC-3 | 95.5 ± 2.8 1, 68.9 ± 6.6 2 | [114] | |||
| 3 |  23 | ds2-TryB | H520 | 88.3 ± 8.4 1, 0.1 ± 0.1 2 | [114] |
| MCF7 | 73.6 ± 5.3 1, 0.0 ± 0.0 2 | [114] | |||
| PC-3 | 59.3 ± 3.9 1, 0.2 ± 0.0 2 | [114] | |||
| H520 | 11.9 3 | [115] | |||
| MCF7 | 17.0 3 | [115] | |||
| PC-3 | 12.3 3 | [115] | |||
| 4 |  24 | Piscarinin A | L929 | >50 4 | [116] |
| HeLa | >50 4 | [116] | |||
| LNCAP | 2.195 5 | [117] | |||
| 5 |  25 | Piscarinin B | L929 | >50 4 | [116] |
| HeLa | >50 4 | [116] | |||
| LNCAP | 1.914 5 | [117] | |||
| 6 |  26 | Notoamide C | HeLa | 50 5 | [118] |
| L1210 | 22 5 | [118] | |||
| 7 |  27 | Notoamide M | 22Rv1 | 55 6 | [119] |
| 8 |  28 | 17-O-ethylnotoamide M | 22Rv1 | 25 6 | [119] |
| 9 |  29 | Brevianamide W | P388 | n.a. | [120] |
| BEL-7402 | n.a. | [120] | |||
| MOLT | n.a. | [120] | |||
| 10 |  30 | Brevianamide Q | P388 | n.a. | [120] |
| BEL-7402 | n.a. | [120] | |||
| MOLT | n.a. | [120] | |||
| 11 |  31 | Brevianamide R | P388 | n.a. | [120] |
| BEL-7402 | n.a. | [120] | |||
| MOLT | n.a. | [120] | |||
| 12 |  32 | Brevianamide K | P388 | n.a. | [120] |
| BEL-7402 | n.a. | [120] | |||
| MOLT | n.a. | [120] | |||
| 13 |  33 | Brevianamide E | P388 | n.a. | [120] |
| BEL-7402 | n.a. | [120] | |||
| MOLT | n.a. | [120] |
| Entry | Structure | Name | Cell Line | IC50 [μM] | Reference |
|---|---|---|---|---|---|
| Tetracyclic | |||||
| 1. |  34 | Drimentine G | A2780 | 2.81 ± 0.09 | [121] |
| 1.38 ± 0.27 | [121] | ||||
| >10 | [121] | ||||
| 1.01 ± 0.04 | [121] | ||||
| 2.54 ± 0.18 | [121] | ||||
| 2. |  35 | Drimentine F | BGC-823 | >10 | [121] |
| A549 | >10 | [121] | |||
| A2780 | >10 | [121] | |||
| 3. |  36 | Drimentine H | Bel-7402 | >10 | [122] |
| BGC-823 | >10 | [122] | |||
| A549 | >10 | [122] | |||
| A2780 | >10 | [122] | |||
| 4. |  37 | Okaramine C | L5178Y | 14.7 | [123] |
| 5. |  38 | Okaramine G | L5178Y | 12.8 | [123] |
| 6. |  39–41 | Okaramine S (39) (R1 = R2 = prenyl) Okaramine T (40) (R1 = prenyl, R2 = H) Okaramine U (41) (R1 = R2 = H) | (for 39) | 0.78 22.4 | [124] |
| 7 |  42 | Roquefortine F | A-549 | 14.0 | [125] |
| HL-60 | 33.6 | [125] | |||
| BEL-7402 | 13.0 | [125] | |||
| MOLT-4 | 21.2 | [125] | |||
| 8. |  43 | Roquefortine G | A-549 | 42.5 | [125] |
| HL-60 | 36.6 | [125] | |||
| BEL-7402 | >50 | [125] | |||
| MOLT-4 | >50 | [125] | |||
| 9. |  44 | Fructigenine A | MCF-7 | >100, 63.1 1 | [126] |
| K562 | >100, 40.2 1 | [126] | |||
| HL-60 | >100, 47.7 1 | [126] | |||
| 10. |  45 | Fructigenine B | HeLa | >100, 26.6 1 | [126] |
| BGC-823 | >100, 35.4 1 | [126] | |||
| MCF-7 | >100, 39.6 1 | [126] | |||
| K562 | >100,49.9 1 | [126] | |||
| HL-60 | >100, 34.2 1 | [126] | |||
| 11. |  46 | Rugulosuvine A | HeLa | >100, 52.2 1 | [126] |
| BGC-823 | >100, 52.9 1 | [126] | |||
| MCF-7 | >100, 62.2 1 | [126] | |||
| K562 | >100, 75.6 1 | [126] | |||
| HL-60 | >100, 81.1 1 | [126] | |||
| 12. |  Penicimutanin A (47) | BGC-823 | 8.3, 88.0 1 | [126] | |
| MCF-7 | 7.3, 84.4 1 | [126] | |||
| K562 | 10.7, 87.4 1 | [126] | |||
| HL-60 | 6.1, 85.2 1 | [126] | |||
| 13. |  Penicimutanin C (48) | HeLa | 8.6, 88.1 1 | [126] | |
| BGC-823 | 8.7, 83.9 1 | [126] | |||
| MCF-7 | 6.0, 80.5 1 | [126] | |||
| K562 | 11.9, 87.7 1 | [126] | |||
| HL-60 | 5.0, 87.3 1 | [126] | |||
| 14. |  49 | Eurotiumin A | SF-268 | >100 | [127] |
| HepG2 | >100 | [127] | |||
| 15. |  50 | Eurotiumin B | SF-268 | >100 | [127] |
| HepG2 | >100 | [127] | |||
| Entry | Structure | Name | Cell Line | IC50 [μM] | Reference |
|---|---|---|---|---|---|
| Pentacyclic | |||||
| 1. |  51 | Spirotryprostatin A | tsFT210 | 197.5 | [128] |
| 2. |  52 | Spirotryprostatin B | tsFT210 | 14.0, 12.5 1 | [128,129] |
| K562 | 35 2 | [129] | |||
| HL-60 | 10 2 | [129] | |||
| 3Y1 | 14.0 | [130] | |||
| 3. |  53 | Spirotryprostatin L | HL-60 | 6.0 | [131] |
| 4. |  54 | - | HL-60 | 7.9 | [131] |
| 5. |  55 | Cyclotryprostatin B | MCF-7 | 5.1 | [131] |
| 6. |  56 | Cyclotryprostatin F | MCF-7 | 7.6 | [131] |
| 7. |  57 | Cyclotryprostatin G | MCF-7 | 10.8 | [131] |
| 8. |  58 | Cyclotryprostatin E | A549 | n.a. | [132] |
| A375 | n.a. | [132] | |||
| HeLa | n.a. | [132] | |||
| 9. |  59 | Versicamide G | HeLa | n.a. | [133] |
| HCT-116 | n.a. | [133] | |||
| HL-60 | n.a. | [133] | |||
| K562 | n.a. | [133] | |||
| Hexacyclic | |||||
| 10. |  60 | Versicamide A | HeLa | n.a. | [133] |
| HCT-116 | n.a. | [133] | |||
| HL-60 | n.a. | [133] | |||
| K562 | n.a. | [133] | |||
| 11. |  61 | Versicamide B | HeLa | n.a. | [133] |
| HCT-116 | n.a. | [133] | |||
| HL-60 | n.a. | [133] | |||
| K562 | n.a. | [133] | |||
| 12. |  62 | Versicamide C | HeLa | n.a. | [133] |
| HCT-116 | n.a. | [133] | |||
| HL-60 | n.a. | [133] | |||
| K562 | n.a. | [133] | |||
| 13. |  63 | Versicamide D | HeLa | n.a. | [133] |
| HCT-116 | n.a. | [133] | |||
| HL-60 | n.a. | [133] | |||
| K562 | n.a. | [133] | |||
| 14. |  64 | Versicamide E | HeLa | n.a. | [133] |
| HCT-116 | n.a. | [133] | |||
| HL-60 | n.a. | [133] | |||
| K562 | n.a. | [133] | |||
| 15. |  65 | Versicamide F | HeLa | n.a. | [133] |
| HCT-116 | n.a. | [133] | |||
| HL-60 | n.a. | [133] | |||
| K562 | n.a. | [133] | |||
| 16. |  66 | Versicamide H | HeLa | 19.4 | [133] |
| HCT-116 | 17.7 | [133] | |||
| HL-60 | 8.7 | [133] | |||
| K562 | 22.4 | [133] | |||
| 17. |  67 | Asperversiamide I | HeLa | 7.3 | [134] |
| 18. |  68 | Speramide B | PC3 | >40 | [135] |
| DU145 | >40 | [135] | |||
| LNCaP | >40 | [135] | |||
| Entry | Structure | Name | Cell Line | IC50 [μM] | Reference |
|---|---|---|---|---|---|
| Heptacyclic | |||||
| 1 |  Speramide A (69) | PC3 | >40 | [135] | |
| DU145 | >40 | [135] | |||
| LNCaP | >40 | [135] | |||
| 2 |  Stephacidin A (70) | PC3 | 2.1 | [136] | |
| LNCaP | 1 | [136] | |||
| A2780 | 4 | [136] | |||
| A2780/DDP | 6.8 | [136] | |||
| A2780/Tax | 3.6 | [136] | |||
| HCT116 | 2.1 | [136] | |||
| HCT116/mdr+ | 6.7 | [136] | |||
| HCT116/topo | 13.1 | [136] | |||
| MCF-7 | 4.2 | [136] | |||
| SKBR3 | 2.15 | [136] | |||
| LX-1 | 4.22 | [136] | |||
| 3 |  71 | Drimentine I | HeLa | 16.73 | [137] |
| A549 | n.a. | [137] | |||
| 4 |  72 | Gartryprostatin A | MV4-11 | 7.2 | [138] |
| K562 | > 10, 19.7 1 | [138] | |||
| HL-60 | > 10, 20.6 1 | [138] | |||
| A549 | > 10, 15.2 1 | [138] | |||
| 5 |  73 | Gartryprostatin B | MV4-11 | 10 | [138] |
| K562 | > 10, 18.5 1 | [138] | |||
| HL-60 | > 10, 23.2 1 | [138] | |||
| A549 | > 10, 18.2 1 | [138] | |||
| 6 |  74 | (+)-Avrainvillamide | HeLa | 1.1 2 | [87] |
| T-47D | 0.33 3 | [139] | |||
| LNCaP | 0.423 | [139] | |||
| NB4 | 1.1 | [140] | |||
| HL-60 | 0.643 | [140] | |||
| MV4-11 | 0.116 | [140] | |||
| OCI-AML3 | 0.112 | [140] | |||
| Molm-13 | 0.078 | [140] | |||
| 7 |  75 | (−)-Avrainvillamide | T-47D | 0.91 3 | [139] |
| LNCaP | 1.4 3 | [139] | |||
| Polycyclic | |||||
| 8 |  76 | Waikikiamide A | HT1080 | 0.519 | [141] |
| PC3 | 1.855 | [141] | |||
| Jurkat | 0.62 | [141] | |||
| A2780S | 0.78 | [141] | |||
| Dimeric | |||||
| 9 |  Waikikiamide B (77) | HT1080 | 1.135 | [141] | |
| PC3 | 1.805 | [141] | |||
| Jurkat | 1.79 | [141] | |||
| A2780S | 1.127 | [141] | |||
| 10 |  Stephacidin B (78) | PC3 | 0.37 | [136] | |
| LNCaP | 0.06 | [136] | |||
| A2780 | 0.33 | [136] | |||
| A2780/DDP | 0.43 | [136] | |||
| A2780/Tax | 0.26 | [136] | |||
| HCT116 | 0.46 | [136] | |||
| HCT116/mdr+ | 0.46 | [136] | |||
| HCT116/topo | 0.42 | [136] | |||
| MCF-7 | 0.27 | [136] | |||
| SKBR3 | 0.32 | [136] | |||
| LX-1 | 0.38 | [136] | |||
| 11 |  Naseseazine A (79) | AGS | n.a. | [142] | |
| SH-SY5Y | n.a. | [142] | |||
| TF-1 | n.a. | [142] | |||
| HT-29 | n.a. | [142] | |||
| 12 |  Naseseazine B (80) | AGS | n.a. | [142] | |
| SH-SY5Y | n.a. | [142] | |||
| TF-1 | n.a. | [142] | |||
| HT-29 | n.a. | [142] | |||
| 13 |  Asperflocin (81) | HT-29 | >20 | [143] | |
| A375 | 10.29 ± 2.37 | [143] | |||
| MCF-7 | >20 | [143] | |||
| HepG2 | >20 | [143] | |||
| 14 |  WIN 64821 (82) | HT-29 | >20 | [143] | |
| A375 | >20 | [143] | |||
| MCF-7 | >20 | [143] | |||
| HepG2 | >20 | [143] | |||
| Entry | Structure | Name | Cell Line | IC50 [μM] | Reference |
|---|---|---|---|---|---|
| Tricyclic | |||||
| 1. |  83 | Gliotoxin | SF-268 | 0.24 ± 0.10 | [144] |
| MCF-7 | 0.08 ± 0.0 | [144] | |||
| NCI-H460 | 0.24 ± 0.01 | [144] | |||
| HepG-2 | 0.21 ± 0.01 | [144] | |||
| P388 | 0.024 | [145] | |||
| HEK293 | 1.58 ± 0.03 | [146] | |||
| HCT-116 | 1.24 ± 0.38 | [146] | |||
| RKO | 0.80 ± 0.20 | [146] | |||
| tsFT210 | 0.15 1 | [147] | |||
| U937 | 0.20 ± 0.03 | [148] | |||
| PC-3 | 0.39 ± 0.03 | [148] | |||
| 2. |  84 | Acetylgliotoxin | SF-268 | 0.25 ± 0.03 | [144] |
| MCF-7 | 0.22 ± 0.04 | [144] | |||
| NCI-H460 | 0.32 ± 0.02 | [144] | |||
| HepG-2 | 0.49 ± 0.07 | [144] | |||
| HEK293 | 4.49 ± 0.24 | [146] | |||
| HCT-116 | 0.89 ± 0.04 | [146] | |||
| RKO | 1.24 ± 0.18 | [146] | |||
| 3. |  85 | 6-deoxy-5a,6-didehydrogliotoxin | SF-268 | 1.35 ± 0.05 | [144] |
| MCF-7 | 0.68 ± 0.02 | [144] | |||
| NCI-H460 | 1.27 ± 0.04 | [144] | |||
| HepG-2 | 1.52 ± 0.03 | [144] | |||
| P388 | 0.058 | [145] | |||
| 4. |  86 | bisdethiobis(methylthio)gliotoxin | SF-268 | >100 | [144] |
| MCF-7 | >100 | [144] | |||
| NCI-H460 | >100 | [144] | |||
| HepG-2 | >100 | [144] | |||
| P388 | 0.11 | [145] | |||
| HEK293 | 16.39 ± 0.38 | [146] | |||
| HCT-116 | 8.59 ± 0.96 | [146] | |||
| RKO | 10.32 ± 0.04 | [146] | |||
| U937 | 0.52 ± 0.07 | [148] | |||
| PC-3 | 15.87 ± 1.38 | [148] | |||
| B16 | 82 2 | [149] | |||
| 5. |  87 | 6-acetylbisdethiobis(methylthio)gliotoxin | SF-268 | 34.0 ± 3.6 | [144] |
| MCF-7 | 3.1 ± 0.10 | [144] | |||
| NCI-H460 | 5.4 ± 0.60 | [144] | |||
| HepG-2 | 7.0 ± 0.17 | [144] | |||
| HEK293 | >50 | [146] | |||
| HCT-116 | >50 | [146] | |||
| RKO | >50 | [146] | |||
| 6. |  88 | Dichotocejpin A | SF-268 | 35.7 ± 2.1 | [144] |
| MCF-7 | 29.5 ± 2.3 | [144] | |||
| NCI-H460 | >100 | [144] | |||
| HepG-2 | 28.9 ± 3.0 | [144] | |||
| 7. |  89 | Dichotocejpin B | SF-268 | >100 | [144] |
| MCF-7 | >100 | [144] | |||
| NCI-H460 | >100 | [144] | |||
| HepG-2 | >100 | [144] | |||
| 8. |  90 | bis(dethio)-10a-methylthio-3a-deoxy-3,3a-didehydrogliotoxin | P388 | 3.4 | [145] |
| 9. |  91 | bis(dethio)bis- (methylthio)-5a,6-didehydrogliotoxin | P388 | 0.11 | [145] |
| tsFT210 | 89 1 | [147] | |||
| U937 | >100 | [148] | |||
| PC-3 | >100 | [148] | |||
| B16 | 78 2 | [149] | |||
| 10. |  92 | 5a,6-didehydrogliotoxin | P388 | 0.056 | [145] |
| 11. |  93 | Gliotoxin G | P388 | 0.020 | [145] |
| 12. |  94 | reduced Gliotoxin | HEK293 | 1.26 ± 0.04 | [146] |
| HCT-116 | 0.43 ± 0.04 | [146] | |||
| RKO | 0.41 ± 0.07 | [146] | |||
| 13. |  95 | 6-deoxy-bis(dethio)bis- (methylthio)-5a,6-didehydrogliotoxin | HEK293 | >50 | [146] |
| HCT-116 | >50 | [146] | |||
| RKO | >50 | [146] | |||
| 14. |  96 | 3-dehydroxymethylbisdethio-3,10a-bis(methylthio)gliotoxin | B16 | 86 2 | [149] |
| 15. |  97 | Glionitrin A | HCT-116 | 0.82 | [150] |
| A549 | 0.55 | [150] | |||
| AGS | 0.45 | [150] | |||
| DU145 | 0.24 | [150] | |||
| MCF-7 | 2.0 | [150] | |||
| HepG2 | 2.3 | [150] | |||
| 16. |  98 | Glionitrin B | DU145 | n.a. | [151] |
| 17. |  99 | Phomazine B | HL-60 | >10 | [152] |
| HCT-116 | >10 | [152] | |||
| A549 | >10 | [152] | |||
| K562 | >10 | [152] | |||
| MGC-803 | 8.5 | [152] | |||
| tetracyclic | |||||
| 18. |  100 | Luteoalbusin A | SF-268 | 0.46 ± 0.05 | [154] |
| MCF-7 | 0.23 ± 0.03 | [154] | |||
| NCI-H460 | 1.15 ± 0.03 | [154] | |||
| HepG-2 | 0.91 ± 0.03 | [154] | |||
| 19. |  101 | Luteoalbusin B | SF-268 | 0.59 ± 0.03 | [154] |
| MCF-7 | 0.25 ± 0.00 | [154] | |||
| NCI-H460 | 1.31 ± 0.12 | [154] | |||
| HepG-2 | 1.29 ± 0.16 | [154] | |||
| 20. |  102 | T988A | SF-268 | 1.04 ± 0.03 | [154] |
| MCF-7 | 0.91 ± 0.03 | [154] | |||
| NCI-H460 | 5.60 ± 0.58 | [154] | |||
| HepG-2 | 3.52 ± 0.74 | [154] | |||
| 21. |  103 | Gliocladine C | SF-268 | 0.73 ± 0.05 | [154] |
| MCF-7 | 0.23 ± 0.03 | [154] | |||
| NCI-H460 | 6.57 ± 0.81 | [154] | |||
| HepG-2 | 0.53 ± 0.04 | [154] | |||
| 22. |  104 | Gliocladine D | SF-268 | 2.49 ± 0.07 | [154] |
| MCF-7 | 0.65 ± 0.07 | [154] | |||
| NCI-H460 | 17.78 ± 0.27 | [154] | |||
| HepG-2 | 2.03 ± 0.07 | [154] | |||
| 23. |  105 | Sporidesmin A | E006AA-hT | 2.5 | [155] |
| pentacyclic | |||||
| 24. |  106 | Brocazine A | Du145 | 4.2 | [156] |
| HeLa | 6.8 | [156] | |||
| HepG2 | 6.4 | [156] | |||
| MCF-7 | 5.5 | [156] | |||
| NCI-H460 | 4.9 | [156] | |||
| SGC-7901 | 2.6 | [156] | |||
| SW1990 | 6.0 | [156] | |||
| SW480 | 2.0 | [156] | |||
| U251 | 5.2 | [156] | |||
| 25. |  107 | Brocazine B | Du145 | 3.6 | [156] |
| HeLa | 5.3 | [156] | |||
| HepG2 | 5.5 | [156] | |||
| MCF-7 | 6.1 | [156] | |||
| NCI-H460 | 4.0 | [156] | |||
| SGC-7901 | 2.4 | [156] | |||
| SW1990 | 6.4 | [156] | |||
| SW480 | 1.2 | [156] | |||
| U251 | 3.5 | [156] | |||
| 26. |  108 | Brocazine C | Du145 | >20 | [156] |
| HeLa | >20 | [156] | |||
| HepG2 | >20 | [156] | |||
| MCF-7 | >20 | [156] | |||
| NCI-H460 | >20 | [156] | |||
| SGC-7901 | >20 | [156] | |||
| SW1990 | >20 | [156] | |||
| SW480 | >20 | [156] | |||
| U251 | >20 | [156] | |||
| 27. |  109 | Brocazine D | Du145 | >20 | [156] |
| HeLa | >20 | [156] | |||
| HepG2 | >20 | [156] | |||
| MCF-7 | >20 | [156] | |||
| NCI-H460 | >20 | [156] | |||
| SGC-7901 | >20 | [156] | |||
| SW1990 | >20 | [156] | |||
| SW480 | >20 | [156] | |||
| U251 | >20 | [156] | |||
| 28. |  110 | Brocazine E | Du145 | 11.2 | [156] |
| HeLa | 4.3 | [156] | |||
| HepG2 | 5.6 | [156] | |||
| MCF-7 | 9.0 | [156] | |||
| NCI-H460 | 12.4 | [156] | |||
| SGC-7901 | 3.3 | [156] | |||
| SW1990 | 2.1 | [156] | |||
| SW480 | n.t. | [156] | |||
| U251 | 6.1 | [156] | |||
| 29. |  111 | Brocazine F | Du145 | 1.7 | [156] |
| HeLa | 6.9 | [156] | |||
| HepG2 | 2.9 | [156] | |||
| MCF-7 | 3.0 | [156] | |||
| NCI-H460 | 0.89 | [156] | |||
| SGC-7901 | 8.0 | [156] | |||
| SW1990 | 5.9 | [156] | |||
| SW480 | n.t. | [156] | |||
| U251 | 5.3 | [156] | |||
| 31. |  112 | Epicorazine A | HL-60 | 0.05 | [152] |
| HCT-116 | 0.33 | [152] | |||
| A549 | 2.3 | [152] | |||
| K562 | 1.5 | [152] | |||
| MGC-803 | 2.7 | [152] | |||
| 32 |  113 | Epicorazine B | HL-60 | 1.6 | [152] |
| HCT-116 | >10 | [152] | |||
| A549 | >10 | [152] | |||
| K562 | 4.6 | [152] | |||
| MGC-803 | 5.2 | [152] | |||
| 33. |  114 | Epicorazine C | HL-60 | 3.5 | [152] |
| HCT-116 | 7.1 | [152] | |||
| A549 | >10 | [152] | |||
| K562 | >10 | [152] | |||
| MGC-803 | 3.4 | [152] | |||
| 34. |  115 | Exserohilone A | HL-60 | 3.5 | [152] |
| HCT-116 | 4.0 | [152] | |||
| A549 | >10 | [152] | |||
| K562 | >10 | [152] | |||
| MGC-803 | 4.2 | [152] | |||
| 35. |  116 | Brocazine G | A2780 | 0.664 | [157] |
| A2780 cisR | 0.661 | [157] | |||
| 36. |  117 | Spirobrocazine A | A2780 | n.a. | [157] |
| A2780 cisR | n.a. | [157] | |||
| 37. |  118 | Spirobrocazine B | A2780 | n.a. | [157] |
| A2780 cisR | n.a. | [157] | |||
| 38. |  119 | 5′-hydroxy-6′-ene-epicoccin G | Huh.7.5 | n.a. | [158] |
| 39. |  120 | 7-methoxy-7′-hydroxyepicoccin G | Huh.7.5 | n.a. | [158] |
| 40. |  121 | 8′-acetoxyepicoccin D | Huh.7.5 | n.a. | [158] |
| 41. |  122 | 7′-demethoxyrostratin C | Huh.7.5 | 9.52 | [158] |
| 42. |  123 | Epicoccin E | Huh.7.5 | n.a. | [158] |
| 43. |  124 | Epicoccin G | Huh.7.5 | n.a. | [158] |
| 44. |  125 | Rostracin C | Huh.7.5 | 4.88 | [158] |
| 45. |  126 | - | KB | >30.9 | [159] |
| BC-1 | 29.2 | [159] | |||
| NCI-H187 | 22.9 | [159] | |||
| 46. |  127 | - | KB | >31.0 | [159] |
| BC-1 | >31.0 | [159] | |||
| NCI-H187 | 20.3 | [159] | |||
| 47. |  128 | - | KB | >46.0 | [159] |
| BC-1 | >46.0 | [159] | |||
| NCI-H187 | >46.0 | [159] | |||
| dimeric | |||||
| 48. |  129 | Cristazine | HeLa | 0.5 | [160] |
| A431 | ~0.5 | [161] | |||
| 49. |  130 | Chaetocin | HL-60 | 0.153 ± 0.22 | [162] |
| U937 | 0.096 ± 0.18 | [162] | |||
| KG-1a | 0.103 ± 0.34 | [162] | |||
| 50. |  131 | Verticillin A | HT-29 | 0.02 | [163] |
| H460 | 0.20 | [163] | |||
| SF-268 | 0.25 | [163] | |||
| MCF-7 | 0.37 | [163] | |||
| MDAMB-435 | 0.070, 0.018 | [163,164] | |||
| MDA-MB-231 | 0.023 | [164] | |||
| OVCAR3 | 0.036 | [164] | |||
| AGS | 0.070 | [165] | |||
| HeLa | 0.319 | [165] | |||
| OVSAHO | 0.060 | [166] | |||
| OVCAR4 | 0.047 | [166] | |||
| OVCAR8 | 0.045 | [166] | |||
| 51. |  132 | Verticillin H | HT-29 | 0.04 | [163] |
| H460 | 0.30 | [163] | |||
| SF-268 | 0.33 | [163] | |||
| MCF-7 | 0.49 | [163] | |||
| MDAMB-435 | 0.10, 0.044 | [163,164] | |||
| MDA-MB-231 | 0.031 | [164] | |||
| OVCAR3 | 0.229 | [164] | |||
| 52. |  133 | Sch 52900 | HT-29 | 0.01 | [163] |
| H460 | 0.29 | [163] | |||
| SF-268 | 0.37 | [163] | |||
| MCF-7 | 0.58 | [163] | |||
| MDAMB-435 | 0.48 | [163] | |||
| 53. |  134 | Sch 52901 | HT-29 | 0.19 | [163] |
| H460 | 1.20 | [163] | |||
| SF-268 | 0.75 | [163] | |||
| MCF-7 | 1.11 | [163] | |||
| MDAMB-435 | 0.03 | [163] | |||
| 54. |  135 | Gliocladicillin A | HT-29 | n.t. | [163] |
| H460 | 0.03 | [163] | |||
| SF-268 | 0.09 | [163] | |||
| MCF-7 | 0.09 | [163] | |||
| MDAMB-435 | n.t. | [163] | |||
| 55. |  136 | Gliocladicillin C | HT-29 | 0.03 | [163] |
| H460 | 0.52 | [163] | |||
| SF-268 | 0.38 | [163] | |||
| MCF-7 | 0.61 | [163] | |||
| MDAMB-435 | 0.08 | [163] | |||
| 56. |  137 | 11′-Deoxyverticillin | HT-29 | n.t. | [163] |
| H460 | 0.01 | [163] | |||
| SF-268 | 0.04 | [163] | |||
| MCF-7 | 0.03 | [163] | |||
| MDAMB-435 | n.t. | [163] | |||
| HCT-116 | 0.030 1 | [167] | |||
| 57. |  138 | 11,11′-dideoxyverticillin | HCT-116 | 0.030 1 | [167] |
| MDAMB-468 | 0.281 ± 0.022 | [168] | |||
| MCF-7 | 0.158 ± 0.070 | [168] | |||
| MDA-MB-435 | 0.223 ± 0.099 | [168] | |||
| MDA-MB-231 | 0.138 ± 0.025 | [168] | |||
| Entry | Structure | Name | Bacterial Strains | MIC [μg/mL] | Reference |
|---|---|---|---|---|---|
| 1. |  | cyclo(l-Leu-l-Pro) | Enterococcus faecalis (VRE) Micrococcus luteus, Staphylococcus aureus | 12.5 | [214] |
| 2.9 mmol/L | [218] | ||||
| 2. |  | cyclo(l-Phe-l-Pro) | Micrococcus luteus, Staphylococcus aureus Staphylococcus aureus | 2.9 mmol/L | [218] |
| 10 mm zone of inhibition | [222] | ||||
| 3. |  | Combination of cyclo(l-Leu-l-Pro) and cyclo(l-Phe-l-Pro) | Enterococcus faecalis (VRE) Micrococcus luteus, Escherichia coli, Candida albicans, Staphylococcus aureus, Cryptococcus neoformans | 0.25–1 | [215] |
| 0.25–0.5 | [215] | ||||
| 4. |  | cyclo(l-Pro-l-Pro) | Pseudomonas aeruginosa | 150 AU/mL | [219] |
| Micrococcus luteus | 100 AU/mL | [219] | |||
| 5. |  | Avrainvillamide | Staphylococcus aureus | 12.5 | [87] |
| Streptococcus pyogenes | 12.5 | [87] | |||
| Enterococcus faecalis | 25 | [87] | |||
| 6. |  | Speramide A | Pseudomonas aeruginosa | 0.36 | [41] |
| 7. |  | Fumitremorgin C | Staphylococcus aureus Bacillus subtilis | 2.1–3.3 | [41] |
| 8. |  | Tryprostatin B | Staphylococcus aureus Bacillus subtilis | 2.1–3.3 | [41] |
| 9. |  | Brevianamide F, cyclo(l-Trp-l-Pro) | Micrococcus luteus Staphylococcus aureus | Reported as zones of inhibition of 14 mm | [216] |
| 10. |  | cyclo(l-Phe-d-Pro) | Vibrio anguillarum | 0.10 | [220] |
| Staphylococcus aureus | 14 mm zone of inhibition | [108] | |||
| 11. |  | cyclo(d-Phe-d-Pro) | Vibrio anguillarum | 0.03 | [220] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bojarska, J.; Mieczkowski, A.; Ziora, Z.M.; Skwarczynski, M.; Toth, I.; Shalash, A.O.; Parang, K.; El-Mowafi, S.A.; Mohammed, E.H.M.; Elnagdy, S.; et al. Cyclic Dipeptides: The Biological and Structural Landscape with Special Focus on the Anti-Cancer Proline-Based Scaffold. Biomolecules 2021, 11, 1515. https://doi.org/10.3390/biom11101515
Bojarska J, Mieczkowski A, Ziora ZM, Skwarczynski M, Toth I, Shalash AO, Parang K, El-Mowafi SA, Mohammed EHM, Elnagdy S, et al. Cyclic Dipeptides: The Biological and Structural Landscape with Special Focus on the Anti-Cancer Proline-Based Scaffold. Biomolecules. 2021; 11(10):1515. https://doi.org/10.3390/biom11101515
Chicago/Turabian StyleBojarska, Joanna, Adam Mieczkowski, Zyta M. Ziora, Mariusz Skwarczynski, Istvan Toth, Ahmed O. Shalash, Keykavous Parang, Shaima A. El-Mowafi, Eman H. M. Mohammed, Sherif Elnagdy, and et al. 2021. "Cyclic Dipeptides: The Biological and Structural Landscape with Special Focus on the Anti-Cancer Proline-Based Scaffold" Biomolecules 11, no. 10: 1515. https://doi.org/10.3390/biom11101515