UPLC–MS Triglyceride Profiling in Sunflower and Rapeseed Seeds

 , , , ,
, , , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Reagents
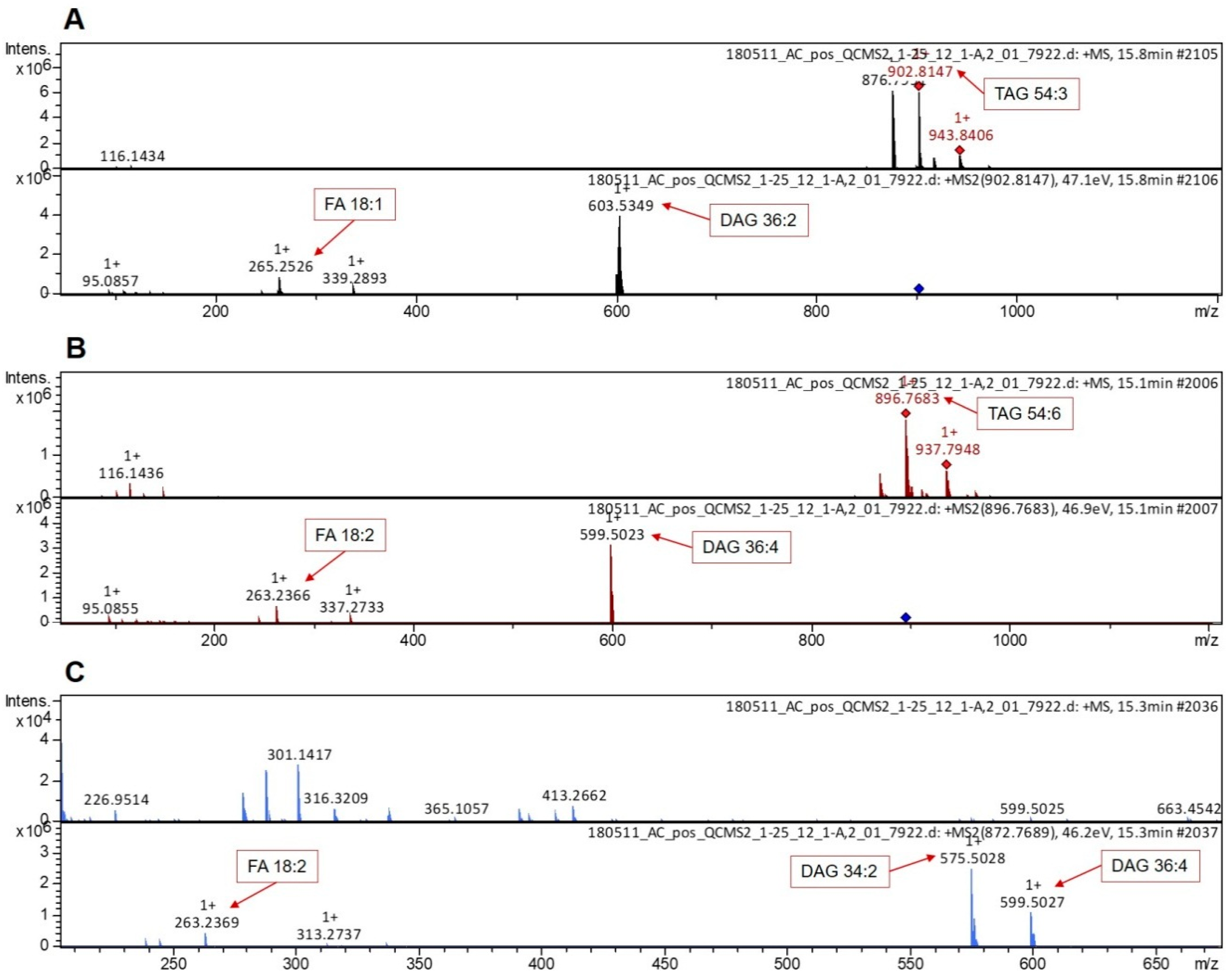
2.3. UPLC–MS Analysis
2.4. Data Processing and Analysis
3. Results and Discussion
3.1. Sunflower versus Rapeseed
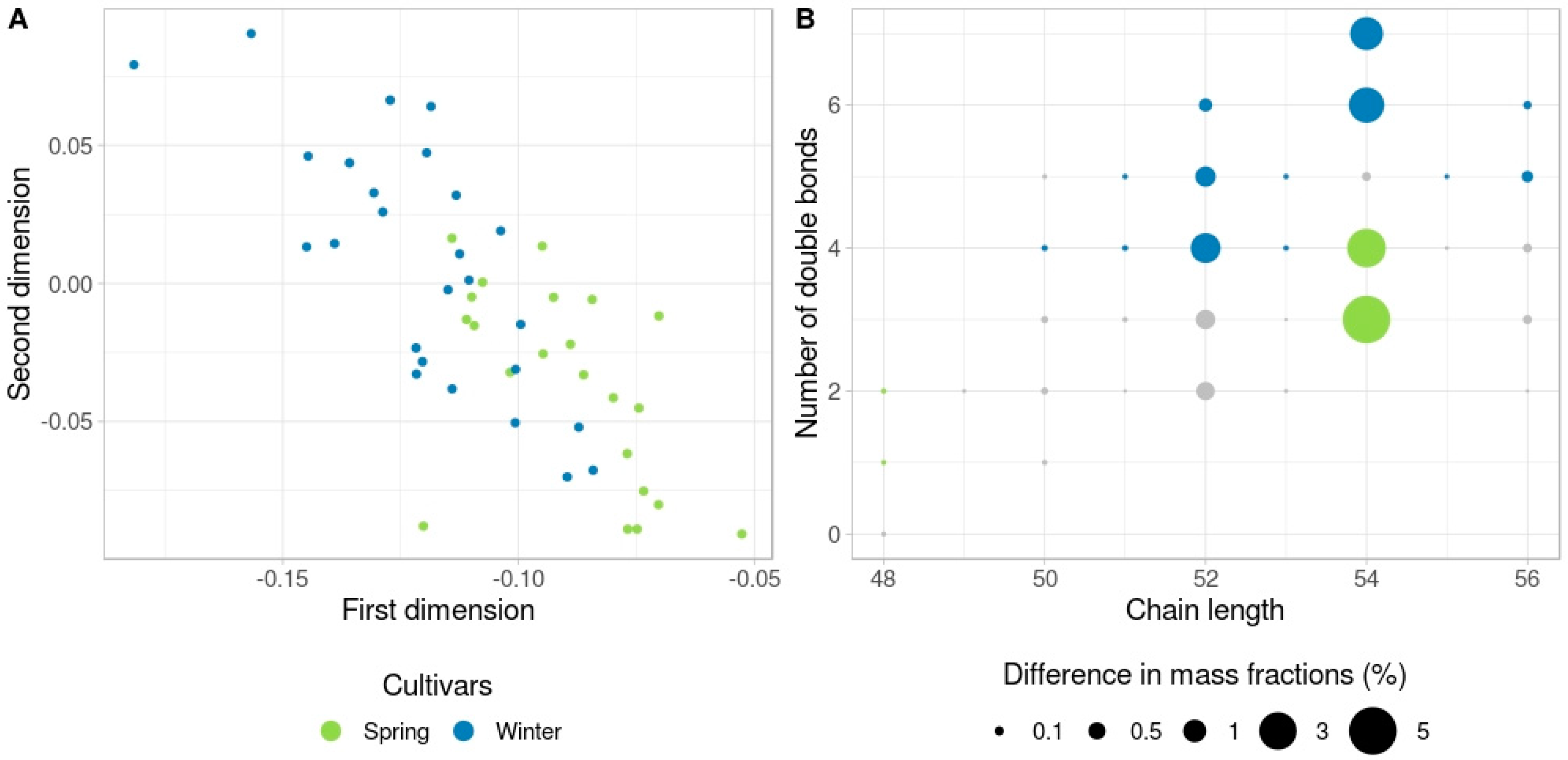
3.2. Spring-Type versus Winter-Type Rapeseed
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Thomas, A. Fats and Fatty Oils. In Ullmann’s Encyclopedia of Industrial Chemistry; Wiley-VCH Verlag GmbH & Co. KGaA, Ed.; Wiley-VCH Verlag GmbH & Co. KGaA: Weinheim, Germany, 2000; ISBN 978-3-527-30673-2. [Google Scholar]
- Durrett, T.P.; Benning, C.; Ohlrogge, J. Plant triacylglycerols as feedstocks for the production of biofuels. Plant J. 2008, 54, 593–607. [Google Scholar] [CrossRef]
- Montero de Espinosa, L.; Meier, M.A.R. Plant oils: The perfect renewable resource for polymer science?! Eur. Polym. J. 2011, 47, 837–852. [Google Scholar] [CrossRef]
- Xu, C.; Shanklin, J. Triacylglycerol metabolism, function, and accumulation in plant vegetative tissues. Annu. Rev. Plant Biol. 2016, 67, 179–206. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Benning, C. Functions of triacylglycerols during plant development and stress. Curr. Opin. Biotechnol. 2018, 49, 191–198. [Google Scholar] [CrossRef] [PubMed]
- Cagliari, A.; Margis, R.; Dos Santos Maraschin, F.; Turchetto-Zolet, A.C.; Loss, G.; Margis-Pinheiro, M. Biosynthesis of triacylglycerols (TAGs) in plants and algae. Int. J. Plant Biol. 2011, 2, e10. [Google Scholar] [CrossRef]
- Voelker, T.A.; Kinney, A.J. Variations in the biosynthesis of seed-storage lipids. Annu. Rev. Plant Physiol. Plant Mol. Biol. 2001, 52, 335–361. [Google Scholar] [CrossRef]
- Rauf, S.; Jamil, N.; Tariq, S.A.; Khan, M.; Kausar, M.; Kaya, Y. Progress in modification of sunflower oil to expand its industrial value. J. Sci. Food Agric. 2017, 97, 1997–2006. [Google Scholar] [CrossRef]
- Velasco, L.; Fernández-martínez, J.M. Breeding oilseed crops for improved oil quality. J. Crop Prod. 2002, 5, 309–344. [Google Scholar] [CrossRef]
- Sakhno, L. Variability in the fatty acid composition of rapeseed oil: Classical breeding and biotechnology. Cytol. Genet. 2010, 44, 389–397. [Google Scholar] [CrossRef]
- Chapman, M.A.; Burke, J.M. Evidence of selection on fatty acid biosynthetic genes during the evolution of cultivated sunflower. Theor. Appl. Genet. 2012, 125, 897–907. [Google Scholar] [CrossRef]
- Joksimović, J.; Atlagić, J.; Marinković, R.; Jovanović, D. Genetic control of oleic and linoleic acid contents in sunflower/control genético del contenido de aceite oleico y linólico en girasol/contrôle génétique des contenus d’acide oléique et linoléique chez le tournesol. Helia 2006, 29, 33–40. [Google Scholar]
- Bocianowski, J.; Mikołajczyk, K.; Bartkowiak-Broda, I. Determination of fatty acid composition in seed oil of rapeseed (Brassica napus L.) by mutated alleles of the FAD3 desaturase genes. J. Appl. Genet. 2012, 53, 27–30. [Google Scholar] [CrossRef]
- Fuller, G.; Diamond, M.J.; Applewhite, T.H. High-oleic safflower oil. Stability and chemical modification. J. Am. Oil Chem. Soc. 1967, 44, 264–266. [Google Scholar] [CrossRef]
- Premnath, A.; Narayana, M.; Ramakrishnan, C.; Kuppusamy, S.; Chockalingam, V. Mapping quantitative trait loci controlling oil content, oleic acid and linoleic acid content in sunflower (Helianthus annuus L.). Mol. Breed. 2016, 36, 106. [Google Scholar] [CrossRef]
- Anushree, S.; André, M.; Guillaume, D.; Frédéric, F. Stearic sunflower oil as a sustainable and healthy alternative to palm oil. A review. Agron. Sustain. Dev. 2017, 37, 18. [Google Scholar] [CrossRef]
- Fernández-Moya, V.; Martínez-Force, E.; Garcés, R. Identification of triacylglycerol species from high-saturated sunflower (Helianthus annuus) mutants. J. Agric. Food Chem. 2000, 48, 764–769. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Moya, V.; Martínez-Force, E.; Garcés, R. Metabolism of triacylglycerol species during seed germination in fatty acid sunflower (Helianthus annuus) Mutants. J. Agric. Food Chem. 2000, 48, 770–774. [Google Scholar] [CrossRef] [PubMed]
- Guan, M.; Chen, H.; Xiong, X.; Lu, X.; Li, X.; Huang, F.; Guan, C. A Study on triacylglycerol composition and the structure of high-oleic rapeseed oil. Engineering 2016, 2, 258–262. [Google Scholar] [CrossRef]
- Hawkins, G.P.; Deng, Z.; Kubik, T.J.; Johnson-Flanagan, A.M. Characterization of freezing tolerance and vernalization in Vern-, a spring-type Brassica napus line derived from a winter cross. Planta 2002, 216, 220–226. [Google Scholar] [CrossRef]
- Huang, Z.; Zhang, X.; Jiang, S.; Qin, M.; Zhao, N.; Lang, L.; Liu, Y.; Tian, Z.; Liu, X.; Wang, Y.; et al. Analysis of cold resistance and identification of SSR markers linked to cold resistance genes in Brassica rapa L. Breed. Sci. 2017, 67, 213–220. [Google Scholar] [CrossRef]
- Rihan, H.Z.; Al-Issawi, M.; Fuller, M.P. Advances in physiological and molecular aspects of plant cold tolerance. J. Plant Interact. 2017, 12, 143–157. [Google Scholar] [CrossRef] [Green Version]
- Giavalisco, P.; Li, Y.; Matthes, A.; Eckhardt, A.; Hubberten, H.-M.; Hesse, H.; Segu, S.; Hummel, J.; Köhl, K.; Willmitzer, L. Elemental formula annotation of polar and lipophilic metabolites using 13C, 15N and 34S isotope labelling, in combination with high-resolution mass spectrometry. Plant J. 2011, 68, 364–376. [Google Scholar] [CrossRef] [PubMed]
- Smith, C.A.; Want, E.J.; O’Maille, G.; Abagyan, R.; Siuzdak, G. XCMS: Processing mass spectrometry data for metabolite profiling using nonlinear peak alignment, matching, and identification. Anal. Chem. 2006, 78, 779–787. [Google Scholar] [CrossRef] [PubMed]
- Libiseller, G.; Dvorzak, M.; Kleb, U.; Gander, E.; Eisenberg, T.; Madeo, F.; Neumann, S.; Trausinger, G.; Sinner, F.; Pieber, T.; et al. IPO: A tool for automated optimization of XCMS parameters. BMC Bioinform. 2015, 16, 118. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2013; Available online: http://www.R-project.org/ (accessed on 10 October 2018).
- Hummel, J.; Segu, S.; Li, Y.; Irgang, S.; Jueppner, J.; Giavalisco, P. Ultra performance liquid chromatography and high resolution mass spectrometry for the analysis of plant lipids. Front. Plant Sci. 2011, 2, 54. [Google Scholar] [CrossRef]
- Himawan, C.; Starov, V.M.; Stapley, A.G.F. Thermodynamic and kinetic aspects of fat crystallization. Adv. Colloid Interface Sci. 2006, 122, 3–33. [Google Scholar] [CrossRef]
- Teutonico, R.A.; Yandell, B.; Satagopan, J.M.; Ferreira, M.E.; Palta, J.P.; Osborn, T.C. Genetic analysis and mapping of genes controlling freezing tolerance in oilseed Brassica. Mol. Breed. 1995, 1, 329–339. [Google Scholar] [CrossRef]
- Roughan, P.G. Phosphatidylglycerol and chilling sensitivity in plants. Plant Physiol. 1985, 77, 740–746. [Google Scholar] [CrossRef]




© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chernova, A.; Gubaev, R.; Mazin, P.; Goryunova, S.; Demurin, Y.; Gorlova, L.; Vanushkina, A.; Mair, W.; Anikanov, N.; Martynova, E.; et al. UPLC–MS Triglyceride Profiling in Sunflower and Rapeseed Seeds. Biomolecules 2019, 9, 9. https://doi.org/10.3390/biom9010009
Chernova A, Gubaev R, Mazin P, Goryunova S, Demurin Y, Gorlova L, Vanushkina A, Mair W, Anikanov N, Martynova E, et al. UPLC–MS Triglyceride Profiling in Sunflower and Rapeseed Seeds. Biomolecules. 2019; 9(1):9. https://doi.org/10.3390/biom9010009
Chicago/Turabian StyleChernova, Alina, Rim Gubaev, Pavel Mazin, Svetlana Goryunova, Yakov Demurin, Lyudmila Gorlova, Anna Vanushkina, Waltraud Mair, Nikolai Anikanov, Elena Martynova, and et al. 2019. "UPLC–MS Triglyceride Profiling in Sunflower and Rapeseed Seeds" Biomolecules 9, no. 1: 9. https://doi.org/10.3390/biom9010009