
Bag-1 Protects Nucleus Pulposus Cells from Oxidative Stress by Interacting with HSP70
 , ,
, ,  , , , ,
, , , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation of NP Cells, Hypoxic Culture Conditions, and Cell Treatments
2.2. Bag-1 Overexpression
2.3. Immunohistological Studies
2.4. Cell Viability Assay
2.5. Measurement of Intracellular ROS
2.6. Detection of Mitochondrial Membrane Potential
2.7. Real-Time RT-PCR Analysis
2.8. Protein Extraction, Western Blot Analysis, and Immunoprecipitation
2.9. Statistical Analysis
3. Results
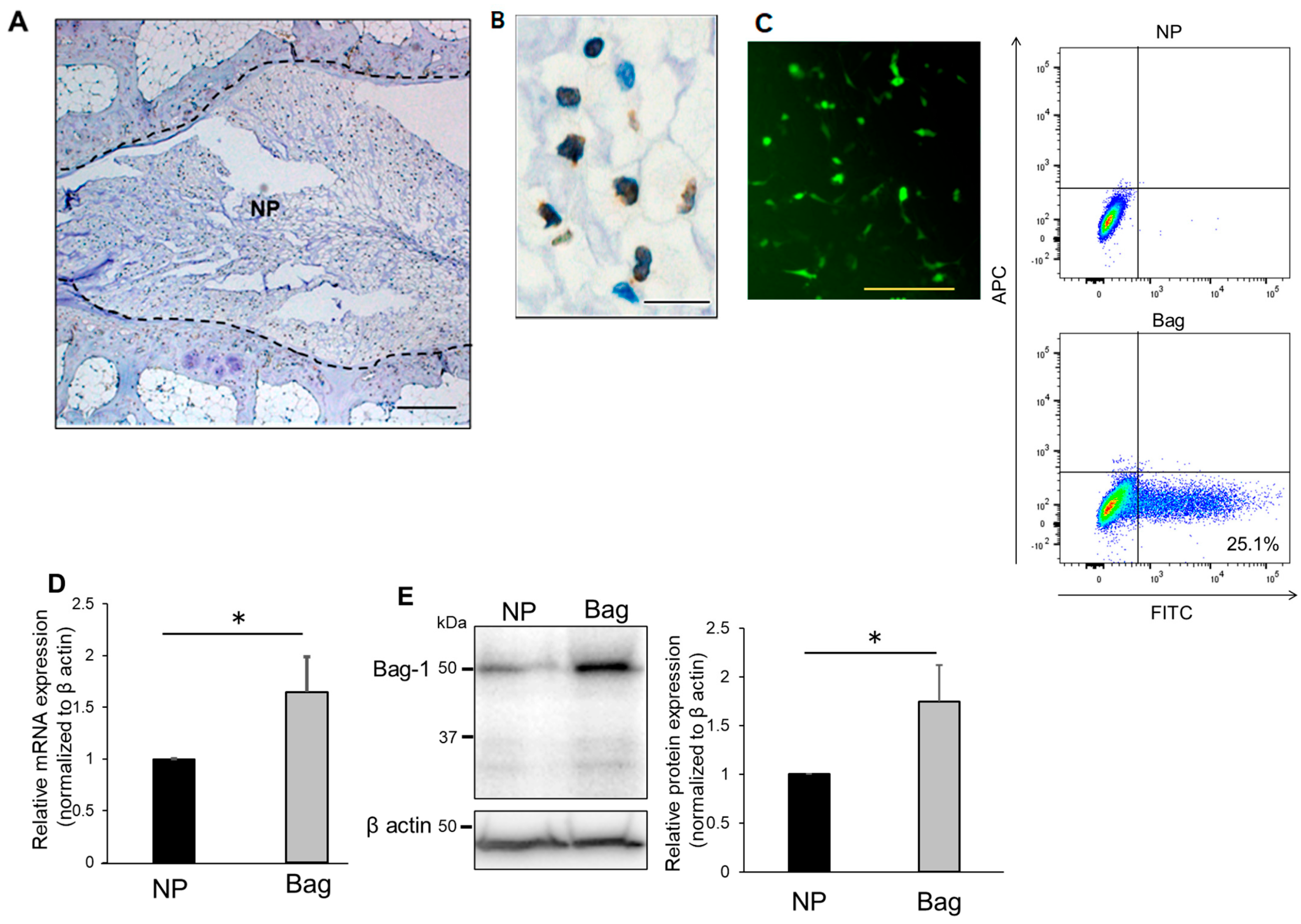
3.1. Evaluation of Bag-1 Expression in NP Cells
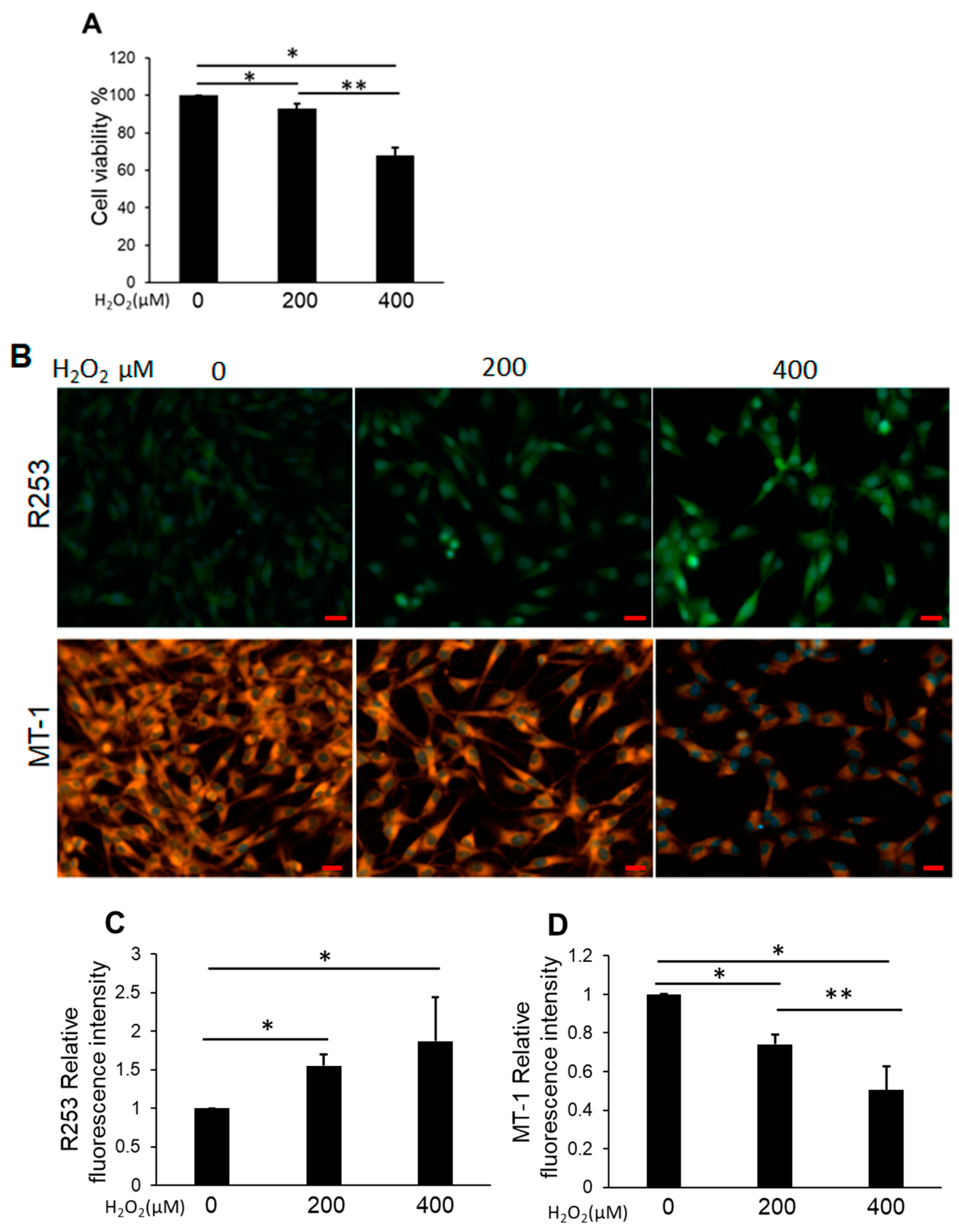
3.2. H2O2 Impaired NP Cell Viability and Mitochondrial Function and Increased Intracellular ROS
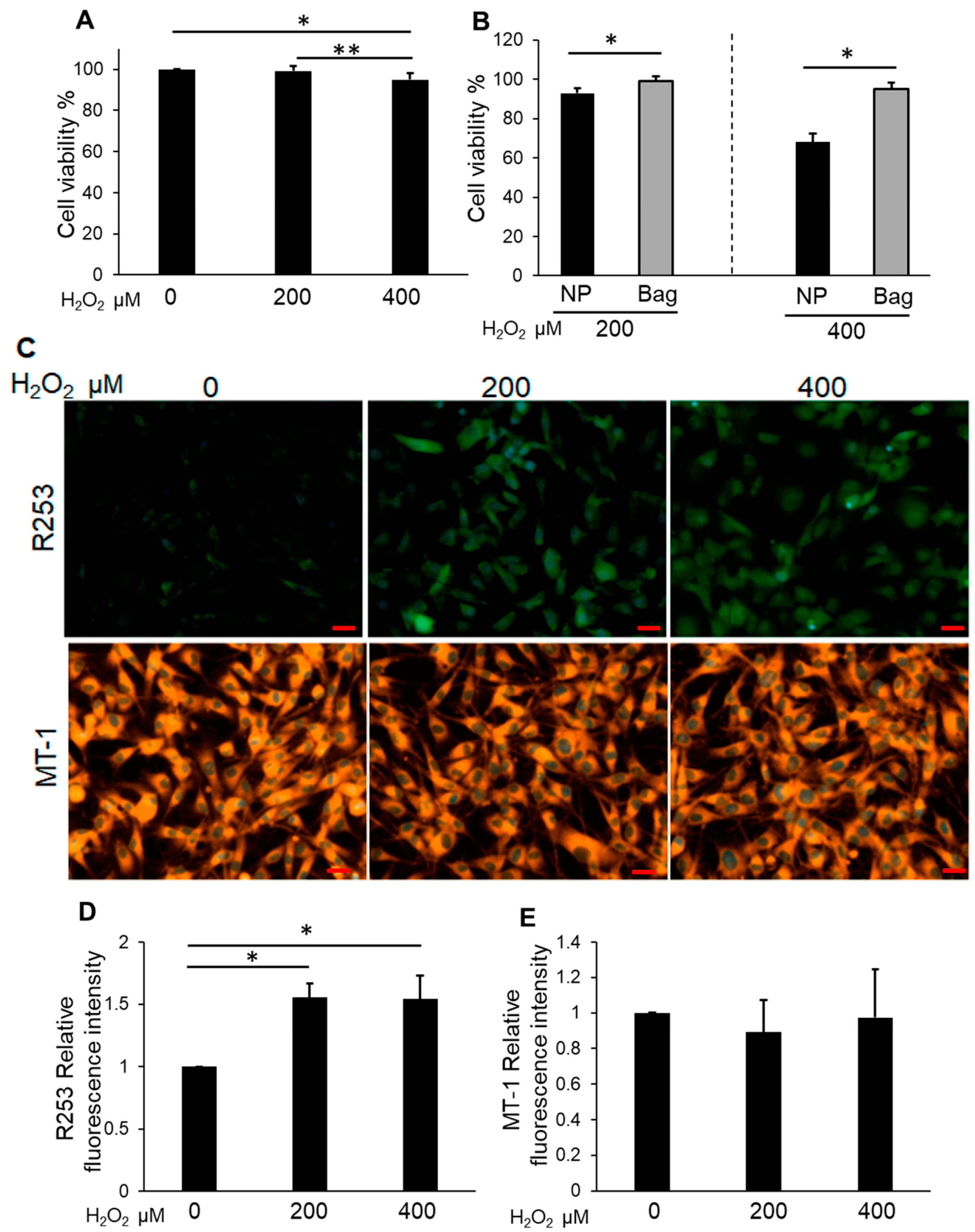
3.3. Bag-1-Overexpressing NP Cells Attenuate the Effect of H2O2 on Cell Viability, Mitochondrial Function, and Increased Intracellular ROS Levels
3.4. Bag-1 Binds to HSP70 in NP Cells, but Does Exhibit Obvious Raf-1 Binding
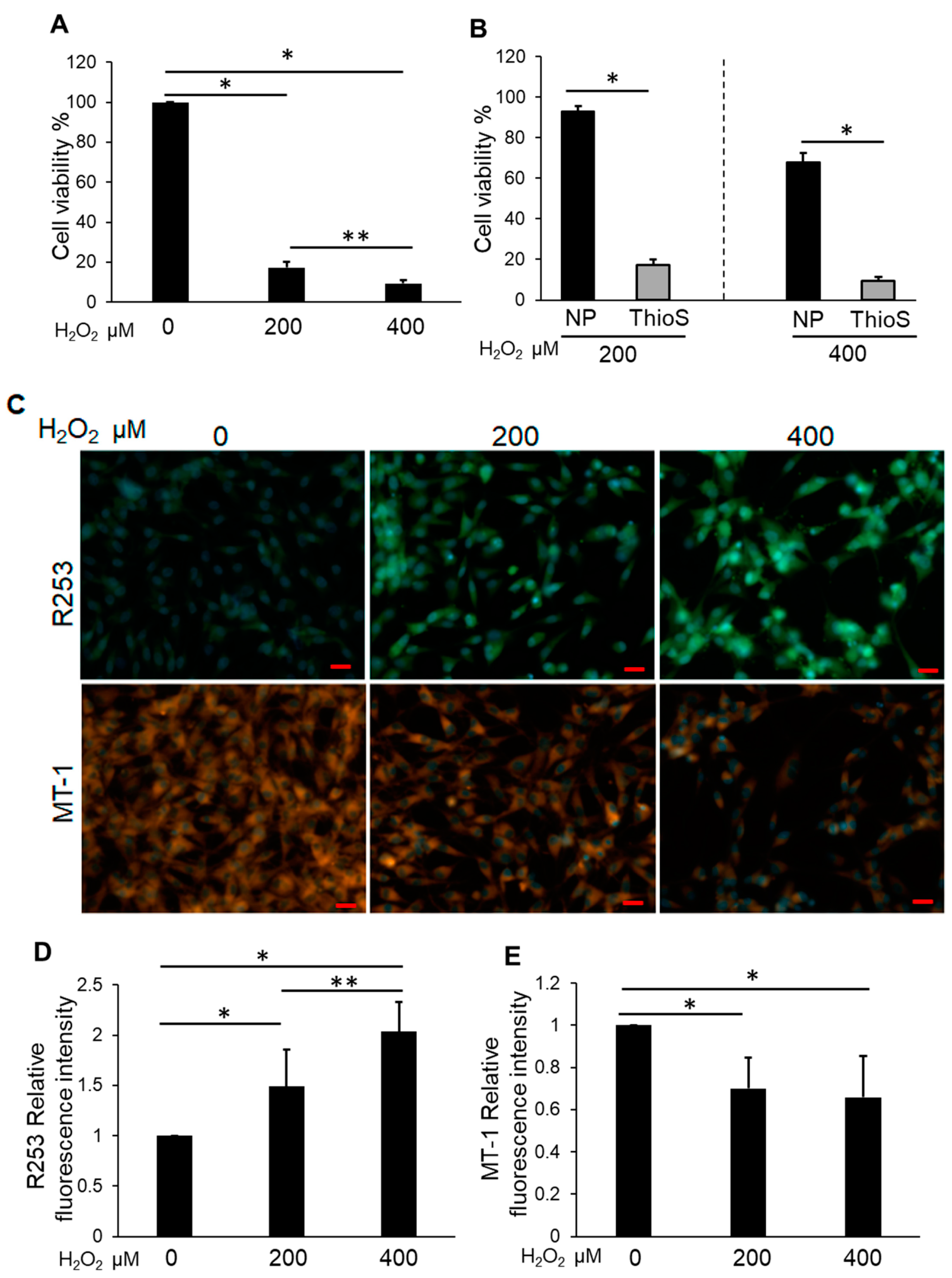
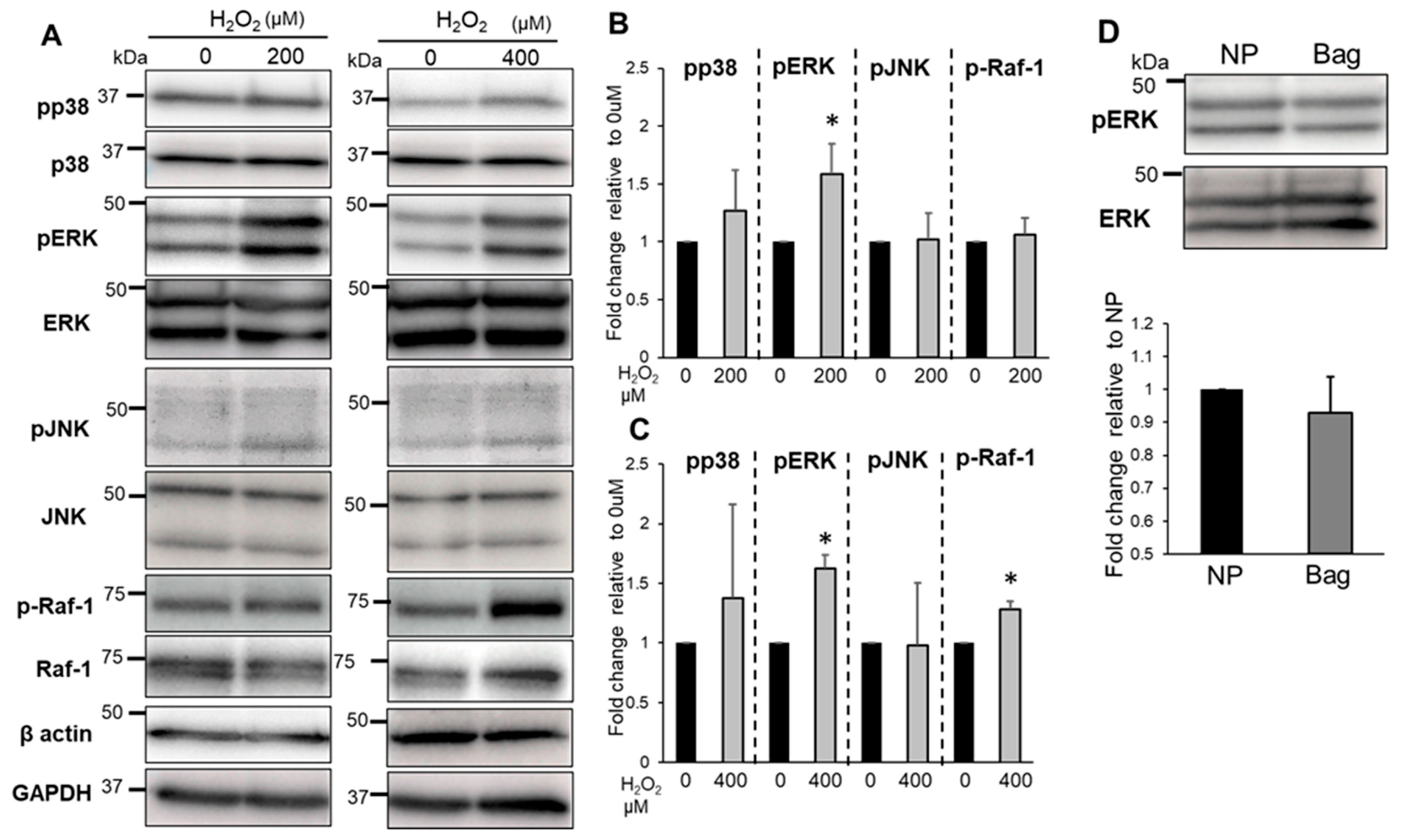
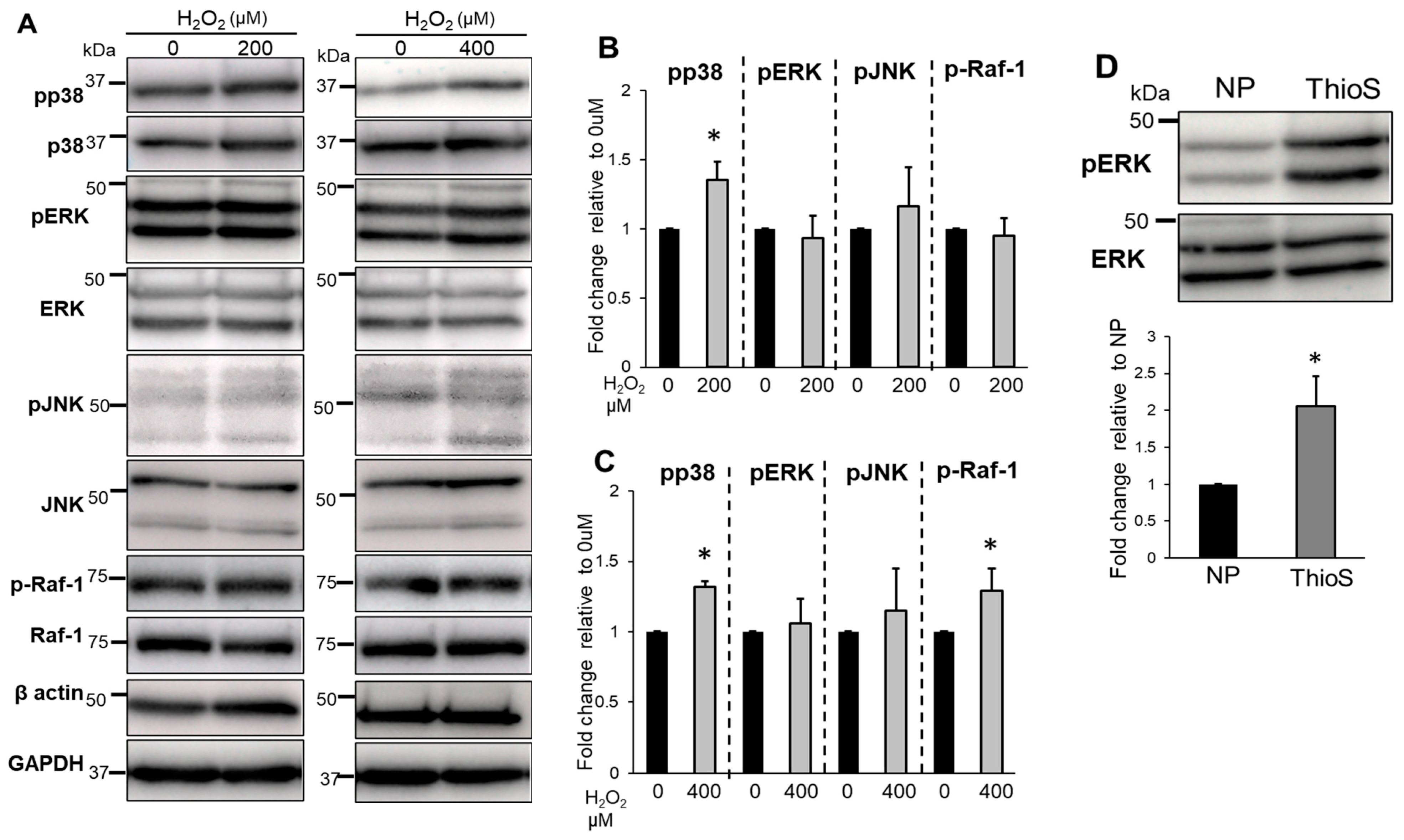
3.5. Treatment of Bag-1 with an Inhibitor of the Binding Site for HSP70 and Raf-1 Attenuates NP Cell Viability, Mitochondrial Function, and Increased Intracellular ROS Levels
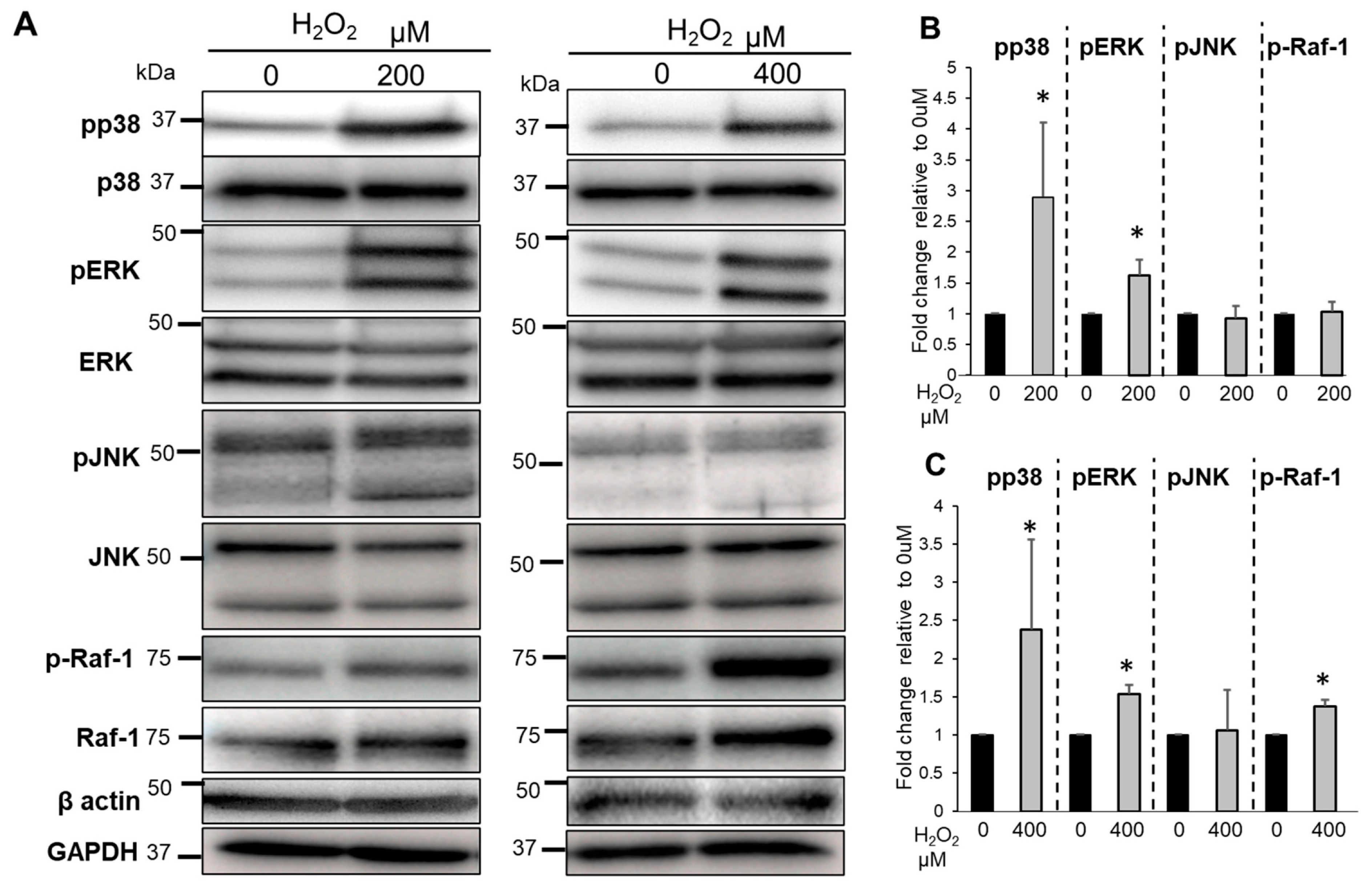
3.6. The Effects of Bag-1 on MAPKs and Raf-1 Activation
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Martin, B.I.; Deyo, R.A.; Mirza, S.K.; Turner, J.A.; Comstock, B.A.; Hollingworth, W.; Sullivan, S.D. Expenditures and Health Status among Adults with Back and Neck Problems. JAMA 2008, 299, 656–664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vo, N.V.; Hartman, R.A.; Patil, P.R.; Risbud, M.V.; Kletsas, D.; Iatridis, J.C.; Hoyland, J.A.; Le Maitre, C.L.; Sowa, G.A.; Kang, J.D. Molecular mechanisms of biological aging in intervertebral discs. J. Orthop. Res. 2016, 34, 1289–1306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Urban, J.P.; Roberts, S. Degeneration of the intervertebral disc. Arthritis Res. Ther. 2003, 5, 120–130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, F.; Zhao, X.; Shen, H.; Zhang, C. Molecular mechanisms of cell death in intervertebral disc degeneration (Review). Int. J. Mol. Med. 2016, 37, 1439–1448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patil, P.; Falabella, M.; Saeed, A.; Lee, D.; Kaufman, B.; Shiva, S.; Croix, C.S.; Van Houten, B.; Niedernhofer, L.J.; Robbins, P.D.; et al. Oxidative stress-induced senescence markedly increases disc cell bioenergetics. Mech. Ageing Dev. 2019, 180, 97–106. [Google Scholar] [CrossRef]
- Zhou, J.; Liu, Q.; Yang, Z.; Xie, C.; Ling, L.; Hu, H.; Cao, Y.; Huang, Y.; Hua, Y. Rutin maintains redox balance to relieve oxidative stress induced by TBHP in nucleus pulposus cells. In Vitro Cell. Dev. Biol. Anim. 2021, 57, 448–456. [Google Scholar] [CrossRef]
- Feng, C.; Liu, H.; Yang, M.; Zhang, Y.; Huang, B.; Zhou, Y. Disc cell senescence in intervertebral disc degeneration: Causes and molecular pathways. Cell Cycle 2016, 15, 1674–1684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Yang, B.; Wang, J.; Cheng, F.; Shi, K.; Ying, L.; Wang, C.; Xia, K.; Huang, X.; Gong, Z.; et al. Cell Senescence: A Nonnegligible Cell State under Survival Stress in Pathology of Intervertebral Disc Degeneration. Oxid. Med. Cell. Longev. 2020, 2020, 9503562. [Google Scholar] [CrossRef]
- Xiang, Q.; Cheng, Z.; Wang, J.; Feng, X.; Hua, W.; Luo, R.; Wang, B.; Liao, Z.; Ma, L.; Li, G.; et al. Allicin Attenuated Advanced Oxidation Protein Product-Induced Oxidative Stress and Mitochondrial Apoptosis in Human Nucleus Pulposus Cells. Oxid. Med. Cell. Longev. 2020, 2020, 6685043. [Google Scholar] [CrossRef]
- Song, Y.; Lu, S.; Geng, W.; Feng, X.; Luo, R.; Li, G.; Yang, C. Mitochondrial quality control in intervertebral disc degeneration. Exp. Mol. Med. 2021, 53, 1124–1133. [Google Scholar] [CrossRef]
- Zhang, S.; Liu, W.; Wang, P.; Hu, B.; Lv, X.; Chen, S.; Wang, B.; Shao, Z. Activation of HSP70 impedes tert-butyl hydroperoxide (t-BHP)-induced apoptosis and senescence of human nucleus pulposus stem cells via inhibiting the JNK/c-Jun pathway. Mol. Cell. Biochem. 2021, 476, 1979–1994. [Google Scholar] [CrossRef] [PubMed]
- Dimozi, A.; Mavrogonatou, E.; Sklirou, A.; Kletsas, D. Oxidative stress inhibits the proliferation, induces premature senescence and promotes a catabolic phenotype in human nucleus pulposus intervertebral disc cells. Eur. Cell. Mater. 2015, 30, 89–102, discussion 103. [Google Scholar] [CrossRef] [PubMed]
- Takayama, S.; Sato, T.; Krajewski, S.; Kochel, K.; Irie, S.; Milian, J.A.; Reed, J.C. Cloning and functional analysis of BAG-1: A novel Bcl-2-binding protein with anti-cell death activity. Cell 1995, 80, 279–284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lv, J.; Shi, L.; Zhai, C.; Wang, G.; Qu, Y. Bag-1L Protects against Cell Apoptosis in an In Vitro Model of Lung Ischemia-Reperfusion Injury through the C-Terminal “Bag” Domain. BioMed Res. Int. 2021, 2021, 8822807. [Google Scholar] [CrossRef]
- Mariotto, E.; Viola, G.; Zanon, C.; Aveic, S. A BAG’s life: Every connection matters in cancer. Pharmacol. Ther. 2020, 209, 107498. [Google Scholar] [CrossRef]
- Denizce Can, N.; Id, E.B.; Kizilboga, T.; Mehmet Akcay, I.; Dingiloglu, B.; Tatli, O.; Acar, S.; Kilbas, P.O.; Elbeyli, E.; Muratcioglu, S.; et al. Interactome analysis of Bag-1 isoforms reveals novel interaction partners in endoplasmic reticulum-associated degradation. PLoS ONE 2021, 16, e0256640. [Google Scholar] [CrossRef]
- Anderson, L.R.; Sutherland, R.L.; Butt, A.J. BAG-1 overexpression attenuates luminal apoptosis in MCF-10A mammary epithelial cells through enhanced RAF-1 activation. Oncogene 2010, 29, 527–538. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Townsend, P.A.; Cutress, R.I.; Sharp, A.; Brimmell, M.; Packham, G. BAG-1: A multifunctional regulator of cell growth and survival. Biochim. Biophys. Acta (BBA)—Rev. Cancer 2003, 1603, 83. [Google Scholar] [CrossRef]
- Takayama, S.; Reed, J.C. Molecular chaperone targeting and regulation by BAG family proteins. Nat. Cell Biol. 2001, 3, E237–E241. [Google Scholar] [CrossRef]
- Takayama, S.; Bimston, D.N.; Matsuzawa, S.; Freeman, B.C.; Aime-Sempe, C.; Xie, Z.; Morimoto, R.I.; Reed, J.C. BAG-1 modulates the chaperone activity of Hsp70/Hsc70. EMBO J. 1997, 16, 4887–4896. [Google Scholar] [CrossRef] [Green Version]
- Lepetsos, P.; Papavassiliou, A.G. ROS/oxidative stress signaling in osteoarthritis. Biochim. Biophys. Acta Rev. Cancer. 2016, 1862, 576–591. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Gong, W.; Wu, S.; Perrett, S. Hsp70 in Redox Homeostasis. Cells 2022, 11, 829. [Google Scholar] [CrossRef] [PubMed]
- Garbarino, V.R.; Orr, M.E.; Rodriguez, K.A.; Buffenstein, R. Mechanisms of oxidative stress resistance in the brain: Lessons learned from hypoxia tolerant extremophilic vertebrates. Arch. Biochem. 2015, 576, 8–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, H.-c.; Xu, X.; Xing, Y.; Wei, Z.-l.; Takahashi, H.; Masuda, S.; Takano, Y. Nuclear or cytoplasmic localization of Bag-1 distinctly correlates with pathologic behavior and outcome of gastric carcinomas. Hum. Pathol. 2010, 41, 724–736. [Google Scholar] [CrossRef] [PubMed]
- Ozfiliz, P.; Arisan, E.D.; Coker-Gurkan, A.; Obakan, P.; Eralp, T.N.; Dinler-Doganay, G.; Palavan-Unsal, N. Bag-1L is a Stress-withstand Molecule Prevents the Downregulation of Mcl-1 and c-Raf Under Control of Heat Shock Proteins in Cisplatin Treated HeLa Cervix Cancer Cells. Asian Pac. J. Cancer Prev. 2014, 15, 4475–4482. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.G.; Takayama, S.; Rapp, U.R.; Reed, J.C. Bcl-2 Interacting Protein BAG-1, Binds to and Activates the Kinase Raf-1. Proc. Natl. Acad. Sci. USA 1996, 93, 7063–7068. [Google Scholar] [CrossRef] [Green Version]
- Song, J.; Takeda, M.; Morimoto, R.I. Bag-1-Hsp70 mediates a physiological stress signalling pathway that regulates Raf-1/ERK and cell growth. Nat. Cell Biol. 2001, 3, 276–282. [Google Scholar] [CrossRef]
- Sun, R.; Zhou, Y.; Cai, Y.; Shui, C.; Wang, X.; Zhu, J. circ_0000045 promotes proliferation, migration, and invasion of head and neck squamous cell carcinomas via regulating HSP70 and MAPK pathway. BMC Cancer 2022, 22, 799. [Google Scholar] [CrossRef]
- Fan, W.; Gao, X.K.; Rao, X.S.; Shi, Y.P.; Liu, X.C.; Fei, Y.; Wang; Liu, Y.F.; Cong, X.X.; He, M.Y.; et al. Hsp70 Interacts with Mitogen-Activated Protein Kinase (MAPK)-Activated Protein Kinase 2 To Regulate p38MAPK Stability and Myoblast Differentiation during Skeletal Muscle Regeneration. Mol. Cell Biol. 2018, 38, e00211-18. [Google Scholar] [CrossRef] [Green Version]
- Risbud, M.V.; Guttapalli, A.; Stokes, D.G.; Hawkins, D.; Danielson, K.G.; Schaer, T.P.; Albert, T.J.; Shapiro, I.M. Nucleus pulposus cells express HIF-1α under normoxic culture conditions: A metabolic adaptation to the intervertebral disc microenvironment. J. Cell. Biochem. 2006, 98, 152–159. [Google Scholar] [CrossRef]
- Enthammer, M.; Papadakis, E.S.; Salomé Gachet, M.; Deutsch, M.; Schwaiger, S.; Koziel, K.; Ashraf, M.I.; Khalid, S.; Wolber, G.; Packham, G.; et al. Isolation of a Novel Thioflavin S–Derived Compound That Inhibits BAG-1–Mediated Protein Interactions and Targets BRAF Inhibitor–Resistant Cell Lines. Mol. Cancer Ther. 2013, 12, 2400–2414. [Google Scholar] [CrossRef] [Green Version]
- Sharp, A.; Crabb, S.J.; Johnson, P.W.M.; Hague, A.; Cutress, R.; Townsend, P.A.; Ganesan, A.; Packham, G. Thioflavin S (NSC71948) Interferes with Bcl-2-Associated Athanogene (BAG-1)-Mediated Protein-Protein Interactions. J. Pharmacol. Exp. Ther. 2009, 331, 680–689. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Jia, C.; Li, Q.; Xie, C.; Zhang, N.; Qu, Y. BAG-1L Protects SH-SY5Y Neuroblastoma Cells Against Hypoxia/Re-oxygenation Through Up-Regulating HSP70 and Activating PI3K/AKT Signaling Pathway. Neurochem. Res. 2017, 42, 2861–2868. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Shen, J.; Chen, Y.; Liu, H.; Zhou, H.; Bai, Z.; Hu, Z.; Guo, X. PINK1 protects against oxidative stress induced senescence of human nucleus pulposus cells via regulating mitophagy. Biochem. Biophys. Res. Commun. 2018, 504, 406–414. [Google Scholar] [CrossRef]
- Cedikova, M.; Pitule, P.; Kripnerova, M.; Markova, M.; Kuncova, J. Multiple Roles of Mitochondria in Aging Processes. Physiol. Res. 2016, 65, S519–S531. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Liao, H.; Bai, D.; Wang, Z.; Xie, X. MAPK/ERK signaling pathway: A potential target for the treatment of intervertebral disc degeneration. Biomed. Pharmacother. 2021, 143, 112170. [Google Scholar] [CrossRef]
- Freund, A.; Patil, C.K.; Campisi, J. p38MAPK is a novel DNA damage response-independent regulator of the senescence-associated secretory phenotype. EMBO J. 2011, 30, 1536–1548. [Google Scholar] [CrossRef] [Green Version]
- Xu, W.; Zhang, X.; Liu, G.; Zhu, M.; Wu, Y.; Jie, Z.; Xie, Z.; Wang, S.; Ma, Q.; Fan, S.; et al. Oxidative stress abrogates the degradation of KMT2D to promote degeneration in nucleus pulposus. Biochim. Biophys. Acta Mol. Cell Res. 2020, 1866, 165888. [Google Scholar] [CrossRef]
- Han, Y.; Li, X.; Yan, M.; Yang, M.; Wang, S.; Pan, J.; Li, L.; Tan, J. Oxidative damage induces apoptosis and promotes calcification in disc cartilage endplate cell through ROS/MAPK/NF-κB pathway: Implications for disc degeneration. Biochem. Biophys. Res. Commun. 2019, 516, 1026–1032. [Google Scholar] [CrossRef]
- Xu, Y.; Yao, H.; Wang, Q.; Xu, W.; Liu, K.; Zhang, J.; Zhao, H.; Hou, G. Aquaporin-3 Attenuates Oxidative Stress-Induced Nucleus Pulposus Cell Apoptosis Through Regulating the P38 MAPK Pathway. Cell. Physiol. Biochem. 2018, 50, 1687–1697. [Google Scholar] [CrossRef]
- Zuiderweg, E.R.P.; Bertelsen, E.B.; Rousaki, A.; Mayer, M.P.; Gestwicki, J.E.; Ahmad, A. Allostery in the Hsp70 Chaperone Proteins. Top. Curr. Chem. 2013, 328, 99–153. [Google Scholar] [CrossRef] [Green Version]
- Kizilboga, T.; Baskale, E.A.; Yildiz, J.; Akcay, I.M.; Zemheri, E.; Can, N.D.; Ozden, C.; Demir, S.; Ezberci, F.; Dinler-Doganay, G. Bag-1 stimulates Bad phosphorylation through activation of Akt and Raf kinases to mediate cell survival in breast cancer. BMC Cancer 2019, 19, 1254. [Google Scholar] [CrossRef]
- Tekari, A.; Marazza, A.; Crump, K.; Bermudez-lekerika, P.; Gantenbein, B. Inhibition of the extracellular signal-regulated kinase pathway reduces the inflammatory component in nucleus pulposus cells. J. Orthop. Res. 2022, 40, 2362. [Google Scholar] [CrossRef]
- Risbud, M.V.; Guttapalli, A.; Albert, T.J.; Shapiro, I.M. Hypoxia activates MAPK activity in rat nucleus pulposus cells: Regulation of integrin expression and cell survival. Spine 2005, 30, 2503–2509. [Google Scholar] [CrossRef]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Suyama, K.; Sakai, D.; Hayashi, S.; Qu, N.; Terayama, H.; Kiyoshima, D.; Nagahori, K.; Watanabe, M. Bag-1 Protects Nucleus Pulposus Cells from Oxidative Stress by Interacting with HSP70. Biomedicines 2023, 11, 863. https://doi.org/10.3390/biomedicines11030863
Suyama K, Sakai D, Hayashi S, Qu N, Terayama H, Kiyoshima D, Nagahori K, Watanabe M. Bag-1 Protects Nucleus Pulposus Cells from Oxidative Stress by Interacting with HSP70. Biomedicines. 2023; 11(3):863. https://doi.org/10.3390/biomedicines11030863
Chicago/Turabian StyleSuyama, Kaori, Daisuke Sakai, Shogo Hayashi, Ning Qu, Hayato Terayama, Daisuke Kiyoshima, Kenta Nagahori, and Masahiko Watanabe. 2023. "Bag-1 Protects Nucleus Pulposus Cells from Oxidative Stress by Interacting with HSP70" Biomedicines 11, no. 3: 863. https://doi.org/10.3390/biomedicines11030863