The Function of Lgr5+ Cells in the Gastric Antrum Does Not Require Fzd7 or Myc In Vivo
by
 and
and
Dustin Flanagan
1,†, 
Nick Barker
2,3,
Matthias Ernst
4,
Elizabeth Vincan
1,5,* and
Toby Phesse
1,6,* 1
University of Melbourne & Victorian Infectious Diseases Reference Laboratory, Doherty Institute of Infection and Immunity, Melbourne VIC 3000, Australia
2
Institute of Medical Biology, Singapore 138648, Singapore
3
MRC Centre for Regenerative Medicine, University of Edinburgh, Edinburgh EH8 9YL, UK
4
Cancer and Inflammation Laboratory, Olivia Newton-John Cancer Research Institute and School of Cancer Medicine, La Trobe University, Heidelberg VIC 3084, Australia
5
School of Pharmacy and Biomedical Sciences, Curtin University, Perth WA 6845, Australia
6
European Cancer Stem Cell Research Institute, Cardiff University, Cardiff CF24 4HQ, UK
*
Authors to whom correspondence should be addressed.
†
Current address: Cancer Research UK Beatson Institute, Glasgow G61 1BD, UK.
Biomedicines 2019, 7(3), 50; https://doi.org/10.3390/biomedicines7030050
Submission received: 7 June 2019
/
Revised: 2 July 2019
/
Accepted: 5 July 2019
/
Published: 8 July 2019
(This article belongs to the Special Issue Gastric Cancer Research: From Basic Science to the Clinic)
{kind=link}
{kind=link}
{kind=link}
Abstract
:The extreme chemical and mechanical forces endured by the gastrointestinal tract drive a constant renewal of the epithelial lining. Stem cells of the intestine and stomach, marked by the cell surface receptor Lgr5, preserve the cellular status-quo of their respective tissues through receipt and integration of multiple cues from the surrounding niche. Wnt signalling is a critical niche component for gastrointestinal stem cells and we have previously shown that the Wnt receptor, Frizzled-7 (Fzd7), is required for gastric homeostasis and the function of Lgr5+ intestinal stem cells. Additionally, we have previously shown a requirement for the Wnt target gene Myc in intestinal homeostasis, regeneration and tumourigenesis. However, it is unknown whether Fzd7 or Myc have conserved functions in gastric Lgr5+ stem cells. Here we show that gastric Lgr5+ stem cells do not require Fzd7 or Myc and are able to maintain epithelial homeostasis, highlighting key differences in the way Wnt regulates homeostasis and Lgr5+ stem cells in the stomach compared to the intestinal epithelium. Furthermore, deletion of Myc throughout the epithelium of the gastric antrum has no deleterious effects suggesting therapeutic targeting of Myc in gastric cancer patients will be well tolerated by the surrounding normal tissue.
1. Introduction
The epithelium of the gastrointestinal tract encounters substantial chemical and physical stresses. One of the mechanisms that has evolved to help cope with these harsh conditions is the continuous turnover of the epithelium, in which new cells are generated from populations of stem cells. These new cells then differentiate and migrate towards the lumen and are eventually sloughed off so that any damaged cells are not retained, and new healthy cells are being produced constantly to replace them. Lgr5 was first identified as a Wnt target gene and a marker of highly proliferative stem cells located at the base of the intestinal crypts that fuel the constant turnover of cells [1]. It has since been confirmed as a stem cell marker in several epithelial tissues including the stomach [2,3], hair follicle [4], ovary [5], mammary gland [6] and kidney [7].
Lgr5 is a receptor for secreted Wnt agonist R-spondins, which acts to modify the strength of Wnt signalling in cells, including those of the intestinal crypt [8]. Wnt signalling regulates several cell functions including proliferation, migration, apoptosis and differentiation, and is critical during embryonic development, and the homeostasis of several tissues including the intestine, stomach, liver and mammary gland [9]. Wnts are a family of 19 glycoproteins that are modified during the secretion process by an O-acyltransferase called Porcupine, which can then bind to Frizzled receptors of which there are 10 in mammals [10]. The fatty acid modification of Wnt ligands is critical for their bind to the U-shape of Frizzled dimers, which also associate with other co-receptors, including Lrp5/6 to form a signalosome with cytoplasmic Dishevelled. This signalosome then inhibits the action of a multimeric degradation complex which allows the co-transcription factor β-catenin to escape degradation and translocate into the nucleus and associate with TCF/Lef transcription factors to regulate target genes [11].
In the intestine, Wnt signalling is most active at the base of the crypts, which contain the Lgr5+ stem cells and Paneth cells [1], with Wnt3a secreted from the Paneth cells [12], and Wnt2b from the underlying stroma [13]. We recently demonstrated that the deletion of Fzd7 in Lgr5+ intestinal epithelial cells was deleterious and triggered rapid repopulation with Fzd7 proficient cells, indicating its requirement for this population of stem cells [14]. Similarly, the deletion of Fzd7 throughout the antrum of the gastric epithelium also triggered repopulation, indicating that Fzd7 regulates a population of stem cells in the antrum [15]. Furthermore, Fzd7 was highly expressed in gastric tumours, and transmitted Wnt signalling to upregulate the transcription factor Myc, to promote tumour initiation and growth [16]. This is consistent with the role of Myc in the intestinal epithelium in which it is required for homeostasis [17], regeneration [18] and tumorigenesis [19].
As Lgr5 marks stem cells in several tissues it is important to understand how these populations of cells are regulated, and therefore here we asked if Fzd7 and Myc regulate Lgr5+ cells in the gastric antrum.
2. Materials and Methods
2.1. Mice
The Tff1CreERT2 [20], Fzd7fl/fl [14], c-Mycfl/fl [21], Rosa26LacZ [22] and Lgr5CreERT2 [1] mice are previously described. Mice were interbred to generate compound mice with appropriate alleles on an inbred C57Bl/6 genetic background. Mice were co-housed using appropriate littermates as controls. All animal experiments were approved by the Animal Ethics Committee, Office for Research Ethics and Integrity, University of Melbourne (1513488, approved on 18th May 2015).
2.2. Treatments
For short-term labelling, mice received a single daily intraperitoneal (IP) injection of 2 mg of tamoxifen. For long-term (>14 days) labelling, mice received single IP injections of 2 mg tamoxifen over three consecutive days.
2.3. Tissue Collection and Histological Analysis
2.4. β-Galactosidase (X-gal) Staining
Mouse stomachs were prepared and stained for X-gal as previously described [15].
2.5. Gland Isolation, Cell Dissociation and Organoid Culture
2.6. Genomic Recombination PCR
2.7. RNA Extraction and Analysis
Whole antral glands and FACS-isolated single antral cells were homogenized in TRizol and total RNA purified, DNAse treated, quantified and subjected to quantitative reverse transcriptase PCR (qRT-PCR). qRT-PCR and calculating gene expression levels relative to the house-keeping gene 18S (2−∆∆Ct) were performed as previously described [25].
2.8. MTT Assay
2.9. Statistical Analysis
Data are expressed as mean ± SEM, where the mean represents the number of mice (≥3 per genotype) or number of independent experiments (≥3). Statistical tests used were two-way ANOVA with Prism7 (GraphPad software) where p values of ≤0.05 were considered significant.
3. Results
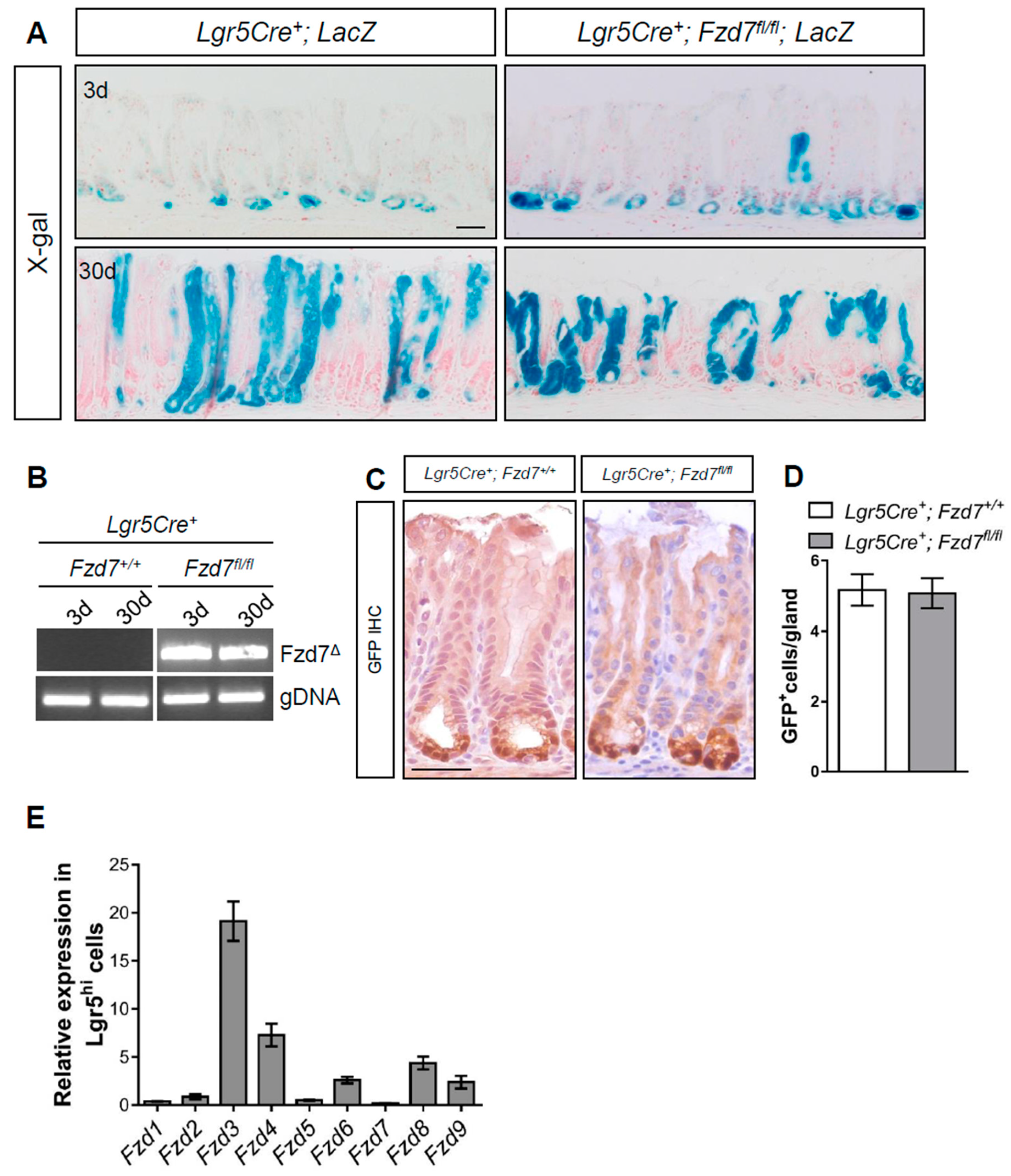
To determine the requirement for Fzd7 in antral Lgr5+ cells we deleted Fzd7 specifically in Lgr5+ cells using Lgr5CreERT2; Fzd7fl/fl; LacZLSL mice and performed lineage tracing via X-gal staining. At 3 days post tamoxifen induction recombined cells of Fzd7 proficient Lgr5CreERT2; Fzd7+/+; LacZLSL mice can be seen at the base of the antral crypts where the Lgr5+ cells are located, and 30 days after tamoxifen many of the crypts are composed entirely of lineage traced cells indicating the variegated Lgr5 locus gives rise to entire gastric units in the antrum as previously reported [2] (Figure 1A). Surprisingly, lineage tracing also proceeded in the Fzd7 deficient Lgr5CreERT2; Fzd7fl/fl; LacZLSL mice indicating that Fzd7 loss is not deleterious to Lgr5+ cells in the gastric antrum, and these stem cells can function without Fzd7. To confirm robust deletion of Fzd7 we performed PCR for the recombined product which gave a very strong band 3 and 30 days after tamoxifen induction in Lgr5CreERT2; Fzd7fl/fl; LacZLSL mice, whilst in Lgr5CreERT2; Fzd7+/+; LacZLSL mice the recombined product was undetectable as expected (Figure 1B). The Lgr5CreERT2 locus also has an EGFP cassette and therefore we performed immunohistochemistry (IHC) for GFP which demonstrated no difference in the number of Lgr5+ cells in the gastric antrum after Fzd7 deletion (Figure 1C,D). Together these data demonstrate that, in contrast to the intestinal epithelium, Fzd7 is not required for the activity of Lgr5+ stem cells in the gastric antral epithelium. Indeed, RT-qPCR for Fzd genes in cells FACS sorted for high GFP expression from the antrum epithelium of Lgr5CreEGFP-ERT2 mice revealed that Fzd7 expression was undetectable (Figure 1E), thus supporting our in vivo observations that Lgr5+ cells in the gastric antrum are regulated differently from those in the intestinal epithelium, and do not require Fzd7.
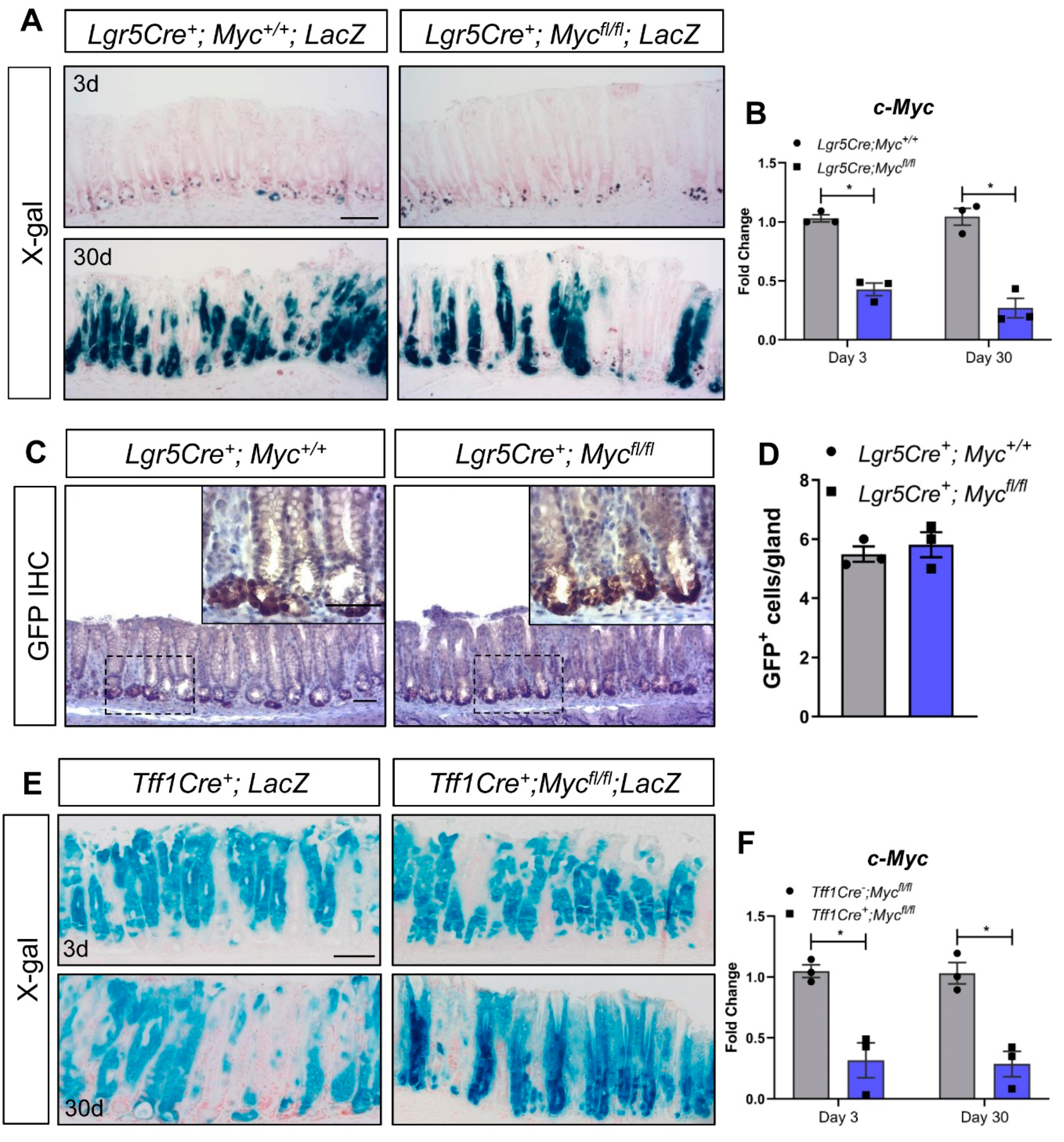
We have previously shown that deletion of Myc in the intestinal epithelium phenocopies deletion of Fzd7 and results in rapid repopulation with Myc proficient cells [17]. Furthermore, we recently demonstrated that Wnt regulates Myc expression, via Fzd7, to control tumour initiation and growth in the stomach [16], regeneration in the intestine [14] and homeostasis in the stomach [15]. To investigate if Lgr5+ cells in the antrum require Myc, we deleted Myc in Lgr5+ cells in vivo. Surprisingly, and similar to deletion of Fzd7, fully lineage traced gastric units were observed in the antrum of Lgr5CreERT2; Mycfl/fl; LacZLSL mice 30 days after tamoxifen induction (Figure 2A), with no difference in the number of Lgr5+ cells between Lgr5CreERT2; Mycfl/fl and Lgr5CreERT2; Myc+/+ mice (Figure 2B,C). These data demonstrate that Lgr5+ antral cells do not require Myc for their stem cell activity. However, there are several stem cell populations identified in the antrum based on expression of distinct maker genes including Lrig1 [26], Sox2 [27] and CCK2R [28] and therefore Myc could be required for activity of one of these populations of stem cells which would have been missed in the analysis of our Lgr5Cre mice.
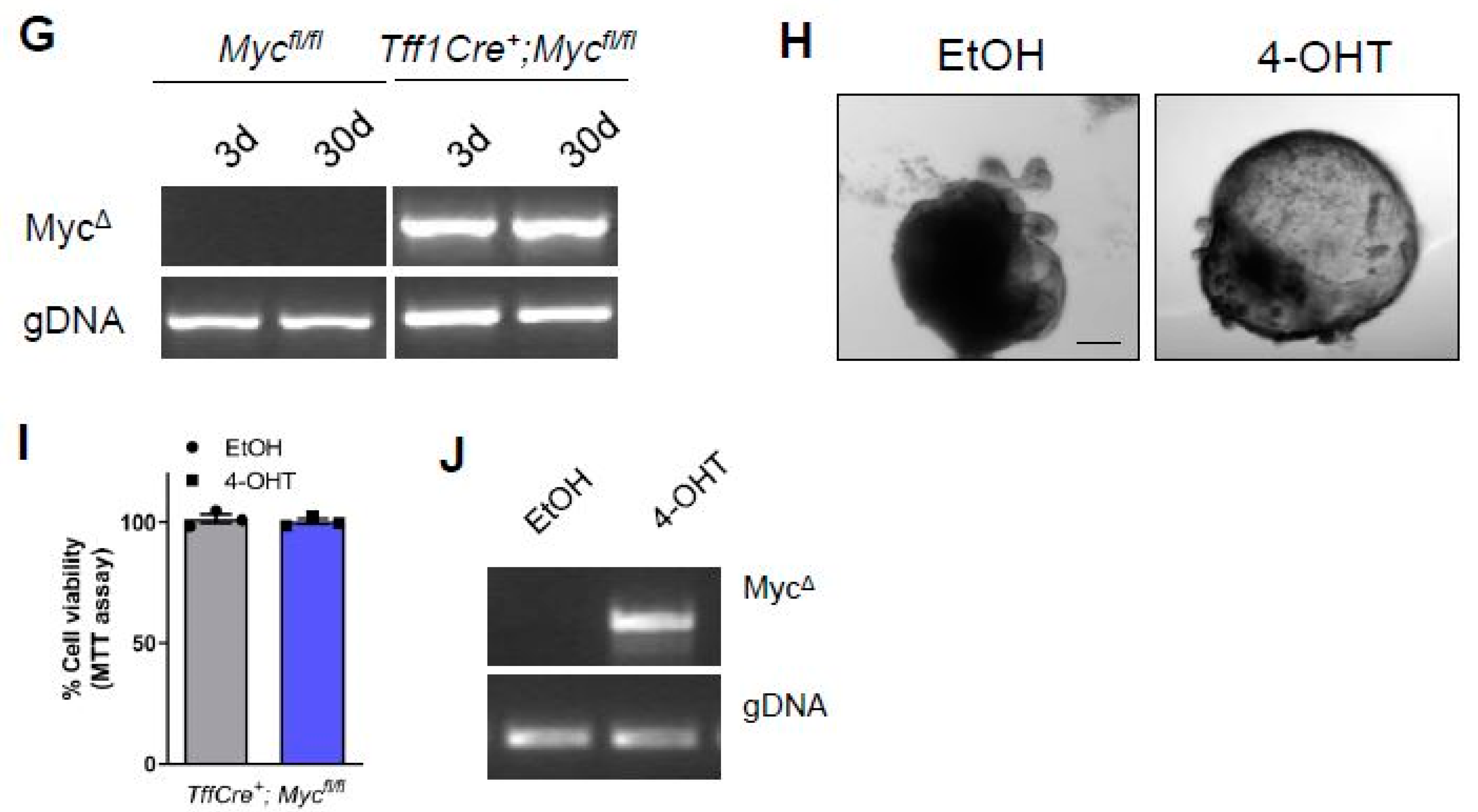
To investigate if Myc is required generically in the antral epithelium we deleted Myc throughout the antral epithelium using Tff1CreERT2 mice [15]. X-Gal staining revealed robust recombination throughout the antral epithelium of Tff1Cre; LacZSLS mice 3 days after tamoxifen induction (Figure 2D). Recombination was still observed at 30 days post induction (Figure 2D), demonstrating that recombination has occurred in at least one population of antral stem cells as previously published [15]. Remarkably, deletion of Myc throughout the antral epithelium of Tff1Cre; Mycfl/fl; LacZSLS mice did not perturb stem cell activity and lineage tracing was able to proceed as per Tff1Cre; Myc+/+; LacZSLS mice (Figure 2D) despite confirming robust deletion of Myc in the antral epithelium via RT-qPCR (Figure 2E), and retention of a strong recombined band via PCR 3 days and 30 days after tamoxifen induction of Tff1Cre; Mycfl/fl; LacZSLS mice (Figure 2F). To help confirm this observation that the antral epithelium can function in the absence of Myc, we cultured gastric organoids from the antrum of Tff1Cre; Mycfl/fl mice, and deleted Myc via treatment with 4-OHT (we have previously demonstrated that 4-OHT does not adversely affect gastric organoids [15]. Myc deleted organoids continued to thrive (Figure 2G) and MTT assays showed no difference in viability compared to vehicle-treated, Myc proficient organoids (Figure 2H), despite confirming robust recombination of the Myc flox allele (Figure 2I). Together these data demonstrate that the epithelium of the gastric antrum does not require Myc in vivo, or in cultured organoids.
4. Discussion
Fzd7 is required for intestinal stem cell activity during homeostasis, regeneration and survival of cultured organoids [29]. Here we show for the first time that deletion of Fzd7 does not inhibit the capacity of Lgr5+ cells to lineage trace full gastric units in the antral epithelium illustrating a substantial difference for Fzd7 in regulating Lgr5+ stem cells in the intestine compared to the antrum. We also show that the number of Lgr5+ cells is maintained in the Fzd7 deficient antrum at similar levels to that of Fzd7 proficient mice, demonstrating that lineage tracing is not due to a small population of Lgr5+ cells that were resistant to Fzd7 deletion, but rather that Fzd7 deletion has not affected Lgr5+ survival and activity. We also observe that other Fzd genes are expressed in Lgr5HI cells in the antrum, with Fzd3 and Fzd4 the highest, suggesting one of these may be transmitting Wnt signalling in antral cells. Interestingly, the expression pattern for Fzd3 seems to be consistent with that of Lgr5 in the gastric antrum whilst Fzd4 is expressed broadly throughout the gastric units [30]. However, Fzd3 is significantly upregulated when Fzd7 is deleted in gastric antrum organoids, but this increased expression is unable to compensate for the loss of Fzd7 in vitro and Fzd7-deficient organoids undergo apoptosis [15]. Thus, the exact Fzd receptors required for antral Lgr5+ cell activity will require additional functional studies in the future including analysis of Wnt pathway activity. We have previously shown that the deletion of Fzd7 throughout the epithelium of the gastric antrum is deleterious and triggers rapid repopulation [15]. This demonstrates that Fzd7 is required for at least one population of stem cells in the gastric antrum, but this population has yet to be identified, and our data here illustrate it is not Lgr5+ antral stem cells. A possible candidate for this population is Axin2+/Lgr5- cells which are located in a similar location as Fzd7 expressing cells in the lower half of the antral glands [30]. Furthermore, the Axin2+/Lgr5- population in the antrum expands in response to Helicobacter pylori infection via upregulation of Rspo3 in the underlying myofibroblasts [30], whilst inhibition of Fzd7 can suppress H. pylori-induced Wnt signalling and proliferation [31]. These data suggest Fzd7 may be regulating Axin2+/Lgr5- cells in the antrum, although this is yet to be confirmed.
We have previously shown that Myc is required for all the tumourigenic phenotypes following truncation of Apc in the intestine [19], and also for homeostasis [17], regeneration [18] and apoptosis [32] in this tissue. Furthermore, we recently demonstrated that tumour initiation and growth in the gastric antrum requires Fzd7 dependant upregulation of Myc [16]. Myc is upregulated in many cancers and thus represents a potential target for therapy which has been the subject of intense research for several years. However, Myc is not required in all the adult tissues it is expressed in, for example, both liver zonation and Apc loss induced hepatomegaly are regulated by Wnt signalling via Myc independent mechanisms [33,34]. Given its differential requirement between different tissues it is important to gain a full insight into how the loss of Myc affects separate tissues. To our surprise, Myc deletion did not inhibit stem cell activity in Lgr5+ stem cells of the gastric antrum or when genetically deleted throughout the entire epithelium of the gastric antrum. Furthermore, organoids cultured from the antrum were able to thrive and showed no difference in viability when Myc was deleted compared to Myc proficient organoids. These data demonstrate that Myc is not only dispensable for antral Lgr5+ stem cells, but the entire antral epithelium.
These data identify a distinct difference in the way Wnt regulates homeostasis and stem cells in the gastric antrum compared to the intestinal epithelium. As Myc is required for gastric tumour growth, it also informs future clinical trials with putative Myc inhibitors that the gastric epithelium can tolerate the loss of Myc. Future investigations into the differences between the intestinal and gastric epithelium could help uncover why the intestine is more sensitive to deregulated Wnt, and explore if Wnt inhibitors elicit a different response from these two tissues thus identifying new oncogenes/tumour suppressors in both cancer types.
Intestinal-type gastric tumours undergo metaplasia to become more intestinal-like, with upregulation of intestinal specific genes and morphological features including tubular structures. This could explain why gastric adenomas are sensitive to deletion of Myc, whilst the normal gastric epithelium is not, as the adenoma has acquired features of the intestinal epithelium which is sensitive to Myc loss. However, the exact molecular mechanism behind this observation has yet to be identified and requires further comparative experiments to delineate the role of Wnt/Fzd/Myc in both the normal and transformed tissue of the stomach and intestine.
Author Contributions
Conception and design—D.F., E.V. and T.P. Methodology—D.F., N.B., E.V. and T.P. Data acquisition—D.F., E.V. and T.P. Data analysis—D.F., N.B., E.V. and T.P. Writing, review, editing and submission—D.F., N.B., E.V. and T.P. Supervision—M.E., E.V. and T.P.
Funding
Funding is gratefully acknowledged from the following; National Health and Medical Research Council of Australia (NHMRC, 566679 and APP1099302 awarded to E.V./T.P./N.B.), Melbourne Health project grants (605030 and PG-002 awarded to E.V./T.P./N.B.), Medical Research Council (MR/R026424/1 to T.P.), and Early career researcher grant (GIA-033 awarded to D.F.), Cancer Council of Victoria project grants (CCV, APP1020716 awarded to E.V./T.P./N.B.), CCV Fellowship awarded to D.F. and Cardiff University/CMU Research Fellowship awarded to T.P.
Acknowledgments
We thank Damian Neate and Jean Moselen for technical assistance. We also thank Cameron Nowell, Matthias Ernst and Owen Sansom for gifting mutant mice and microscopy assistance.
Conflicts of Interest
The authors have no conflicts of interests to declare. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
References
- Barker, N.; Van Es, J.H.; Kuipers, J.; Kujala, P.; Van den Born, M.; Cozijnsen, M.; Haegebarth, A.; Korving, J.; Begthel, H.; Peters, P.J.; et al. Identification of stem cells in small intestine and colon by marker gene Lgr5. Nature 2007, 449, 1003–1007. [Google Scholar] [CrossRef] [PubMed]
- Barker, N.; Huch, M.; Kujala, P.; van de Wetering, M.; Snippert, H.J.; van Es, J.H.; Sato, T.; Stange, D.E.; Begthel, H.; van den Born, M.; et al. Lgr5(+ve) stem cells drive self-renewal in the stomach and build long-lived gastric units in vitro. Cell Stem Cell 2010, 6, 25–36. [Google Scholar] [CrossRef] [PubMed]
- Leushacke, M.; Tan, S.H.; Wong, A.; Swathi, Y.; Hajamohideen, A.; Tan, L.T.; Goh, J.; Wong, E.; Denil, S.L.I.J.; Murakami, K.; et al. Lgr5-Expressing Chief Cells Drive Epithelial Regeneration and Cancer in the Oxyntic Stomach. Nat. Cell Biol. 2017, 19, 774–786. [Google Scholar] [CrossRef] [PubMed]
- Jaks, V.; Barker, N.; Kasper, M.; Van Es, J.H.; Snippert, H.J.; Clevers, H.; Toftgård, R. Lgr5 marks cycling, yet long-lived, hair follicle stem cells. Nat. Genet. 2008, 40, 1291–1299. [Google Scholar] [CrossRef] [PubMed]
- Ng, A.; Tan, S.; Singh, G.; Rizk, P.; Swathi, Y.; Tan, T.Z.; Huang, R.Y.-J.; Leushacke, M.; Barker, N. Lgr5 marks stem/progenitor cells in ovary and tubal epithelia. Nat. Cell Biol. 2014, 16, 745–757. [Google Scholar] [CrossRef] [PubMed]
- de Visser, K.E.; Ciampricotti, M.; Michalak, E.M.; Tan, D.W.M.; Speksnijder, E.N.; Hau, C.S.; Clevers, H.; Barker, N.; Jonkers, J. Developmental stage-specific contribution of LGR5(+) cells to basal and luminal epithelial lineages in the postnatal mammary gland. J. Pathol. 2012, 228, 300–309. [Google Scholar] [CrossRef]
- Barker, N.; Rookmaaker, M.B.; Kujala, P.; Ng, A.; Leushacke, M.; Snippert, H.; de Wetering, M.; Tan, S.; Van Es, J.H.; Huch, M. Lgr5(+ve) stem/progenitor cells contribute to nephron formation during kidney development. Cell Rep. 2012, 2, 540–552. [Google Scholar] [CrossRef]
- de Lau, W.; Barker, N.; Low, T.Y.; Koo, B.K.; Li, V.S.; Teunissen, H.; Kujala, P.; Haegebarth, A.; Peters, P.J.; van de Wetering, M.; et al. Lgr5 homologues associate with Wnt receptors and mediate R-spondin signalling. Nature 2011, 476, 293–297. [Google Scholar] [CrossRef]
- Nusse, R.; Clevers, H. Wnt/beta-Catenin Signaling, Disease, and Emerging Therapeutic Modalities. Cell 2017, 169, 985–999. [Google Scholar] [CrossRef]
- Janda, C.Y.; Waghray, D.; Levin, A.M.; Thomas, C.; Garcia, K.C. Structural basis of Wnt recognition by Frizzled. Science 2012, 337, 59–64. [Google Scholar] [CrossRef]
- Wiese, K.E.; Nusse, R.; van Amerongen, R. Wnt signalling: conquering complexity. Development 2018, 145, dev165902. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sato, T.; Van Es, J.H.; Snippert, H.J.; Stange, D.E.; Vries, R.G.; Van Den Born, M.; Barker, N.; Shroyer, N.F.; van de Wetering, M.; Clevers, H. Paneth cells constitute the niche for Lgr5 stem cells in intestinal crypts. Nature 2011, 469, 415–418. [Google Scholar] [CrossRef] [PubMed]
- Farin, H.F.; van Es, J.H.; Clevers, H. Redundant sources of Wnt regulate intestinal stem cells and promote formation of Paneth cells. Gastroenterology 2012, 143, 1518–1529. [Google Scholar] [CrossRef] [PubMed]
- Flanagan, D.J.; Phesse, T.J.; Barker, N.; Schwab, R.H.; Amin, N.; Malaterre, J.; Stange, D.E.; Nowell, C.J.; Currie, S.A.; Saw, J.T.S.; et al. Frizzled7 Functions as a Wnt Receptor in Intestinal Epithelial Lgr5(+) Stem Cells. Stem Cell Rep. 2015, 4, 759–767. [Google Scholar] [CrossRef] [PubMed]
- Flanagan, D.J.; Barker, N.; Nowell, C.; Clevers, H.; Ernst, M.; Phesse, T.J.; Vincan, E. Loss of the Wnt receptor Frizzled 7 in the gastric epithelium is deleterious and triggers rapid repopulation in vivo. Dis. Model Mech. 2017, 10, 971–980. [Google Scholar] [CrossRef] [PubMed]
- Flanagan, D.J.; Barker, N.; Di Costanzo, N.S.; Mason, E.A.; Gurney, A.; Meniel, V.S.; Koushyar, S.; Austin, C.R.; Ernst, M.; Pearson, H.B.; et al. Frizzled-7 Is Required for Wnt Signaling in Gastric Tumors with and Without Apc Mutations. Cancer Res. 2019, 79, 970–981. [Google Scholar] [CrossRef] [PubMed]
- Muncan, V.; Sansom, O.J.; Tertoolen, L.; Phesse, T.J.; Begthel, H.; Sancho, E.; Cole, A.M.; Gregorieff, A.; de Alboran, I.M.; Clevers, H.; et al. Rapid loss of intestinal crypts upon conditional deletion of the Wnt/Tcf-4 target gene c-Myc. Mol. Cell. Biol. 2006, 26, 8418–8426. [Google Scholar] [CrossRef]
- Ashton, G.H.; Morton, J.P.; Myant, K.; Phesse, T.J.; Ridgway, R.A.; Marsh, V.; Wilkins, J.A.; Athineos, D.; Muncan, V.; Kemp, R.; et al. Focal adhesion kinase is required for intestinal regeneration and tumorigenesis downstream of Wnt/c-Myc signaling. Dev. Cell 2010, 19, 259–269. [Google Scholar] [CrossRef]
- Sansom, O.J.; Meniel, V.S.; Muncan, V.; Phesse, T.J.; Wilkins, J.A.; Reed, K.R.; Vass, J.K.; Athineos, D.; Clevers, H.; Clarke, A.R. Myc deletion rescues Apc deficiency in the small intestine. Nature 2007, 446, 676–679. [Google Scholar] [CrossRef]
- Thiem, S.; Eissmann, M.F.; Elzer, J.; Jonas, A.; Putoczki, T.L.; Poh, A.; Nguyen, P.; Preaudet, A.; Flanagan, D.; Vincan, E. Stomach-Specific Activation of Oncogenic KRAS and STAT3-Dependent Inflammation Cooperatively Promote Gastric Tumorigenesis in a Preclinical Model. Cancer Res. 2016, 76, 2277–2287. [Google Scholar] [CrossRef] [Green Version]
- Bettess, M.D.; Dubois, N.; Murphy, M.J.; Dubey, C.; Roger, C.; Robine, S.; Trumpp, A. c-Myc is required for the formation of intestinal crypts but dispensable for homeostasis of the adult intestinal epithelium. Mol. Cell Biol. 2005, 25, 7868–7878. [Google Scholar] [CrossRef] [PubMed]
- Soriano, P. Generalized lacZ expression with the ROSA26 Cre reporter strain. Nat. Genet. 1999, 21, 70–71. [Google Scholar] [CrossRef] [PubMed]
- Broutier, L.; Andersson-Rolf, A.; Hindley, C.J.; Boj, S.F.; Clevers, H.; Koo, B.K.; Huch, M. Culture and establishment of self-renewing human and mouse adult liver and pancreas 3D organoids and their genetic manipulation. Nat. Protoc. 2016, 11, 1724–1743. [Google Scholar] [CrossRef] [PubMed]
- Flanagan, D.J.; Schwab, R.H.; Tran, B.M.; Phesse, T.J.; Vincan, E. Isolation and Culture of Adult Intestinal, Gastric, and Liver Organoids for Cre-recombinase-Mediated Gene Deletion. Methods Mol. Biol. 2016. [Google Scholar] [CrossRef]
- Vincan, E.; Darcy, P.K.; Farrelly, C.A.; Faux, M.C.; Brabletz, T.; Ramsay, R.G. Frizzled-7 dictates three-dimensional organization of colorectal cancer cell carcinoids. Oncogene 2007, 26, 2340–2352. [Google Scholar] [CrossRef] [PubMed]
- Wong, V.W.; Stange, D.E.; Page, M.E.; Buczacki, S.; Wabik, A.; Itami, S.; van de Wetering, M.; Poulsom, R.; Wright, N.A.; Trotter, M.W.B.; et al. Lrig1 controls intestinal stem-cell homeostasis by negative regulation of ErbB signalling. Nat. Cell Biol. 2012, 14, 401–408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arnold, K.; Sarkar, A.; Yram, M.A.; Polo, J.M.; Bronson, R.; Sengupta, S.; Seandel, M.; Geijsen, N.; Hochedlinger, K. Sox2(+) adult stem and progenitor cells are important for tissue regeneration and survival of mice. Cell Stem Cell 2011, 9, 317–329. [Google Scholar] [CrossRef]
- Hayakawa, Y.; Jin, G.; Wang, H.; Chen, X.; Westphalen, C.B.; Asfaha, S.; Renz, B.W.; Ariyama, H.; Dubeykovskaya, Z.K.; Takemoto, Y.; et al. CCK2R identifies and regulates gastric antral stem cell states and carcinogenesis. Gut 2015, 64, 544–553. [Google Scholar] [CrossRef]
- Flanagan, D.J.; Austin, C.; Vincan, E.; Phesse, T. Wnt Signalling in Gastrointestinal Epithelial Stem Cells. Genes (Basel) 2018, 9, 178. [Google Scholar] [CrossRef]
- Sigal, M.; Logan, C.Y.; Kapalczynska, M.; Mollenkopf, H.J.; Berger, H.; Wiedenmann, B.; Nusse, R.; Amieva, M.R.; Meyer, T.F. Stromal R-spondin orchestrates gastric epithelial stem cells and gland homeostasis. Nature 2017, 548, 451–455. [Google Scholar] [CrossRef]
- Geng, Y.; Lu, X.; Wu, X.; Xue, L.; Wang, X.; Xu, J. MicroRNA-27b suppresses Helicobacter pylori-induced gastric tumorigenesis through negatively regulating Frizzled7. Oncol. Rep. 2016, 35, 2441–2450. [Google Scholar] [CrossRef] [PubMed]
- Phesse, T.J.; Myant, K.B.; Cole, A.M.; Ridgway, R.A.; Pearson, H.; Muncan, V.; van den Brink, G.R.; Vousden, K.H.; Sears, R.; Vassilev, L.T. Endogenous c-Myc is essential for p53-induced apoptosis in response to DNA damage in vivo. Cell Death Differ. 2014, 21, 956–966. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burke, Z.D.; Reed, K.R.; Phesse, T.J.; Sansom, O.J.; Clarke, A.R.; Tosh, D. Liver Zonation Occurs Through a beta-Catenin-Dependent, c-Myc-Independent Mechanism. Gastroenterology 2009, 136, 2316–2324. [Google Scholar] [CrossRef] [PubMed]
- Reed, K.R.; Athineos, D.; Meniel, V.S.; Wilkins, J.A.; Ridgway, R.A.; Burke, Z.D.; Muncan, V.; Clarke, A.R.; Sansom, O.J. B-catenin deficiency, but not Myc deletion, suppresses the immediate phenotypes of APC loss in the liver. Proc. Natl. Acad. Sci. USA 2008, 105, 18919–18923. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Figure 1.
Conditional deletion of Fzd7 from Lgr5+ gastric stem cells does not perturb homeostasis. (A) X-gal stained tissue sections from Lgr5Cre+; LacZ and Lgr5Cre+; Fzd7fl/fl; LacZ mice 3 and 30 days following tamoxifen. Scale bars = 100 μm. (B) Conventional PCR for recombined Fzd7 (Fzd7Δ) alleles in mice described in (A). (C) Representative IHC staining for GFP, which permits detection of Lgr5GFP+ stem cells, on sections from mice described in A at 30 days post tamoxifen. Scale bars = 100 µm. (D) Quantification of GFP+ cells in sections from mice described in A (mean ± SEM, n = 3 mice, at least 30 glands scored/mouse, Unpaired student t-test). (E) qPCR for Fzd receptors in FACS-isolated GFP+ (Lgr5hi) cells. Expression was normalized to Lgr5lo cells.
Figure 1.
Conditional deletion of Fzd7 from Lgr5+ gastric stem cells does not perturb homeostasis. (A) X-gal stained tissue sections from Lgr5Cre+; LacZ and Lgr5Cre+; Fzd7fl/fl; LacZ mice 3 and 30 days following tamoxifen. Scale bars = 100 μm. (B) Conventional PCR for recombined Fzd7 (Fzd7Δ) alleles in mice described in (A). (C) Representative IHC staining for GFP, which permits detection of Lgr5GFP+ stem cells, on sections from mice described in A at 30 days post tamoxifen. Scale bars = 100 µm. (D) Quantification of GFP+ cells in sections from mice described in A (mean ± SEM, n = 3 mice, at least 30 glands scored/mouse, Unpaired student t-test). (E) qPCR for Fzd receptors in FACS-isolated GFP+ (Lgr5hi) cells. Expression was normalized to Lgr5lo cells.

Figure 2.
Conditional deletion of Myc from the gastric epithelium does not trigger epithelial repopulation. (A) X-gal stained tissue sections from Lgr5Cre+; LacZ and Lgr5Cre+; Mycfl/fl; LacZ mice 3 and 30 days following tamoxifen. Scale bars = 100 μm. (B) qRT-PCR for c-Myc on gastric epithelial cells isolated from Lgr5Cre+; Myc+/+ and Lgr5Cre+; Mycfl/fl mice following tamoxifen at 3 and 30 days post tamoxifen as indicated (* p < 0.05, mean ± SEM, n = 3 mice, Two-way ANOVA). (C) Representative IHC for GFP, which permits detection of Lgr5GFP+ stem cells, on sections from mice described in A. Scale bars = 100 µm. (D) Quantification of GFP+ cells in sections from mice described in A (mean ±SEM, n = 3 mice, at least 30 glands scored/mouse, unpaired student t-test). (E) X-gal stained tissue sections from Tff1Cre+; LacZ and Tff1Cre+; Mycfl/fl; LacZ mice 3 and 30 days following tamoxifen. Scale bars = 100 μm. (F) qRT-PCR for c-Myc on gastric epithelial cells isolated from Tff1Cre-; Mycfl/fl and Tff1Cre+; Mycfl/fl mice following tamoxifen (* p < 0.05, mean ± SEM, n = 3 mice, two-way ANOVA). (G) Conventional PCR for recombined Myc (MycΔ) alleles in mice described in (E). (H) Representative DIC images of gastric organoids derived from Tff1Cre+; Mycfl/fl mice 5 days after treatment with vehicle (EtOH) or tamoxifen (4-OHT). Scale bars = 100 µm. (I) MTT viability assay performed on organoid cultures described in (G) (mean ±SEM, n = 3 biological replicates, unpaired student t-test). Individual experiments were repeated twice. (J) Conventional PCR for recombined Myc (MycΔ) alleles on organoid cultures described in (H).
Figure 2.
Conditional deletion of Myc from the gastric epithelium does not trigger epithelial repopulation. (A) X-gal stained tissue sections from Lgr5Cre+; LacZ and Lgr5Cre+; Mycfl/fl; LacZ mice 3 and 30 days following tamoxifen. Scale bars = 100 μm. (B) qRT-PCR for c-Myc on gastric epithelial cells isolated from Lgr5Cre+; Myc+/+ and Lgr5Cre+; Mycfl/fl mice following tamoxifen at 3 and 30 days post tamoxifen as indicated (* p < 0.05, mean ± SEM, n = 3 mice, Two-way ANOVA). (C) Representative IHC for GFP, which permits detection of Lgr5GFP+ stem cells, on sections from mice described in A. Scale bars = 100 µm. (D) Quantification of GFP+ cells in sections from mice described in A (mean ±SEM, n = 3 mice, at least 30 glands scored/mouse, unpaired student t-test). (E) X-gal stained tissue sections from Tff1Cre+; LacZ and Tff1Cre+; Mycfl/fl; LacZ mice 3 and 30 days following tamoxifen. Scale bars = 100 μm. (F) qRT-PCR for c-Myc on gastric epithelial cells isolated from Tff1Cre-; Mycfl/fl and Tff1Cre+; Mycfl/fl mice following tamoxifen (* p < 0.05, mean ± SEM, n = 3 mice, two-way ANOVA). (G) Conventional PCR for recombined Myc (MycΔ) alleles in mice described in (E). (H) Representative DIC images of gastric organoids derived from Tff1Cre+; Mycfl/fl mice 5 days after treatment with vehicle (EtOH) or tamoxifen (4-OHT). Scale bars = 100 µm. (I) MTT viability assay performed on organoid cultures described in (G) (mean ±SEM, n = 3 biological replicates, unpaired student t-test). Individual experiments were repeated twice. (J) Conventional PCR for recombined Myc (MycΔ) alleles on organoid cultures described in (H).


© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Flanagan, D.; Barker, N.; Ernst, M.; Vincan, E.; Phesse, T. The Function of Lgr5+ Cells in the Gastric Antrum Does Not Require Fzd7 or Myc In Vivo. Biomedicines 2019, 7, 50. https://doi.org/10.3390/biomedicines7030050
AMA Style
Flanagan D, Barker N, Ernst M, Vincan E, Phesse T. The Function of Lgr5+ Cells in the Gastric Antrum Does Not Require Fzd7 or Myc In Vivo. Biomedicines. 2019; 7(3):50. https://doi.org/10.3390/biomedicines7030050
Chicago/Turabian StyleFlanagan, Dustin, Nick Barker, Matthias Ernst, Elizabeth Vincan, and Toby Phesse. 2019. "The Function of Lgr5+ Cells in the Gastric Antrum Does Not Require Fzd7 or Myc In Vivo" Biomedicines 7, no. 3: 50. https://doi.org/10.3390/biomedicines7030050
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.