S-Methylcysteine (SMC) Ameliorates Intestinal, Hepatic, and Splenic Damage Induced by Cryptosporidium parvum Infection Via Targeting Inflammatory Modulators and Oxidative Stress in Swiss Albino Mice

 ,
,  ,
,  , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethical Considerations
2.2. Materials
2.3. Parasite Material and Parasite Preparation
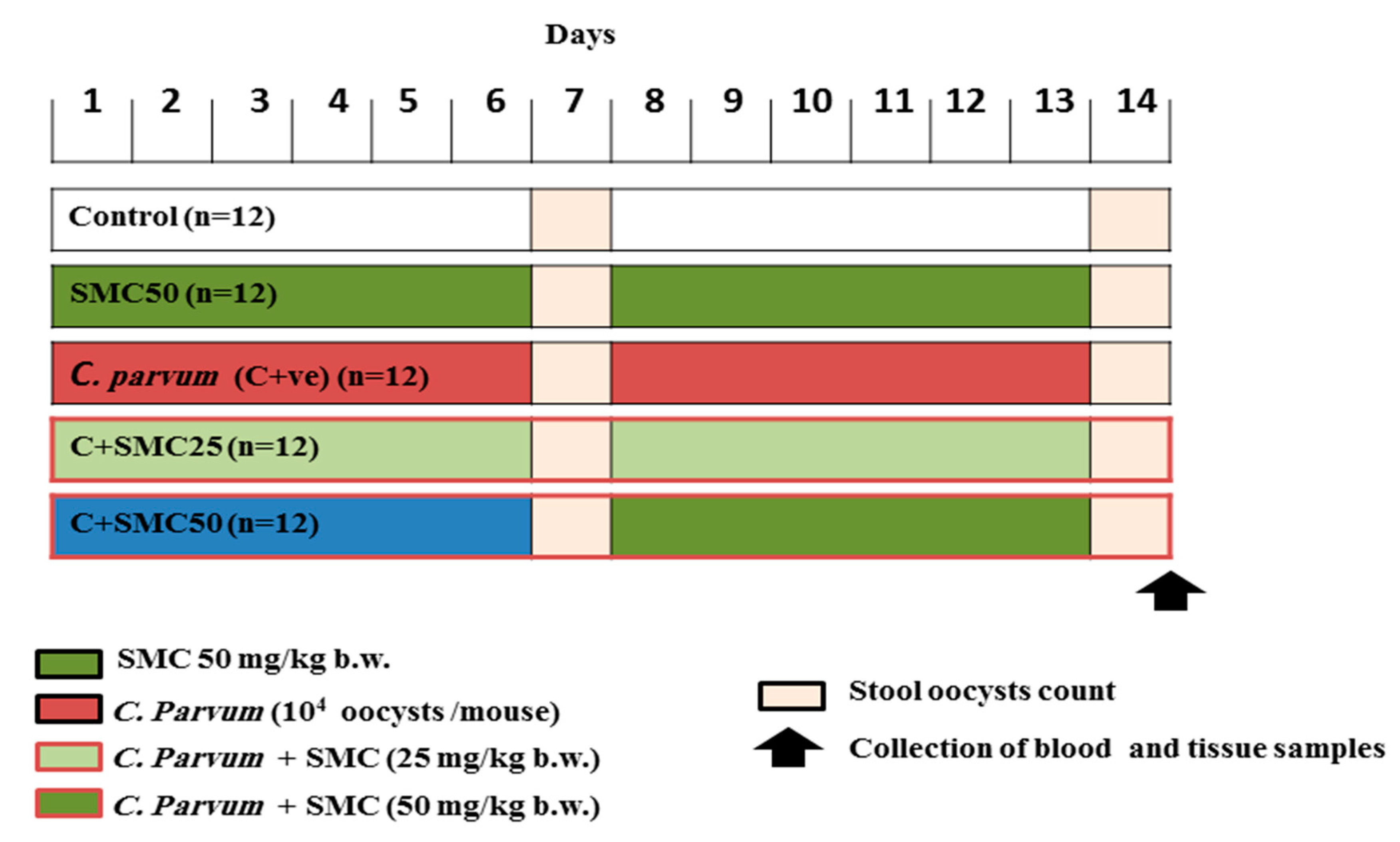
2.4. Animals and Experimental Protocol
2.5. Measurement of Serum Biochemical Parameters
2.6. Histopathological Examination
2.7. Evaluation of Oxidative Stress and Antioxidant Markers
2.8. Statistical Analysis
3. Results
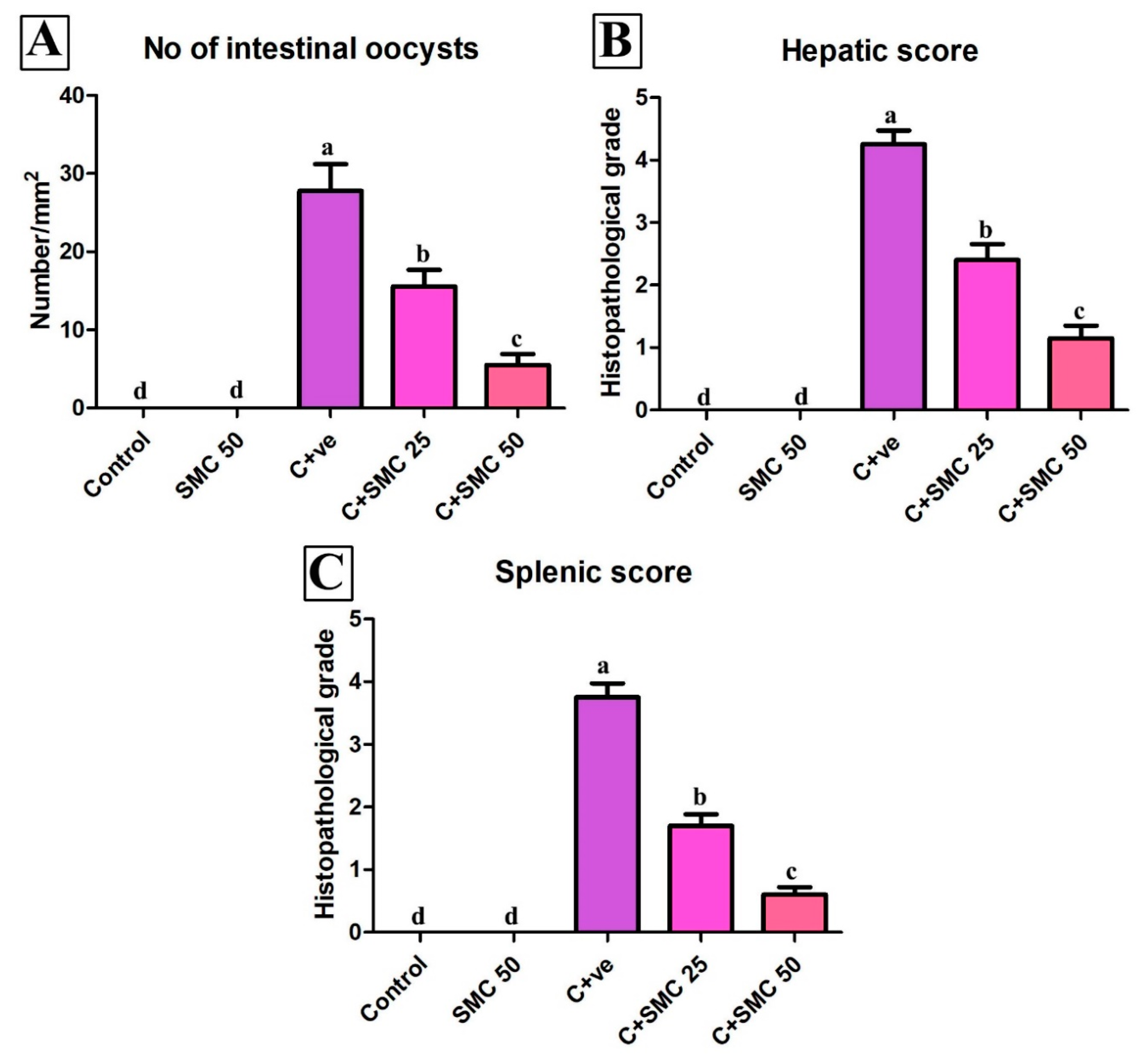
3.1. Effects of s-Methylcysteine (SMC) on Shedding of Cryptosporidium Oocysts
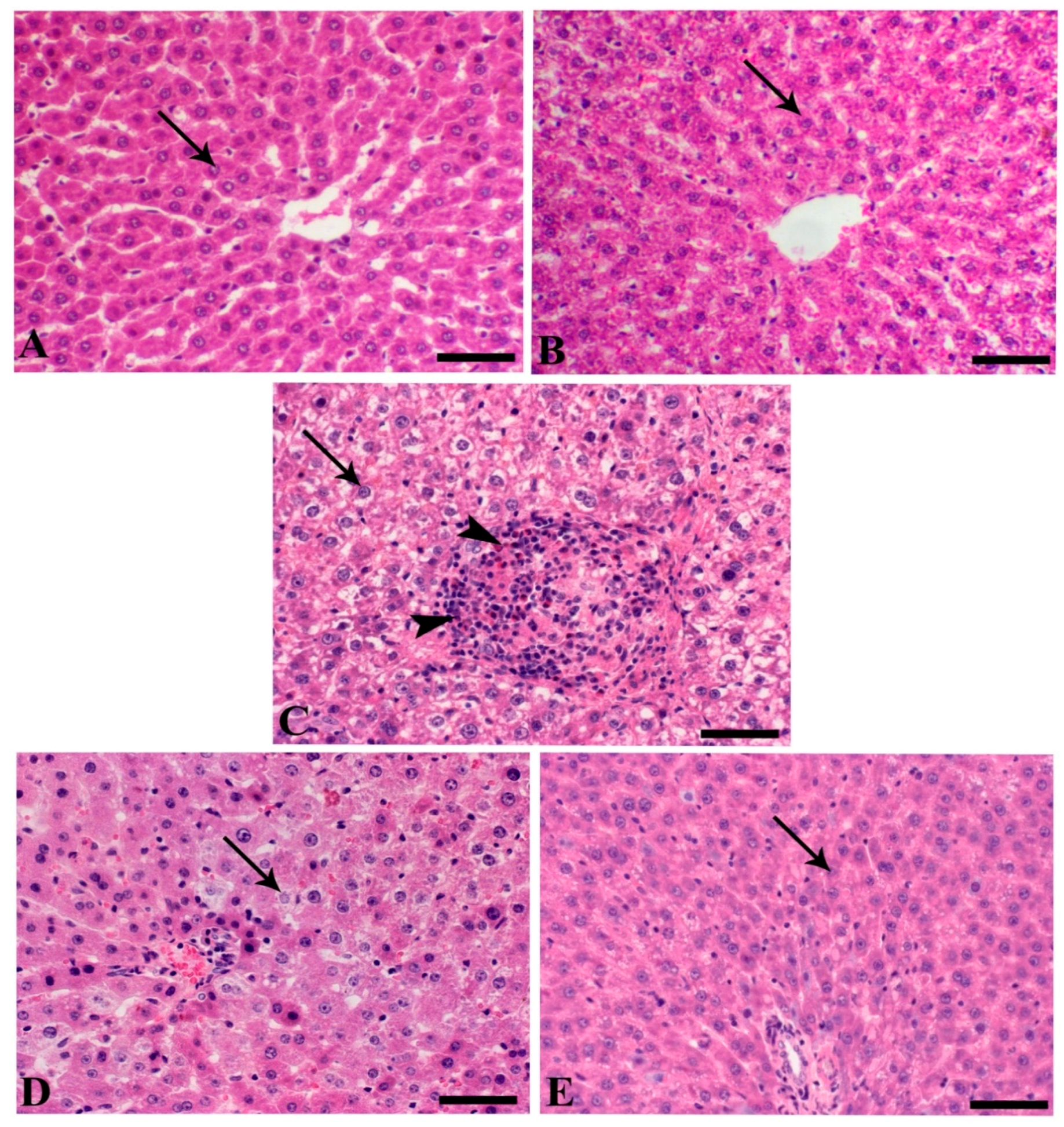
3.2. SMC Attenuates Cryptosporidium-Induced Liver Damage
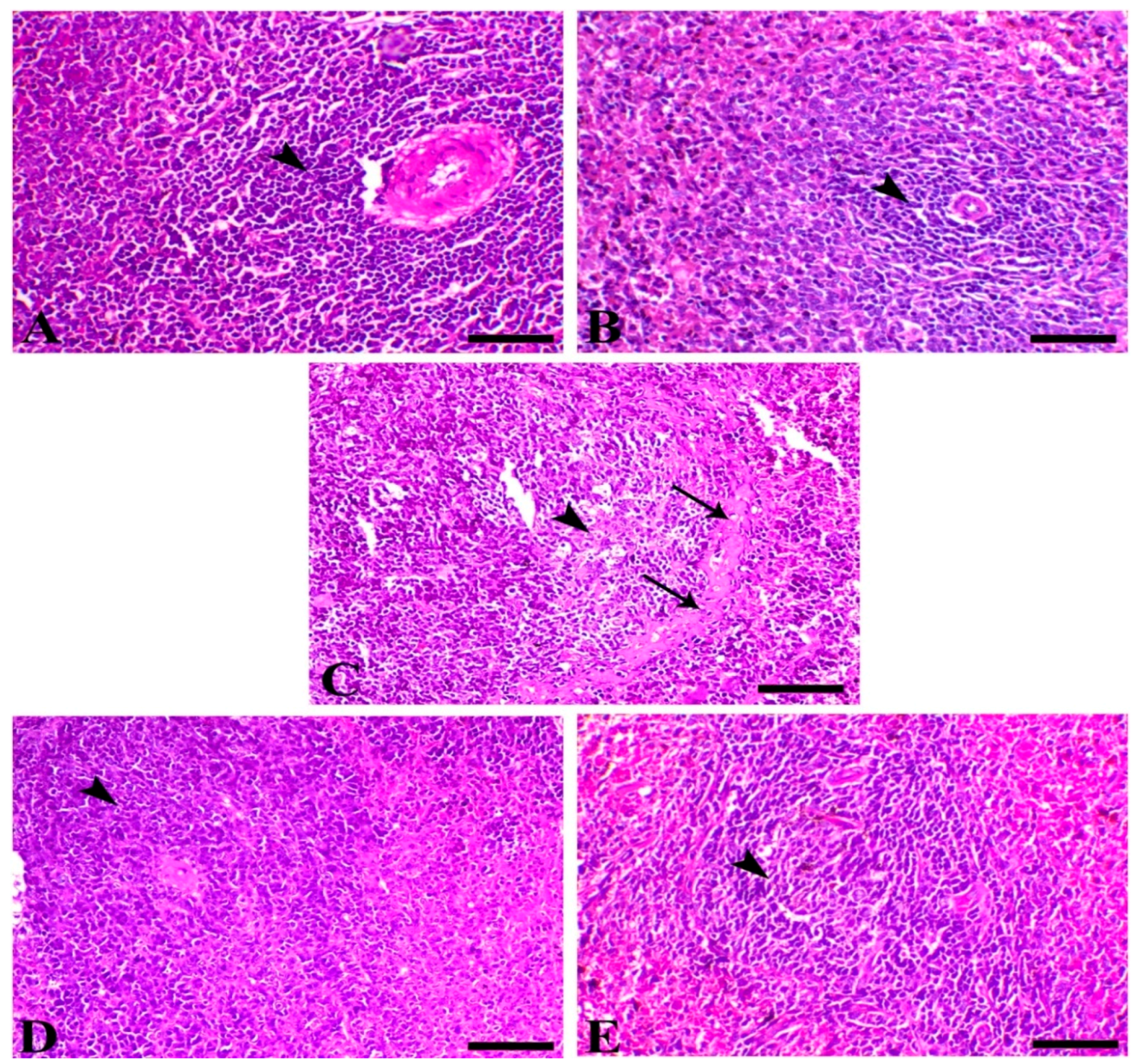
3.3. SMC Modulates Cryptosporidium-Induced Inflammation
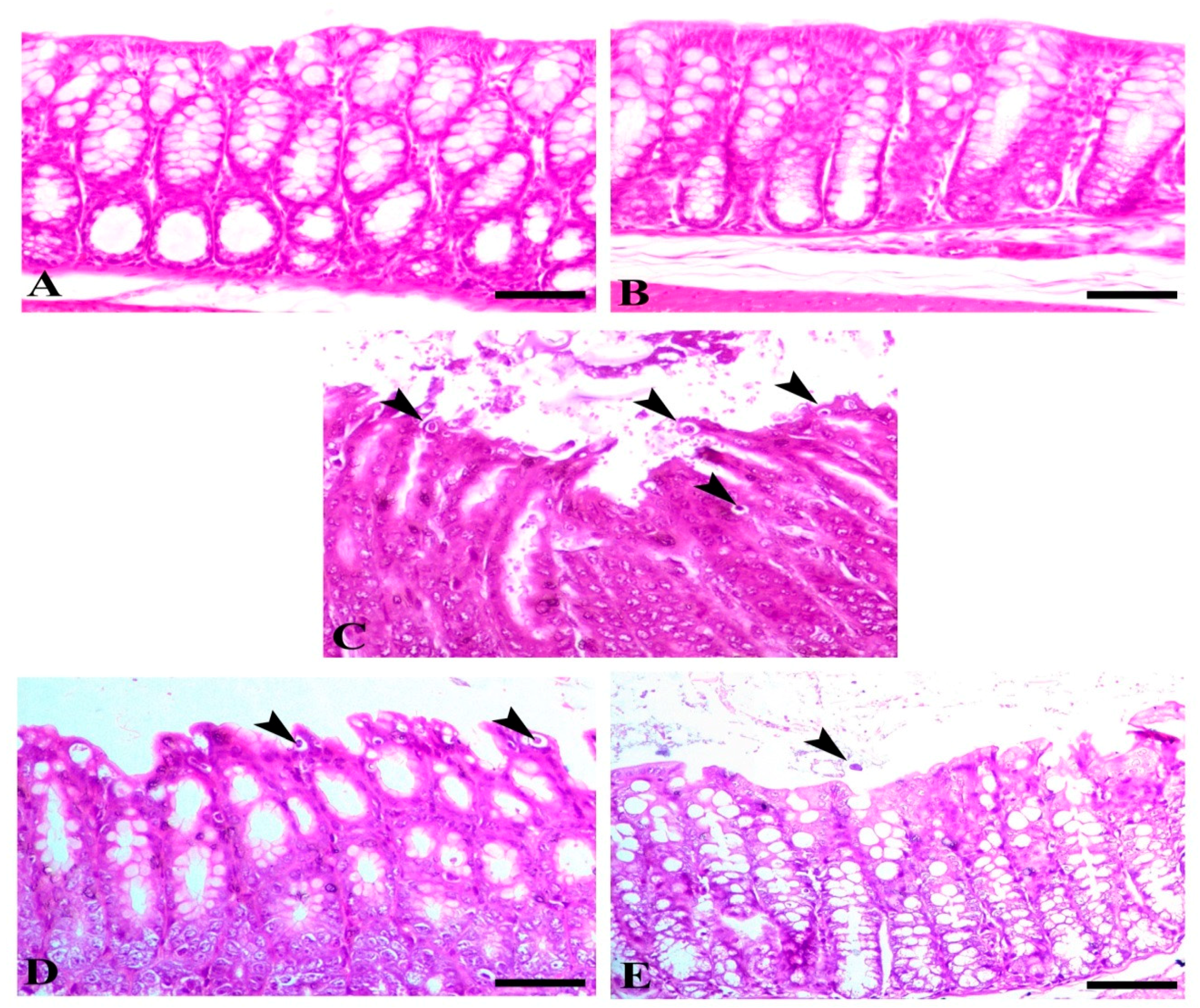
3.4. Histopathology
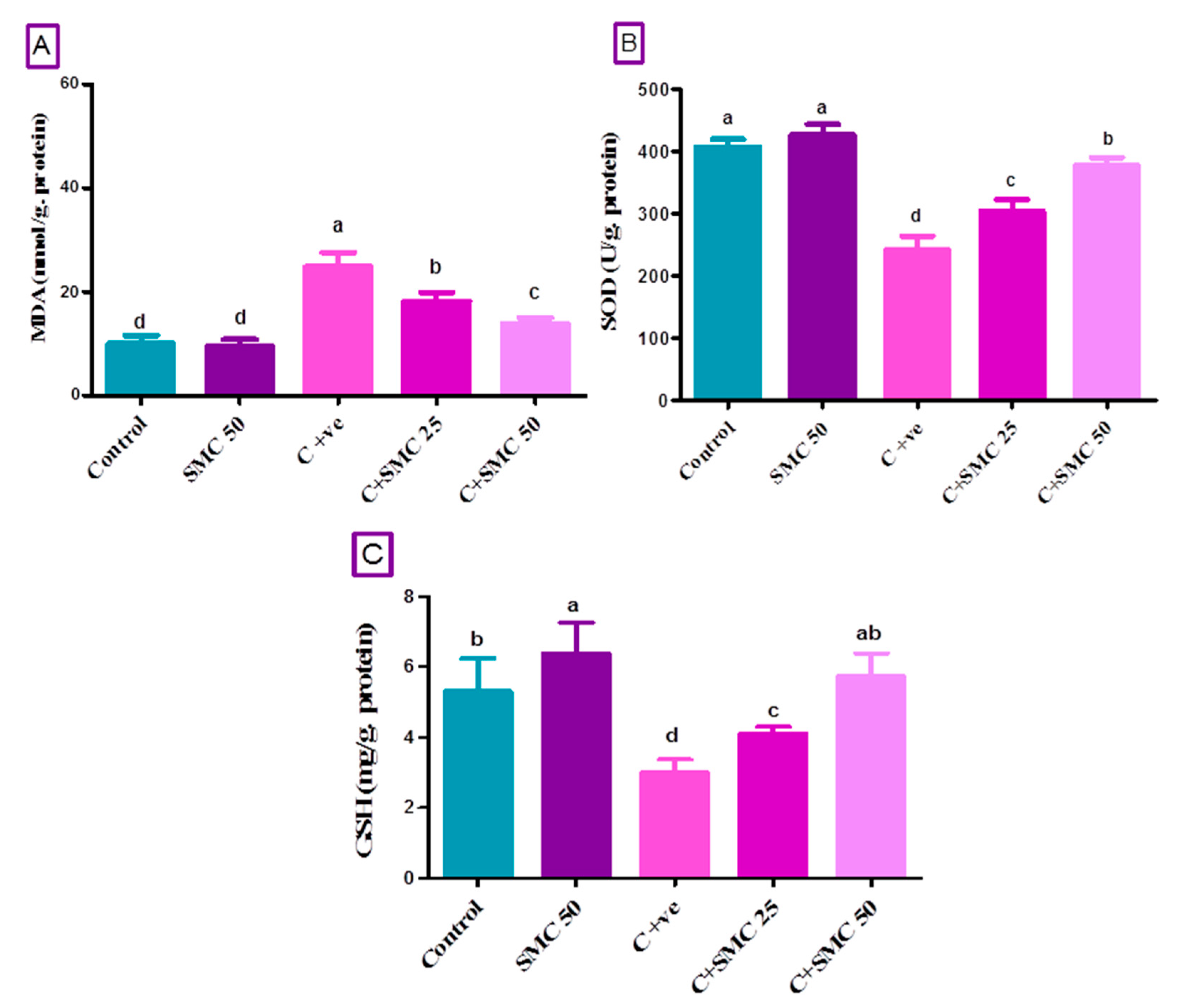
3.5. SMC Restores Intestinal Oxidative Injury Generated by Cryptosporidium
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| Cryptosporidium parvum | C. parvum |
| SMC | s-Methylcysteine |
| SMC25 | s-Methylcysteine (25 mg/kg b.w.) |
| SMC50 | s-Methylcysteine (50 mg/kg b.w.) |
| PI | post-infection |
| ALT | alanine aminotransferase |
| AST | aspartate aminotransferase |
| ALP | alkaline phosphatase |
| SAP | serum amyloid P |
| TNF-α | tumor necrosis factor alpha. |
| IL-6 | Interleukin-6 |
| IFN-γ | Interferon gamma |
| MDA | malondialdehyde |
| GSH | glutathione |
| SOD | superoxide dismutase |
References
- Fletcher, S.M.; Stark, D.; Harkness, J.; Ellis, J. Enteric protozoa in the developed world: A public health perspective. Clin. Microbiol. Rev. 2012, 25, 420–449. [Google Scholar] [CrossRef] [Green Version]
- Sack, D.A.; Freij, L.; Holmgren, J. Prospects for Public Health Benefits in Developing Countries from New Vaccines against Enteric Infections. J. Infect. Dis. 1991, 163, 503–506. [Google Scholar] [CrossRef] [PubMed]
- McCormick, B.J.J.; Lang, D.R. Diarrheal disease and enteric infections in LMIC communities: How big is the problem? Trop. Dis. Travel. Med. Vaccines 2016, 2, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, L.; Ryan, U.M. Cryptosporidiosis: An update in molecular epidemiology. Curr. Opin. Infect. Dis. 2004, 17, 483–490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marshall, M.M.; Naumovitz, D.; Ortega, Y.; Sterling, C.R. Waterborne protozoan pathogens. Clin. Microbiol. Rev. 1997, 10, 67–85. [Google Scholar] [CrossRef] [PubMed]
- Pumipuntu, N.; Piratae, S. Cryptosporidiosis: A zoonotic disease concern. Vet. World 2018, 11, 681–686. [Google Scholar] [CrossRef] [Green Version]
- Fayer, R. Taxonomy and species delimitation in Cryptosporidium. Exp. Parasitol. 2010, 124, 90–97. [Google Scholar] [CrossRef]
- Fayer, R. Cryptosporidium: A water-borne zoonotic parasite. Vet. Parasitol. 2004, 126, 37–56. [Google Scholar] [CrossRef]
- Chen, X.M.; Keithly, J.S.; Paya, C.V.; LaRusso, N.F. Cryptosporidiosis. N. Engl. J. Med. 2002, 346. [Google Scholar] [CrossRef]
- Shane, A.L.; Mody, R.K.; Crump, J.A.; Tarr, P.I.; Steiner, T.S.; Kotloff, K.; Langley, J.M.; Wanke, C.; Warren, C.A.; Cheng, A.C. 2017 Infectious Diseases Society of America clinical practice guidelines for the diagnosis and management of infectious diarrhea. Clin. Infect. Dis. 2017, 65, e45–e80. [Google Scholar] [CrossRef] [Green Version]
- Bouzid, M.; Kintz, E.; Hunter, P.R. Risk factors for Cryptosporidium infection in low and middle income countries: A systematic review and meta-analysis. PLoS Negl. Trop. Dis. 2018, 12, e0006553. [Google Scholar] [CrossRef] [PubMed]
- Mor, S.M.; Tzipori, S. Cryptosporidiosis in children in Sub-Saharan Africa: A lingering challenge. Clin. Infect. Dis. 2008, 47, 915–921. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shirley, D.A.; Moonah, S.N.; Kotloff, K.L. Burden of disease from cryptosporidiosis. Curr. Opin. Infect. Dis. 2012, 25, 555–563. [Google Scholar] [CrossRef] [PubMed]
- Khalil, I.A.; Troeger, C.; Rao, P.C.; Blacker, B.F.; Brown, A.; Brewer, T.G.; Colombara, D.V.; De Hostos, E.L.; Engmann, C.; Guerrant, R.L.; et al. Morbidity, mortality, and long-term consequences associated with diarrhoea from Cryptosporidium infection in children younger than 5 years: A meta-analyses study. Lancet Glob. Health 2018, 6, e758–e768. [Google Scholar] [CrossRef] [Green Version]
- Shoultz, D.A.; de Hostos, E.L.; Choy, R.K. Addressing Cryptosporidium Infection among Young Children in Low-Income Settings: The Crucial Role of New and Existing Drugs for Reducing Morbidity and Mortality. PLoS Negl. Trop. Dis. 2016, 10, e0004242. [Google Scholar] [CrossRef] [Green Version]
- Rossle, N.F.; Latif, B. Cryptosporidiosis as threatening health problem: A review. Asian Pac. J. Trop. Biomed. 2013, 3, 916–924. [Google Scholar] [CrossRef]
- Gargala, G. Drug treatment and novel drug target against Cryptosporidium. Parasite 2008, 15, 275–281. [Google Scholar] [CrossRef] [Green Version]
- Chavez, M.A.; White, A.C., Jr. Novel treatment strategies and drugs in development for cryptosporidiosis. Expert Rev. Anti Infect. 2018, 16, 655–661. [Google Scholar] [CrossRef]
- Manjunatha, U.H.; Chao, A.T.; Leong, F.J.; Diagana, T.T. Cryptosporidiosis Drug Discovery: Opportunities and Challenges. ACS Infect. Dis. 2016, 2, 530–537. [Google Scholar] [CrossRef]
- Cabada, M.M.; White, A.C., Jr. Treatment of cryptosporidiosis: Do we know what we think we know? Curr. Opin. Infect. Dis. 2010, 23, 494–499. [Google Scholar] [CrossRef]
- Maw, G.A. Biochemistry of S-Methyl-L-Cysteine and its Principal Derivatives. Sulfur Rep. 1982, 2, 1–26. [Google Scholar] [CrossRef]
- Takada, N.; Yano, Y.; Wanibuchi, H.; Otani, S.; Fukushima, S. S-methylcysteine and cysteine are inhibitors of induction of glutathione S-transferase placental form-positive foci during initiation and promotion phases of rat hepatocarcinogenesis. Jpn. J. Cancer Res. 1997, 88, 435–442. [Google Scholar] [CrossRef]
- Amagase, H. Clarifying the real bioactive constituents of garlic. J. Nutr. 2006, 136, 716S–725S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Magd, M.A.; Abdo, W.S.; El-Maddaway, M.; Nasr, N.M.; Gaber, R.A.; El-Shetry, E.S.; Saleh, A.A.; Alzahrani, F.A.A.; Abdelhady, D.H. High doses of S-methylcysteine cause hypoxia-induced cardiomyocyte apoptosis accompanied by engulfment of mitochondaria by nucleus. Biomed. Pharm. 2017, 94, 589–597. [Google Scholar] [CrossRef]
- Swiderski, F.; Dabrowska, M.; Rusaczonek, A.; Waszkiewicz-Robak, B. Bioactive substances of garlic and their role in dietoprophylaxis and dietotherapy. Rocz. Państw. Zakł. 2007, 58, 41–46. [Google Scholar]
- Amagase, H.; Petesch, B.L.; Matsuura, H.; Kasuga, S.; Itakura, Y. Intake of garlic and its bioactive components. J. Nutr. 2001, 131, 955S–962S. [Google Scholar] [CrossRef]
- Vijayaraghavan, M.; Wanibuchi, H.; Takada, N.; Yano, Y.; Otani, S.; Yamamoto, S.; Fukushima, S. Inhibitory effects of S-methylcysteine and cysteine on the promoting potential of sodium phenobarbital on rat liver carcinogenesis. Jpn. J. Cancer Res. 2000, 91, 780–785. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.M.; Yin, M.C.; Hsu, C.C.; Liu, T.C. Antioxidative and anti-inflammatory effects of four cysteine-containing agents in striatum of MPTP-treated mice. Nutrition 2007, 23, 589–597. [Google Scholar] [CrossRef]
- Thomas, S.; Senthilkumar, G.P.; Sivaraman, K.; Bobby, Z.; Paneerselvam, S.; Harichandrakumar, K.T. Effect of s-methyl-L-cysteine on oxidative stress, inflammation and insulin resistance in male wistar rats fed with high fructose diet. Iran. J. Med. Sci 2015, 40, 45–50. [Google Scholar]
- Senthilkumar, G.P.; Thomas, S.; Sivaraman, K.; Sankar, P.; Bobby, Z. Study the effect of s-methyl L-cysteine on lipid metabolism in an experimental model of diet induced obesity. J. Clin. Diagn. Res. 2013, 7, 2449–2451. [Google Scholar] [CrossRef] [PubMed]
- Kosuge, Y. Neuroprotective mechanisms of S-allyl-L-cysteine in neurological disease. Exp. Med. 2020, 19, 1565–1569. [Google Scholar] [CrossRef]
- Hazaa, I.; Al-Taai, N.; Khalil, N.; Zakri, A. Efficacy of garlic and onion oils on murin experimental Cryptosporidium parvum infection. Al-Anbar J. Vet. Sci. 2016, 9, 69–74. [Google Scholar]
- Gaafar, M.R. Efficacy of Allium sativum (garlic) against experimental cryptosporidiosis. Alex. J. Med. 2012, 48, 59–66. [Google Scholar] [CrossRef]
- Galicia-Moreno, M.; Rodriguez-Rivera, A.; Reyes-Gordillo, K.; Segovia, J.; Shibayama, M.; Tsutsumi, V.; Vergara, P.; Moreno, M.G.; Muriel, P. N-acetylcysteine prevents carbon tetrachloride-induced liver cirrhosis: Role of liver transforming growth factor-beta and oxidative stress. Eur J. Gastroenterol. Hepatol. 2009, 21, 908–914. [Google Scholar] [CrossRef] [PubMed]
- Maksimchik, Y.Z.; Lapshina, E.A.; Sudnikovich, E.Y.; Zabrodskaya, S.V.; Zavodnik, I.B. Protective effects of N-acetyl-L-cysteine against acute carbon tetrachloride hepatotoxicity in rats. Cell Biochem. Funct. 2008, 26, 11–18. [Google Scholar] [CrossRef]
- Wei, M.; Wanibuchi, H.; Yamamoto, S.; Iwai, S.; Morimura, K.; Nomura, T.; Takayama, R.; Fukushima, S. Chemopreventive effects of S-methylcysteine on rat hepatocarcinogenesis induced by concurrent administration of sodium nitrite and morpholine. Cancer Lett. 2000, 161, 97–103. [Google Scholar] [CrossRef]
- Khalifa, A.M.; El Temsahy, M.M.; Abou El Naga, I.F. Effect of ozone on the viability of some protozoa in drinking water. J. Egypt Soc. Parasitol. 2001, 31, 603–616. [Google Scholar]
- Gaafar, M.R. Effect of solar disinfection on viability of intestinal protozoa in drinking water. J. Egypt Soc. Parasitol. 2007, 37, 65–86. [Google Scholar] [PubMed]
- Rasmussen, K.R.; Healey, M.C. Experimental Cryptosporidium parvum infections in immunosuppressed adult mice. Infect. Immun. 1992, 60, 1648–1652. [Google Scholar] [CrossRef] [Green Version]
- Bancroft, J.D.; Gamble, M. Theory and Practice of Histological Techniques, 6th ed.; Churchill Livingstone; Elsevier: London, UK, 2008. [Google Scholar]
- Ellman, G.L. Tissue sulfhydryl groups. Arch. Biochem. Biophys. 1959, 82, 70–77. [Google Scholar] [CrossRef]
- Innes, E.A.; Chalmers, R.M.; Wells, B.; Pawlowic, M.C. A One Health Approach to Tackle Cryptosporidiosis. Trends Parasitol. 2020, 36, 290–303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gerace, E.; Lo Presti, V.D.M.; Biondo, C. Cryptosporidium Infection: Epidemiology, Pathogenesis, and Differential Diagnosis. Eur J. Microbiol Immunol (Bp) 2019, 9, 119–123. [Google Scholar] [CrossRef] [PubMed]
- El-Saber Batiha, G.; Magdy Beshbishy, A.; Wasef, L.G.; Elewa, Y.H.; A. Al-Sagan, A.; El-Hack, A.; Mohamed, E.; Taha, A.E.; Abd-Elhakim, Y.M.; Prasad Devkota, H. Chemical constituents and pharmacological activities of garlic (Allium sativum L.): A review. Nutrients 2020, 12, 872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richard, S.; Tsuyoshi, K.; Cheol-Koo, L. Historical perspective on the use of garlic. J. Nutr. 2001, 131, 951–954. [Google Scholar]
- Abdel-Hafeez, E.H.; Ahmad, A.K.; Kamal, A.M.; Abdellatif, M.Z.; Abdelgelil, N.H. In vivo antiprotozoan e_ects of garlic (Allium sativum) and ginger (Zingiber officinale) extracts on experimentally infected mice with Blastocystis spp. Parasitol. Res. 2015, 114, 3439–3444. [Google Scholar] [CrossRef]
- AbouGabal, A.; Aboul-Ela, H.M.; Ali, E.; Khaled, A.E.M.; Shalaby, O.K. Hepatoprotective, DNA damage prevention and antioxidant potential of spirulina platensis on CCl4-induced hepatotoxicity in mice. Am. J. Biomed. Res. 2015, 3, 29–34. [Google Scholar]
- Fernandes, M.d.S.; Iano, F.G.; Rocia, V.; Yanai, M.M.; Leite, A.d.L.; Furlani, T.A.; Buzalaf, M.A.R.; Oliveira, R.C.d. Alkaline phosphatase activity in plasma and liver of rats submitted to chronic exposure to fluoride. Braz. Arch. Biol. Technol. 2011, 54, 1187–1192. [Google Scholar] [CrossRef] [Green Version]
- Aboelsoued, D.; Abo-Aziza, F.; Mahmoud, M.; Megeed, K.A.; El Ezz, N.A.; Abu-Salem, F. Anticryptosporidial effect of pomegranate peels water extract in experimentally infected mice with special reference to some biochemical parameters and antioxidant activity. J. Parasit. Dis. 2019, 43, 215–228. [Google Scholar] [CrossRef]
- Chalmers, R.M.; Davies, A.P. Minireview: Clinical cryptosporidiosis. Exp. Parasitol. 2010, 124, 138–146. [Google Scholar] [CrossRef]
- Bhagat, M.; Sood, S.; Yadav, A.; Verma, P.; Manzoor, N.; Chakraborty, D.; Katoch, R.; Sangha, N. Alterations in oxidative stress parameters and its associated correlation with clinical disease on experimental Cryptosporidium parvum infection in Swiss albino mice. J. Parasit. Dis. 2017, 41, 707–712. [Google Scholar] [CrossRef]
- Ruot, B.; Breuillé, D.; Rambourdin, F.; Bayle, G.; Capitan, P.; Obled, C. Synthesis rate of plasma albumin is a good indicator of liver albumin synthesis in sepsis. Am. J. Physiol. Endocrinol. Metab. 2000, 279, E244–E251. [Google Scholar] [CrossRef] [Green Version]
- Soufy, H.; Nadia, M.; Nasr, S.M.; Abd El-Aziz, T.H.; Khalil, F.A.; Ahmed, Y.F.; Abou Zeina, H.A. Effect of Egyptian propolis on cryptosporidiosis in immunosuppressed rats with special emphasis on oocysts shedding, leukogram, protein profile and ileum histopathology. Asian Pac. J. Trop. Med. 2017, 10, 253–262. [Google Scholar] [CrossRef]
- Mostafa, M.G.; Mima, T.; Ohnishi, S.T.; Mori, K. S-allylcysteine ameliorates doxorubicin toxicity in the heart and liver in mice. Planta Med. 2000, 66, 148–151. [Google Scholar] [CrossRef]
- Numagami, Y.; Ohnishi, S.T. S-allylcysteine inhibits free radical production, lipid peroxidation and neuronal damage in rat brain ischemia. J. Nutr. 2001, 131, 11005–11055. [Google Scholar] [CrossRef]
- Maldonado, P.D.; Barrera, D.; Rivero, I.; Mata, R.; Medina-Campos, O.N.; Hernández-Pando, R.; Pedraza-Chaverrí, J. Antioxidant S-allylcysteine prevents gentamicin-induced oxidative stress and renal damage. Free Radic. Biol. Med. 2003, 35, 317–324. [Google Scholar] [CrossRef]
- Pilling, D.; Gomer, R.H. The development of serum amyloid P as a possible therapeutic. Front. Immunol. 2018, 9, 2328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.-M.; An, J. Cytokines, inflammation and pain. Int. Anesthesiol. Clin. 2007, 45, 27. [Google Scholar] [CrossRef] [Green Version]
- Vartanian, T.; Li, Y.; Zhao, M.; Stefansson, K. Interferon-γ-Induced Oligodendrocyte Cell Death: Implications for the Pathogenesis of Multiple Sclerosis. Mol. Med. 1995, 1, 732–742. [Google Scholar] [CrossRef] [Green Version]
- Aliberti, J.; Cardoso, M.; Martins, G.A.; Gazzinelli, R.T.; Vieira, L.Q.; Silva, J.S. Interleukin-12 mediates resistance to Trypanosoma cruzi in mice and is produced by murine macrophages in response to live trypomastigotes. Infect. Immun. 1996, 64, 1961–1967. [Google Scholar] [CrossRef] [Green Version]
- Cardillo, F.; Voltarelli, J.C.; Reed, S.G.; Silva, J.S. Regulation of Trypanosoma cruzi infection in mice by gamma interferon and interleukin 10: Role of NK cells. Infect. Immun. 1996, 64, 128–134. [Google Scholar] [CrossRef] [Green Version]
- Kitani, H.; Yagi, Y.; Naessens, J.; Sekikawa, K.; Iraqi, F. The secretion of acute phase proteins and inflammatory cytokines during Trypanosoma congolense infection is not affected by the absence of the TNF-α gene. Acta Trop. 2004, 92, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Vasileiadou, K.; Pantazidis, G.; Papadopoulou, K.; Ligoudistianou, C.; Kourelis, A.; Petrakis, S.; Masmanidou, E.; Testa, T.; Kourounakis, A.; Hadjipetrou, L. α1-Acid glycoprotein production in rat dorsal air pouch in response to inflammatory stimuli, dexamethasone and honey bee venom. Exp. Mol. Pathol. 2010, 89, 63–71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Atwa, M.T.; Salem, N.I.; Mohamed, H.R. Effect of Cryptosporidium parvum infection on dna damage and cytokines gene expression in blood of immunocompromised mice. Egypt. J. Zool. 2020, 73, 16–25. [Google Scholar]
- Petry, F.; Jakobi, V.; Tessema, T.S. Host immune response to Cryptosporidium parvum infection. Exp. Parasitol. 2010, 126, 304–309. [Google Scholar] [CrossRef]
- Codices, V.; Martins, C.; Novo, C.; de Sousa, B.; Lopes, Â.; Borrego, M.; Matos, O. Dynamics of cytokines and immunoglobulins serum profiles in primary and secondary Cryptosporidium parvum infection: Usefulness of Luminex® xMAP technology. Exp. Parasitol. 2013, 133, 106–113. [Google Scholar] [CrossRef]
- Hsia, T.C.; Wang, Z.H.; Yin, M.C. s-Ethyl Cysteine and s-Methyl Cysteine Post-intake Attenuated LPS-induced Lung Injury in Mice. J. Adv. Med. Med. Res. 2017, 23, 1–10. [Google Scholar] [CrossRef]
- You, B.R.; Yoo, J.-M.; Baek, S.Y.; Kim, M.R. Anti-inflammatory effect of aged black garlic on 12-O-tetradecanoylphorbol-13-acetate-induced dermatitis in mice. Nutr. Res. Pract. 2019, 13, 189–195. [Google Scholar] [CrossRef]
- Masuno, K.; Yanai, T.; Sakai, H.; Satoh, M.; Kai, C.; Nakai, Y. Pathological features of Cryptosporidium andersoni-induced lesions in SCID mice. Exp. Parasitol. 2013, 134, 381–383. [Google Scholar] [CrossRef]
- Masuno, K.; Fukuda, Y.; Kubo, M.; Ikarashi, R.; Kuraishi, T.; Hattori, S.; Kimura, J.; Kai, C.; Yanai, T.; Nakai, Y. Infectivity of Cryptosporidium andersoni and Cryptosporidium muris to Normal and Immunosuppressive Cynomolgus Monkeys. J. Vet. Med Sci. Jpn. Soc. Vet. Sci. 2013, 76, 169–172. [Google Scholar] [CrossRef] [Green Version]
- Masuno, K.; Yanai, T.; Hirata, A.; Yonemaru, K.; Sakai, H.; Satoh, M.; Masegi, T.; Nakai, Y. Morphological and immunohistochemical features of Cryptosporidium andersoni in cattle. Vet. Pathol. 2006, 43, 202–207. [Google Scholar] [CrossRef] [Green Version]
- Yanai, T.; Simon, M.A.; Doddy, F.D.; Mansfield, K.G.; Pauley, D.; Lackner, A.A. Nodular Pneumocystis carinii pneumonia in SIV-infected macaques. Vet. Pathol 1999, 36, 471–474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Certad, G.; Ngouanesavanh, T.; Guyot, K.; Gantois, N.; Chassat, T.; Mouray, A.; Fleurisse, L.; Pinon, A.; Cailliez, J.-C.; Dei-Cas, E. Cryptosporidium parvum, a potential cause of colic adenocarcinoma. Infect. Agents Cancer 2007, 2, 22. [Google Scholar] [CrossRef]
- Abdou, A.G.; Harba, N.M.; Afifi, A.F.; Elnaidany, N.F. Assessment of Cryptosporidium parvum infection in immunocompetent and immunocompromised mice and its role in triggering intestinal dysplasia. Int. J. Infect. Dis. 2013, 17, e593–e600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ide, N.; Lau, B.H.S. Garlic compounds minimize intracellular oxidative stress and inhibit nuclear factor-kB activation. J. Nutr. 2001, 131, 1020–1026. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.M.; Chun, S.B.; Koo, M.S.; Choi, W.J.; Kim, T.W.; Kwon, Y.G.; Chung, H.T.; Billiar, T.R.; Kim, Y.M. Differential regulation of NO availability from macrophages and endothelial cells by the garlic component of S-allylcysteine. Free Radic. Biol. Med. 2001, 30, 747–756. [Google Scholar] [CrossRef]
- Keshavarzian, A.; Banan, A.; Farhadi, A.; Komanduri, S.; Mutlu, E.; Zhang, Y.; Fields, J. Increases in free radicals and cytoskeletal protein oxidation and nitration in the colon of patients with inflammatory bowel disease. Gut 2003, 52, 720–728. [Google Scholar] [CrossRef] [Green Version]
- Quílez, J.; Vergara-Castiblanco, C.; Monteagudo, L.; del Cacho, E.; Sánchez-Acedo, C. Host association of Cryptosporidium parvum populations infecting domestic ruminants in Spain. Appl. Environ. Microbiol. 2013, 79, 5363–5371. [Google Scholar] [CrossRef] [Green Version]
- Castro, M.C.; Francini, F.; Schinella, G.; Caldiz, C.I.; Zubiría, M.G.; Gagliardino, J.J.; Massa, M.L. Apocynin administration prevents the changes induced by a fructose-rich diet on rat liver metabolism and the antioxidant system. Clin. Sci. 2012, 123, 681–692. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Wu, Y.; Qin, J.; Sun, H.; He, H. Induced susceptibility of host is associated with an impaired antioxidant system following infection with Cryptosporidium parvum in Se-deficient mice. PLoS ONE 2009, 4, e4628. [Google Scholar] [CrossRef]
- Abd El-Aziz, T.H.; El-Beih, N.; Soufy, H.; Nasr, S.M.; Khalil, F.A.; Sharaf, M. Effect of Egyptian propolis on lipid profile and oxidative status in comparison with nitazoxanide in immunosuppressed rats infected with Cryptosporidium spp. Glob. Vet. 2014, 13, 17–27. [Google Scholar]
- Abo-Aziza, F.A.; Hendawy, S.; El-Kader, A.; Oda, S.; El-Namaky, A. Clinicohistopathological and immunological alterations in Egyptian donkeys infested by Rhinoestrus spp. during the winter season. Egypt. J. Vet. Sci. 2017, 48, 61–71. [Google Scholar] [CrossRef] [Green Version]
- Abdel-Daim, M.M.; Shaheen, H.M.; Abushouk, A.I.; Toraih, E.A.; Fawzy, M.S.; Alansari, W.S.; Aleya, L.; Bungau, S. Thymoquinone and diallyl sulfide protect against fipronil-induced oxidative injury in rats. Environ. Sci. Pollut. Res. 2018, 25, 23909–23916. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Days Post Infection | Experimental Groups | ||||
|---|---|---|---|---|---|
| Control | SMC50 | C + ve | C + SMC25 | C + SMC50 | |
| Day 7 | 0.00 ± 0.00 d | 0.00 ± 0.00 d | 41.20 ± 3.11 a | 32.60 ± 3.05 b | 23.60 ± 2.51 c |
| Day 14 | 0.00 ± 0.00 d | 0.00 ± 0.00 d | 45.40 ± 4.34 a | 18.80 ± 2.59 b | 12.00 ± 3.54 c |
| Serum Biochemical Parameters | Experimental Groups | ||||
|---|---|---|---|---|---|
| Control | SMC50 | C + ve | C + SMC25 | C + SMC50 | |
| ALT (U/L) | 42.72 ± 2.08 d | 39.28 ± 3.52 d | 109.22 ± 13.56 a | 71.18 ± 7.35 b | 53.35 ± 6.77 c |
| AST (U/L) | 58.48 ± 8.70 c | 54.34 ± 5.46 c | 151.68 ± 14.69 a | 83.96 ± 8.97 b | 58.52 ± 9.53 c |
| ALP (U/L) | 138.60 ± 13.43 cd | 130.60 ± 21.14 d | 240.20 ± 10.89 a | 179.00 ± 8.60 b | 80.154 ± 9.36 c |
| Albumin (g/dL) | 4.42 ± 0.32 a | 4.64 ± 0.23 a | 2.82 ± 0.19 d | 3.38 ± 0.33 c | 3.98 ± 0.19 b |
| Globulin (g/dL) | 4.88 ± 0.29 bc | 5.04 ± 0.21 b | 2.94 ± 0.18 d | 4.44 ± 0.58 c | 5.70 ± 0.36 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Elmahallawy, E.K.; Elshopakey, G.E.; Saleh, A.A.; Agil, A.; El-Morsey, A.; EL-shewehy, D.M.M.; Sad, A.S.; Yanai, T.; Abdo, W. S-Methylcysteine (SMC) Ameliorates Intestinal, Hepatic, and Splenic Damage Induced by Cryptosporidium parvum Infection Via Targeting Inflammatory Modulators and Oxidative Stress in Swiss Albino Mice. Biomedicines 2020, 8, 423. https://doi.org/10.3390/biomedicines8100423
Elmahallawy EK, Elshopakey GE, Saleh AA, Agil A, El-Morsey A, EL-shewehy DMM, Sad AS, Yanai T, Abdo W. S-Methylcysteine (SMC) Ameliorates Intestinal, Hepatic, and Splenic Damage Induced by Cryptosporidium parvum Infection Via Targeting Inflammatory Modulators and Oxidative Stress in Swiss Albino Mice. Biomedicines. 2020; 8(10):423. https://doi.org/10.3390/biomedicines8100423
Chicago/Turabian StyleElmahallawy, Ehab Kotb, Gehad E. Elshopakey, Amira A. Saleh, Ahmad Agil, Ahmed El-Morsey, Dina M. M. EL-shewehy, Ahmed S. Sad, Tokuma Yanai, and Walied Abdo. 2020. "S-Methylcysteine (SMC) Ameliorates Intestinal, Hepatic, and Splenic Damage Induced by Cryptosporidium parvum Infection Via Targeting Inflammatory Modulators and Oxidative Stress in Swiss Albino Mice" Biomedicines 8, no. 10: 423. https://doi.org/10.3390/biomedicines8100423