
mRNA Profiles of Porcine Parathyroid Glands Following Variable Phosphorus Supplies throughout Fetal and Postnatal Life

 , , , , , , and
, , , , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Diets, Animals and Sample Collection
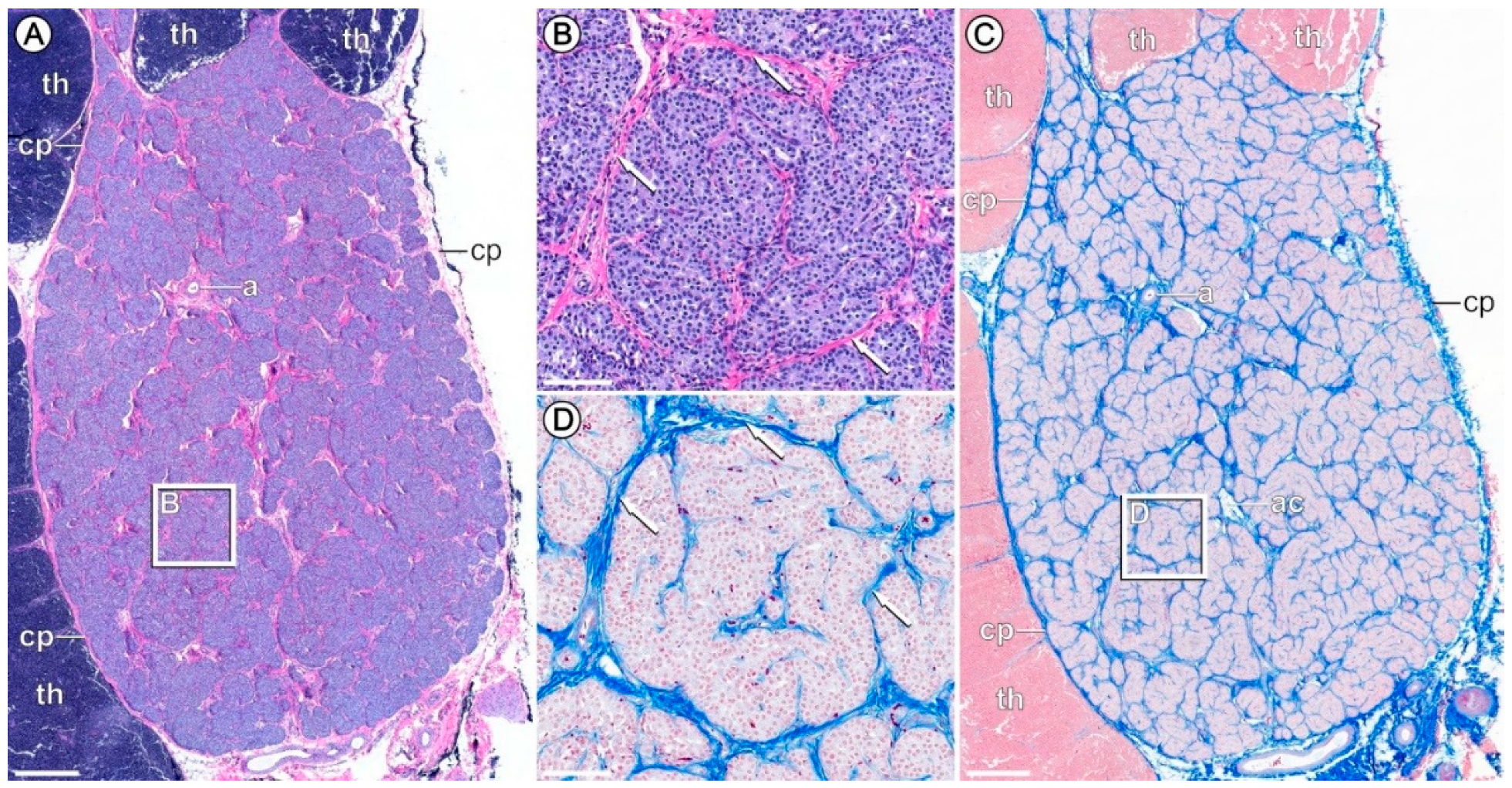
2.2. Histology
2.3. Growth and Performance
2.4. Serum Parameters of P Homeostasis
2.5. RNA Isolation and RNA Sequencing
2.6. Data Analyses
3. Results
3.1. Growth and Performance Data
3.2. Parameters of P Homeostasis in Serum
3.3. Verification of PTG Sampling
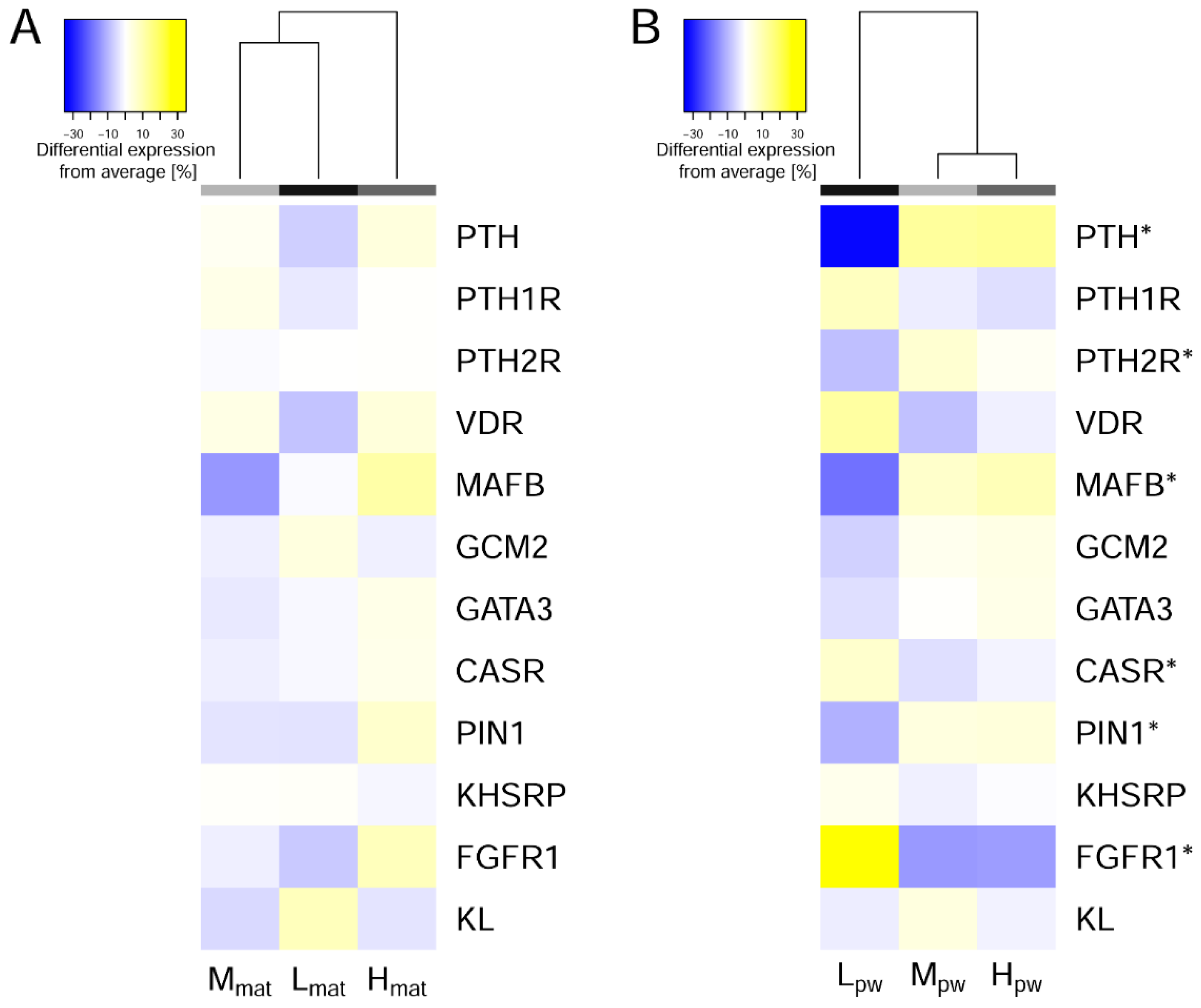
3.4. Transcriptome Profiling of PTGs
4. Discussion
4.1. Gene Expression Patterns at Physiological Conditions of Porcine PTGs
4.2. Endogenous Responses to Balance PTH Expression, Stability and Secretion
4.3. Gene Expression Patterns at Physiological Conditions of Porcine PTGs
4.4. Early Life Conditioning and Implications on Long-Term Health
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Shapiro, R.; Heaney, R. Co-dependence of calcium and phosphorus for growth and bone development under conditions of varying deficiency. Bone 2003, 32, 532–540. [Google Scholar] [CrossRef]
- Gerlinger, C.; Oster, M.; Reyer, H.; Polley, C.; Vollmar, B.; Muráni, E.; Wimmers, K.; Wolf, P. Effects of excessive or restricted phosphorus and calcium intake during early life on markers of bone architecture and composition in pigs. J. Anim. Physiol. Anim. Nutr. 2020. [Google Scholar] [CrossRef] [Green Version]
- Pokharel, B.B.; Regassa, A.; Nyachoti, C.M.; Kim, W.K. Effect of low levels of dietary available phosphorus on phosphorus utilization, bone mineralization, phosphorus transporter mRNA expression and performance in growing pigs. J. Environ. Sci. Heal. Part B 2017, 52, 395–401. [Google Scholar] [CrossRef]
- Sørensen, K.U.; Kruger, M.C.; Hansen-Møller, J.; Poulsen, H.D. Bone biochemical markers for assessment of bone responses to differentiated phosphorus supply in growing-finishing pigs. J. Anim. Sci. 2018, 96, 4693–4703. [Google Scholar] [CrossRef]
- Ji, Y.; Wu, Z.; Dai, Z.; Wang, X.; Li, J.; Wang, B.; Wu, G. Fetal and neonatal programming of postnatal growth and feed efficiency in swine. J. Anim. Sci. Biotechnol. 2017, 8, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Hales, C.N.; Barker, D.J.P. Type 2 (non-insulin-dependent) diabetes mellitus: The thrifty phenotype hypothesis. Diabetologia 1992, 35, 595–601. [Google Scholar] [CrossRef] [PubMed]
- Snoeck, A.; Remacle, C.; Reusens, B.; Hoet, J.J. Effect of a Low Protein Diet during Pregnancy on the Fetal Rat Endocrine Pancreas. Biol. Neonateol. 1990, 57, 107–118. [Google Scholar] [CrossRef]
- McMillen, I.C.; Robinson, J.S. Developmental Origins of the Metabolic Syndrome: Prediction, Plasticity, and Programming. Physiol. Rev. 2005, 85, 571–633. [Google Scholar] [CrossRef]
- Zhang, W.; A Kroscher, K.; Murray, R.L.; Gagliardi, R.; Guiltinan, C.; Rhoads, R.; Stahl, C.H. Dietary Calcium and Phosphorus Amounts Affect Development and Tissue-Specific Stem Cell Characteristics in Neonatal Pigs. J. Nutr. 2020, 150, 1086–1092. [Google Scholar] [CrossRef] [PubMed]
- Uy, H.L. Effects of parathyroid hormone (PTH)-related protein and PTH on osteoclasts and osteoclast precursors in vivo. Endocrinology 1995, 136, 3207–3212. [Google Scholar] [CrossRef]
- Lee, M.; Partridge, N.C. Parathyroid hormone signaling in bone and kidney. Curr. Opin. Nephrol. Hypertens. 2009, 18, 298–302. [Google Scholar] [CrossRef]
- Brenza, H.L.; Kimmel-Jehan, C.; Jehan, F.; Shinki, T.; Wakino, S.; Anazawa, H.; Suda, T.; DeLuca, H.F. Parathyroid hormone activation of the 25-hydroxyvitamin D3-1 -hydroxylase gene promoter. Proc. Natl. Acad. Sci. USA 1998, 95, 1387–1391. [Google Scholar] [CrossRef] [Green Version]
- Ritter, C.S.; Haughey, B.H.; Miller, B.; Brown, A.J. Differential Gene Expression by Oxyphil and Chief Cells of Human Parathyroid Glands. J. Clin. Endocrinol. Metab. 2012, 97, E1499–E1505. [Google Scholar] [CrossRef]
- Shi, Y.; Hogue, J.; Dixit, D.; Koh, J.; Olson, J.A. Functional and genetic studies of isolated cells from parathyroid tumors reveal the complex pathogenesis of parathyroid neoplasia. Proc. Natl. Acad. Sci. USA 2014, 111, 3092–3097. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DeMay, M.B.; Kiernan, M.S.; DeLuca, H.F.; Kronenberg, H.M. Sequences in the human parathyroid hormone gene that bind the 1,25-dihydroxyvitamin D3 receptor and mediate transcriptional repression in response to 1,25-dihydroxyvitamin D3. Proc. Natl. Acad. Sci. USA 1992, 89, 8097–8101. [Google Scholar] [CrossRef] [Green Version]
- Ben-Dov, I.Z.; Galitzer, H.; Lavi-Moshayoff, V.; Goetz, R.; Kuro-O, M.; Mohammadi, M.; Sirkis, R.; Naveh-Many, T.; Silver, J. The parathyroid is a target organ for FGF23 in rats. J. Clin. Investig. 2007, 117, 4003–4008. [Google Scholar] [CrossRef] [Green Version]
- Brown, E.M.; Gamba, G.; Riccardi, D.; Lombardi, M.; Butters, R.; Kifor, O.; Sun, A.; Hediger, M.A.; Lytton, J.; Hebert, S.C. Cloning and characterization of an extracellular Ca2+-sensing receptor from bovine parathyroid. Nat. Cell Biol. 1993, 366, 575–580. [Google Scholar] [CrossRef]
- Geng, Y.; Mosyak, L.; Kurinov, I.; Zuo, H.; Sturchler, E.; Cheng, T.C.; Subramanyam, P.; Brown, A.P.; Brennan, S.C.; Mun, H.-C.; et al. Structural mechanism of ligand activation in human calcium-sensing receptor. eLife 2016, 5, e13662. [Google Scholar] [CrossRef]
- Canalejo, A.; Rodríguez, M.; Almadén, Y. PTH Regulation by Phosphate and miRNAs. In Parathyroid Glands in Chronic Kidney Disease; Covic, A., Goldsmith, D., Ureña Torres, P., Eds.; Springer: Cham, Switzerland, 2020; pp. 79–91. [Google Scholar] [CrossRef]
- Nechama, M.; Ben-Dov, I.Z.; Briata, P.; Gherzi, R.; Naveh-Many, T. The mRNA decay promoting factor K-homology splicing regulator protein post-transcriptionally determines parathyroid hormone mRNA levels. FASEB J. 2008, 22, 3458–3468. [Google Scholar] [CrossRef]
- Shilo, V.; Levi, I.M.-Y.; Abel, R.; Mihailović, A.; Wasserman, G.; Naveh-Many, T.; Ben-Dov, I.Z. Let-7andMicroRNA-148Regulate Parathyroid Hormone Levels in Secondary Hyperparathyroidism. J. Am. Soc. Nephrol. 2017, 28, 2353–2363. [Google Scholar] [CrossRef] [Green Version]
- Naveh-Many, T.; Silver, J. Transcription factors that determine parathyroid development power PTH expression. Kidney Int. 2018, 93, 7–9. [Google Scholar] [CrossRef] [Green Version]
- Morito, N.; Yoh, K.; Usui, T.; Oishi, H.; Ojima, M.; Fujita, A.; Koshida, R.; Shawki, H.H.; Hamada, M.; Muratani, M.; et al. Transcription factor MafB may play an important role in secondary hyperparathyroidism. Kidney Int. 2018, 93, 54–68. [Google Scholar] [CrossRef] [Green Version]
- AfBN (Ausschuss für Bedarfsnormen der Gesellschaft für Ernährungsphysiologie). Empfehlungen zur Energie- und Nährstoffversorgung von Schweinen; DLG-Verlag: Frankfurt am Main, Germany, 2006. [Google Scholar]
- Wubuli, A.; Gerlinger, C.; Reyer, H.; Oster, M.; Muráni, E.; Trakooljul, N.; Ponsuksili, S.; Wolf, P.; Wimmers, K. Reduced phosphorus intake throughout gestation and lactation of sows is mitigated by transcriptional adaptations in kidney and intestine. BMC Genom. 2020, 21, 1–11. [Google Scholar] [CrossRef]
- Oster, M.; Keiler, J.; Schulze, M.; Reyer, H.; Wree, A.; Wimmers, K. Fast and reliable dissection of porcine parathyroid glands—A protocol for molecular and histological analyses. Ann. Anat. Anat. Anz. 2018, 219, 76–81. [Google Scholar] [CrossRef]
- Helvich, K. Official Methods of Analysis, 15th ed.; Association of Official Analytical Chemists: Washington, DC, USA, 1990. [Google Scholar]
- Pertea, M.; Kim, D.; Pertea, G.M.; Leek, J.T.; Salzberg, S.L. Transcript-level expression analysis of RNA-seq experiments with HISAT, StringTie and Ballgown. Nat. Protoc. 2016, 11, 1650–1667. [Google Scholar] [CrossRef]
- Anders, S.; Pyl, P.T.; Huber, W. HTSeq—A Python framework to work with high-throughput sequencing data. Bioinformatics 2015, 31, 166–169. [Google Scholar] [CrossRef]
- Kauffmann, A.; Gentleman, R.; Huber, W. arrayQualityMetrics—A bioconductor package for quality assessment of microarray data. Bioinformatics 2009, 25, 415–416. [Google Scholar] [CrossRef] [Green Version]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- D’amico, M.A.; Ghinassi, B.; Izzicupo, P.; Manzoli, L.; Di Baldassarre, A. Biological function and clinical relevance of chromogranin A and derived peptides. Endocr. Connect. 2014, 3, R45–R54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oster, M.; Just, F.; Büsing, K.; Wolf, P.; Polley, C.; Vollmar, B.; Muráni, E.; Ponsuksili, S.; Wimmers, K. Toward improved phosphorus efficiency in monogastrics—Interplay of serum, minerals, bone, and immune system after divergent dietary phosphorus supply in swine. Am. J. Physiol. Integr. Comp. Physiol. 2016, 310, R917–R925. [Google Scholar] [CrossRef] [Green Version]
- Lopez-Hilker, S.; Dusso, A.S.; Rapp, N.S.; Martin, K.J.; Slatopolsky, E. Phosphorus restriction reverses hyperparathyroidism in uremia independent of changes in calcium and calcitriol. Am. J. Physiol. Physiol. 1990, 259, F432–F437. [Google Scholar] [CrossRef]
- Brown, E.M. Role of the calcium-sensing receptor in extracellular calcium homeostasis. Best Pract. Res. Clin. Endocrinol. Metab. 2013, 27, 333–343. [Google Scholar] [CrossRef]
- Han, S.-I.; Tsunekage, Y.; Kataoka, K. Gata3 cooperates with Gcm2 and MafB to activate parathyroid hormone gene expression by interacting with SP1. Mol. Cell. Endocrinol. 2015, 411, 113–120. [Google Scholar] [CrossRef] [PubMed]
- Hernández, A.; Concepción, M.T.; Rodríguez, M.; Salido, E.; Torres, A. High phosphorus diet increases preproPTH mRNA independent of calcium and calcitriol in normal rats. Kidney Int. 1996, 50, 1872–1878. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silver, J.; Naveh-Many, T. Vitamin D and the parathyroids. In Vitamin D, 4th ed.; Feldman, D., Pike, J.W., Bouillon, R., Giovannucci, E., Goltzman, D., Hewison, M., Eds.; Academic Press: San Diego, CA, USA, 2018; pp. 461–475. [Google Scholar]
- Hendy, G.; D’Souza-Li, L.; Yang, B.; Canaff, L.; Cole, D. Mutations of the calcium-sensing receptor (CASR) in familial hypocalciuric hypercalcemia, neonatal severe hyperparathyroidism, and autosomal dominant hypocalcemia. Hum. Mutat. 2000, 16, 281–296. [Google Scholar] [CrossRef]
- Brown, A.J.; Ritter, C.S.; Finch, J.L.; Slatopolsky, E.A. Decreased calcium-sensing receptor expression in hyperplastic parathyroid glands of uremic rats: Role of dietary phosphate. Kidney Int. 1999, 55, 1284–1292. [Google Scholar] [CrossRef] [Green Version]
- Just, F.; Öster, M.; Büsing, K.; Borgelt, L.; Murani, E.; Ponsuksili, S.; Wolf, P.; Wimmers, K. Lowered dietary phosphorus affects intestinal and renal gene expression to maintain mineral homeostasis with immunomodulatory implications in weaned piglets. BMC Genom. 2018, 19, 207. [Google Scholar] [CrossRef] [Green Version]
- Nechama, M.; Uchida, T.; Yosef-Levi, I.M.; Silver, J.; Naveh-Many, T. The peptidyl-prolyl isomerase Pin1 determines parathyroid hormone mRNA levels and stability in rat models of secondary hyperparathyroidism. J. Clin. Investig. 2009, 119, 3102–3114. [Google Scholar] [CrossRef]
- Kifor, O.; Diaz, R.; Butters, R.; Brown, E.M. The Ca2+-Sensing Receptor (CaR) Activates Phospholipases C, A2, and D in Bovine Parathyroid and CaR-Transfected, Human Embryonic Kidney (HEK293) Cells. J. Bone Miner. Res. 1997, 12, 715–725. [Google Scholar] [CrossRef]
- Ritter, C.S.; Slatopolsky, E.; Santoro, S.; Brown, A.J. Parathyroid Cells Cultured in Collagen Matrix Retain Calcium Responsiveness: Importance of Three-Dimensional Tissue Architecture. J. Bone Miner. Res. 2004, 19, 491–498. [Google Scholar] [CrossRef]
- Sun, F.; Maercklein, P.; Fitzpatrick, L.A. Paracrine interactions among parathyroid cells: Effect of cell density on cell secretion. J. Bone Miner. Res. 2009, 9, 971–976. [Google Scholar] [CrossRef]
- Verdelli, C.; Avagliano, L.; Creo, P.; Guarnieri, V.; Scillitani, A.; Vicentini, L.; Steffano, G.B.; Beretta, E.; Soldati, L.; Costa, E.; et al. Tumour-associated fibroblasts contribute to neoangiogenesis in human parathyroid neoplasia. Endocr. Relat. Cancer 2015, 22, 87–98. [Google Scholar] [CrossRef] [Green Version]
- Dawodu, A.; Salameh, K.M.; Al-Janahi, N.S.; Bener, A.; Elkum, N. The Effect of High-Dose Postpartum Maternal Vitamin D Supplementation Alone Compared with Maternal Plus Infant Vitamin D Supplementation in Breastfeeding Infants in a High-Risk Population. A Randomized Controlled Trial. Nutrients 2019, 11, 1632. [Google Scholar] [CrossRef] [Green Version]
- Reyer, H.; Oster, M.; Wittenburg, D.; Murani, E.; Ponsuksili, S.; Wimmers, K. Genetic Contribution to Variation in Blood Calcium, Phosphorus, and Alkaline Phosphatase Activity in Pigs. Front. Genet. 2019, 10, 590. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Maternal Diet | Offspring Diet | BW [kg] | DFI [kg/d] | ADG [kg/d] | FCR [g/g] | Ash (MLD) [%] |
|---|---|---|---|---|---|---|
| L | L | 60.72 | 1.67 | 0.74 | 2.27 | 1.20 |
| L | M | 69.30 | 1.88 | 0.85 | 2.18 | 1.19 |
| L | H | 65.54 | 1.73 | 0.81 | 2.14 | 1.20 |
| M | L | 47.70 | 1.30 | 0.60 | 2.16 | 1.22 |
| M | M | 58.09 | 1.56 | 0.74 | 2.08 | 1.22 |
| M | H | 62.19 | 1.67 | 0.81 | 2.07 | 1.20 |
| H | L | 55.30 | 1.50 | 0.68 | 2.18 | 1.20 |
| H | M | 58.17 | 1.54 | 0.73 | 2.11 | 1.22 |
| H | H | 62.83 | 1.62 | 0.80 | 2.02 | 1.20 |
| Pooled SEM | 9.00 | 0.28 | 0.11 | 0.22 | 0.03 | |
| p-Value | Maternal diet | 0.327 | 0.453 | 0.117 | 0.798 | 0.657 |
| post-weaning diet | <0.001 | 0.015 | <0.001 | 0.015 | 0.640 | |
| Mat diet × pw diet | 0.207 | 0.188 | 0.193 | 0.941 | 0.302 |
| Maternal Diet | Offspring Diet | IP [mg/dL] | Ca [mg/dL] | Mg [mg/dL] | ALP [U/L] |
|---|---|---|---|---|---|
| L | L | 8.94 | 10.97 | 2.21 | 164.8 |
| L | M | 10.75 | 10.21 | 2.20 | 135.1 |
| L | H | 10.66 | 10.40 | 2.12 | 158.4 |
| M | L | 8.05 | 11.44 | 2.22 | 237.5 |
| M | M | 10.66 | 10.54 | 2.14 | 142.8 |
| M | H | 10.60 | 10.42 | 2.11 | 153.3 |
| H | L | 8.66 | 11.04 | 2.22 | 169.0 |
| H | M | 11.28 | 10.64 | 2.24 | 144.9 |
| H | H | 10.75 | 10.52 | 2.20 | 143.6 |
| Pooled SEM | 0.90 | 0.57 | 0.20 | 36.5 | |
| p-Value | Maternal diet | 0.305 | 0.517 | 0.902 | 0.053 |
| Post-weaning diet | <0.001 | <0.001 | 0.327 | <0.001 | |
| Mat diet × pw diet | 0.346 | 0.466 | 0.873 | 0.011 |
| Canonical Pathway | Adjusted p-Value | z-Score † (Lpw vs. Hpw) | z-Score † (Lpw vs. Mpw) | z-Score † (Mpw vs. Hpw) |
|---|---|---|---|---|
| Axonal guidance signaling | 0.0003 | - | - | - |
| Protein kinase A signaling | 0.0015 | 1.07 | 1.07 | −1.07 |
| RhoGDI signaling | 0.0016 | 1.15 | 1.15 | −0.69 |
| GP6 signaling pathway | 0.0016 | 2.99 | 2.99 | 0.43 |
| Cardiac hypertrophy signaling (enhanced) | 0.0016 | 3.58 | 3.58 | 0.66 |
| NRF2-mediated oxidative stress response | 0.0021 | −1.04 | −1.04 | −1.04 |
| Phospholipase C signaling | 0.0021 | 0.78 | 0.78 | 0.39 |
| Cardiac β-adrenergic signaling | 0.0021 | 2.67 | 2.67 | −0.73 |
| cAMP-mediated signaling | 0.0021 | 2.12 | 2.12 | 0.00 |
| LPS/IL-1 mediated inhibition of RXR function | 0.0032 | 0.71 | 0.71 | 0.71 |
| Thrombin signaling | 0.0042 | 1.21 | 1.21 | 0.24 |
| Adipogenesis pathway | 0.0046 | - | - | - |
| Glycogen degradation III | 0.0068 | −0.82 | −0.82 | 0.82 |
| α-Adrenergic signaling | 0.0083 | 0.91 | 0.91 | −1.51 |
| G-Protein coupled receptor signaling | 0.0083 | - | - | - |
| Ephrin B signaling | 0.0093 | −1.00 | −1.00 | 0.33 |
| Gαi signaling | 0.0093 | 1.50 | 1.50 | 0.50 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oster, M.; Reyer, H.; Gerlinger, C.; Trakooljul, N.; Siengdee, P.; Keiler, J.; Ponsuksili, S.; Wolf, P.; Wimmers, K. mRNA Profiles of Porcine Parathyroid Glands Following Variable Phosphorus Supplies throughout Fetal and Postnatal Life. Biomedicines 2021, 9, 454. https://doi.org/10.3390/biomedicines9050454
Oster M, Reyer H, Gerlinger C, Trakooljul N, Siengdee P, Keiler J, Ponsuksili S, Wolf P, Wimmers K. mRNA Profiles of Porcine Parathyroid Glands Following Variable Phosphorus Supplies throughout Fetal and Postnatal Life. Biomedicines. 2021; 9(5):454. https://doi.org/10.3390/biomedicines9050454
Chicago/Turabian StyleOster, Michael, Henry Reyer, Christian Gerlinger, Nares Trakooljul, Puntita Siengdee, Jonas Keiler, Siriluck Ponsuksili, Petra Wolf, and Klaus Wimmers. 2021. "mRNA Profiles of Porcine Parathyroid Glands Following Variable Phosphorus Supplies throughout Fetal and Postnatal Life" Biomedicines 9, no. 5: 454. https://doi.org/10.3390/biomedicines9050454