
Neurobiological Responses towards Stimuli Depicting Aggressive Interactions in Delinquent Young Adults and Controls: No Relation to Reactive and Proactive Aggression

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Materials
2.2.1. Questionnaires
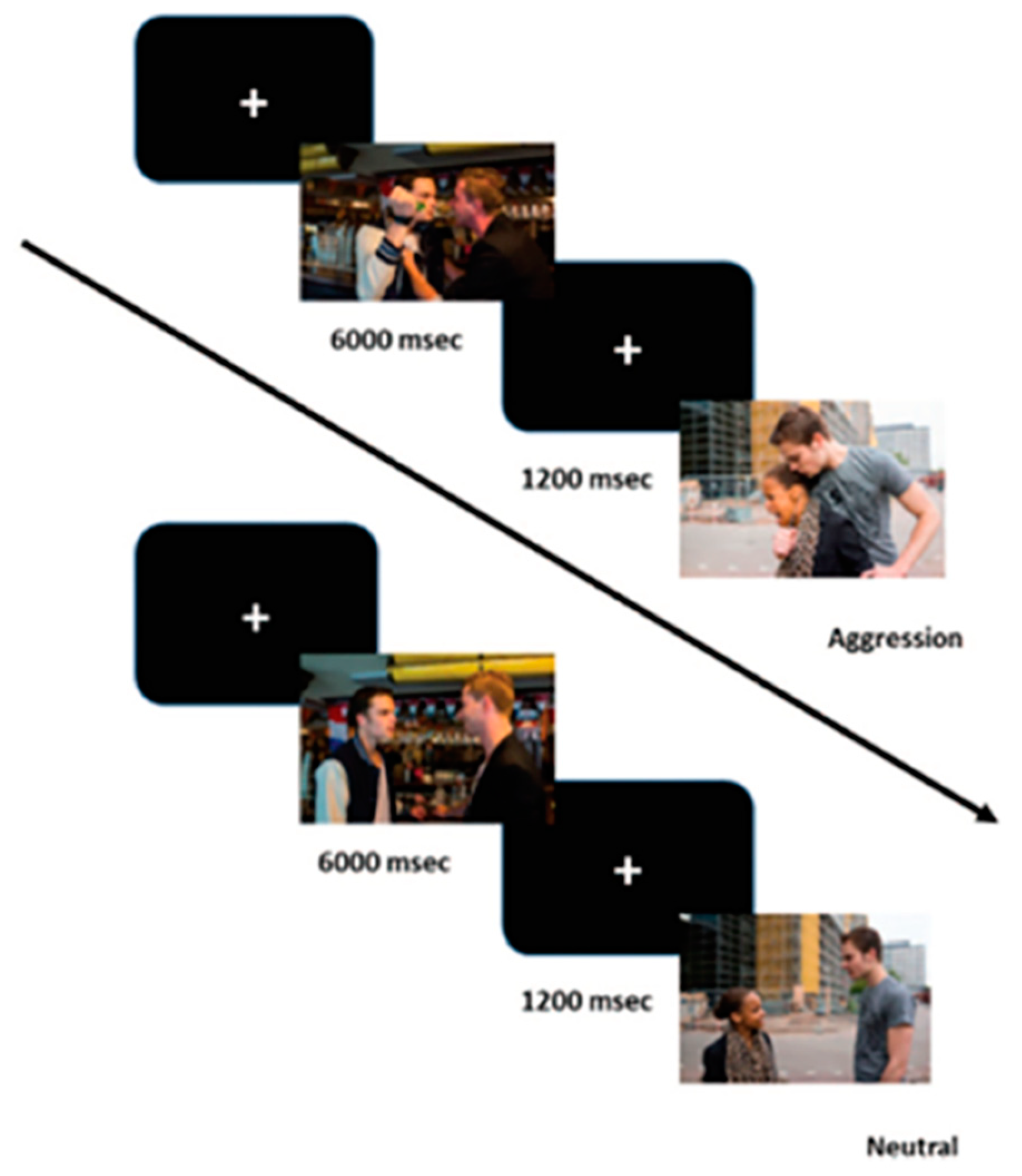
2.2.2. Passive Picture Viewing Task
2.2.3. Psychophysiological Measures
2.2.4. Electrophysiological Measures
2.2.5. Data-Analysis
3. Results
3.1. Descriptive Analysis
3.2. Task Validity
3.3. Group Differences
3.3.1. Psychophysiological Measures
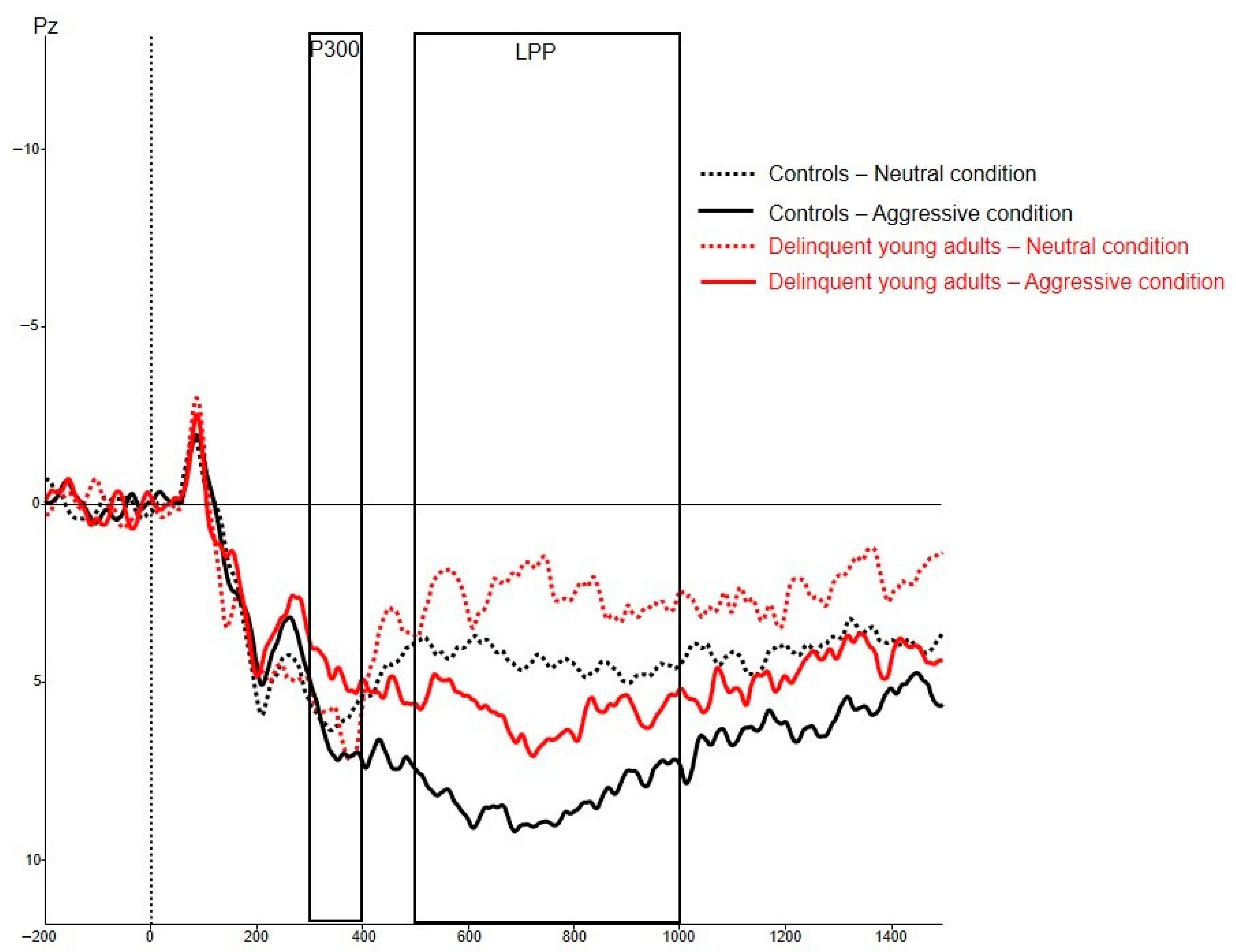
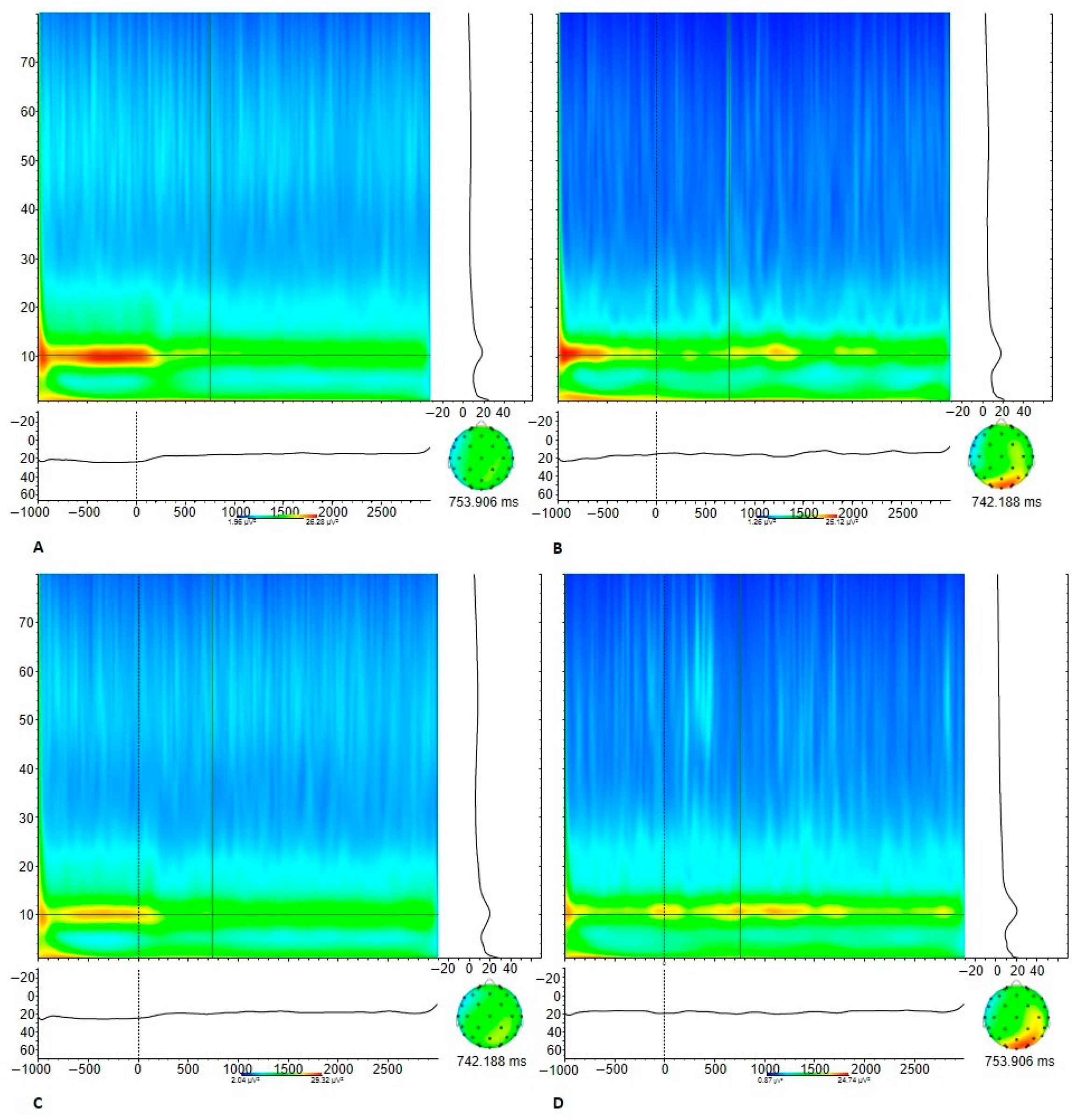
3.3.2. Electrophysiological Measures
3.4. Regression Analyses
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cohen, M.A. The Costs of Crime and Justice; Routledge: London, UK, 2020. [Google Scholar]
- De Looff, P.C.; Cornet, L.J.; De Kogel, C.H.; Fernández-Castilla, B.; Embregts, P.J.; Didden, R.; Nijman, H.L. Heart Rate and Skin Conductance Associations with Physical Aggression, Psychopathy, Antisocial Personality Disorder and Conduct Disorder: An Updated Meta-Analysis. Neurosci. Biobehav. Rev. 2021, 132, 553–582. [Google Scholar] [CrossRef] [PubMed]
- Henwood, K.S.; Chou, S.; Browne, K.D. A systematic review and meta-analysis on the effectiveness of CBT informed anger management. Aggress. Violent Behav. 2015, 25, 280–292. [Google Scholar] [CrossRef]
- Brännström, L.; Kaunitz, C.; Andershed, A.-K.; South, S.; Smedslund, G. Aggression replacement training (ART) for reducing antisocial behavior in adolescents and adults: A systematic review. Aggress. Violent Behav. 2016, 27, 30–41. [Google Scholar] [CrossRef] [Green Version]
- McIntosh, L.G.; Janes, S.; O’Rourke, S.; Thomson, L.D. Effectiveness of psychological and psychosocial interventions for forensic mental health inpatients: A meta-analysis. Aggress. Violent Behav. 2021, 58, 101551. [Google Scholar] [CrossRef]
- Wagels, L.; Habel, U.; Raine, A.; Clemens, B. Neuroimaging, hormonal and genetic biomarkers for pathological aggression—success or failure? Curr. Opin. Behav. Sci. 2022, 43, 101–110. [Google Scholar] [CrossRef]
- Popma, A.; Raine, A. Will future forensic assessment be neurobiologic? Child Adolesc. Psychiatr. Clin. 2006, 15, 429–444. [Google Scholar] [CrossRef] [Green Version]
- Smeijers, D.; Benbouriche, M.; Garofalo, C. The association between emotion, social information processing, and aggressive behavior: A systematic review. Eur. Psychol. 2020, 25, 81–91. [Google Scholar] [CrossRef]
- Garofalo, C.; Neumann, C.S.; Velotti, P. Psychopathy and aggression: The role of emotion dysregulation. J. Interpers. Violence 2020, 36, NP12640–NP12664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blankenstein, N.E.; de Rooij, M.; van Ginkel, J.; Wilderjans, T.F.; de Ruigh, E.L.; Oldenhof, H.C.; Zijlmans, J.; Jambroes, T.; Platje, E.; de Vries-Bouw, M. Neurobiological correlates of antisociality across adolescence and young adulthood: A multi-sample, multi-method study. Psychol. Med. 2021, 1–16. [Google Scholar] [CrossRef]
- Johanson, M.; Vaurio, O.; Tiihonen, J.; Lähteenvuo, M. A systematic literature review of neuroimaging of psychopathic traits. Front. Psychiatry 2020, 10, 1027. [Google Scholar] [CrossRef] [Green Version]
- Dodge, K.A.; Coie, J.D. Social-information-processing factors in reactive and proactive aggression in children’s peer groups. J. Personal. Soc. Psychol. 1987, 53, 1146. [Google Scholar] [CrossRef]
- Berkowitz, L. Aggression: Its Causes, Consequences, and Control; McGraw-Hill Book Company: New York, NY, USA, 1993. [Google Scholar]
- Bandura, A. Social learning theory of aggression. J. Commun. 1978, 28, 12–29. [Google Scholar] [CrossRef] [PubMed]
- Raine, A.; Jones, F. Attention, autonomic arousal, and personality in behaviorally disordered children. J. Abnorm. Child Psychol. 1987, 15, 583–599. [Google Scholar] [CrossRef] [PubMed]
- Eysenck, H.J. Personality and experimental psychology: The unification of psychology and the possibility of a paradigm. J. Personal. Soc. Psychol. 1997, 73, 1224. [Google Scholar] [CrossRef]
- Cima, M.; Raine, A. Distinct characteristics of psychopathy relate to different subtypes of aggression. Personal. Individ. Differ. 2009, 47, 835–840. [Google Scholar] [CrossRef]
- Raine, A.; Dodge, K.; Loeber, R.; Gatzke-Kopp, L.; Lynam, D.; Reynolds, C.; Stouthamer-Loeber, M.; Liu, J. The reactive–proactive aggression questionnaire: Differential correlates of reactive and proactive aggression in adolescent boys. Aggress. Behav. Off. J. Int. Soc. Res. Aggress. 2006, 32, 159–171. [Google Scholar] [CrossRef] [Green Version]
- Hubbard, J.A.; McAuliffe, M.D.; Morrow, M.T.; Romano, L.J. Reactive and proactive aggression in childhood and adolescence: Precursors, outcomes, processes, experiences, and measurement. J. Personal. 2010, 78, 95–118. [Google Scholar] [CrossRef]
- Lobbestael, J.; Cima, M.; Arntz, A. The relationship between adult reactive and proactive aggression, hostile interpretation bias, and antisocial personality disorder. J. Personal. Disord. 2013, 27, 53–66. [Google Scholar] [CrossRef]
- Latzman, R.D.; Vaidya, J.G. Common and distinct associations between aggression and alcohol problems with trait disinhibition. J. Psychopathol. Behav. Assess. 2013, 35, 186–196. [Google Scholar] [CrossRef]
- Fite, P.J.; Raine, A.; Stouthamer-Loeber, M.; Loeber, R.; Pardini, D.A. Reactive and proactive aggression in adolescent males: Examining differential outcomes 10 years later in early adulthood. Crim. Justice Behav. 2010, 37, 141–157. [Google Scholar] [CrossRef]
- Brugman, S.; Lobbestael, J.; Arntz, A.; Cima, M.; Schuhmann, T.; Dambacher, F.; Sack, A.T. Identifying cognitive predictors of reactive and proactive aggression. Aggress. Behav. 2015, 41, 51–64. [Google Scholar] [CrossRef] [PubMed]
- Portnoy, J.; Farrington, D.P. Resting heart rate and antisocial behavior: An updated systematic review and meta-analysis. Aggress. Violent Behav. 2015, 22, 33–45. [Google Scholar] [CrossRef]
- Ortiz, J.; Raine, A. Heart rate level and antisocial behavior in children and adolescents: A meta-analysis. J. Am. Acad. Child Adolesc. Psychiatry 2004, 43, 154–162. [Google Scholar] [CrossRef]
- Lorber, M.F. Psychophysiology of aggression, psychopathy, and conduct problems: A meta-analysis. Psychol. Bull. 2004, 130, 531. [Google Scholar] [CrossRef] [PubMed]
- Crozier, J.C.; Dodge, K.A.; Fontaine, R.G.; Lansford, J.E.; Bates, J.E.; Pettit, G.S.; Levenson, R.W. Social information processing and cardiac predictors of adolescent antisocial behavior. J. Abnorm. Psychol. 2008, 117, 253. [Google Scholar] [CrossRef] [Green Version]
- Wagner, C.R.; Abaied, J.L. Relational victimization and proactive versus reactive relational aggression: The moderating effects of respiratory sinus arrhythmia and skin conductance. Aggress. Behav. 2015, 41, 566–579. [Google Scholar] [CrossRef] [PubMed]
- Puhalla, A.A.; McCloskey, M.S. The relationship between physiological reactivity to provocation and emotion dysregulation with proactive and reactive aggression. Biol. Psychol. 2020, 155, 107931. [Google Scholar] [CrossRef]
- Zijlmans, J.; Marhe, R.; van Duin, L.; Luijks, M.-J.; Bevaart, F.; Popma, A. No association between autonomic functioning and psychopathy and aggression in multi-problem young adults. Front. Psychol. 2021, 12, 686. [Google Scholar] [CrossRef]
- Murray-Close, D.; Holterman, L.A.; Breslend, N.L.; Sullivan, A. Psychophysiology of proactive and reactive relational aggression. Biol. Psychol. 2017, 130, 77–85. [Google Scholar] [CrossRef]
- Hubbard, J.A.; Smithmyer, C.M.; Ramsden, S.R.; Parker, E.H.; Flanagan, K.D.; Dearing, K.F.; Relyea, N.; Simons, R.F. Observational, physiological, and self–report measures of children’s anger: Relations to reactive versus proactive aggression. Child Dev. 2002, 73, 1101–1118. [Google Scholar] [CrossRef]
- Wagner, C.R.; Abaied, J.L. Skin conductance level reactivity moderates the association between parental psychological control and relational aggression in emerging adulthood. J. Youth Adolesc. 2016, 45, 687–700. [Google Scholar] [CrossRef]
- Armstrong, T.; Wells, J.; Boisvert, D.L.; Lewis, R.; Cooke, E.M.; Woeckener, M.; Kavish, N. Skin conductance, heart rate and aggressive behavior type. Biol. Psychol. 2019, 141, 44–51. [Google Scholar] [CrossRef] [PubMed]
- Thomson, N.D.; Kevorkian, S.; Blair, J.; Farrell, A.; West, S.J.; Bjork, J.M. Psychophysiological underpinnings of proactive and reactive aggression in young men and women. Physiol. Behav. 2021, 242, 113601. [Google Scholar] [CrossRef] [PubMed]
- Moore, C.C.; Hubbard, J.; Morrow, M.T.; Barhight, L.R.; Lines, M.M.; Sallee, M.; Hyde, C.T. The simultaneous assessment of and relations between children’s sympathetic and parasympathetic psychophysiology and their reactive and proactive aggression. Aggress. Behav. 2018, 44, 614–623. [Google Scholar] [CrossRef]
- Centifanti, L.C.M.; Kimonis, E.R.; Frick, P.J.; Aucoin, K.J. Emotional reactivity and the association between psychopathy-linked narcissism and aggression in detained adolescent boys. Dev. Psychopathol. 2013, 25, 473–485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gatzke-Kopp, L.M.; Greenberg, M.; Bierman, K. Children’s parasympathetic reactivity to specific emotions moderates response to intervention for early-onset aggression. J. Clin. Child Adolesc. Psychol. 2015, 44, 291–304. [Google Scholar] [CrossRef]
- Beauchaine, T.P.; Thayer, J.F. Heart rate variability as a transdiagnostic biomarker of psychopathology. Int. J. Psychophysiol. 2015, 98, 338–350. [Google Scholar] [CrossRef]
- Zhang, W.; Gao, Y. Interactive effects of social adversity and respiratory sinus arrhythmia activity on reactive and proactive aggression. Psychophysiology 2015, 52, 1343–1350. [Google Scholar] [CrossRef]
- Patrick, C.J. Physiological correlates of psychopathy, antisocial personality disorder, habitual aggression, and violence. Electrophysiol. Psychophysiol. Psychiatry Psychopharmacol. 2014, 21, 197–227. [Google Scholar]
- Fanti, K.A.; Eisenbarth, H.; Goble, P.; Demetriou, C.; Kyranides, M.N.; Goodwin, D.; Zhang, J.; Bobak, B.; Cortese, S. Psychophysiological activity and reactivity in children and adolescents with conduct problems: A systematic review and meta-analysis. Neurosci. Biobehav. Rev. 2019, 100, 98–107. [Google Scholar] [CrossRef] [PubMed]
- Branje, S.; Koot, H.M. Psychophysiology of aggression. In Handbook of Child and Adolescent Aggression; Guilford Press: New York, NY, USA, 2018; pp. 84–106. [Google Scholar]
- El-Sheikh, M.; Kouros, C.D.; Erath, S.; Cummings, E.M.; Keller, P.; Staton, L. Marital conflict and children’s externalizing behavior: Pathways involving interactions between parasympathetic and sympathetic nervous system activity. Monogr. Soc. Res. Child Dev. 2009, 74, vii. [Google Scholar]
- Raine, A. From genes to brain to antisocial behavior. Curr. Dir. Psychol. Sci. 2008, 17, 323–328. [Google Scholar] [CrossRef]
- Lang, P.J.; Bradley, M.M. Emotion and the motivational brain. Biol. Psychol. 2010, 84, 437–450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schupp, H.T.; Flaisch, T.; Stockburger, J.; Junghöfer, M. Emotion and attention: Event-related brain potential studies. Prog. Brain Res. 2006, 156, 31–51. [Google Scholar]
- Schubring, D.; Schupp, H.T. Affective picture processing: Alpha-and lower beta-band desynchronization reflects emotional arousal. Psychophysiology 2019, 56, e13386. [Google Scholar] [CrossRef]
- Gao, Y.; Raine, A.; Venables, P.H.; Mednick, S.A. The association between P3 amplitude at age 11 and criminal offending at age 23. J. Clin. Child Adolesc. Psychol. 2013, 42, 120–130. [Google Scholar] [CrossRef] [Green Version]
- Bernat, E.M.; Hall, J.R.; Steffen, B.V.; Patrick, C.J. Violent offending predicts P300 amplitude. Int. J. Psychophysiol. 2007, 66, 161–167. [Google Scholar] [CrossRef] [Green Version]
- Gerstle, J.E.; Mathias, C.W.; Stanford, M.S. Auditory P300 and self-reported impulsive aggression. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 1998, 22, 575–583. [Google Scholar] [CrossRef]
- Barratt, E.S.; Stanford, M.S.; Kent, T.A.; Alan, F. Neuropsychological and cognitive psychophysiological substrates of impulsive aggression. Biol. Psychiatry 1997, 41, 1045–1061. [Google Scholar] [CrossRef]
- Stanford, M.S.; Houston, R.J.; Villemarette-Pittman, N.R.; Greve, K.W. Premeditated aggression: Clinical assessment and cognitive psychophysiology. Personal. Individ. Differ. 2003, 34, 773–781. [Google Scholar] [CrossRef]
- Helfritz-Sinville, L.E.; Stanford, M.S. Looking for trouble? Processing of physical and social threat words in impulsive and premeditated aggression. Psychol. Rec. 2015, 65, 301–314. [Google Scholar] [CrossRef]
- Kunaharan, S.; Halpin, S.; Sitharthan, T.; Walla, P. Do EEG and startle reflex modulation vary with self-reported aggression in response to violent images? Brain Sci. 2019, 9, 298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gagnon, J.; Aubin, M.; Emond, F.C.; Derguy, S.; Brochu, A.F.; Bessette, M.; Jolicoeur, P. An ERP study on hostile attribution bias in aggressive and nonaggressive individuals. Aggress. Behav. 2017, 43, 217–229. [Google Scholar] [CrossRef]
- Cavanagh, J.F.; Frank, M.J. Frontal theta as a mechanism for cognitive control. Trends Cogn. Sci. 2014, 18, 414–421. [Google Scholar] [CrossRef] [Green Version]
- Peylo, C.; Hilla, Y.; Sauseng, P. Cause or consequence? Alpha oscillations in visuospatial attention. Trends Neurosci. 2021, 44, 705–713. [Google Scholar] [CrossRef] [PubMed]
- Otten, M.; Jonas, K.J. Humiliation as an intense emotional experience: Evidence from the electro-encephalogram. Soc. Neurosci. 2014, 9, 23–35. [Google Scholar] [CrossRef] [PubMed]
- Balconi, M.; Mazza, G. Brain oscillations and BIS/BAS (behavioral inhibition/activation system) effects on processing masked emotional cues.: ERS/ERD and coherence measures of alpha band. Int. J. Psychophysiol. 2009, 74, 158–165. [Google Scholar] [CrossRef]
- Furl, N.; Lohse, M.; Pizzorni-Ferrarese, F. Low-frequency oscillations employ a general coding of the spatio-temporal similarity of dynamic faces. Neuroimage 2017, 157, 486–499. [Google Scholar] [CrossRef]
- Knyazev, G.G.; Bocharov, A.V.; Levin, E.A.; Savostyanov, A.N.; Slobodskoj-Plusnin, J.Y. Anxiety and oscillatory responses to emotional facial expressions. Brain Res. 2008, 1227, 174–188. [Google Scholar] [CrossRef]
- Popov, T.; Miller, G.A.; Rockstroh, B.; Weisz, N. Modulation of α power and functional connectivity during facial affect recognition. J. Neurosci. 2013, 33, 6018–6026. [Google Scholar] [CrossRef] [Green Version]
- Popov, T.; Steffen, A.; Weisz, N.; Miller, G.A.; Rockstroh, B. Cross-frequency dynamics of neuromagnetic oscillatory activity: Two mechanisms of emotion regulation. Psychophysiology 2012, 49, 1545–1557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, Z.; Zhong, S.; Xu, P.; Gong, G.; He, Y. PANDA: A pipeline toolbox for analyzing brain diffusion images. Front. Hum. Neurosci. 2013, 7, 42. [Google Scholar] [CrossRef] [Green Version]
- De Cesarei, A.; Codispoti, M. Affective modulation of the LPP and α-ERD during picture viewing. Psychophysiology 2011, 48, 1397–1404. [Google Scholar] [CrossRef]
- Mennella, R.; Patron, E.; Palomba, D. Frontal alpha asymmetry neurofeedback for the reduction of negative affect and anxiety. Behav. Res. Ther. 2017, 92, 32–40. [Google Scholar] [CrossRef] [PubMed]
- Schubring, D.; Schupp, H.T. Emotion and brain oscillations: High arousal is associated with decreases in alpha-and lower beta-band power. Cereb. Cortex 2021, 31, 1597–1608. [Google Scholar] [CrossRef]
- Calzada-Reyes, A.; Alvarez-Amador, A.; Galán-García, L.; Valdés-Sosa, M. EEG abnormalities in psychopath and non-psychopath violent offenders. J. Forensic Leg. Med. 2013, 20, 19–26. [Google Scholar] [CrossRef]
- Wang, Y.; Lin, Y.; Fu, C.; Huang, Z.; Xiao, S.; Yu, R. Effortless retaliation: The neural dynamics of interpersonal intentions in the Chicken Game using brain–computer interface. Soc. Cogn. Affect. Neurosci. 2021, 16, 1138–1149. [Google Scholar] [CrossRef]
- Zebarjadi, N.; Adler, E.; Kluge, A.; Jääskeläinen, I.P.; Sams, M.; Levy, J. Rhythmic neural patterns during empathy to vicarious pain: Beyond the affective-cognitive empathy dichotomy. Front. Hum. Neurosci. 2021, 15, 380. [Google Scholar] [CrossRef]
- Luijks, M.-J.A.; Bevaart, F.; Zijlmans, J.; Van Duin, L.; Marhe, R.; Doreleijers, T.A.; Tiemeier, H.; Asscher, J.J.; Popma, A. A multimodal day treatment program for multi-problem young adults: Study protocol for a randomized controlled trial. Trials 2017, 18, 225. [Google Scholar] [CrossRef] [PubMed]
- Zijlmans, J.; van Duin, L.; Jorink, M.; Marhe, R.; Luijks, M.-J.A.; Crone, M.; Popma, A.; Bevaart, F. Disentangling multiproblem behavior in male young adults: A cluster analysis. Dev. Psychopathol. 2021, 33, 149–159. [Google Scholar] [CrossRef]
- Cima, M.; Raine, A.; Meesters, C.; Popma, A. Validation of the Dutch Reactive Proactive Questionnaire (RPQ): Differential correlates of reactive and proactive aggression from childhood to adulthood. Aggress. Behav. 2013, 39, 99–113. [Google Scholar] [CrossRef]
- Van Dongen, J.D.; Brazil, I.A.; Van der Veen, F.M.; Franken, I.H. Electrophysiological correlates of empathic processing and its relation to psychopathic meanness. Neuropsychology 2018, 32, 996. [Google Scholar] [CrossRef] [PubMed]
- Klaver, C.; De Geus, E.; De Vries, J. Ambulatory monitoring system. Comput. Psychol. 1994, 5, 254–268. [Google Scholar]
- Gratton, G.; Coles, M.G.; Donchin, E. A new method for off-line removal of ocular artifact. Electroencephalogr. Clin. Neurophysiol. 1983, 55, 468–484. [Google Scholar] [CrossRef]
- Cuthbert, B.N.; Schupp, H.T.; Bradley, M.M.; Birbaumer, N.; Lang, P.J. Brain potentials in affective picture processing: Covariation with autonomic arousal and affective report. Biol. Psychol. 2000, 52, 95–111. [Google Scholar] [CrossRef] [Green Version]
- Hajcak, G.; Dunning, J.P.; Foti, D. Motivated and controlled attention to emotion: Time-course of the late positive potential. Clin. Neurophysiol. 2009, 120, 505–510. [Google Scholar] [CrossRef]
- Shamay-Tsoory, S.G.; Tomer, R.; Berger, B.D.; Aharon-Peretz, J. Characterization of empathy deficits following prefrontal brain damage: The role of the right ventromedial prefrontal cortex. J. Cogn. Neurosci. 2003, 15, 324–337. [Google Scholar] [CrossRef]
- Singer, T.; Kiebel, S.J.; Winston, J.S.; Dolan, R.J.; Frith, C.D. Brain responses to the acquired moral status of faces. Neuron 2004, 41, 653–662. [Google Scholar] [CrossRef] [Green Version]
- Herrmann, C.S.; Grigutsch, M.; Busch, N.A. 11 EEG oscillations and wavelet analysis. In Event-Related Potentials: A Methods Handbook; The MIT Press: Cambridge, MA, USA, 2005; p. 229. [Google Scholar]
- White, I.R.; Royston, P.; Wood, A.M. Multiple imputation using chained equations: Issues and guidance for practice. Stat. Med. 2011, 30, 377–399. [Google Scholar] [CrossRef]
- Van Buuren, S.; Groothuis-Oudshoorn, K. mice: Multivariate imputation by chained equations in R. J. Stat. Softw. 2011, 45, 1–67. [Google Scholar] [CrossRef] [Green Version]
- Myers, R.H.; Myers, R.H. Classical and Modern Regression with Applications; Duxbury Press: Belmont, CA, USA, 1990; Volume 2. [Google Scholar]
- Cohen, J. Statistical Power Analysis for the Behavioral Sciences, 2nd ed.; Erlbaum: Hillsdale, NJ, USA, 1988. [Google Scholar]
- Sergiou, C.S.; Santarnecchi, E.; Romanella, S.M.; Wieser, M.J.; Franken, I.H.; Rassin, E.G.; van Dongen, J.D. tDCS targeting the Ventromedial Prefrontal Cortex reduces reactive aggression and modulates electrophysiological responses in a forensic population. Biol. Psychiatry Cogn. Neurosci. Neuroimaging 2021, 7, 95–107. [Google Scholar]
- Cheng, Y.; Hung, A.-Y.; Decety, J. Dissociation between affective sharing and emotion understanding in juvenile psychopaths. Dev. Psychopathol. 2012, 24, 623–636. [Google Scholar] [CrossRef] [PubMed]
- Venables, N.C.; Hall, J.R.; Yancey, J.R.; Patrick, C.J. Factors of psychopathy and electrocortical response to emotional pictures: Further evidence for a two-process theory. J. Abnorm. Psychol. 2015, 124, 319. [Google Scholar] [CrossRef] [PubMed]
- Klee, D.; Colgan, D.D.; Hanes, D.; Oken, B. The effects of an internet-based mindfulness meditation intervention on electrophysiological markers of attention. Int. J. Psychophysiol. 2020, 158, 103–113. [Google Scholar] [CrossRef] [PubMed]
- De Wied, M.; Van Boxtel, A.; Matthys, W.; Meeus, W. Verbal, facial and autonomic responses to empathy-eliciting film clips by disruptive male adolescents with high versus low callous-unemotional traits. J. Abnorm. Child Psychol. 2012, 40, 211–223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beauchaine, T.P.; Gatzke-Kopp, L.; Mead, H.K. Polyvagal theory and developmental psychopathology: Emotion dysregulation and conduct problems from preschool to adolescence. Biol. Psychol. 2007, 74, 174–184. [Google Scholar] [CrossRef] [Green Version]
- Barry, C.T.; Lui, J.H.; Anderson, A.C. Adolescent narcissism, aggression, and prosocial behavior: The relevance of socially desirable responding. J. Personal. Assess. 2017, 99, 46–55. [Google Scholar] [CrossRef] [PubMed]
- Smeets, K.; Oostermeijer, S.; Lappenschaar, M.; Cohn, M.; Van der Meer, J.; Popma, A.; Jansen, L.; Rommelse, N.; Scheepers, F.; Buitelaar, J.K. Are proactive and reactive aggression meaningful distinctions in adolescents? A variable-and person-based approach. J. Abnorm. Child Psychol. 2017, 45, 1–14. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Delinquent Young Adults (N = 118) | Aged-Matched Controls (N = 25) | |
|---|---|---|
| Age | 22.54 (2.41) | 22.86 (2.55) |
| Ethnicity | ||
| Western | 21 (17.8) | 11 (44.0) |
| Surinamese | 23 (19.5) | 6 (24.0) |
| Caribbean | 29 (24.6) | 2 (8.0) |
| Moroccan | 20 (16.9) | 1 (4.0) |
| Cape Verdean | 8 (6.8) | - |
| Other non-Western | 17 (14.4) | 5 (20.0) |
| Education | ||
| Senior secondary education | 31 (26.3) | 17 (68.0) |
| Junior secondary education | 37 (31.3) | 7 (28.0) |
| Primary education | 43 (35.4) | 1 (4.0) |
| None | 7 (5.9) | - |
| Past offenses (official records) | ||
| 0 | 23 (19.5) | - |
| 1–5 | 55 (46.6) | - |
| 5–10 | 24 (20.3) | - |
| >10 | 16 (13.6) | - |
| Lifetime delinquency (self-report) | ||
| Destruction/public order offense | 79 (67.5) | - |
| Property offense | 98 (85.2) | - |
| Aggression/violent offense | 79 (68.7) | - |
| Weapon offense | 49 (42.6) | - |
| Drug offense | 71 (61.2) | - |
| Any offense | 112 (95.7) | - |
| Delinquent Young Adults Mean (SD) | Aged-Matched Controls Mean (SD) | Group Differences p-Value | |
|---|---|---|---|
| Behavioral measure | (N = 118) | (N = 25) | |
| Reactive aggression | 11.38 (4.51) | 8.76 (5.23) | 0.026 * |
| Proactive aggression | 5.02 (3.89) | 4.00 (4.27) | 0.280 |
| Total aggression | 16.40 (7.34) | 12.76 (9.05) | 0.069 |
| Psychophysiological measures | (N = 96) | (N = 22) | |
| HR-aggressive | 65.70 (8.82) | 65.75 (8.75) | 0.981 |
| HR-neutral | 66.44 (8.70) | 66.51 (8.63) | 0.975 |
| RSA-aggressive | 91.24 (34.45) | 95.58 (45.28) | 0.677 |
| RSA-neutral | 95.33 (44.83) | 93.40 (41.67) | 0.849 |
| SCL-aggressive | 5.75 (2.65) | 4.08 (2.13) | 0.004 ** |
| SCL-neutral | 5.78 (2.68) | 4.09 (2.13) | 0.004 ** |
| Electrophysiological measures | (N = 110) | (N = 19) | |
| P3-aggressive | 4.43 (6.19) | 7.63 (4.57) | 0.012 * |
| P3-neutral | 4.72 (6.34) | 6.25 (4.64) | 0.221 |
| LPP-aggressive | 5.63 (4.67) | 7.66 (4.47) | 0.082 |
| LPP-neutral | 2.96 (4.70) | 3.13 (4.16) | 0.868 |
| Mu power-aggressive | 16.72 (14.32) | 14.68 (9.27) | 0.480 |
| Mu power-neutral | 17.58 (12.92) | 15.90 (10.44) | 0.574 |
| Outcome | Predictor | β | SE (β) | p | 95% CI β | |
|---|---|---|---|---|---|---|
| Lower | Upper | |||||
| Reactive aggression | Intercept | 11.565 | 0.661 | 0.000 | 10.252 | 12.879 |
| Δ HR | 0.035 | 0.287 | 0.902 | −0.535 | 0.606 | |
| Δ RSA | 0.002 | 0.015 | 0.909 | −0.028 | 0.031 | |
| Δ SCL | −3.113 | 3.274 | 0.346 | −9.667 | 3.443 | |
| Δ P3 | 0.093 | 0.134 | 0.489 | −0.174 | 0.360 | |
| Δ LPP | −0.136 | 0.136 | 0.319 | −0.405 | 0.133 | |
| Δ Mu power | 0.029 | 0.084 | 0.733 | −0.140 | 0.198 | |
| Proactive aggression | Intercept | 5.432 | 0.557 | 0.000 | 4.325 | 6.538 |
| Δ HR | 0.083 | 0.254 | 0.745 | −0.423 | 0.589 | |
| Δ RSA | −0.003 | 0.013 | 0.828 | −0.029 | 0.023 | |
| Δ SCL | −1.239 | 2.572 | 0.631 | −6.365 | 3.886 | |
| Δ P3 | 0.122 | 0.127 | 0.337 | −0.130 | 0.375 | |
| Δ LPP | −0.171 | 0.124 | 0.170 | −0.418 | 0.075 | |
| Δ Mu power | 0.000 | 0.090 | 0.996 | −0.182 | 0.183 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
ter Harmsel, J.F.; van Dongen, J.D.M.; Zijlmans, J.; van der Pol, T.M.; Marhe, R.; Popma, A. Neurobiological Responses towards Stimuli Depicting Aggressive Interactions in Delinquent Young Adults and Controls: No Relation to Reactive and Proactive Aggression. Brain Sci. 2022, 12, 124. https://doi.org/10.3390/brainsci12020124
ter Harmsel JF, van Dongen JDM, Zijlmans J, van der Pol TM, Marhe R, Popma A. Neurobiological Responses towards Stimuli Depicting Aggressive Interactions in Delinquent Young Adults and Controls: No Relation to Reactive and Proactive Aggression. Brain Sciences. 2022; 12(2):124. https://doi.org/10.3390/brainsci12020124
Chicago/Turabian Styleter Harmsel, Janna F., Josanne D. M. van Dongen, Josjan Zijlmans, Thimo M. van der Pol, Reshmi Marhe, and Arne Popma. 2022. "Neurobiological Responses towards Stimuli Depicting Aggressive Interactions in Delinquent Young Adults and Controls: No Relation to Reactive and Proactive Aggression" Brain Sciences 12, no. 2: 124. https://doi.org/10.3390/brainsci12020124