Connexin 43 Controls the Astrocyte Immunoregulatory Phenotype

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Mice
2.2. Study Approval
2.3. Antibodies
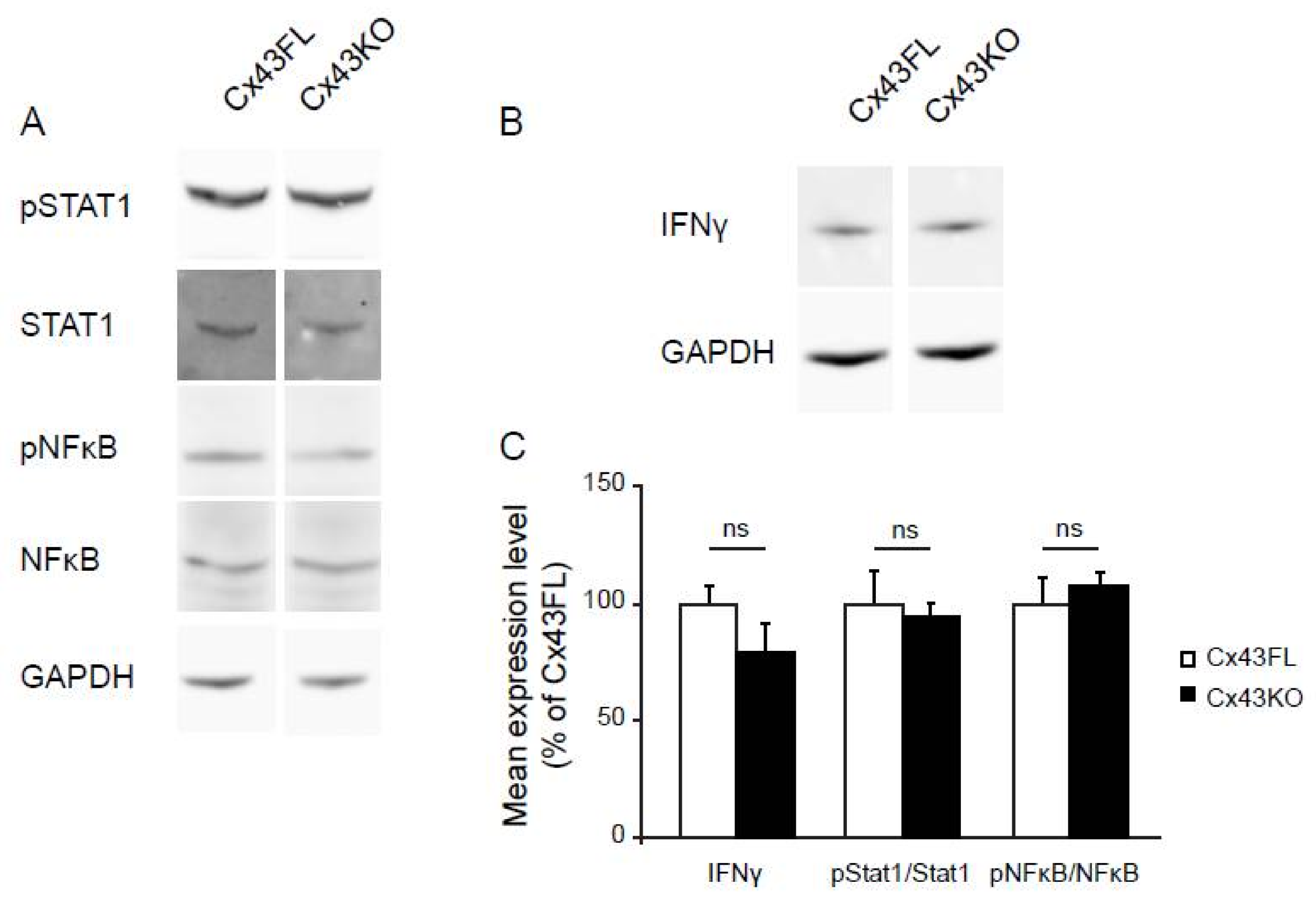
2.4. Western-Blot
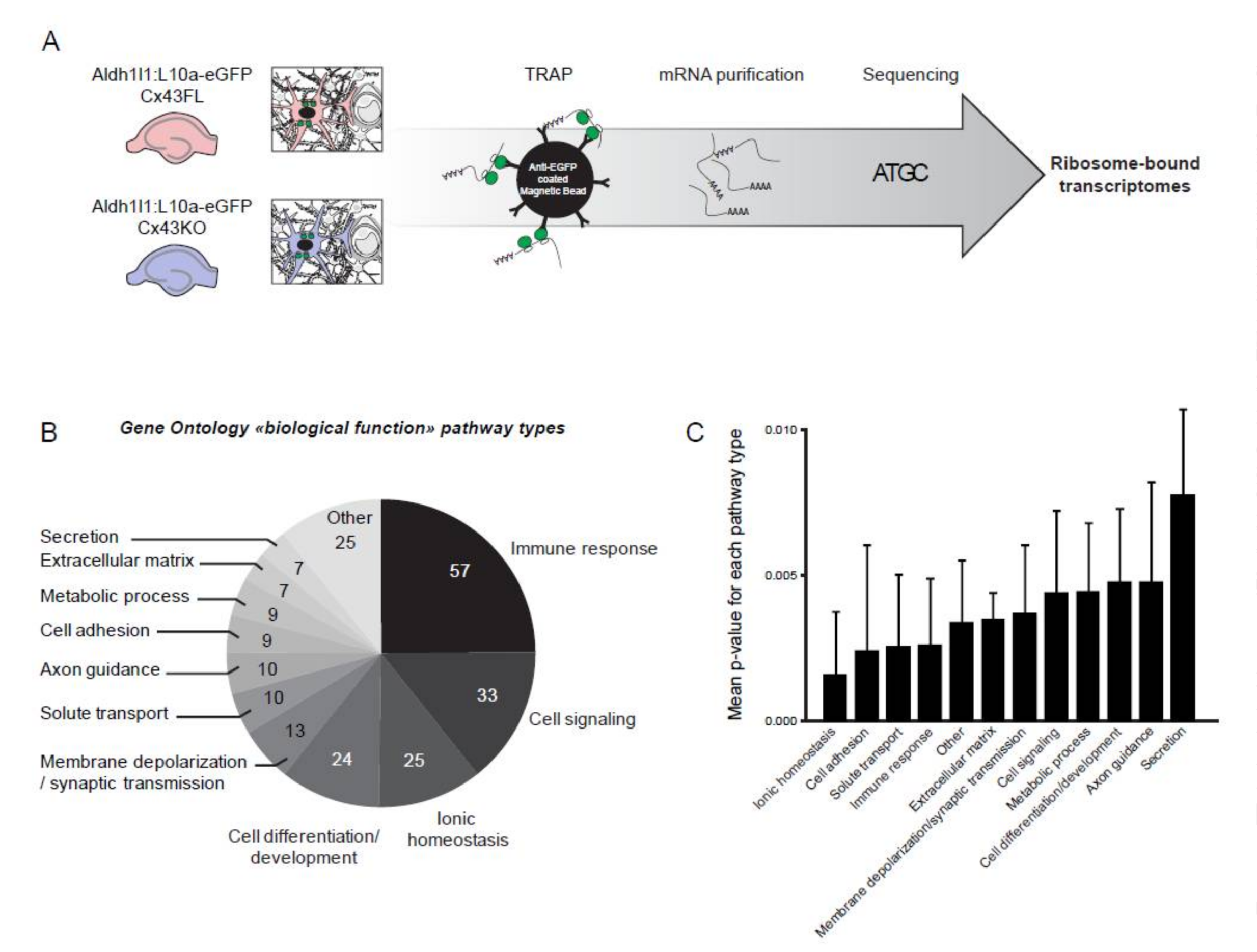
2.5. Translating Ribosome Affinity Purification (TRAP) in Astrocytes, RNA Sequencing and Analysis
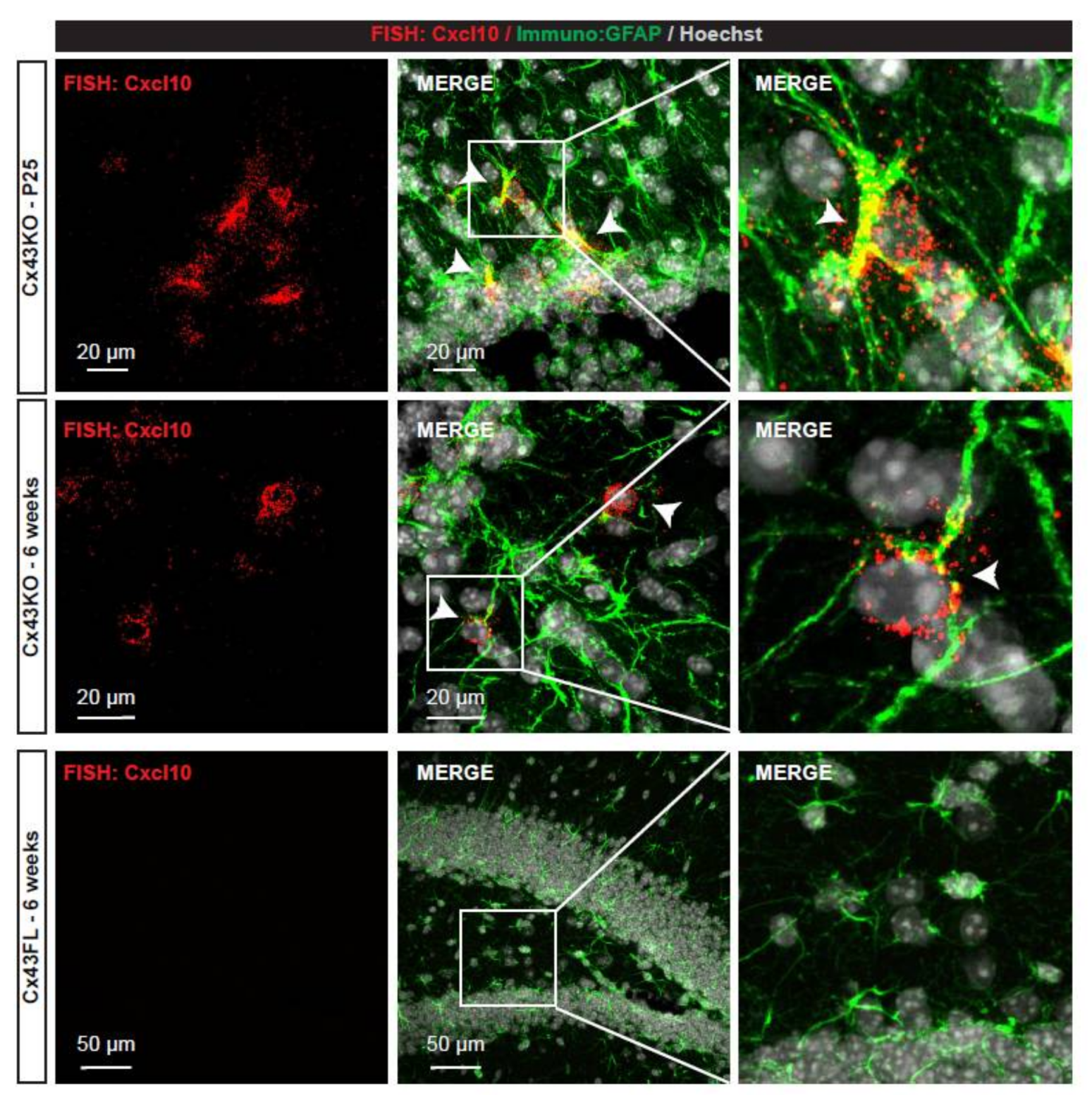
2.6. High Resolution Fluorescent In Situ Hybridization
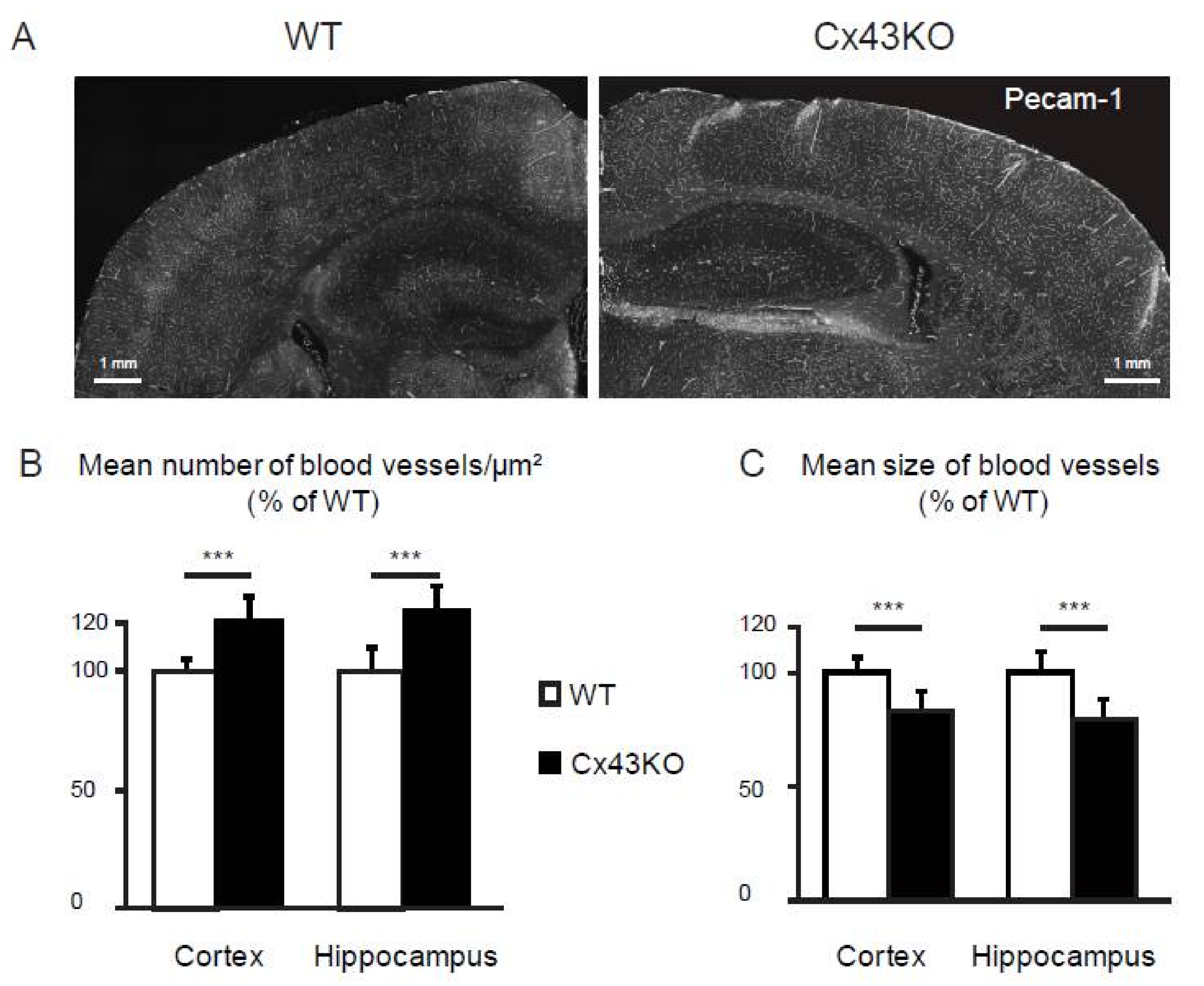
2.7. Vessel Density Calculation
3. Results
3.1. Characterization of the Ribosome-Bound Transcriptome in Cx43-Deleted Hippocampal Astrocytes
3.2. Cx43KO Astrocyte Reactive Profile Is Not Related to Classical Signaling Inflammatory Pathways
3.3. Microvascular Density Is Increased in Cx43KO Brain
4. Discussion
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Engelhardt, B.; Vajkoczy, P.; Weller, R.O. The movers and shapers in immune privilege of the CNS. Nat. Immunol. 2017, 18, 123–131. [Google Scholar] [CrossRef] [PubMed]
- Alvarez, J.I.; Katayama, T.; Prat, A. Glial influence on the blood brain barrier. Glia 2013, 61, 1939–1958. [Google Scholar] [CrossRef] [PubMed]
- Armulik, A.; Genove, G.; Mae, M.; Nisancioglu, M.H.; Wallgard, E.; Niaudet, C.; He, L.; Norlin, J.; Lindblom, P.; Strittmatter, K.; et al. Pericytes regulate the blood-brain barrier. Nature 2010, 468, 557–561. [Google Scholar] [CrossRef] [PubMed]
- Engelhardt, B.; Coisne, C. Fluids and barriers of the CNS establish immune privilege by confining immune surveillance to a two-walled castle moat surrounding the CNS castle. Fluids Barriers CNS 2011, 8, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zamanian, J.L.; Xu, L.; Foo, L.C.; Nouri, N.; Zhou, L.; Giffard, R.G.; Barres, B.A. Genomic analysis of reactive astrogliosis. J. Neurosci. 2012, 32, 6391–6410. [Google Scholar] [CrossRef] [PubMed]
- Liddelow, S.A.; Guttenplan, K.A.; Clarke, L.E.; Bennett, F.C.; Bohlen, C.J.; Schirmer, L.; Bennett, M.L.; Munch, A.E.; Chung, W.S.; Peterson, T.C.; et al. Neurotoxic reactive astrocytes are induced by activated microglia. Nature 2017, 541, 481–487. [Google Scholar] [CrossRef] [PubMed]
- Anderson, M.A.; Burda, J.E.; Ren, Y.; Ao, Y.; O’Shea, T.M.; Kawaguchi, R.; Coppola, G.; Khakh, B.S.; Deming, T.J.; Sofroniew, M.V. Astrocyte scar formation aids central nervous system axon regeneration. Nature 2016, 532, 195–200. [Google Scholar] [CrossRef] [PubMed]
- Broux, B.; Gowing, E.; Prat, A. Glial regulation of the blood-brain barrier in health and disease. Semin. Immunopathol. 2015, 37, 577–590. [Google Scholar] [CrossRef] [PubMed]
- Boulay, A.C.; Cisternino, S.; Cohen-Salmon, M. Immunoregulation at the gliovascular unit in the healthy brain: A focus on connexin 43. Brain Behav. Immun. 2016, 56, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Ezan, P.; Andre, P.; Cisternino, S.; Saubamea, B.; Boulay, A.C.; Doutremer, S.; Thomas, M.A.; Quenech’du, N.; Giaume, C.; Cohen-Salmon, M. Deletion of astroglial connexins weakens the blood-brain barrier. J. Cereb. Blood Flow Metab. 2012, 32, 1457–1467. [Google Scholar] [CrossRef] [PubMed]
- Lynn, B.D.; Tress, O.; May, D.; Willecke, K.; Nagy, J.I. Ablation of connexin30 in transgenic mice alters expression patterns of connexin26 and connexin32 in glial cells and leptomeninges. Eur. J. Neurosci. 2011, 34, 1783–1793. [Google Scholar] [CrossRef] [PubMed]
- Herve, J.C.; Derangeon, M.; Sarrouilhe, D.; Giepmans, B.N.; Bourmeyster, N. Gap junctional channels are parts of multiprotein complexes. Biochim. Biophys. Acta 2012, 1818, 1844–1865. [Google Scholar] [CrossRef] [PubMed]
- Boulay, A.C.; Mazeraud, A.; Cisternino, S.; Saubamea, B.; Mailly, P.; Jourdren, L.; Blugeon, C.; Mignon, V.; Smirnova, M.; Cavallo, A.; et al. Immune quiescence of the brain is set by astroglial connexin 43. J. Neurosci. 2015, 35, 4427–4439. [Google Scholar] [CrossRef] [PubMed]
- Theis, M.; Mas, C.; Doring, B.; Kruger, O.; Herrera, P.; Meda, P.; Willecke, K. General and conditional replacement of connexin43-coding DNA by a lacz reporter gene for cell-autonomous analysis of expression. Cell Commun. Adhes. 2001, 8, 383–386. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Dublin, P.; Griemsmann, S.; Klein, A.; Brehm, R.; Bedner, P.; Fleischmann, B.K.; Steinhauser, C.; Theis, M. Germ-line recombination activity of the widely used hgfap-cre and nestin-cre transgenes. PLoS ONE 2013, 8, e82818. [Google Scholar] [CrossRef] [PubMed]
- Doyle, J.P.; Dougherty, J.D.; Heiman, M.; Schmidt, E.F.; Stevens, T.R.; Ma, G.; Bupp, S.; Shrestha, P.; Shah, R.D.; Doughty, M.L.; et al. Application of a translational profiling approach for the comparative analysis of CNS cell types. Cell 2008, 135, 749–762. [Google Scholar] [CrossRef] [PubMed]
- Heiman, M.; Schaefer, A.; Gong, S.; Peterson, J.D.; Day, M.; Ramsey, K.E.; Suarez-Farinas, M.; Schwarz, C.; Stephan, D.A.; Surmeier, D.J.; et al. A translational profiling approach for the molecular characterization of CNS cell types. Cell 2008, 135, 738–748. [Google Scholar] [CrossRef] [PubMed]
- Heiman, M.; Kulicke, R.; Fenster, R.J.; Greengard, P.; Heintz, N. Cell type-specific mrna purification by translating ribosome affinity purification (trap). Nat. Protoc. 2014, 9, 1282–1291. [Google Scholar] [CrossRef] [PubMed]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed]
- Subramanian, A.; Tamayo, P.; Mootha, V.K.; Mukherjee, S.; Ebert, B.L.; Gillette, M.A.; Paulovich, A.; Pomeroy, S.L.; Golub, T.R.; Lander, E.S.; et al. Gene set enrichment analysis: A knowledge-based approach for interpreting genome-wide expression profiles. Proc. Natl. Acad. Sci. USA 2005, 102, 15545–15550. [Google Scholar] [CrossRef] [PubMed]
- Bachoo, R.M.; Kim, R.S.; Ligon, K.L.; Maher, E.A.; Brennan, C.; Billings, N.; Chan, S.; Li, C.; Rowitch, D.H.; Wong, W.H.; et al. Molecular diversity of astrocytes with implications for neurological disorders. Proc. Natl. Acad. Sci. USA 2004, 101, 8384–8389. [Google Scholar] [CrossRef] [PubMed]
- Nikitin, A.; Egorov, S.; Daraselia, N.; Mazo, I. Pathway studio—The analysis and navigation of molecular networks. Bioinformatics 2003, 19, 2155–2157. [Google Scholar] [CrossRef] [PubMed]
- Pannasch, U.; Vargova, L.; Reingruber, J.; Ezan, P.; Holcman, D.; Giaume, C.; Sykova, E.; Rouach, N. Astroglial networks scale synaptic activity and plasticity. Proc. Natl. Acad. Sci. USA 2011, 108, 8467–8472. [Google Scholar] [CrossRef] [PubMed]
- Clarke, L.E.; Liddelow, S.A.; Chakraborty, C.; Munch, A.E.; Heiman, M.; Barres, B.A. Normal aging induces a1-like astrocyte reactivity. Proc. Natl. Acad. Sci. USA 2018, 115, E1896–E1905. [Google Scholar] [CrossRef] [PubMed]
- Han, R.; Luo, J.; Shi, Y.; Yao, Y.; Hao, J. Pd-l1 (programmed death ligand 1) protects against experimental intracerebral hemorrhage-induced brain injury. Stroke 2017, 48, 2255–2262. [Google Scholar] [CrossRef] [PubMed]
- Rosciszewski, G.; Cadena, V.; Murta, V.; Lukin, J.; Villarreal, A.; Roger, T.; Ramos, A.J. Toll-like receptor 4 (TLR4) and triggering receptor expressed on myeloid cells-2 (TREM-2) activation balance astrocyte polarization into a proinflammatory phenotype. Mol. Neurobiol. 2017, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Colombo, E.; Farina, C. Astrocytes: Key regulators of neuroinflammation. Trends Immunol. 2016, 37, 608–620. [Google Scholar] [CrossRef] [PubMed]
- Kudriaeva, A.; Galatenko, V.V.; Maltseva, D.V.; Khaustova, N.A.; Kuzina, E.; Tonevitsky, A.G.; Gabibov, A.; Belogurov, A. The transcriptome of type I murine astrocytes under interferon-gamma exposure and remyelination stimulus. Molecules 2017, 22, 808. [Google Scholar] [CrossRef] [PubMed]
- Hidano, S.; Randall, L.M.; Dawson, L.; Dietrich, H.K.; Konradt, C.; Klover, P.J.; John, B.; Harris, T.H.; Fang, Q.; Turek, B.; et al. Stat1 signaling in astrocytes is essential for control of infection in the central nervous system. mBio 2016, 7, e01881-16. [Google Scholar] [CrossRef] [PubMed]
- Ben Haim, L.; Ceyzeriat, K.; Carrillo-de Sauvage, M.A.; Aubry, F.; Auregan, G.; Guillermier, M.; Ruiz, M.; Petit, F.; Houitte, D.; Faivre, E.; et al. The jak/stat3 pathway is a common inducer of astrocyte reactivity in alzheimer’s and Huntington’s diseases. J. Neurosci. 2015, 35, 2817–2829. [Google Scholar] [CrossRef] [PubMed]
- Navankasattusas, S.; Whitehead, K.J.; Suli, A.; Sorensen, L.K.; Lim, A.H.; Zhao, J.; Park, K.W.; Wythe, J.D.; Thomas, K.R.; Chien, C.B.; et al. The netrin receptor unc5b promotes angiogenesis in specific vascular beds. Development 2008, 135, 659–667. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.K.; Chen, H.; Mao, F.; Xiao, Q.G.; Xie, R.F.; Lei, T. Downregulation of LRIG2 expression inhibits angiogenesis of glioma via EGFR/VEGF-A pathway. Oncol. Lett. 2017, 14, 4021–4028. [Google Scholar] [CrossRef] [PubMed]
- Anderson, M.A.; Ao, Y.; Sofroniew, M.V. Heterogeneity of reactive astrocytes. Neurosci. Lett. 2014, 565, 23–29. [Google Scholar] [CrossRef] [PubMed]
- Liddelow, S.A.; Barres, B.A. Reactive astrocytes: Production, function, and therapeutic potential. Immunity 2017, 46, 957–967. [Google Scholar] [CrossRef] [PubMed]
- Sofroniew, M.V. Molecular dissection of reactive astrogliosis and glial scar formation. Trends Neurosci. 2009, 32, 638–647. [Google Scholar] [CrossRef] [PubMed]
- Wilcox, R.A.; Feldman, A.L.; Wada, D.A.; Yang, Z.Z.; Comfere, N.I.; Dong, H.; Kwon, E.D.; Novak, A.J.; Markovic, S.N.; Pittelkow, M.R.; et al. B7-H1 (PD-L1, CD274) suppresses host immunity in T-cell lymphoproliferative disorders. Blood 2009, 114, 2149–2158. [Google Scholar] [CrossRef] [PubMed]
- Van Beek, J.; van Meurs, M.; Ba, T.H.; Brok, H.P.; Neal, J.W.; Chatagner, A.; Harris, C.L.; Omidvar, N.; Morgan, B.P.; Laman, J.D.; et al. Decay-accelerating factor (CD55) is expressed by neurons in response to chronic but not acute autoimmune central nervous system inflammation associated with complement activation. J. Immunol. 2005, 174, 2353–2365. [Google Scholar] [CrossRef] [PubMed]
- Argaw, A.T.; Asp, L.; Zhang, J.; Navrazhina, K.; Pham, T.; Mariani, J.N.; Mahase, S.; Dutta, D.J.; Seto, J.; Kramer, E.G.; et al. Astrocyte-derived VEGF-A drives blood-brain barrier disruption in CNS inflammatory disease. J. Clin. Investig. 2012, 122, 2454–2468. [Google Scholar] [CrossRef] [PubMed]
- Horng, S.; Therattil, A.; Moyon, S.; Gordon, A.; Kim, K.; Argaw, A.T.; Hara, Y.; Mariani, J.N.; Sawai, S.; Flodby, P.; et al. Astrocytic tight junctions control inflammatory CNS lesion pathogenesis. J. Clin. Investig. 2017, 127, 3136–3151. [Google Scholar] [CrossRef] [PubMed]
- Ries, M.; Sastre, M. Mechanisms of abeta clearance and degradation by glial cells. Front Aging Neurosci. 2016, 8, 160. [Google Scholar] [CrossRef] [PubMed]
- Toyn, J.H.; Lin, X.A.; Thompson, M.W.; Guss, V.; Meredith, J.E., Jr.; Sankaranarayanan, S.; Barrezueta, N.; Corradi, J.; Majumdar, A.; Small, D.L.; et al. Viable mouse gene ablations that robustly alter brain abeta levels are rare. BMC Neurosci. 2010, 11, 143. [Google Scholar] [CrossRef] [PubMed]
- Yi, C.; Koulakoff, A.; Giaume, C. Astroglial connexins as a therapeutic target for Alzheimer’s disease. Curr. Pharm. Des. 2017, 23, 4958–4968. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.J.; Cheng, C.; Qiu, Z.; Zhou, W.H.; Cheng, G.Q. Decreased connexin 43 in astrocytes inhibits the neuroinflammatory reaction in an acute mouse model of neonatal sepsis. Neurosci. Bull. 2015, 31, 763–768. [Google Scholar] [CrossRef] [PubMed]
- Lioy, D.T.; Garg, S.K.; Monaghan, C.E.; Raber, J.; Foust, K.D.; Kaspar, B.K.; Hirrlinger, P.G.; Kirchhoff, F.; Bissonnette, J.M.; Ballas, N.; et al. A role for glia in the progression of Rett’s syndrome. Nature 2011, 475, 497–500. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Astrocyte Ribosome-Bound mRNAs (Hippocampus) | Astrocyte Total mRNAs (Spinal Cord) | ||||||
|---|---|---|---|---|---|---|---|
| Gene Name | Molecule Name | Mean Reads Cx43FL | Mean Reads Cx43KO | Fold Change in Cx43KO Astrocytes | Padj | Fold Change in WT SCI Astrocytes | Fold Change in STAT3 KO SCI Astrocytes |
| Inflammation | |||||||
| Ccl5 | chemokine (C-C motif) ligand 5 | 0.4 | 139.3 | 17.9 | 1.2 × 107 | 9.6 | 10.34 |
| Itgam | integrin alpha M | 4.8 | 314.8 | 14.2 | 9.1 × 105 | 7 | 8.02 |
| C3ar1 | complement component 3a receptor 1 | 0.8 | 63.2 | 13.5 | 6.7 × 104 | 11.8 | 12.22 |
| Cd34 | CD34 antigen | 2.8 | 140.4 | 13.1 | 4.3 × 104 | 9 | 6.82 |
| Cd53 | chemokine (C-C motif) ligand 5 | 2.0 | 151.0 | 14.2 | 1.2 × 104 | 7.4 | 7.5 |
| Cspg4 | chondroitin sulfate proteoglycan 4 | 4.1 | 127.9 | 11.4 | 7.2 × 103 | 4.5 | 5.54 |
| Lyz2 | lysozyme 2 | 39.5 | 764.4 | 11.3 | 1.5 × 104 | 17.8 | 16.52 |
| Gbp4 | guanylate binding protein 4 | 3.7 | 83.2 | 11.2 | 1.8 × 103 | 10.34 | 19.5 |
| Slfn5 | schlafen 5 | 5.2 | 93.6 | 9.6 | 4.1 × 102 | 14.24 | 12.1 |
| Igtp | interferon gamma induced GTPase | 66.6 | 706.6 | 9.7 | 1.6 × 107 | 5.4 | 10.2 |
| Iigp1 | interferon inducible GTPase 1 | 54.8 | 803.7 | 9.5 | 2.1 × 102 | 9.2 | 17.34 |
| Psmb9 | proteasome (prosome, macropain) subunit, beta type 9 (large multifunctional peptidase 2) | 78.7 | 920.7 | 9.0 | 1.8 × 102 | 5.98 | 9.12 |
| C1qb | complement component 1, q subcomponent, beta polypeptide | 35.2 | 328.6 | 8.3 | 2.9 × 102 | 8.02 | 8.84 |
| Scrt1 | scratch homolog 1, zinc finger protein (Drosophila) | 76.9 | 598.0 | 8.2 | 9.0 × 104 | ns | ns |
| Cxcl10 | chemokine (C-X-C motif) ligand 10 | 80.8 | 663.5 | 8.1 | 1.4 × 102 | 11.56 | 12.52 |
| Fyb | FYN binding protein | 27.0 | 185.8 | 7.6 | 9.8 × 103 | ns | 10.08 |
| Faah | fatty acid amide hydrolase | 124.9 | 862.3 | 7.5 | 2.7 × 102 | 6.4 | ns |
| Anti-inflammation | |||||||
| Cd55 | CD55 antigen | 12.4 | 511.9 | 13.2 | 1.2 × 104 | ns | ns |
| Cd274 | CD274 antigen | 43.8 | 657.4 | 10.5 | 2.3 × 104 | ns | 6.94 |
| L1cam | L1 cell adhesion molecule | 61.2 | 757.7 | 9.9 | 2.2 × 104 | ns | ns |
| Best3 | bestrophin 3 | 7.4 | 110.9 | 9.6 | 1.7 × 102 | ns | ns |
| Serpina3n | serine (or cysteine) peptidase inhibitor, clade A, member 3N | 202.7 | 1709.4 | 8.3 | 7.8 × 103 | 8.14 | ns |
| Cellular junction | |||||||
| Cldn3 | claudin 3 | 0.0 | 60.2 | 15.2 | 2.8 × 104 | ns | ns |
| Cldn9 | claudin 9 | 2.0 | 104.3 | 12.9 | 1.1 × 103 | ns | ns |
| Pcdhb18 | protocadherin beta 18 | 27.4 | 249.1 | 8.7 | 2.7 × 103 | ns | ns |
| Cdh12 | cadherin 12 | 75.2 | 579.6 | 7.8 | 2.1 × 102 | ns | ns |
| Unc5b | unc-5 homolog B (C. elegans) | 17.3 | 264.5 | 9.8 | 1.1 × 102 | ns | ns |
| Angiogenesis | |||||||
| Lrig2 | leucine-rich repeats and immunoglobulin-like domains 2 | 137.8 | 953.6 | 7.7 | 8.6 × 103 | ns | ns |
| Ccbe1 | collagen and calcium binding EGF domains 1 | 49.8 | 394.3 | 8.1 | 6.3 × 103 | ns | ns |
| Glioma | |||||||
| Cmtm7 | CKLF-like MARVEL transmembrane domain containing 7 | 3.6 | 66.2 | 9.9 | 2.4 × 102 | 9.2 | 7.66 |
| Rbl1 | retinoblastoma-like 1 (p107) | 21.3 | 266.9 | 9.7 | 1.5 × 103 | ns | ns |
| Cd109 | CD109 antigen | 4.0 | 58.1 | 9.2 | 3.4 × 102 | 3.74 | 3.22 |
| Ion Transport | |||||||
| Slc9a4 | solute carrier family 9 (sodium/hydrogen exchanger), member 4 | 6.0 | 115.2 | 10.2 | 1.3 × 102 | ns | ns |
| Slc8a2 | solute carrier family 8 (sodium/calcium exchanger), member 2 | 94.9 | 740.6 | 8.4 | 1.1 × 104 | ns | ns |
| Scnn1a | sodium channel, nonvoltage-gated 1 alpha | 24.8 | 208.2 | 8.4 | 4.5 × 103 | ns | ns |
| Slc16a4 | solute carrier family 16 (monocarboxylic acid transporters), member 4 | 42.7 | 397.4 | 8.3 | 2.6 × 102 | ns | ns |
| Cacna2d3 | calcium channel, voltage-dependent, alpha2/delta subunit 3 | 56.8 | 407.6 | 7.8 | 1.0 × 102 | ns | ns |
| Aβ degradation | |||||||
| Mme | membrane metallo endopeptidase | 6.0 | 141.3 | 11.8 | 1.2 × 104 | ns | ns |
| Pigz | phosphatidylinositol glycan anchor biosynthesis, class Z | 6.4 | 122.5 | 10.3 | 1.2 × 102 | ns | ns |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Boulay, A.-C.; Gilbert, A.; Oliveira Moreira, V.; Blugeon, C.; Perrin, S.; Pouch, J.; Le Crom, S.; Ducos, B.; Cohen-Salmon, M. Connexin 43 Controls the Astrocyte Immunoregulatory Phenotype. Brain Sci. 2018, 8, 50. https://doi.org/10.3390/brainsci8040050
Boulay A-C, Gilbert A, Oliveira Moreira V, Blugeon C, Perrin S, Pouch J, Le Crom S, Ducos B, Cohen-Salmon M. Connexin 43 Controls the Astrocyte Immunoregulatory Phenotype. Brain Sciences. 2018; 8(4):50. https://doi.org/10.3390/brainsci8040050
Chicago/Turabian StyleBoulay, Anne-Cécile, Alice Gilbert, Vanessa Oliveira Moreira, Corinne Blugeon, Sandrine Perrin, Juliette Pouch, Stéphane Le Crom, Bertrand Ducos, and Martine Cohen-Salmon. 2018. "Connexin 43 Controls the Astrocyte Immunoregulatory Phenotype" Brain Sciences 8, no. 4: 50. https://doi.org/10.3390/brainsci8040050