Natural Killer Cells: Tumor Surveillance and Signaling


1
The Walter and Eliza Hall Institute of Medical Research, Parkville, VIC 3052, Australia
2
Department of Medical Biology, The University of Melbourne, Parkville, VIC 3010, Australia
*
Authors to whom correspondence should be addressed.
Cancers 2020, 12(4), 952; https://doi.org/10.3390/cancers12040952
Submission received: 4 March 2020
/
Revised: 3 April 2020
/
Accepted: 9 April 2020
/
Published: 11 April 2020
(This article belongs to the Special Issue Cancer Immunotherapy Using Natural Killer Cells: Successes and Challenges of the Field)
Abstract
:Natural killer (NK) cells play a pivotal role in cancer immunotherapy due to their innate ability to detect and kill tumorigenic cells. The decision to kill is determined by the expression of a myriad of activating and inhibitory receptors on the NK cell surface. Cell-to-cell engagement results in either self-tolerance or a cytotoxic response, governed by a fine balance between the signaling cascades downstream of the activating and inhibitory receptors. To evade a cytotoxic immune response, tumor cells can modulate the surface expression of receptor ligands and additionally, alter the conditions in the tumor microenvironment (TME), tilting the scales toward a suppressed cytotoxic NK response. To fully harness the killing power of NK cells for clinical benefit, we need to understand what defines the threshold for activation and what is required to break tolerance. This review will focus on the intracellular signaling pathways activated or suppressed in NK cells and the roles signaling intermediates play during an NK cytotoxic response.
1. Introduction
Natural Killer (NK) cells are bone marrow–derived innate lymphocytes that are found in most organs, with the largest population of NK cells residing in the blood [1]. NK cells are large granular lymphocytes that were initially defined by their ability to kill tumor cells without prior sensitization [2,3]. The role of NK cells has since been expanded to include the elimination of virally infected cells and secretion of cytokines that mediate crosstalk and regulation of other immune cells [4].
Following their discovery in the 1970s, immunologists have been fascinated by the ability of NK cells to detect and kill tumorigenic or virally-infected cells, whilst tolerating healthy self-tissue [5,6,7]. However, it wasn’t until the early 1990s that scientists started to explore the mechanisms by which NK cells distinguished “self” from “non-self,” an area of research instigated by Klas Kärre’s exposition of the “missing self” theory [8]. Karre hypothesized that NK cells could recognize loss or reduction in surface expression of major histocompatibility complex (MHC) class I proteins (human leukocyte antigens (HLA) class I in humans), triggering recognition as non-self. This hypothesis was based on earlier studies by Strokus et al. [9,10] that described protection of susceptible cells with experimentally expressed MHC-I. The ‘missing-self’ theory was further validated by Karlhofer et al. [11], who showed that the murine Lymphocyte Ag 49A (Ly49A) receptor recognized and discriminated between different MHC-I molecules, with tumor cells from H2d and H2K backgrounds resistant to killing by Ly49A expressing NK cells [11,12]. Soon after, Moretta et al. [13] discovered the first human inhibitory NK receptor Killer cell immunoglobulin-like receptor 2DL1 (KIR2DL). Inhibitory receptors are now known to not only sense reduction in expression of MHC class I proteins, but also recognize non-MHC-I molecules, such as glycans and collagen, which are crucial for NK cell discrimination of self [14]. Upon engagement of cognate ligands, the various NK cell receptors send activating and inhibitory signals, which collectively determine NK cell action.
NK cells kill infected and transformed cells via a variety of mechanisms, including the delivery of lytic granules loaded with proteases and pore-forming proteins such as granzymes and perforin, release of cytokines such as tumor necrosis factor alpha (TNFα) and interferon gamma (IFNγ), upregulation of FASL and TNF-related apoptosis-inducing ligand (TRAIL) and by antibody-dependent cellular cytotoxicity (ADCC) [15,16,17,18,19]. There are multiple steps between NK cell: target cell engagement and cell killing, with receptor-ligand interactions thought to be the initiating step in the formation of an immunological synapse (IS) (Figure 1A,B) [20,21]. This is followed by recruitment of filamentous actin (F-actin) to the IS (Figure 1B) and polarization of the lytic granules and the microtubule-organizing center (MTOC) toward the IS (Figure 1C). Then, the granules dock at the synapse and are ready for the final step: granule-membrane fusion and release of the cytotoxic contents at the center of the IS (Figure 1D) [22,23]. NK cell signaling and killing is considered to be localized to the IS [24], with each NK cell thought to reach exhaustion after killing four to seven target cells [25].
NK cells represent approximately 10% of the circulating lymphocyte population and thus predominantly control hematologic malignancies and tumor metastasis, rather than solid tumors [26]. This is reflected by only minor NK cell infiltration in nascent and fully developed tumor microenvironments, such as in colorectal cancer “encapsulated” by tissue barriers [27,28]. Importantly, the presence of circulating NK cells is inversely correlated with metastatic burden in patients suffering from different carcinomas [29,30,31], gastrointestinal sarcoma (GIST) [32], melanoma [33], and breast cancer [34] and suggests that enhancing NK infiltration into tumors and/or activity would have clinical benefit. Early clinical trials for non-Hodgkin’s lymphoma and chronic lymphocytic leukemia (CLL) have shown positive patient responses following adoptive transfer of allogeneic chimeric antigen receptor (CAR)-NK cells, without significant toxicities [35]. This recent advance by Liu et al. [35] underscores the clinical utility of NK cell–based therapies.
The remainder of this review highlights the importance of individual NK receptors and our current understanding of how they function. Understanding the interplay between NK cell, tumor, and TME has the potential to not only lead to new cancer therapies, but inform which patients are likely to respond to receptor-focused interventions.
2. Receptor Mediated Inhibition of NK Cells: Inhibitory Receptors
NK cell self-tolerance in humans and mice is mostly mediated by inhibitory receptors that recognize either MHC-I complexes or non-MHC-I surface molecules [36]. In general, inhibitory receptors belong to receptor families comprised of both activating and inhibitory members, and signal through a cytoplasmic signaling tail containing an immunoreceptor tyrosine-based inhibitory motif (ITIM) (Table 1, Figure 2) [37]. NK killing of target cells can be viewed as an internal decision-making process using a “pros and cons” list; when activating signals outweigh inhibitory signals, the NK cell becomes cytotoxic. Interestingly, NK cells lacking inhibitory receptors are unable to become cytotoxic, thus acquiring functional maturation is also dependent on inhibitory signals. This requirement is commonly referred to as NK cell “education” [38].
2.1. MHC-I Recognizing Receptors
MHC-I complexes can be divided into classical (MHC-Ia) and non-classical (MHC-Ib) complexes [39]. The up-regulation of particular inhibitory receptors leads to NK cell education by either classical MHC-Ia-dependent (licensing), non-classical MHC-Ib-dependent recognition, or MHC-independent recognition [40].
Inhibitory MHC receptors are divided into the following three categories: the KIR and Leukocyte Immunoglobulin-Like Receptor (LILR, LIR, ILT, CD85) family in humans; the Ly49 family, also known as the Killer Cell Lectin-like Receptor subfamily A (klra) in mice; and lastly, the CD94/NKG2 receptor family, found in both mouse and human (Figure 2A). KIRs and Ly49s are so-called functional homologues as they bind MHC-I molecules yet are different at the protein and genetic sequence level [41,42]. Despite these differences, they both appear to have arisen from multiple duplication events, making KIRs and Ly49s highly polymorphic.
Inhibitory (i)KIRs are composed of a long (L) intracellular ITIM-containing region, a transmembrane domain, and an extracellular region with either two or three C2-type immunoglobulin (Ig)-like domains (2/3 D) [43,44]. iKIR family members are named based on their domain architecture, thus iKIRs can be either KIR2DL1-3,5 or KIR3DL1-5. Human NK cells express up to six iKIRs that are able to differentiate between HLA allotypes, as well as recognize different moieties on HLA molecules (Table 2). For example, KIR3DL2 recognizes HLA-A3 and -A11 allotypes, KIR3DL1 recognizes HLA-B allotypes containing a Bw4 epitope, and KIR2DL1-3 recognizes HLA-C [45,46,47]. In combination, these KIRs cover all HLA-isoforms.
The LILR family includes many inhibitory receptors, but only LILRB1 is expressed on NK cells and is capable of recognizing both classical and non-classical HLA-I molecules [48,49]. LILRB1 has four Ig-like domains in the extracellular domain and four cytoplasmic ITIMs [50,51]. Unlike other inhibitory receptors, LILRB1 is mostly known as a receptor for viral infection and until recently, was thought to have little or no effect in tumor immunity. However, given that different breast and colorectal patient cancers upregulate the LILRB1 ligand HLA-G as a means of evading NK targeting and killing, LILRB1 is likely to be important in tumor immunity [52,53,54,55].
The Ly49 receptor family contains 23 different members that are mostly encoded by a highly polymorphic Ly49 gene cluster located on chromosome 6 [90]. Although most members are expressed on NK cells, not all are expressed in all mice strains. NOD and 129 mice express most Ly49 members (15 and 13), with C57BL/6 (11) and BALB/c mice expressing relatively few members (8) [56,58,91]. Inhibitory Ly49 receptors (iLy49s) function as homodimers; each subunit is composed of a single C-type lectin domain (CTLD), an extracellular stalk region, a transmembrane region and a C-terminal cytoplasmic tail containing an ITIM. iLy49s bind both classical and non-classical MHC-I molecules through a binding site created by homodimerization of the two single CTLDs [56,57,58]. Although iLy49 binding requires MHC-I loaded with peptide, binding occurs in a peptide-independent manner, except for Ly49C, which has been shown to confer protection by binding H2-Kb haplotype loaded with specific peptides (from ovalbumin, vesicular stomatitis virus, and elongation factor, but not peptide from Sendai virus) [57,92]. Of the iLy49s present on murine NK cells, only Ly49A and Ly49C bind non-classical MHC-I ligands H2-M3 and H2-Q10, respectively [77,78,93].
The CD94:NKG2 receptor complex consists of a heterodimer between CD94 (also known as killer cell lectin-like receptor subfamily D; KLRD1) and a member of the greater NKG2 family and contributes to non-classical MHC-I-dependent education [59]. When the complex contains NKG2A or NKG2B (a splice variant of NKG2A), the receptor complex is inhibitory; however, when CD94 is complexed with other NKG2 family members it can generate an activating signal [94]. The inhibitory complex signals through two ITIMs present in the cytoplasmic tail of NKG2A and B [95,96]. NKG2A and B are the only two inhibitory NKG2 family members and recognize the same non-classical MHC-1b ligand: HLA-E in humans or its homologue Qa-1 in mice [59,60]. NKG2A and B expressing NK cells degranulate better and produce more IFNγ in the absence of HLA-E or Qa-1 [97,98] and are better killers compared to NKG2A and B negative NK cells which are considered to be uneducated. Although the activating receptor subunit NKG2C binds the same non-classical MHC-1b ligand as iNKG2A and B, the iNKG2A and B receptors display considerably higher ligand affinity, explaining why the NKG2-HLA-E/Qa-1 interaction is mostly considered relevant for maintaining self-tolerance and not activation [99].
2.2. Non-MHC-I Recognizing Receptors
NK cells also express various inhibitory receptors that recognize non-MHC molecules on healthy cells. These include T cell immunoglobulin and ITIM domain (TIGIT); carcinoembryonic Ag cell adhesion molecule 1 (CEACAM1); soluble leukocyte-associated Ig-like receptor-1 (LAIR-1); Killer cell lectin-like receptor G1 (KLRG1) and NKR-P1(A/B); sialic acid-binding immunoglobulin-like lectin (Siglec); and the Tyro3, Axl, and MerTK (TAM) receptors [100,101]. Like the KIRs, the TIGIT, TACTILE (CD96), LAIR-1, and CEACAM1 receptors are members of the Ig-like superfamily and share a similar domain architecture comprised of an extracellular region containing the Ig-like domains, a transmembrane domain and a cytoplasmic signaling tail (Figure 2A).
TIGIT and TACTILE bind to the poliovirus receptor (CD155, PVR) and nectin-2 (CD112, PVRL2), ligands which are normally expressed by antigen presenting cells and T cells [82,83]. However, CD155 and CD112 are also frequently expressed by tumorigenic cells, presumably as an immune escape mechanism. NK cells try to counter this evasion tactic by expressing DNAX accessory molecule-1 (DNAM-1; also known as CD226), which competes with TIGIT and TACTILE for binding to CD155 and CD112 [102]. TIGIT has the highest affinity for the ligands, followed by TACTILE, with DNAM-1 having the lowest affinity, suggesting that NK expression of DNAM-1 may not be a particularly effective counter-tactic [103,104]. The cytoplasmic signaling tails of human and murine TIGIT contain an ITIM and ITT motif, whereas murine TACTILE has only an ITIM motif, and human TACTILE has an additional YxxM activating motif (Table 3) [105]. TIGIT is the most well understood and blocking TIGIT gives rise to a potent NK anti-tumor effect, which is attributed to both loss of NK inhibition and enhanced activation. Recently, Zhang et al. [83] showed that TIGIT blockade appeared to reverse NK cell exhaustion, as well as enhance NK mediated anti-tumor responses in an experimental colon cancer model, both alone and in combination with anti-PD1 (programmed cell death protein 1) and anti-PD-L1 (programmed death-ligand 1).
PD-1 and PD-L1 are well-known T cell checkpoints commonly targeted in cancer immunotherapy [106]. PD-1 recognizes two ligands, PD-L1 (B7-H1) and PD-L2 (programmed death-ligand 2), but only PD-L1 is constitutively expressed on hematopoietic and non-hematopoietic cells, explaining why intervention has focused on the PD-1:PD-L1 interaction [107,108]. PD-1 and PD-L1 contain a similar domain architecture encompassing an Ig-like extracellular binding domain, a transmembrane region, and a cytoplasmic tail [109,110]. The cytoplasmic tail of PD-1 contains an ITIM and ITSM motif, while the cytoplasmic tail of PD-L1 is short, lacks the motifs and does not have a known function.
In recent years, multiple studies have investigated a role for these checkpoints in NK cells, with no clear consensus reached in the field. Some studies suggest that PD-1 is expressed solely on activated NK cells [111], while others have identified PD-1 expression as a marker for dysfunctional or exhausted NK cells [112,113,114]. Most recently, Judge et al. [115] compared PD-1 expression on mouse, human- and canine, T cells and NK cells under various conditions in vitro and investigated intratumoral NK cells in sarcoma, colon cancer, and head and neck squamous carcinoma, concluding that PD-1 was not significantly expressed on mouse or human NK cells. However, there may still be an argument for combining anti-PD-1 therapy with other checkpoint blockers to rescue exhausted NK cells [114].
T cell immunoglobulin and mucin domain-containing protein 3 (TIM-3) and Lymphocyte-activation gene 3 (LAG-3) are newly established T cell checkpoints and targets for cancer therapy, with studies suggesting an emerging role for these proteins in NK cells [116]. TIM-3 recognizes galectin-9, phospholipid phosphatidylserine (PtdSer), alarmin high mobility group box 1 (HMGB1) and carcinoembryonic antigen-related cell adhesion molecule 1 (CEACAM-1) via its Ig-like extracellular domain [117,118,119,120]. Although the intracellular tail of TIM-3 contains five tyrosines that are conserved between humans and mice and are potential phosphorylation sites, they don’t conform to known binding motifs [121]. TIM-3 is expressed by NK cells and is reported to not only inhibit NK cell cytotoxicity [122], but also mediate IFNγ production [123]. So et al. were unable to reproduce these specific findings and argued that TIM-3 could be used as a marker for end-stage NK cell activation, with TIM-3 blockade likely to affect NK cell activation [124].
LAG-3 binds to MHC-II on APCs via four Ig-like domains in the extracellular region and signals via a “KIELE” motif present in the cytoplasmic tail [125,126,127]. Although NK cells express LAG-3, studies exploring its biological relevance are still limited. LAG-3 deficiency in mice resulted in decreased NK cell cytotoxicity [128], while anti-LAG-3 blocking antibodies had no impact on human NK cells [129], suggesting a species-specific role. Regardless, LAG-3 in NK cells requires further investigation before being considered as a candidate for immune checkpoint therapy.
LAIR1 binds to all collagens and is thought to aid in NK cell discrimination of damaged tissue, where the collagen-rich extracellular matrix of cells has been reduced [130]. Tumor cells have been observed to up-regulate localized collagen expression as a means of forming adhesive structures in the TME. Coincidentally, this also hides tumor cells from LAIR-1-expressing immune cells such as NK cells, with LAIR-1 inhibitory signals correlating with dampened in NK cytotoxicity [85].
Siglec-7 and 9 receptors recognize sialylated glycans [131,132], which are found on glycoproteins and glycolipids on the outer membrane of mammalian cells and are believed to act as a signal of self [133]. An abnormally high sialic acid coat (hypersialylation) is a hallmark of many tumors and may facilitate evasion of Siglec-expressing NK cells [134,135]. Consistent with this, several in vitro studies using blocking antibodies or chemical de-sialylation of proteins on the surface of tumor cells have demonstrated enhanced NK-mediated anti-tumor responses [136,137,138]. Although a murine orthologue for Siglec-9 exists (Siglec-E), it is not expressed on NK cells, so there is no role for Siglec 9 NK responses in mice to date.
The CEACAM family is composed of over 10 members that have multiple roles in differentiation, proliferation and signaling. CEACAM1 is the only family member expressed on NK cells and has approximately 12 isoforms with varying extracellular and intracellular domains. The CEACAM1-L isoform predominates and contains three to four extracellular Ig-like domains and two cytoplasmic ITIMs in humans, or one ITIM and one ITSM in rodents [143]. NK CEACAM1 binds other CEACAM molecules present on nearby cells, including tumor cells; this interaction inhibits NKG2D-mediated tumor control [84,149].
The C-type lectin receptor superfamily includes inhibitory members KLRG1 on both human and mice, NKRP1A in humans, and NKRP1B and gp49B1 in mice. KLRG1 is mostly known for its role in NK cell maturation, development and maintenance of peripheral homeostasis [150]. Recently, a new role for KLRG1 has emerged as an inhibitory receptor in the context of tumor control. The KLRG1 ligands E- and N-cadherin were found to be upregulated in tumor samples from patients suffering from melanoma, prostate, breast or colorectal cancer [86]. Furthermore, blocking KLGR1 in mice showed enhanced tumor control compared to controls, with even better protection observed when used in combination with anti-PD1 [86]. Little research has been performed to understand the inhibitory role of KLRG1 in NK cells and this latter study highlights the therapeutic potential of anti-NKLRG1 antibodies.
The TAMs are a relatively new family of NK inhibitory receptors [88,151,152]. TAMs contain an extracellular region composed of two Ig-like domains followed by two fibronectin type II domains, a transmembrane region and an intracellular region containing a tyrosine kinase domain, an ITIM and ITIM-like motif. TAMs are classified as receptor tyrosine kinases (RTKs) and bind via their extracellular domain to growth arrest-specific gene 6 (Gas6) and Protein S (Pros1). TAM inhibition in the mouse B16F10 melanoma model has shown promising results and mechanistically, TAM receptors are proposed to act via inhibition of NKG2D signaling [88]. TAM association with Casitas B lineage lymphoma b (Cbl-b) has recently uncovered a new inhibitory signaling pathway in NK cells [88,153] (discussed in more detail in a later section; Figure 2A).
3. Current Therapies Harnessing the Power of Inhibitory NK Receptors
The success of current immunotherapies that block immune checkpoints, such as CTLA4 and PD1 in CD8 cells, has spurred interest in targeting other immune populations, such as NK cells. Given that blocking NK inhibitory receptors has proven beneficial in experimental tumor models, a number of blocking antibodies have been developed, with several progressing to clinical trials. Currently, KIR2D and NKG2A are the only two MHC-I-dependent receptors targeted to mimic a “missing-self” situation and enhance anti-tumor activity, with two agents that block KIR2D (Lirilumab and IPH2101) in clinical phase I or II trials. Lirilumab alone or in combination with other agents is being trialed against solid tumors (NCT03203876), hematological malignancies [154], chronic lymphocytic leukemia (NCT02557516), multiple myeloma (NCT01592370), resectable squamous cell carcinoma of the head and neck (NCT03341936), and resectable bladder cancer (NCT03532451), while IPH2101 is being trialed against acute myeloid leukemia [155]. Monalizumab is the only antibody being trialed for NKG2A, both alone and in combination with other agents for advanced gynecological solid tumors [156], advanced squamous cell carcinoma of the head and neck (NCT02643550), and resectable non-small cell lung cancer (NCT03794544).
In addition to KIR2D and NKG2A, anti-TIGIT antibodies are in multiple phase I and II clinical trials against different advanced solid tumors (NCT029133133, NCT02794571 and NCT02964013). Anti-TIGIT antibodies are also being used in combination with other therapies against solid tumors (NCT04150965, NCT03119428 and NCT04047862), notably against advanced non-small cell lung cancer (NCT03563716). These are just a handful of the clinical trials that are taking advantage of blocking inhibitory NK signaling, with no doubt many more to come.
4. Inhibitory NK Signaling
The full complement of events that transduce inhibitory receptor signals remains unclear, with much work still required to fully understand the different pathways. However, for some receptors a clearer picture of the signaling events following ligand engagement is now emerging, as well as identification of alternative signaling cascades that can lead to inhibition.
Inhibitory NK receptors signal via the different tyrosine-containing motifs: ITIM, ITT, ITSM, ITIM-like, ITT-like, or ITSM-like motifs. Upon engagement of the respective ligands, tyrosines within the signaling motifs are phosphorylated by members of the Src-family kinases (SFKs), with Lyn and Lck the most likely candidates in NK cells (Figure 3A) [157,158,159]. Tyrosine phosphorylation within the inhibitory motif enables recruitment of Src homology 2 (SH2)-containing protein tyrosine phosphatases such as SHP-1 and SHP-2, which then function to de-phosphorylate signaling intermediates and negatively regulate NK cell activity.
4.1. SHP1 and SHP-2 Signaling
SHP-1 is required for inhibition of NK cell effector function, whereas SHP-2, despite binding to most inhibitory receptors, appears to selectively inhibit cytokine production [160,161]. Most inhibitory NK receptor signaling tails are comprised of at least of two ITIMs or are found as homodimers creating two parallel ITIM sites within one receptor complex. Recruitment of SHP-1 and SHP-2 is dependent on both the residues flanking the tyrosine and the spacing between phosphorylated tyrosines. So far, the only known NK cell substrate de-phosphorylated by SHP-1 is the guanine nucleotide exchange factor Vav1. However, in other cell types SHP-1 has been shown to interact with CD3ζ, Syk, ZAP-70, LAT and SLP-76, and these may also prove to be targets in NK cells [159,162,163,164]. Phosphorylated Vav1 (pVav1) is a major intermediate downstream of multiple activating signals and is crucial for MTOC organization, thus dephosphorylation of pVav1 at the IS essentially attenuates NK activation. Signaling intermediates downstream of Src-kinase activity and SHP-1 and -2 recruitment remain to be elucidated in most KIRs, most Ly49s, CD94:NKG2A and B, NKRP1B, and CEACAM1 [165,166,167], whereas others such as KIR2DL1,3–4, Ly49C and gp49B1 bind to β-arrestin 2, enhancing recruitment of SHP-1 and SHP-2 [168]. For some receptors, recruitment of SHP-1 and SHP-2 is species dependent; take for instance human LAIR-1 associating with SHP-1 and SHP-2 and murine LAIR-1 only interacting with SHP-2 [145,169,170]. Regardless, the Src-kinase inhibitor PPI is shown to inhibit LAIR-1 tyrosine phosphorylation despite the Src-kinase responsible remaining unknown. Although LILRB1 contains four ITIM motifs, a study by Bellón et al. [171] showed that only Y614 and Y644 are required for inhibitory signals via SHP-1 association; Siglec-7 and -9 have also been shown to associate with SHP-1 and SHP-2 [172,173]. Inhibitory signaling by Siglec 7 is regulated by Suppressor of Cytokine Signaling (SOCS)3, which binds to the ITIM, blocking docking of SHP-1 and SHP-2, and potentially targeting Siglec-7 for proteasomal degradation via its associated E3 ubiquitin ligase complex [174]. In contrast, TIGIT contains an ITT-like motif that allows association with Grb2, which in turn recruits SHIP-1 (SH-2 containing inositol 5’ polyphosphatase 1) [175]. SHIP-1 is known to inhibit PI3K signaling by hydrolyzing PI(3,4,5)P3, inhibiting the accumulation and activation of Akt, Btk and PLC-γ (Figure 2A).
4.2. c-Abl Signaling
In both human and murine NK cells, a SHP-1/2-independent inhibitory mechanism has also been described, involving phosphorylation of a small adaptor protein named Crk by the c-Abl tyrosine kinase [176,177]. Active Crk is non-phosphorylated, is found in many different complexes together with c-Cbl, C3G and p130CAS, and is known to confer activating signals. Following engagement of inhibitory receptors and phosphorylation of the signaling motifs, the c-Abl kinase phosphorylates Crk, rendering it inactive and resulting in its dissociation [178] from activating complexes [179]. This inhibitory pathway has been shown to be KIR- and CD94:NKG2A-mediated.
4.3. TAM Signaling
A novel TAM/Cbl-b-mediated inhibitory signaling pathway has also been described. TAMs exert their inhibitory function by phosphorylation of human Cbl-b on tyrosine residues 133 and 363 [153]. Phosphorylation of Y363 prompts a conformational change, activating Cbl-b E3 ubiquitin ligase activity [180]. Cbl-b is thought to then target LAT1, a key molecule downstream of activating receptors NKG2D and NK1.1, for ubiquitination and subsequent proteasomal degradation, resulting in inhibition of NK cell activity [153].
5. Activating NK Cell Receptors
NK cells express activating receptors on their surface which recognize different stress molecules and ligands on ‘unhealthy’ cells (Figure 2B). Unlike the inhibitory receptors, most activating receptors lack a cytoplasmic signaling tail and instead associate with membrane bound adaptor proteins which contain immunoreceptor tyrosine-based activation motifs (ITAMs), to propagate their signals [181]. To date, CD16, NKG2D, NKp46 (natural cytotoxicity receptor 1; NCR1), DNAM-1, 2B4, NTB-A and CRACC have been found to be critical for tumor surveillance in both human and murine NK cells [182]. Human NK cells additionally express and rely on NKp30 (NCR3) and NKp44 (NCR2) for tumor surveillance (Figure 2 and Table 4) [183].
CD16 (FcgRIII) is a low-affinity receptor for IgG and is expressed at high levels on human NK cells and at lower densities on murine NK cells [194,195,196]. NK cells express a transmembrane form of CD16 that lacks a cytoplasmic signaling tail [197] and, in humans, associates with either CD3ζ homodimers [194,198], FCRγ homodimers [199] or CD3ζ:FCRγ heterodimers to transduce activating signals, while murine CD16 associates with FCRγ homodimers [200]. CD16 expression enables NK cells and other immune cells to recognize the Fc portion of an antibody bound to an antigen on the surface of tumor cells and trigger NK-mediated lysis of the target cell (known as ADCC). After cytolysis, CD16 is proteolytically cleaved by ADAM17 [201] or MMP25 [202] and shed from the NK cell surface, with shedding not only important for NK cell detachment and subsequent targeting of tumor cells but also enhancement of subsequent NK cell signaling [203]. This has resulted in a new field focused on generating monoclonal antibodies against tumor antigens to enhance immune-mediated ADCC [204]. Additionally, endogenous antibodies to tumor antigens have been observed in patients with various cancers, including human papillary thyroid cancer, some soft tissue sarcomas and melanoma [205,206]. Although these antibodies are predominantly IgG and are located within neoplastic tissue in the TME [207], they apparently fail to elicit tumor control; why this is the case remains unknown.
NKG2D is a member of the NKG2 receptor family and is one of the main activating receptors involved in NK cell tumor surveillance [208]. In humans, its ligands include MICA and B and UL16-binding proteins (ULBP)1–6, and in mice, Rae1α-ε, MULT1 and H60a-c (with H60a-c restricted to BALB/C mice) [188,189,190]. Structurally, NKG2D ligands resemble MHC-I proteins, with MICA and B containing three extracellular α(1–3) domains in which α3 is an Ig-like domain. ULBP1-6 have two extracellular α(1–2) domains and ULBP4 and 5 are similar to murine Rae1 ligands [191,209]. ULBP1-3 and -6 are attached to the membrane via a glycophosphatidylinositol (GPI)-anchor, whereas MICA and B and ULBP4–5 all have a transmembrane domain and cytoplasmic tail.
NKG2D has two isoforms that dictate NKG2D signaling: the short isoform NKG2D-S associates with the DNAX-activating protein (DAP)10 and 12 adaptor proteins, while the long isoform (NKG2D-L) only associates with DAP10 [210]. Homodimeric NKG2D forms a hexamer with each subunit associated with either a DAP10 or DAP12 homodimer, respectively [211]. Additionally, NKG2D-S is restricted to mice, meaning that human NKG2D only signals via DAP10, while engagement of the murine NKG2D receptor can lead to signaling by either DAP10 or DAP12 [212,213]. The differences between mouse and human are further amplified by the presence of an ITAM signaling motif in DAP12 and a YxxM motif in DAP10 (Figure 2B and Table 5) [214,215].
Human NKG2D ligands are expressed in ovarian cancer, leukemia, colorectal cancer and pediatric brain cancers, amongst others [216,217,218,219,220]. Regardless of ligand expression, Groh et al. [221] showed that soluble MIC (sMIC) shed by tumors impaired NKG2D mediated cytotoxicity by blocking receptor engagement and thus signaling [221,222]. This shedding of NKG2D ligands has been observed in many cancers, including ovarian, breast, lung, colon and prostate, as well as lymphoma, myeloma and melanoma [219,220,223,224]. Antibodies targeting sMIC have been shown to enhance NK and T cell-mediated killing of head and neck squamous cell carcinoma [225], in addition to other patient cancers, when combined with PD1/PD-L1 blockade [226].
NKp46, NKp30 and NKp44 (NCR family) were the first activating receptors identified in NK cells [227,228,229]. While many molecules such as heparin, vimentin and viral proteins are suspected ligands, the search for cellular ligands remains a priority in the field [183]. So far, two cellular ligands, B7 homologue 6 (B7H6) and NKp44L, have been identified for NKp30 and NK44, respectively [184,185,186,187]. There is currently no known ligand for NKp46, which is the only NCR conserved in both human and mice and is found on activated and resting NK cells. Despite not knowing which ligands activate NKp46, its engagement has been shown to be critical for control of melanoma and lung carcinoma metastasis in experimental models [230,231]. NKp46 contains two extracellular Ig-like domains, a transmembrane domain and a short, 25-residue cytoplasmic tail [232]. Following NKp46 engagement, signaling occurs through NKp46 interaction with a CD3ζFCRγ heterodimer [229,233] and results in NK cytoskeletal engagement and F-actin accumulation at the lytic synapse, both early events in the killing process (Figure 1) [234]. Tantalizingly, this hints that NKp46 has a critical role in target identification, but without a known ligand or ligands, it is impossible to confirm. In addition, studies have shown that NKp46 engagement results in IFNγ and TNFα production [231,235].
NKp30 is found on mature resting and activated NK cells, with most NK cells expressing one or two of the three major NKp30 isoforms (NKp30A-C), that differ in the composition of their cytoplasmic tails [228,236]. Depending on the isoform engaged, NKp30 transduces different signals via CD3ζ homodimers or CD3ζFCRγ heterodimers. For example, engaging NKp30A or B induces significantly higher IFNγ, TNFα and IL12B production, while NKp30C which induces IL10 production, leads to immune suppression [32,237]. In addition, NKp30B constitutively associates with CD3ζ, while both NKp30A and C associate with CD3ζ upon engagement; however, NKp30A has a tighter association with CD3ζ and gives the dominant response [32]. Furthermore, the predominant expression of NKp30C in patients with GIST is predictive of a poor prognosis [32,237].
NKp44 is only expressed upon activation and associates with ITAM-containing DAP12 to induce IFNγ and TNFα production [229,238,239], and cell lysis of experimental cervix carcinoma and neuroblastoma cell lines [240]. Finally, although NKp44 contains a cytoplasmic ITIM-like motif, this does not appear to be functional [239,241].
DNAM-1 has an important role in NK cell-mediated tumor immunosurveillance and shares CD155 and CD112 ligands with TIGIT and TACTILE (CD96) [242,243]. The role of DNAM-1 in immunosurveillance was discovered using the RMA lymphoma model where NK cells were shown to be responsible, at least in part, for metastatic control of DNAM-1 ligand-expressing RMA tumors [244]. The importance of DNAM-1 is underscored by DNAM-1-deficient NK cells that no longer control experimental metastasis in a spontaneous fibrosarcoma formation model [245,246]. However, if the tumors express NKG2D ligands, then DNAM-1 deficiency has no impact, suggesting a signal hierarchy between the two receptors [247,248]. DNAM-1 ligand CD155 is expressed in a variety of human cancers, including colon, adenocarcinoma, pancreatic and melanoma [249,250,251,252,253]. Interestingly, human melanoma samples have also been shown to express CD112 [254], and expression of CD112 and CD155 on patient-derived neuroblastomas correlated with susceptibility to NK cell killing in vitro [255]. To date, CD155 but not CD112 has been shown to be critical for DNAM-1-mediated NK cell cytotoxicity [255,256]. Furthermore, CD155 association with DNAM-1 promotes NK cell cytotoxicity and IFNγ production [257].
2B4 (SLAMF4), NTB-A (SLAMF6 or Ly108 in mouse) and CRACC (SLAMF7) belong to the signaling lymphocytic activation molecule (SLAM) family and are the only members expressed on NK cells [258,259]. CRACC and NTB-A act in trans as their own ligands, while CD48 (SLAMF2) is the ligand for 2B4 and is thought to act in trans and in cis [192,193,260]. CRACC and 2B4 are potent stimulators of NK cell cytotoxicity; CRACC is already in clinical use and 2B4 is a potential new therapeutic target [261]. The SLAMs contain cytoplasmic ITSM motifs that recruit different signaling molecules to allow for a switch between activating and inhibitory signals following receptor engagement [262,263].
6. Current Therapies Harnessing the Power of Activating NK Receptors
There are several ongoing clinical trials testing antibodies that enhance NK cell activation, mediate direct cell killing (ADCC) or achieve both NK cell activation and ADCC. The latter is exemplified by Elozutumab, an anti-CRACC (SLAM7) antibody currently in pre-clinical testing and phase 1–3 clinical trials for multiple myeloma (NCT01335399) [264,265,266]. Another ongoing trial in non-Hodgkin’s lymphoma is combining anti-CD123 antibody with adoptive transfer of an NK cell line engineered to express high levels of CD16 and potentiate NK responses (NCT03027128) [267]. Adoptively transferred, allogeneic CD19 CAR-NK cells were successfully used in recent phase 1 and 2 trials to treat patients with non-Hodgkin’s lymphoma or chronic lymphocytic leukemia (CLL) without significant toxicities [35]. These studies demonstrate the importance of NK cell therapies and pave the way for further clinical trials using blocking antibodies and/or CAR-NK cells expressing activating receptors [268,269,270].
7. Activating NK Signaling
7.1. ITAM Signaling
Following CD16, NKG2D and NCR family receptor engagement adaptor proteins, DAP12, CD3ζ and FCRγ are rapidly phosphorylated within their ITAM sequences by an as yet unidentified Src-kinase, which leads to adaptor association with Syk or Zap70 tyrosine kinases (Figure 3B) [215,271,272]. Following recruitment to DAP12, Syk is thought to interact with the p58 subunit of PI3K leading to a PI3K → Rac1 → PAK1 → MEK → ERK signaling cascade that drives NK cell cytotoxicity (Figure 3B) [272,273]. Although Zap70 has also been shown to associate with the ITAMs it does not appear to be required for signaling.
CD16 signals through its CD3ζ or FCRγ adaptors and like DAP12, activates PI3K, however, other signaling molecules such as Vav1, PLC-γ1 and PLC-γ2 can also be activated following CD16 engagement [274,275]. Additionally, CD16 engagement has been linked to PIP2 production mediated by PI5K [276], with Galandrini et al. [277] showing that PI5K was required for NK cell degranulation but not granule polarization in primary human NK cells. The combined activation of the PI3K and PI5K pathways could explain why CD16 is the only receptor that can fully activate resting human NK cells [278]. In addition to the ITAM-mediated signaling cascades, NK cells have been shown to signal through transmembrane-bound LAT complexed with PLC-γ1/2; the signaling intermediates remain to be elucidated [279].
7.2. DAP10 (YxxM) Signaling
DAP10 is a small transmembrane adaptor protein containing a ’traditional’ costimulatory PI3K binding motif (YxNM) and a binding site for Grb2 (pYxNx) [280]. Following receptor engagement, the DAP10 motif is phosphorylated by an unknown Src-kinase to recruit a Grb2-Vav1 complex and the p85 subunit of PI3K [281,282]. Phosphorylation of Grb2-Vav1 leads to phosphorylation of Vav1, PLC-γ2 and SLP-76 [281,283]. Presumably, PI3K activation via DAP10 converges on AKT with the end result being an increase in direct cytotoxicity [280,284]. Interestingly, Grb2-Vav1 signaling alone is not sufficient to stimulate full calcium release and cytotoxicity [282], whilst NKG2D:DAP10 activation of Vav1 is important for induction of actin polymerization and polarization of MTOC at the IS [285].
7.3. DNAM-1, 2B4, CRACC and NTB-A Signaling
DNAM-1, 2B4, CRACC and NTB-A contain a cytoplasmic signaling tail, distinguishing them from the NCRs, CD16 and NKG2D.
DNAM-1 has an ITT-like motif that is phosphorylated at Y319 in mouse and Y322 in humans [286] and is required for association with Grb2 and initiation of the PI3K signaling cascade (Grb2→ Vav1 → PI3K → PLC-γ1) (Figure 3B) [102], although further signaling intermediates have not been fully elucidated. Interestingly, DNAM-1 signaling enhances Vav1-mediated actin polymerization and polarization of the lytic granules to the IS, consistent with its role in NKG2D:Dap10 signaling [102,285].
2B4, CRACC and NTB-A: 2B4 contains four ITSM motifs, while CRACC and NTB-A contain two ITSM motifs [287]. Following 2B4 engagement, ITSM tyrosines are phosphorylated recruiting either SAP, EAT2 or 3BP2. Six possible signaling cascades have been elucidated to date: (1) SAP → Fyn → pVAV1 [288,289], (2) EAT → PLC-γ → Ca2+ flux, (3) EAT → pERK, [290], (4) 3BP2 → pVAV1, (5) 3BP2 → PLC-γ and (6) 3BP2 → pERK [291,292,293]. Notably, Saborit-Villarroya et al. [292] showed that cytotoxicity, not cytokine release, is regulated by pVAV1 and ERK, thus cytokine release and cytotoxicity are regulated by different signaling pathways downstream of 2B4.
Similar to 2B4, engagement of NTB-A results in recruitment of SAP and EAT2, and NK cell cytotoxicity and cytokine production [258,294,295]. In contrast, CRACC does not recruit SAP and solely relies on EAT2 for signaling [296,297]. As mentioned, ITSM phosphorylation can transduce either activating or inhibitory signals (Figure 2A). Activation relies on SAP recruitment, which blocks binding site of the phosphatases SHP-1, SHP-2 and SHIP [288,298,299]. Patients suffering from X-linked lymphoproliferative disease (XLP), which is caused by mutations in the gene encoding SAP, show defective NK cell activation and even inhibition [300,301].
8. Releasing the Brake
While the previous sections discuss the inhibitory and activating receptors and their signaling events separately, in reality, inhibitory and activating signaling occurs simultaneously upon each NK cell-to-cell encounter. As mentioned, NK cells are loaded with lytic granules and are ready to kill once a “decision” has been made. It is this ability to release lytic content and kill their targets within 30 minutes of engagement, that makes them such a powerful weapon for the immune system [302,303,304]. These natural killers are constantly surveilling our bodies and more often than not, encounter healthy cells that engage the myriad of inhibitory receptors expressed on NK cells. NK cells can thus be considered in a constant state of inhibition and overcoming that inhibitory threshold requires either “release” of the inhibitory “brake” or a countermanding activating signal such as via CD16, or from co-engagement of various activating receptors [305]. The inhibitory signals appear to act by blocking early activation, for example, NKG2D, DNAM-1, 2B4, NTB-A and CRACC all converge at Vav-1 dephosphorylation (Figure 3A), which would prevent MTOC formation and result in tolerance. This implies that Vav phosphorylation may be a “master” switch and understanding the regulatory events surrounding it could point to new interventions which release the brake on NK cell killing.
9. Conclusion and Future Directions
While numerous inhibitory and activating receptors have been identified, the NK signaling field is still in its infancy. For clarity, this review has focused on the most well-studied aspects and we apologize if we have omitted publications that have contributed to a rapidly moving field.
NK cells play a pivotal role in controlling tumor metastasis and there is enormous potential for the development of new cancer immunotherapies that enhance NK cell activity and infiltration into the TME. If we consider that NK cells are constantly surveying the body for infected or transformed “unhealthy” cells, then essentially, most of the time there should be no NK response and thus constant NK inhibitory signaling in the “normal” state.
In this context, the trigger for response is activation and raises an over-arching question: how does activation signaling switch off or dominate baseline inhibitory signaling? There are still many gaps in our understanding of these activating and inhibitory signaling pathways. For instance, what are the initiating signals post-ligand–receptor engagement? What is the role of the Src-family kinases and how are they activated? Similarly, once the initial signals are transduced, what intermediates and regulatory mechanisms come into play and how does this converge to give the final killing blow? Further delineation of the signals that regulate NK cell responses will be critical not only in the identification of new targets but to understand the full impact of any intervening strategies.
To evade NK cell recognition and killing, tumor cells upregulate inhibitory ligands and signaling, as well as downregulate activating ligands; blocking inhibitory and/or enhancing activating ligands offers attractive opportunities to improve anti-tumor responses. In contrast to T-cells, activated or adoptively transferred NK cells have the added advantage of not inducing cytokine release syndrome or neurotoxicity, and allogeneic NK cells/cell lines have at least proven safe for adoptive immunotherapy [35,306,307]. Further safety and efficacy testing of NK cell therapies, such as adoptive transfer of NK cell lines, CAR-NK cells, checkpoint blockades and ADCC, will no doubt further advance the field of NK therapeutics.
Many studies have shown that NK cell–mediated tumor control results not only from NK cell cytotoxicity but also NK release of cytokines into the TME to marshal other immune cells into a full-scale attack. Therapeutic approaches that not only trigger cytotoxicity, but aid exhaustion recovery and enhance proliferation and directed cytokine production, will be key.
Author Contributions
L.G.M.G. collected the literature and wrote the manuscript. N.K. designed and created all figures for this manuscript. S.E.N. proofread the manuscript, gave suggestions, and modified the language. All authors have read and agreed to the published version of the manuscript.
Funding
This review was made possible in part through a grant from the National Health and Medical Research Council of Australia (1124784) and through Victorian State Government Operational Infrastructure Support and the Australian Government National Health and Medical Research Council Independent Research Institute Infrastructure Support scheme.
Conflicts of Interest
S.N. receives research funding from and has licensed a patent to a pharmaceutical partner with interest in NK cells as therapeutic targets. The funders had no role in the content of the review or the writing of the manuscript.
Abbreviations
Antibody dependent cellular cytotoxicity (ADCC); Programmed cell death protein 1 (PD1); Programmed death-ligand 1 (PD-L1); B7 homologue 6 (B7H6); Bruton’s tyrosine kinase (Btk); Abelson tyrosine kinase (c-Abl); c-Casitas B lineage lymphoma (c-Cbl); Chimeric antigen receptor (CAR); Casitas B lineage lymphoma b (Cbl-b); Carcinoembryonic Ag cell adhesion molecule 1 (CEACAM1); Carcinoembryonic Ag cell adhesion molecule 1 Ligand (CEACAM1-L); Chronic lymphocytic leukemia (CLL); CD2-like receptor-activating cytotoxic cell (CRACC); Cytotoxic T-lymphocyte-associated protein 4 (CTLA4); C-type lectin domain (CTLD); DNAX-activating Protein 10 (DAP10); DNAX-activating Protein 12 (DAP12); DNAX accessory molecule-1 (DNAM-1); Extracellular signal-regulated kinases (ERK); Filamentous actin (F-actin); Low affinity immunoglobulin gamma Fc region receptor III-A (FcgRIII); Growth arrest-specific gene 6 (Gas6); Gastrointestinal sarcoma (GIST); Growth factor receptor-bound protein 2 (Grb2); Human leukocyte antigens (HLA); Interferon gamma (IFNγ); Immunoglobulin (Ig); Inhibitory KIRs (iKIRs); Interleukin-12 subunit beta (IL12B); Inhibitory Ly49 receptors (iLy49s); Inhibitory NKG2 (iNKG2); Immunological synapse (IS); Immunoreceptor tyrosine-based activation motif (ITAM); Immunoreceptor tyrosine-based inhibtory motif (ITIM); Immunoreceptor tyrosine-based switch motif (ITSM); Immunoglobulin tail tyrosine motif (ITT); Killer cell immunoglobulin-like receptor (KIR); Killer cell lectin-like receptor subfamily A (klra); Killer cell lectin-like receptor G1 (KLRG1); Soluble leukocyte-associated Ig-like receptor-1 (LAIR-1); L-type amino acid transporter 1 (LAT1); lymphocyte-specific protein tyrosine kinase (Lck); Leukocyte immunoglobulin-like receptor (LILR); Lymphocyte Ag (Ly(X); Mitogen-activated protein kinase (MAPK); Major histocompatability complex (MHC); MHC class I-related chains (MIC); Matrix metalloproteinase 25 (MMP25); Microtubule-organizing center (MTOC); Murine UL16 binding protein (MULT); Natural cytotoxicity receptor (NCR); Natural killer (NK); Killer cell lectin-like receptor subfamily B, member 1 (NK1.1); Non-obese diabetic (NOD); NK-T-B-Antigen (NTB-A); Phosphatidylinositol 3-kinase (PI3K); Phosphatidylinositol 4,5-bisphosphate or PtdIns(4,5)P2 (PIP2); Phospholipase C (PLC-y); Proton-pump inhibitors (PPI); Protein S (Pros1); Phosphorylated Vav1 (pVav1); Poliovirus receptor (PVR); Poliovirus Receptor-Related 2 (PVRL2); Ras-related C3 botulinum toxin substrate 1 (Rac1); Retinoic Acid Early Transcript 1E (RAET1E); Receptor tyrosine kinases (RTKs); SLAM-associated protein (SAP); Src-family kinases (SFKs); Src homology 2 (SH2); SH2-containing inositol 5-phosphatase (SHIP); SH2 domain- containing protein tyrosine phosphatase-1 (SHP-1); SH2 domain- containing protein tyrosine phosphatase-2 (SHP-2); Sialic acid-binding immunoglobulin-like lectin (Siglec); Signaling lymphocytic activation molecule (SLAM); Lymphocyte cytosolic protein 2 (SLP-76); Soluble MIC (sMIC); Suppressor of Cytokine Signaling (SOCS); Proto-oncogene tyrosine-protein kinase Src (Src); Spleen tyrosine kinase (Syk); Tyro3, Axl and MerTK (TAM); T cell immunoglobulin and ITIM domain (TIGIT); Tumor microenvironment (TME); Tumor necrosis factor alpha (TNFα); Tumor necrosis factor-related apoptosis-inducing ligand (TRAIL); UL16-binding proteins (ULBP); X-linked lymphoproliferative disease (XLP); Zeta Chain Of T Cell Receptor Associated Protein Kinase 70 (ZAP-70).
References
- Gregoire, C.; Chasson, L.; Luci, C.; Tomasello, E.; Geissmann, F.; Vivier, E.; Walzer, T. The trafficking of natural killer cells. Immunol. Rev. 2007, 220, 169–182. [Google Scholar] [CrossRef] [Green Version]
- Ortaldo, J.R.; Wiltrout, R.H.; Reynolds, C.W. Natural killer activity: Early days, advances, and seminal observations. Crit. Rev. Oncog. 2014, 19, 1–13. [Google Scholar] [CrossRef]
- Rosenberg, E.B.; Herberman, R.B.; Levine, P.H.; Halterman, R.H.; McCoy, J.L.; Wunderlich, J.R. Lymphocyte cytotoxicity reactions to leukemia-associated antigens in identical twins. Int. J. Cancer 1972, 9, 648–658. [Google Scholar] [CrossRef]
- Moretta, A.; Bottino, C.; Vitale, M.; Pende, D.; Cantoni, C.; Mingari, M.C.; Biassoni, R.; Moretta, L. Activating receptors and coreceptors involved in human natural killer cell-mediated cytolysis. Annu. Rev. Immunol. 2001, 19, 197–223. [Google Scholar] [CrossRef]
- Greenberg, A.H.; Hudson, L.; Shen, L.; Roitt, I.M. Antibody-dependent cell-mediated cytotoxicity due to a “null” lymphoid cell. Nat. New Biol. 1973, 242, 111–113. [Google Scholar] [CrossRef]
- Herberman, R.B.; Nunn, M.E.; Holden, H.T.; Lavrin, D.H. Natural cytotoxic reactivity of mouse lymphoid cells against syngeneic and allogeneic tumors. II. Characterization of effector cells. Int. J. Cancer 1975, 16, 230–239. [Google Scholar] [CrossRef]
- Borrego, F. The first molecular basis of the “missing self” hypothesis. J. Immunol. 2006, 177, 5759–5760. [Google Scholar] [CrossRef] [Green Version]
- Ljunggren, H.G.; Karre, K. In search of the ‘missing self’: MHC molecules and NK cell recognition. Immunol. Today 1990, 11, 237–244. [Google Scholar] [CrossRef]
- Storkus, W.J.; Alexander, J.; Payne, J.A.; Dawson, J.R.; Cresswell, P. Reversal of natural killing susceptibility in target cells expressing transfected class I HLA genes. Proc. Natl. Acad. Sci. USA 1989, 86, 2361–2364. [Google Scholar] [CrossRef] [Green Version]
- Storkus, W.J.; Alexander, J.; Payne, J.A.; Cresswell, P.; Dawson, J.R. The alpha 1/alpha 2 domains of class I HLA molecules confer resistance to natural killing. J. Immunol. 1989, 143, 3853–3857. [Google Scholar]
- Karlhofer, F.M.; Ribaudo, R.K.; Yokoyama, W.M. MHC Class I alloantigen specificity of Ly-49+ IL-2-activated natural killer cells. Nature 358: 66-70, 1992. J. Immunol. 2006, 177, 5761–5765. [Google Scholar]
- Yokoyama, W.M. The search for the missing ‘missing-self’ receptor on natural killer cells. Scand. J. Immunol. 2002, 55, 233–237. [Google Scholar] [CrossRef]
- Moretta, A.; Vitale, M.; Bottino, C.; Orengo, A.M.; Morelli, L.; Augugliaro, R.; Barbaresi, M.; Ciccone, E.; Moretta, L. P58 molecules as putative receptors for major histocompatibility complex (MHC) class I molecules in human natural killer (NK) cells. Anti-p58 antibodies reconstitute lysis of MHC class I-protected cells in NK clones displaying different specificities. J. Exp. Med. 1993, 178, 597–604. [Google Scholar] [CrossRef] [Green Version]
- Boudreau, J.E.; Hsu, K.C. Natural Killer Cell Education and the Response to Infection and Cancer Therapy: Stay Tuned. Trends Immunol. 2018, 39, 222–239. [Google Scholar] [CrossRef]
- Fauriat, C.; Long, E.O.; Ljunggren, H.G.; Bryceson, Y.T. Regulation of human NK-cell cytokine and chemokine production by target cell recognition. Blood 2010, 115, 2167–2176. [Google Scholar] [CrossRef] [Green Version]
- Takeda, K.; Hayakawa, Y.; Smyth, M.J.; Kayagaki, N.; Yamaguchi, N.; Kakuta, S.; Iwakura, Y.; Yagita, H.; Okumura, K. Involvement of tumor necrosis factor-related apoptosis-inducing ligand in surveillance of tumor metastasis by liver natural killer cells. Nat. Med. 2001, 7, 94–100. [Google Scholar] [CrossRef]
- Backes, C.S.; Friedmann, K.S.; Mang, S.; Knörck, A.; Hoth, M.; Kummerow, C. Natural killer cells induce distinct modes of cancer cell death: Discrimination, quantification, and modulation of apoptosis, necrosis, and mixed forms. J. Biol. Chem. 2018, 293, 16348–16363. [Google Scholar] [CrossRef] [Green Version]
- Bryceson, Y.T.; March, M.E.; Barber, D.F.; Ljunggren, H.G.; Long, E.O. Cytolytic granule polarization and degranulation controlled by different receptors in resting NK cells. J. Exp. Med. 2005, 202, 1001–1012. [Google Scholar] [CrossRef]
- Bryceson, Y.T.; Ljunggren, H.G.; Long, E.O. Minimal requirement for induction of natural cytotoxicity and intersection of activation signals by inhibitory receptors. Blood 2009, 114, 2657–2666. [Google Scholar] [CrossRef] [Green Version]
- Vyas, Y.M.; Mehta, K.M.; Morgan, M.; Maniar, H.; Butros, L.; Jung, S.; Burkhardt, J.K.; Dupont, B. Spatial organization of signal transduction molecules in the NK cell immune synapses during MHC class I-regulated noncytolytic and cytolytic interactions. J. Immunol. 2001, 167, 4358–4367. [Google Scholar] [CrossRef]
- Orange, J.S.; Harris, K.E.; Andzelm, M.M.; Valter, M.M.; Geha, R.S.; Strominger, J.L. The mature activating natural killer cell immunologic synapse is formed in distinct stages. Proc. Natl. Acad. Sci. USA 2003, 100, 14151–14156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, H.S.; Das, A.; Gross, C.C.; Bryceson, Y.T.; Long, E.O. Synergistic signals for natural cytotoxicity are required to overcome inhibition by c-Cbl ubiquitin ligase. Immunity 2010, 32, 175–186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Das, A.; Long, E.O. Lytic granule polarization, rather than degranulation, is the preferred target of inhibitory receptors in NK cells. J. Immunol. 2010, 185, 4698–4704. [Google Scholar] [CrossRef] [Green Version]
- Treanor, B.; Lanigan, P.M.; Kumar, S.; Dunsby, C.; Munro, I.; Auksorius, E.; Culley, F.J.; Purbhoo, M.A.; Phillips, D.; Neil, M.A.; et al. Microclusters of inhibitory killer immunoglobulin-like receptor signaling at natural killer cell immunological synapses. J. Cell Biol. 2006, 174, 153–161. [Google Scholar] [CrossRef] [PubMed]
- Bhat, R.; Watzl, C. Serial killing of tumor cells by human natural killer cells--enhancement by therapeutic antibodies. PLoS ONE 2007, 2, 326. [Google Scholar] [CrossRef] [PubMed]
- Sathe, P.; Delconte, R.B.; Souza-Fonseca-Guimaraes, F.; Seillet, C.; Chopin, M.; Vandenberg, C.J.; Rankin, L.C.; Mielke, L.A.; Vikstrom, I.; Kolesnik, T.B.; et al. Innate immunodeficiency following genetic ablation of Mcl1 in natural killer cells. Nat. Commun. 2014, 5, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Lopez-Soto, A.; Gonzalez, S.; Smyth, M.J.; Galluzzi, L. Control. of metastasis by NK Cells. Cancer Cell 2017, 32, 135–154. [Google Scholar] [CrossRef]
- Larsen, S.K.; Gao, Y.; Basse, P.H. NK cells in the tumor microenvironment. Crit. Rev. Oncog. 2014, 19, 91–105. [Google Scholar] [CrossRef] [Green Version]
- Ishigami, S.; Natsugoe, S.; Tokuda, K.; Nakajo, A.; Che, X.; Iwashige, H.; Aridome, K.; Hokita, S.; Aikou, T. Prognostic value of intratumoral natural killer cells in gastric carcinoma. Cancer 2000, 88, 577–583. [Google Scholar] [CrossRef]
- Donskov, F.; von der Maase, H. Impact of immune parameters on long-term survival in metastatic renal cell carcinoma. J. Clin. Oncol. 2006, 24, 1997–2005. [Google Scholar] [CrossRef]
- Gannon, P.O.; Poisson, A.O.; Delvoye, N.; Lapointe, R.; Mes-Masson, A.M.; Saad, F. Characterization of the intra-prostatic immune cell infiltration in androgen-deprived prostate cancer patients. J. Immunol. Methods 2009, 348, 9–17. [Google Scholar] [CrossRef]
- Delahaye, N.F.; Rusakiewicz, S.; Martins, I.; Ménard, C.; Roux, S.; Lyonnet, L.; Paul, P.; Sarabi, M.; Chaput, N.; Semeraro, M.; et al. Alternatively spliced NKp30 isoforms affect the prognosis of gastrointestinal stromal tumors. Nat. Med. 2011, 17, 700–707. [Google Scholar] [CrossRef]
- Cursons, J.; Souza-Fonseca-Guimaraes, F.; Foroutan, M.; Anderson, A.; Hollande, F.; Hediyeh-Zadeh, S.; Behren, A.; Huntington, N.D.; Davis, M.J. A gene signature predicting natural killer cell infiltration and improved survival in melanoma patients. Cancer. Immunol. Res. 2019, 7, 1162–1174. [Google Scholar] [CrossRef] [Green Version]
- Denkert, C.; von Minckwitz, G.; Darb-Esfahani, S.; Lederer, B.; Heppner, B.I.; Weber, K.E.; Budczies, J.; Huober, J.; Klauschen, F.; Furlanetto, J.; et al. Tumour-infiltrating lymphocytes and prognosis in different subtypes of breast cancer: A pooled analysis of 3771 patients treated with neoadjuvant therapy. Lancet Oncol. 2018, 19, 40–50. [Google Scholar] [CrossRef]
- Liu, E.; Marin, D.; Banerjee, P.; Macapinlac, H.A.; Thompson, P.; Basar, R.; Nassif Kerbauy, L.; Overman, B.; Thall, P.; Kaplan, M.; et al. Use of CAR-transduced natural killer cells in CD19-positive lymphoid tumors. N. Engl. J. Med. 2020, 382, 545–553. [Google Scholar] [CrossRef]
- Long, E.O. Tumor cell recognition by natural killer cells. Semin. Cancer Biol. 2002, 12, 57–61. [Google Scholar] [CrossRef]
- Orr, M.T.; Lanier, L.L. Natural killer cell education and tolerance. Cell 2010, 142, 847–856. [Google Scholar] [CrossRef] [Green Version]
- Anfossi, N.; André, P.; Guia, S.; Falk, C.S.; Roetynck, S.; Stewart, C.A.; Breso, V.; Frassati, C.; Reviron, D.; Middleton, D.; et al. Human NK cell education by inhibitory receptors for MHC class I. Immunity 2006, 25, 331–342. [Google Scholar] [CrossRef]
- Bahram, S.; Bresnahan, M.; Geraghty, D.E.; Spies, T.A. second lineage of mammalian major histocompatibility complex class I genes. Proc. Natl. Acad. Sci. USA 1994, 91, 6259–6263. [Google Scholar] [CrossRef] [Green Version]
- He, Y.; Tian, Z. NK cell education via nonclassical MHC and non-MHC ligands. Cell Mol. Immunol. 2017, 14, 321–330. [Google Scholar] [CrossRef] [Green Version]
- Kelley, J.; Walter, L.; Trowsdale, J. Comparative genomics of natural killer cell receptor gene clusters. PLoS Genet. 2005, 1, 129–139. [Google Scholar] [CrossRef] [Green Version]
- Makrigiannis, A.P.; Pau, A.T.; Saleh, A.; Winkler-Pickett, R.; Ortaldo, J.R.; Anderson, S.K.K. Class I MHC-binding characteristics of the 129/J. Ly49 repertoire. J. Immunol. 2001, 166, 5034–5043. [Google Scholar] [CrossRef]
- Marsh, S.G.; Parham, P.; Dupont, B.; Geraghty, D.E.; Trowsdale, J.; Middleton, D.; Vilches, C.; Carrington, M.; Witt, C.; Guethlein, L.A.; et al. Killer-cell immunoglobulin-like receptor (KIR) nomenclature report, 2002. Immunogenetics 2003, 55, 220–226. [Google Scholar] [CrossRef]
- Campbell, K.S.; Purdy, A.K. Structure/function of human killer cell immunoglobulin-like receptors: Lessons from polymorphisms, evolution, crystal structures and mutations. Immunology 2011, 132, 315–325. [Google Scholar] [CrossRef]
- Parham, P. MHC class I molecules and KIRs in human history, health and survival. Nat. Rev. Immunol. 2005, 5, 201–214. [Google Scholar] [CrossRef]
- Kuroki, K.; Furukawa, A.; Maenaka, K. Molecular recognition of paired receptors in the immune system. Front. Microbiol. 2012, 3, 429. [Google Scholar] [CrossRef] [Green Version]
- Hansasuta, P.; Dong, T.; Thananchai, H.; Weekes, M.; Willberg, C.; Aldemir, H.; Rowland-Jones, S.; Braud, V.M. Recognition of HLA-A3 and HLA-A11 by KIR3DL2 is peptide-specific. Eur. J. Immunol. 2004, 34, 1673–1679. [Google Scholar] [CrossRef]
- Fanger, N.A.; Cosman, D.; Peterson, L.; Braddy, S.C.; Maliszewski, C.R.; Borges, L. The MHC class I binding proteins LIR-1 and LIR-2 inhibit Fc receptor-mediated signaling in monocytes. Eur. J. Immunol. 1998, 28, 3423–3434. [Google Scholar] [CrossRef]
- Colonna, M.; Navarro, F.; Bellón, T.; Llano, M.; García, P.; Samaridis, J.; Angman, L.; Cella, M.; López-Botet, M. A common inhibitory receptor for major histocompatibility complex class I molecules on human lymphoid and myelomonocytic cells. J. Exp. Med. 1997, 186, 1809–1818. [Google Scholar] [CrossRef] [Green Version]
- Borges, L.; Hsu, M.L.; Fanger, N.; Kubin, M.; Cosman, D.A. family of human lymphoid and myeloid Ig-like receptors, some of which bind to MHC class I molecules. J. Immunol. 1997, 159, 5192–5196. [Google Scholar]
- Samaridis, J.; Colonna, M. Cloning of novel immunoglobulin superfamily receptors expressed on human myeloid and lymphoid cells: Structural evidence for new stimulatory and inhibitory pathways. Eur. J. Immunol. 1997, 27, 660–665. [Google Scholar] [CrossRef]
- Lin, A.; Yan, W.H. Heterogeneity of HLA-G Expression in Cancers: Facing the Challenges. Front. Immunol. 2018, 9, 2164. [Google Scholar] [CrossRef]
- Paul, P.; Cabestré, F.A.; Le Gal, F.A.; Khalil-Daher, I.; Le Danff, C.; Schmid, M.; Mercier, S.; Avril, M.F.; Dausset, J.; Guillet, J.G.; et al. Heterogeneity of HLA-G gene transcription and protein expression in malignant melanoma biopsies. Cancer Res. 1999, 59, 1954–1960. [Google Scholar]
- Lefebvre, S.; Antoine, M.; Uzan, S.; McMaster, M.; Dausset, J.; Carosella, E.D.; Paul, P. Specific activation of the non-classical class I histocompatibility HLA-G antigen and expression of the ILT2 inhibitory receptor in human breast cancer. J. Pathol. 2002, 196, 266–274. [Google Scholar] [CrossRef]
- Fukushima, Y.; Oshika, Y.; Nakamura, M.; Tokunaga, T.; Hatanaka, H.; Abe, Y.; Yamazaki, H.; Kijima, H.; Ueyama, Y.; Tamaoki, N. Increased expression of human histocompatibility leukocyte antigen-G in colorectal cancer cells. Int. J. Mol. Med. 1998, 2, 349–351. [Google Scholar] [CrossRef]
- Rahim, M.M.; Tu, M.M.; Mahmoud, A.B.; Wight, A.; Abou-Samra, E.; Lima, P.D.; Makrigiannis, A.P. Ly49 receptors: Innate and adaptive immune paradigms. Front. Immunol. 2014, 5, 145. [Google Scholar] [CrossRef] [Green Version]
- Hanke, T.; Takizawa, H.; McMahon, C.W.; Busch, D.H.; Pamer, E.G.; Miller, J.D.; Altman, J.D.; Liu, Y.; Cado, D.; Lemonnier, F.A.; et al. Direct assessment of MHC class I binding by seven Ly49 inhibitory NK cell receptors. Immunity 1999, 11, 67–77. [Google Scholar] [CrossRef] [Green Version]
- Schenkel, A.R.; Kingry, L.C.; Slayden, R.A. The ly49 gene family. A brief guide to the nomenclature, genetics, and role in intracellular infection. Front. Immunol. 2013, 4, 90. [Google Scholar] [CrossRef] [Green Version]
- Kaiser, B.K.; Pizarro, J.C.; Kerns, J.; Strong, R.K. Structural basis for NKG2A/CD94 recognition of HLA-E. Proc. Natl. Acad. Sci. USA 2008, 105, 6696–6701. [Google Scholar] [CrossRef] [Green Version]
- Kraft, J.R.; Vance, R.E.; Pohl, J.; Martin, A.M.; Raulet, D.H.; Jensen, P.E. Analysis of Qa-1(b) peptide binding specificity and the capacity of CD94/NKG2A to discriminate between Qa-1-peptide complexes. J. Exp. Med. 2000, 192, 613–624. [Google Scholar] [CrossRef]
- Navarro, F.; Llano, M.; Bellón, T.; Colonna, M.; Geraghty, D.E.; López-Botet, M. The ILT2(LIR1) and CD94/NKG2A NK cell receptors respectively recognize HLA-G1 and HLA-E molecules co-expressed on target cells. Eur. J. Immunol. 1999, 29, 277–283. [Google Scholar] [CrossRef]
- Cosman, D.; Fanger, N.; Borges, L.; Kubin, M.; Chin, W.; Peterson, L.; Hsu, M.L. A novel immunoglobulin superfamily receptor for cellular and viral MHC class I molecules. Immunity 1997, 7, 273–282. [Google Scholar] [CrossRef] [Green Version]
- Chapman, T.L.; Heikeman, A.P.; Bjorkman, P.J. The inhibitory receptor LIR-1 uses a common binding interaction to recognize class I MHC molecules and the viral homolog UL18. Immunity 1999, 11, 603–613. [Google Scholar] [CrossRef] [Green Version]
- Angata, T.; Varki, A. Cloning, characterization, and phylogenetic analysis of siglec-9, a new member of the CD33-related group of siglecs. Evidence for co-evolution with sialic acid synthesis pathways. J. Biol. Chem. 2000, 275, 22127–22135. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.Q.; Nicoll, G.; Jones, C.; Crocker, P.R. Siglec-9, a novel sialic acid binding member of the immunoglobulin superfamily expressed broadly on human blood leukocytes. J. Biol. Chem. 2000, 275, 22121–22126. [Google Scholar] [CrossRef] [Green Version]
- Nicoll, G.; Ni, J.; Liu, D.; Klenerman, P.; Munday, J.; Dubock, S.; Mattei, M.G.; Crocker, P.R. Identification and characterization of a novel siglec, siglec-7, expressed by human natural killer cells and monocytes. J. Biol. Chem. 1999, 274, 34089–34095. [Google Scholar] [CrossRef] [Green Version]
- Aldemir, H.; Prod’homme, V.; Dumaurier, M.J.; Retiere, C.; Poupon, G.; Cazareth, J.; Bihl, F.; Braud, V.M. Cutting edge: Lectin-like transcript 1 is a ligand for the CD161 receptor. J. Immunol. 2005, 175, 7791–7795. [Google Scholar] [CrossRef] [Green Version]
- Rosen, D.B.; Bettadapura, J.; Alsharifi, M.; Mathew, P.A.; Warren, H.S.; Lanier, L.L. Cutting edge: Lectin-like transcript-1 is a ligand for the inhibitory human NKR-P1A receptor. J. Immunol. 2005, 175, 7796–7799. [Google Scholar] [CrossRef]
- Johansson, S.; Salmon-Divon, M.; Johansson, M.H.; Pickman, Y.; Brodin, P.; Kärre, K.; Mehr, R.; Höglund, P. Probing natural killer cell education by Ly49 receptor expression analysis and computational modelling in single MHC class I mice. PLoS ONE 2009, 4, 6046. [Google Scholar] [CrossRef]
- Kane, K.P. Ly-49 mediates EL4 lymphoma adhesion to isolated class I major histocompatibility complex molecules. J. Exp. Med. 1994, 179, 1011–1015. [Google Scholar] [CrossRef] [Green Version]
- Takei, F.; Brennan, J.; Mager, D.L. The Ly-49 family: Genes, proteins and recognition of class I MHC. Immunol. Rev. 1997, 155, 67–77. [Google Scholar] [CrossRef]
- Daniels, B.F.; Karlhofer, F.M.; Seaman, W.E.; Yokoyama, W.M. A natural killer cell receptor specific for a major histocompatibility complex class I molecule. J. Exp. Med. 1994, 180, 687–692. [Google Scholar] [CrossRef] [Green Version]
- Mehta, I.K.; Wang, J.; Roland., J.; Marguilies, D.H.; Yokoyama, W.M. Ly49A allelic variation and MHC class I specificity. Immunogenetics 2001, 53, 572–583. [Google Scholar] [CrossRef]
- Scarpellino, L.; Oeschger, F.; Guillaume, P.; Coudert, J.D.; Lévy, F.; Leclercq, G.; Held, W. Interactions of Ly49 family receptors with MHC class I ligands in trans and cis. J. Immunol. 2007, 178, 1277–1284. [Google Scholar] [CrossRef] [Green Version]
- Yu, Y.Y.; George, T.; Dorfman, J.R.; Roland, J.; Kumar, V.; Bennett, M. The role of Ly49A and 5E6(Ly49C) molecules in hybrid resistance mediated by murine natural killer cells against normal T cell blasts. Immunity 1996, 4, 67–76. [Google Scholar] [CrossRef] [Green Version]
- Dimasi, N.; Biassoni, R. Structural and functional aspects of the Ly49 natural killer cell receptors. Immunol. Cell Biol. 2005, 83, 1–8. [Google Scholar] [CrossRef]
- Andrews, D.M.; Sullivan, L.C.; Baschuk, N.; Chan, C.J.; Berry, R.; Cotterell, C.L.; Lin, J.; Halse, H.; Watt, S.V.; Poursine-Laurent, J.; et al. Recognition of the nonclassical MHC class I molecule H2-M3 by the receptor Ly49A regulates the licensing and activation of NK cells. Nat. Immunol. 2012, 13, 1171–1177. [Google Scholar] [CrossRef]
- Sullivan, L.C.; Berry, R.; Sosnin, N.; Widjaja, J.M.; Deuss, F.A.; Balaji, G.R.; LaGruta, N.L.; Mirams, M.; Trapani, J.A.; Rossjohn, J.; et al. Recognition of the Major Histocompatibility Complex (MHC) Class Ib Molecule H2-Q10 by the Natural Killer Cell Receptor Ly49C. J. Biol. Chem. 2016, 291, 18740–18752. [Google Scholar] [CrossRef] [Green Version]
- Abou-Samra, E.; Hickey, Z.; Aguilar, O.A.; Scur, M.; Mahmoud, A.B.; Pyatibrat, S.; Tu, M.M.; Francispillai, J.; Mortha, A.; Carlyle, J.R.; et al. NKR-P1B expression in gut-associated innate lymphoid cells is required for the control of gastrointestinal tract infections. Cell Mol. Immunol. 2019, 16, 868–877. [Google Scholar] [CrossRef]
- Castells, M.C.; Klickstein, L.B.; Hassani, K.; Cumplido, J.A.; Lacouture, M.E.; Austen, K.F.; Katz, H.R. Gp49B1-alpha(v)beta3 interaction inhibits antigen-induced mast cell activation. Nat. Immunol. 2001, 2, 436–442. [Google Scholar] [CrossRef]
- Van Den Broeck, T.; Stevenaert, F.; Taveirne, S.; Debacker, V.; Vangestel, C.; Vandekerckhove, B.; Taghon, T.; Matthys, P.; Plum, J.; Held, W.; et al. Ly49E-dependent inhibition of natural killer cells by urokinase plasminogen activator. Blood 2008, 112, 5046–5051. [Google Scholar] [CrossRef]
- Casado, J.G.; Pawelec, G.; Morgado, S.; Sanchez-Correa, B.; Delgado, E.; Gayoso, I.; Duran, E.; Solana, R.; Tarazona, R. Expression of adhesion molecules and ligands for activating and costimulatory receptors involved in cell-mediated cytotoxicity in a large panel of human melanoma cell lines. Cancer Immunol. Immunother. 2009, 58, 1517–1526. [Google Scholar] [CrossRef]
- Zhang, Q.; Bi, J.; Zheng, X.; Chen, Y.; Wang, H.; Wu, W.; Wang, Z.; Wu, Q.; Peng, H.; Wei, H.; et al. Blockade of the checkpoint receptor TIGIT prevents NK cell exhaustion and elicits potent anti-tumor immunity. Nat. Immunol. 2018, 19, 723–732. [Google Scholar] [CrossRef]
- Chen, Z.; Chen, L.; Baker, K.; Olszak, T.; Zeissig, S.; Huang, Y.-H.; Kuo, T.T.; Mandelboim, O.; Beauchemin, N.; Lanier, L.L.; et al. CEACAM1 dampens antitumor immunity by down-regulating NKG2D ligand expression on tumor cells. J. Exp. Med. 2011, 208, 2633–2640. [Google Scholar] [CrossRef]
- Rygiel, T.P.; Stolte, E.H.; de Ruiter, T.; van de Weijer, M.L.; Meyaard, L. Tumor-expressed collagens can modulate immune cell function through the inhibitory collagen receptor LAIR-1. Mol. Immunol. 2011, 49, 402–406. [Google Scholar] [CrossRef]
- Greenberg, S.A.; Kong, S.W.; Thompson, E.; Gulla, S.V. Co-inhibitory T cell receptor KLRG1: Human cancer expression and efficacy of neutralization in murine cancer models. Oncotarget 2019, 10, 1399–1406. [Google Scholar] [CrossRef]
- Li, Y.; Hofmann, M.; Wang, Q.; Teng, L.; Chlewicki, L.K.; Pircher, H. Mariuzza RA. Structure of natural killer cell receptor KLRG1 bound to E-cadherin reveals basis for MHC-independent missing self recognition. Immunity 2009, 31, 35–46. [Google Scholar] [CrossRef] [Green Version]
- Paolino, M.; Choidas, A.; Wallner, S.; Pranjic, B.; Uribesalgo, I.; Loeser, S.; Jamieson, A.M.; Langdon, W.Y.; Ikeda, F.; Fededa, J.P.; et al. The E3 ligase Cbl-b and TAM receptors regulate cancer metastasis via natural killer cells. Nature 2014, 507, 508–512. [Google Scholar] [CrossRef]
- Stitt, T.N.; Conn, G.; Gore, M.; Lai, C.; Bruno, J.; Radziejewski, C.; Mattsson, K.; Fisher, J.; Gies, D.R.; Jones, P.F.; et al. The anticoagulation factor protein S and its relative, Gas6, are ligands for the Tyro 3/Axl family of receptor tyrosine kinases. Cell 1995, 80, 661–670. [Google Scholar] [CrossRef] [Green Version]
- Yokoyama, W.M.; Kehn, P.J.; Cohen, D.I.; Shevach, E.M. Chromosomal location of the Ly-49 (A1, YE1/48) multigene family. Genetic association with the NK 1.1 antigen. J. Immunol. 1990, 145, 2353–2358. [Google Scholar]
- Zeleznjak, J.; Popovic, B.; Krmpotic, A.; Jonjic, S.; Lisnic, V.J. Mouse cytomegalovirus encoded immunoevasins and evolution of Ly49 receptors-Sidekicks or enemies? Immunol. Lett. 2017, 189, 40–47. [Google Scholar] [CrossRef]
- Franksson, L.; Sundbäck, J.; Achour, A.; Bernlind, J.; Glas, R.; Kärre, K. Peptide dependency and selectivity of the NK cell inhibitory receptor Ly-49C. Eur. J. Immunol. 1999, 29, 2748–2758. [Google Scholar] [CrossRef]
- Thomas, M.L. Of ITAMs and ITIMs: Turning on and off the B cell antigen receptor. J. Exp. Med. 1995, 181, 1953–1956. [Google Scholar] [CrossRef] [Green Version]
- Lazetic, S.; Chang, C.; Houchins, J.P.; Lanier, L.L.; Phillips, J.H. Human natural killer cell receptors involved in MHC class I recognition are disulfide-linked heterodimers of CD94 and NKG2 subunits. J. Immunol. 1996, 157, 4741–4745. [Google Scholar]
- Brooks, A.G.; Posch, P.E.; Scorzelli, C.J.; Borrego, F.; Coligan, J.E. NKG2A complexed with CD94 defines a novel inhibitory natural killer cell receptor. J. Exp. Med. 1997, 185, 795–800. [Google Scholar] [CrossRef] [Green Version]
- Carretero, M.; Cantoni, C.; Bellón, T.; Bottino, C.; Biassoni, R.; Rodríguez, A.; Pérez-Villar, J.J.; Moretta, L.; Moretta, A.; López-Botet, M. The CD94 and NKG2-A C-type lectins covalently assemble to form a natural killer cell inhibitory receptor for HLA class I molecules. Eur. J. Immunol. 1997, 27, 563–567. [Google Scholar] [CrossRef]
- Ivarsson, M.A.; Loh, L.; Marquardt, N.; Kekäläinen, E.; Berglin, L.; Björkström, N.K.; Westgren, M.; Nixon, D.F.; Michaëlsson, J. Differentiation and functional regulation of human fetal NK cells. J. Clin. Investig. 2013, 123, 3889–3901. [Google Scholar] [CrossRef] [Green Version]
- Lisovsky, I.; Isitman, G.; Song, R.; DaFonseca, S.; Tremblay-McLean, A.; Lebouché, B.; Routy, J.P.; Bruneau, J.; Bernard, N.F. A Higher Frequency of NKG2A+ than of NKG2A- NK Cells Responds to Autologous HIV-Infected CD4 Cells irrespective of Whether or Not They Coexpress KIR3DL1. J. Virol. 2015, 89, 9909–9919. [Google Scholar] [CrossRef] [Green Version]
- Braud, V.M.; Allan, D.S.; O’Callaghan, C.A.; Söderström, K.; D’Andrea, A.; Ogg, G.S.; Lazetic, S.; Young, N.T.; Bell, J.I.; Phillips, J.H.; et al. HLA-E binds to natural killer cell receptors CD94/NKG2A, B and C. Nature 1998, 391, 795–799. [Google Scholar] [CrossRef]
- Vivier, E.; Raulet, D.H.; Moretta, A.; Caligiuri, M.A.; Zitvogel, L.; Lanier, L.L.; Yokoyama, W.M.; Ugolini, S. Innate or adaptive immunity? The example of natural killer cells. Science 2011, 331, 44–49. [Google Scholar] [CrossRef] [Green Version]
- Leung, W. Infusions of allogeneic natural killer cells as cancer therapy. Clin. Cancer Res. 2014, 20, 3390–4300. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Wu, N.; Lu, Y.; Davidson, D.; Colonna, M.; Veillette, A. DNAM-1 controls NK cell activation via an ITT-like motif. J. Exp. Med. 2015, 212, 2165–2182. [Google Scholar] [CrossRef]
- Blake, S.J.; Dougall, W.C.; Miles, J.J.; Teng, M.W.; Smyth, M.J. Molecular Pathways: Targeting CD96 and TIGIT for Cancer Immunotherapy. Clin. Cancer Res. 2016, 22, 5183–5188. [Google Scholar] [CrossRef] [Green Version]
- Yu, X.; Harden, K.; Gonzalez, L.C.; Francesco, M.; Chiang, E.; Irving, B.; Tom, I.; Ivelja, S.; Refino, C.J.; Clark, H.; et al. The surface protein TIGIT suppresses T cell activation by promoting the generation of mature immunoregulatory dendritic cells. Nat. Immunol. 2009, 10, 48–57. [Google Scholar] [CrossRef]
- Georgiev, H.; Ravens, I.; Papadogianni, G.; Bernhardt, G. Coming of Age: CD96 Emerges as Modulator of Immune Responses. Front. Immunol. 2018, 9, 1072. [Google Scholar] [CrossRef] [Green Version]
- Sharma, P.; Allison, J.P. The future of immune checkpoint therapy. Science 2015, 348, 56–61. [Google Scholar] [CrossRef]
- Dong, H.; Zhu, G.; Tamada, K.; Chen, L. B7-H1, a third member of the B7 family, co-stimulates T-cell proliferation and interleukin-10 secretion. Nat. Med. 1999, 5, 1365–1369. [Google Scholar] [CrossRef]
- Latchman, Y.; Wood, C.R.; Chernova, T.; Chaudhary, D.; Borde, M.; Chernova, I.; Iwai, Y.; Long, A.J.; Brown, J.A.; Nunes, R.; et al. PD-L2 is a second ligand for PD-1 and inhibits T cell activation. Nat. Immunol. 2001, 2, 261–268. [Google Scholar] [CrossRef]
- Arasanz, H.; Gato-Cañas, M.; Zuazo, M.; Ibáñez-Vea, M.; Breckpot, K.; Kochan, G.; Escors, D. PD1 signal transduction pathways in T cells. Oncotarget 2017, 8, 51936–51945. [Google Scholar] [CrossRef] [Green Version]
- Keir, M.E.; Butte, M.J.; Freeman, G.J.; Sharpe, A.H. PD-1 and its ligands in tolerance and immunity. Annu. Rev. Immunol. 2008, 26, 677–704. [Google Scholar] [CrossRef] [Green Version]
- Lieberman, N.A.P.; DeGolier, K.; Haberthur, K.; Chinn, H.; Moyes, K.W.; Bouchlaka, M.; Walker, K.L.; Capitini, C.M.; Crane, C.A. An uncoupling of canonical phenotypic markers and functional potency of ex vivo-expanded natural killer cells. Front. Immunol. 2018, 9, 150. [Google Scholar] [CrossRef] [Green Version]
- Locatelli, F.; Pende, D.; Maccario, R.; Mingari, M.C.; Moretta, A.; Moretta, L. Haploidentical hemopoietic stem cell transplantation for the treatment of high-risk leukemias: How NK cells make the difference. Clin. Immunol. 2009, 133, 171–178. [Google Scholar] [CrossRef]
- Hsu, J.; Hodgins, J.J.; Marathe, M.; Nicolai, C.J.; Bourgeois-Daigneault, M.-C.; Trevino, T.N.; Azimi, C.S.; Scheer, A.K.; Randolph, H.E.; Thompson, T.W.; et al. Contribution of NK cells to immunotherapy mediated by PD-1/PD-L1 blockade. J. Clin. Investig. 2018, 128, 4654–4668. [Google Scholar] [CrossRef]
- Pesce, S.; Greppi, M.; Tabellini, G.; Rampinelli, F.; Parolini, S.; Olive, D.; Moretta, L.; Moretta, A.; Marcenaro, E. Identification of a subset of human natural killer cells expressing high levels of programmed death 1: A phenotypic and functional characterization. J. Allergy Clin. Immunol. 2017, 139, 335–346. [Google Scholar] [CrossRef] [Green Version]
- Judge, S.J.; Dunai, C.; Aguilar, E.G.; Vick, S.C.; Sturgill, I.R.; Khuat, L.T.; Stoffel, K.M.; Van Dyke, J.; Longo, D.L.; Darrow, M.A.; et al. Minimal PD-1 expression in mouse and human NK cells under diverse conditions. J. Clin. Investig. 2020. [Google Scholar] [CrossRef] [Green Version]
- Anderson, A.C.; Joller, N.; Kuchroo, V.K. Lag-3, Tim-3, and TIGIT: Co-inhibitory Receptors with Specialized Functions in Immune Regulation. Immunity 2016, 44, 989–1004. [Google Scholar] [CrossRef] [Green Version]
- Van De Weyer, P.S.; Muehlfeit, M.; Klose, C.; Bonventre, J.V.; Walz, G.; Kuehn, E.W. A highly conserved tyrosine of Tim-3 is phosphorylated upon stimulation by its ligand galectin-9. Biochem. Biophys. Res. Commun. 2006, 351, 571–576. [Google Scholar] [CrossRef]
- Huang, Y.-H.; Zhu, C.; Kondo, Y.; Anderson, A.C.; Gandhi, A.; Russell, A.; Dougan, S.K.; Löscher, B.-S.; Melum, E.; Pertel, T.; et al. CEACAM1 regulates TIM-3-mediated tolerance and exhaustion. Nature 2015, 517, 386–390. [Google Scholar] [CrossRef] [Green Version]
- DeKruyff, R.H.; Bu, X.; Ballesteros, A.; Santiago, C.; Chim, Y.-L.E.; Lee, H.-H.; Karisola, P.; Pichavant, M.; Kaplan, G.G.; Umetsu, D.T.; et al. T cell/transmembrane, Ig, and mucin-3 allelic variants differentially recognize phosphatidylserine and mediate phagocytosis of apoptotic cells. J. Immunol. 2010, 184, 1918–1930. [Google Scholar] [CrossRef] [Green Version]
- Chiba, S.; Baghdadi, M.; Akiba, H.; Yoshiyama, H.; Kinoshita, I.; Dosaka-Akita, H.; Fujioka, Y.; Ohba, Y.; Gorman, J.V.; Colgan, J.D.; et al. Tumor-infiltrating DCs suppress nucleic acid-mediated innate immune responses through interactions between the receptor TIM-3 and the alarmin HMGB1. Nat. Immunol. 2012, 13, 832–842. [Google Scholar] [CrossRef]
- Wolf, Y.; Anderson, A.C.; Kuchroo, V.K. TIM3 comes of age as an inhibitory receptor. Nat. Rev. Immunol. 2020, 20, 173–185. [Google Scholar] [CrossRef]
- Ndhlovu, L.C.; Lopez-Vergès, S.; Barbour, J.D.; Jones, R.B.; Jha, A.; Long, B.R.; Schoeffler, E.C.; Fujita, T.; Nixon, D.F.; Lanier, L.L. Tim-3 marks human natural killer cell maturation and suppresses cell-mediated cytotoxicity. Blood 2012, 119, 3734–3743. [Google Scholar] [CrossRef] [Green Version]
- Gleason, M.K.; Lenvik, T.R.; McCullar, V.; Felices, M.; O’Brien, M.S.; Cooley, S.A.; Verneris, M.R.; Cichocki, F.; Holman, C.; Panoskaltsis-Mortari, A.; et al. Tim-3 is an inducible human natural killer cell receptor that enhances interferon gamma production in response to galectin-9. Blood 2012, 119, 3064–3072. [Google Scholar] [CrossRef]
- So, E.C.; Khaladj-Ghom, A.; Ji, Y.; Amin, J.; Song, Y.; Burch, E.; Zhou, H.; Sun, H.; Chen, S.; Bentzen, S.; et al. NK cell expression of Tim-3: First impressions matter. Immunobiology 2019, 224, 362–370. [Google Scholar] [CrossRef]
- Huard, B.; Mastrangeli, R.; Prigent, P.; Bruniquel, D.; Donini, S.; El-Tayar, N.; Maigret, B.; Dréano, M.; Triebel, F. Characterization of the major histocompatibility complex class II binding site on LAG-3 protein. Proc. Natl. Acad. Sci. USA 1997, 94, 5744–5749. [Google Scholar] [CrossRef] [Green Version]
- Workman, C.J.; Cauley, L.S.; Kim, I.-J.; Blackman, M.A.; Woodland, D.L.; Vignali, D.A.A. Lymphocyte activation gene-3 (CD223) regulates the size of the expanding T cell population following antigen activation in vivo. J. Immunol. 2004, 172, 5450–5455. [Google Scholar] [CrossRef]
- Triebel, F.; Jitsukawa, S.; Baixeras, E.; Roman-Roman, S.; Genevée, C.; Viegas-Pequignot, E.; Hercend, T. LAG-3, a novel lymphocyte activation gene closely related to CD4. J. Exp. Med. 1990, 171, 1393–1405. [Google Scholar] [CrossRef] [Green Version]
- Miyazaki, T.; Dierich, A.; Benoist, C.; Mathis, D. Independent modes of natural killing distinguished in mice lacking Lag3. Science 1996, 272, 405–408. [Google Scholar] [CrossRef]
- Huard, B.; Tournier, M.; Triebel, F. LAG-3 does not define a specific mode of natural killing in human. Immunol. Lett. 1998, 61, 109–112. [Google Scholar] [CrossRef]
- Lebbink, R.J.; De Ruiter, T.; Adelmeijer, J.; Brenkman, A.B.; Van Helvoort, J.M.; Koch, M.; Farndale, R.W.; Lisman, T.; Sonnenberg, A.; Lenting, P.J.; et al. Collagens are functional, high affinity ligands for the inhibitory immune receptor LAIR-1. J. Exp. Med. 2006, 203, 1419–1425. [Google Scholar] [CrossRef] [Green Version]
- Angata, T.; Varki, A. Siglec-7: A sialic acid-binding lectin of the immunoglobulin superfamily. Glycobiology 2000, 10, 431–438. [Google Scholar] [CrossRef] [Green Version]
- Alphey, M.S.; Attrill, H.; Crocker, P.; Van Aalten, D.M.F. High resolution crystal structures of Siglec-7. Insights into ligand specificity in the Siglec family. J. Biol. Chem. 2003, 278, 3372–3377. [Google Scholar] [CrossRef] [Green Version]
- Varki, A. Since there are PAMPs and DAMPs, there must be SAMPs? Glycan “self-associated molecular patterns” dampen innate immunity, but pathogens can mimic them. Glycobiology 2011, 21, 1121–1214. [Google Scholar] [CrossRef]
- Gasic, G.; Gasic, T. Removal of sialic acid from the cell coat in tumor cells and vascular endothelium, and its effects on metastasis. Proc. Natl. Acad. Sci. USA 1962, 48, 1172–1177. [Google Scholar] [CrossRef] [Green Version]
- Büll, C.; Stoel, M.A.; Brok, M.H.D.; Adema, G.J. Sialic acids sweeten a tumor’s life. Cancer Res. 2014, 74, 3199–3204. [Google Scholar] [CrossRef] [Green Version]
- Nicoll, G.; Avril, T.; Lock, K.; Furukawa, K.; Bovin, N.; Crocker, P. Ganglioside GD3 expression on target cells can modulate NK cell cytotoxicity via siglec-7-dependent and -independent mechanisms. Eur. J. Immunol. 2003, 33, 1642–1648. [Google Scholar] [CrossRef]
- Saito, S.; Orikasa, S.; Satoh, M.; Ohyama, C.; Ito, A.; Takahashi, T. Expression of globo-series gangliosides in human renal cell carcinoma. Jpn. J. Cancer Res. 1997, 88, 652–659. [Google Scholar] [CrossRef]
- Kawasaki, Y.; Ito, A.A.; Withers, D.; Taima, T.; Kakoi, N.; Saito, S.; Arai, Y. Ganglioside DSGb5, preferred ligand for Siglec-7, inhibits NK cell cytotoxicity against renal cell carcinoma cells. Glycobiology 2010, 20, 1373–1379. [Google Scholar] [CrossRef] [Green Version]
- Leaton, L.; Shortt, J.; Kichula, K.M.; Tao, S.; Nemat-Gorgani, N.; Mentzer, A.J.; Oppenheimer, S.J.; Deng, Z.; Hollenbach, J.A.; Gignoux, C.R.; et al. Conservation, extensive heterozygosity, and convergence of signaling potential all indicate a critical role for KIR3DL3 in higher primates. Front. Immunol. 2019, 10, 24. [Google Scholar] [CrossRef]
- Brown, D.; Trowsdale, J.; Allen, R. The LILR family: Modulators of innate and adaptive immune pathways in health and disease. Tissue Antigens 2004, 64, 215–225. [Google Scholar] [CrossRef]
- Kirkham, C.L.; Carlyle, J.R. Complexity and diversity of the NKR-P1:Clr (Klrb1:Clec2) recognition systems. Front. Immunol. 2014, 5, 214. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, M.C.; Niemi, E.C.; Fisher, M.J.; Shultz, L.D.; Seaman, W.E.; Ryan, J.C. Mouse Ly-49A interrupts early signaling events in natural killer cell cytotoxicity and functionally associates with the SHP-1 tyrosine phosphatase. J. Exp. Med. 1997, 185, 673–684. [Google Scholar] [CrossRef] [Green Version]
- Coxon, C.H.; Geer, M.J.; Senis, Y.A. ITIM receptors: More than just inhibitors of platelet activation. Blood 2017, 129, 3407–3418. [Google Scholar] [CrossRef] [Green Version]
- Müller, M.; Klaile, E.; Vorontsova, O.; Singer, B.B.; Öbrink, B. Homophilic adhesion and CEACAM1-S regulate dimerization of CEACAM1-L and recruitment of SHP-2 and c-Src. J. Cell Biol. 2009, 187, 569–581. [Google Scholar] [CrossRef] [Green Version]
- Meyaard, L.; Adema, G.J.; Chang, C.; Woollatt, E.; Sutherland, G.R.; Lanier, L.L.; Phillips, J.H. LAIR-1, a novel inhibitory receptor expressed on human mononuclear leukocytes. Immunity 1997, 7, 283–290. [Google Scholar] [CrossRef]
- Hanke, T.; Corral, L.E.; Vance, R.; Raulet, D.H. 2F1 antigen, the mouse homolog of the rat “mast cell function-associated antigen”, is a lectin-like type II transmembrane receptor expressed by natural killer cells. Eur. J. Immunol. 1998, 28, 4409–4417. [Google Scholar] [CrossRef]
- Guthmann, M.D.; Tal, M.; Pecht, I. A secretion inhibitory signal transduction molecule on mast cells is another C-type lectin. Proc. Natl. Acad. Sci. USA 1995, 92, 9397–9401. [Google Scholar] [CrossRef] [Green Version]
- Rothlin, C.V.; Carrera-Silva, E.A.; Bosurgi, L.; Ghosh, S. TAM receptor signaling in immune homeostasis. Annu. Rev. Immunol. 2015, 33, 355–391. [Google Scholar] [CrossRef] [Green Version]
- Hosomi, S.; Chen, Z.; Baker, K.; Chen, L.; Huang, Y.-H.; Olszak, T.; Zeissig, S.; Wang, J.H.; Mandelboim, O.; Beauchemin, N.; et al. CEACAM1 on activated NK cells inhibits NKG2D-mediated cytolytic function and signaling. Eur. J. Immunol. 2013, 43, 2473–2483. [Google Scholar] [CrossRef]
- Huntington, N.D.; Tabarias, H.; Fairfax, K.A.; Brady, J.; Hayakawa, Y.; Degli-Esposti, M.A.; Smyth, M.J.; Tarlinton, D.; Nutt, S.L. NK Cell Maturation and Peripheral Homeostasis Is Associated with KLRG1 Up-Regulation. J. Immunol. 2007, 178, 4764–4770. [Google Scholar] [CrossRef]
- Park, I.-K.; Giovenzana, C.; Hughes, T.L.; Yu, J.; Trotta, R.; Caligiuri, M.A. The Axl/Gas6 pathway is required for optimal cytokine signaling during human natural killer cell development. Blood 2009, 113, 2470–2477. [Google Scholar] [CrossRef] [Green Version]
- Caraux, A.; Lu, Q.; Fernandez, N.; Riou, S.; Di Santo, J.P.; Raulet, D.H.; Lemke, G.; Roth, C. Natural killer cell differentiation driven by Tyro3 receptor tyrosine kinases. Nat. Immunol. 2006, 7, 747–754. [Google Scholar] [CrossRef]
- Chirino, L.M.; Kumar, S.; Okumura, M.; Sterner, D.E.; Mattern, M.; Butt, T.R.; Kambayashi, T. TAM receptors attenuate murine NK-cell responses via E3 ubiquitin ligase Cbl-b. Eur. J. Immunol. 2020, 50, 48–55. [Google Scholar] [CrossRef]
- Vey, N.; Karlin, L.; Sadot-Lebouvier, S.; Broussais, F.; Berton-Rigaud, D.; Rey, J.; Charbonnier, A.; Marie, D.; André, P.; Paturel, C.; et al. A phase 1 study of lirilumab (antibody against killer immunoglobulin-like receptor antibody KIR2D.; IPH2102) in patients with solid tumors and hematologic malignancies. Oncotarget 2018, 9, 17675–17688. [Google Scholar] [CrossRef] [Green Version]
- Vey, N.; Bourhis, J.-H.; Boissel, N.; Bordessoule, D.; Prebet, T.; Charbonnier, A.; Etienne, A.; Andre, P.; Romagné, F.; Benson, N.; et al. A phase 1 trial of the anti-inhibitory KIR mAb IPH2101 for AML in complete remission. Blood 2012, 120, 4317–4323. [Google Scholar] [CrossRef] [Green Version]
- Tinker, A.V.; Hirte, H.W.; Provencher, D.M.; Butler, M.O.; Ritter, H.; Tu, D.; Azim, H.A.; Paralejas, P.; Grenier, N.; Hahn, S.-A.; et al. Dose-ranging and cohort-expansion study of monalizumab (IPH2201) in patients with advanced gynecologic malignancies: A trial of the canadian cancer trials group (CCTG): IND221. Clin. Cancer Res. 2019, 25, 6052–6060. [Google Scholar] [CrossRef] [Green Version]
- Chan, V.W.; Lowell, C.A.; DeFranco, A.L. Defective negative regulation of antigen receptor signaling in Lyn-deficient B lymphocytes. Curr. Biol. 1998, 8, 545–553. [Google Scholar] [CrossRef] [Green Version]
- Burshtyn, D.; Scharenberg, A.M.; Wagtmann, N.; Rajagopalan, S.; Berrada, K.; Yi, T.; Kinet, J.-P.; Long, E.O. Recruitment of tyrosine phosphatase HCP by the killer cell inhibitor receptor. Immunity 1996, 4, 77–85. [Google Scholar] [CrossRef] [Green Version]
- Binstadt, B.; Brumbaugh, K.M.; Dick, C.J.; Scharenberg, A.M.; Williams, B.L.; Colonna, M.; Lanier, L.L.; Kinet, J.-P.; Abraham, R.T.; Leibson, P.J. Sequential involvement of Lck and SHP-1 with MHC-recognizing receptors on NK cells inhibits FcR-initiated tyrosine kinase activation. Immunity 1996, 5, 629–638. [Google Scholar] [CrossRef] [Green Version]
- Lowin-Kropf, B.; Kunz, B.; Beermann, F.; Held, W. Impaired natural killing of MHC class I-deficient targets by NK cells expressing a catalytically inactive form of SHP-1. J. Immunol. 2000, 165, 1314–1321. [Google Scholar] [CrossRef] [Green Version]
- Viant, C.; Fenis, A.; Chicanne, G.; Payrastre, B.; Ugolini, S.; Vivier, E. SHP-1-mediated inhibitory signals promote responsiveness and anti-tumour functions of natural killer cells. Nat. Commun. 2014, 5, 5108. [Google Scholar] [CrossRef]
- Binstadt, B.; Billadeau, D.D.; Jevremovic, D.; Williams, B.L.; Fang, N.; Yi, T.; Koretzky, G.A.; Abraham, R.T.; Leibson, P.J. SLP-76 is a direct substrate of SHP-1 recruited to killer cell inhibitory receptors. J. Biol. Chem. 1998, 273, 27518–27523. [Google Scholar] [CrossRef] [Green Version]
- Valiante, N.M.; Phillips, J.H.; Lanier, L.L.; Parham, P. Killer cell inhibitory receptor recognition of human leukocyte antigen (HLA) class I blocks formation of a pp36/PLC-gamma signaling complex in human natural killer (NK) cells. J. Exp. Med. 1996, 184, 2243–2250. [Google Scholar] [CrossRef] [Green Version]
- Plas, D.R.; Johnson, R.; Pingel, J.T.; Matthews, R.J.; Dalton, M.; Roy, G.; Chan, A.C.; Thomas, M.L.; Mellors, J.W.; Rinaldo, C.; et al. Direct regulation of ZAP-70 by SHP-1 in T cell antigen receptor signaling. Science 1996, 272, 1173–1176. [Google Scholar] [CrossRef]
- Kabat, J.; Borrego, F.; Brooks, A.; Coligan, J.E. Role that each NKG2A immunoreceptor tyrosine-based inhibitory motif plays in mediating the human CD94/NKG2A inhibitory signal. J. Immunol. 2002, 169, 1948–1958. [Google Scholar] [CrossRef] [Green Version]
- Hosomi, S.; Grootjans, J.; Huang, Y.-H.; Kaser, A.; Blumberg, R.S. New insights into the regulation of natural-killer group 2 member D (NKG2D) and NKG2D-ligands: Endoplasmic reticulum stress and CEA-related cell adhesion molecule 1. Front. Immunol. 2018, 9, 1324. [Google Scholar] [CrossRef]
- Ljutic, B.; Carlyle, J.R.; Filipp, M.; Nakagawa, R.; Julius, M.; Zúñiga-Pflücker, J.C. Functional requirements for signaling through the stimulatory and inhibitory mouse NKR-P1 (CD161) NK cell receptors. J. Immunol. 2005, 174, 4789–4796. [Google Scholar] [CrossRef] [Green Version]
- Yu, M.-C.; Su, L.-L.; Zou, L.; Liu, Y.; Wu, N.; Kong, L.; Zhuang, Z.-H.; Sun, L.; Liu, H.; Hu, J.-H.; et al. An essential function for beta-arrestin 2 in the inhibitory signaling of natural killer cells. Nat. Immunol. 2008, 9, 898–907. [Google Scholar] [CrossRef]
- Fournier, N.; Chalus, L.; Durand, I.; Garcia, E.; Pin, J.-J.; Churakova, T.; Patel, S.; Zlot, C.; Gorman, D.; Zurawski, S.; et al. FDF03, a novel inhibitory receptor of the immunoglobulin superfamily, is expressed by human dendritic and myeloid cells. J. Immunol. 2000, 165, 1197–1209. [Google Scholar] [CrossRef] [Green Version]
- Lebbink, R.J.; De Ruiter, T.; Verbrugge, A.; Bril, W.S.; Meyaard, L. The mouse homologue of the leukocyte-associated Ig-like receptor-1 is an inhibitory receptor that recruits Src homology region 2-containing protein tyrosine phosphatase (SHP)-2, but not SHP-1. J. Immunol. 2004, 172, 5535–5543. [Google Scholar] [CrossRef] [Green Version]
- Bellón, T.; Kitzig, F.; Sayós, J.; López-Botet, M. Mutational analysis of immunoreceptor tyrosine-based inhibition motifs of the Ig-like transcript 2 (CD85j) leukocyte receptor. J. Immunol. 2002, 168, 3351–3359. [Google Scholar] [CrossRef] [Green Version]
- Fong, J.J.; Tsai, C.-M.; Saha, S.; Nizet, V.; Varki, A.; Bui, J. Siglec-7 engagement by GBS beta-protein suppresses pyroptotic cell death of natural killer cells. Proc. Natl. Acad. Sci. USA 2018, 115, 10410–10415. [Google Scholar] [CrossRef] [Green Version]
- Yamaji, T.; Mitsuki, M.; Teranishi, T.; Hashimoto, Y. Characterization of inhibitory signaling motifs of the natural killer cell receptor Siglec-7: Attenuated recruitment of phosphatases by the receptor is attributed to two amino acids in the motifs. Glycobiology 2005, 15, 667–676. [Google Scholar] [CrossRef] [Green Version]
- Orr, S.; Morgan, N.M.; Buick, R.J.; Boyd, C.R.; Elliott, J.; Burrows, J.F.; Jefferies, C.A.; Crocker, P.; Johnston, J.A. SOCS3 targets Siglec 7 for proteasomal degradation and blocks Siglec 7-mediated responses. J. Biol. Chem. 2007, 282, 3418–3422. [Google Scholar] [CrossRef] [Green Version]
- Liu, S.; Zhang, H.; Li, M.; Hu, D.; Li, C.; Ge, B.; Jin, B.; Fan, Z. Recruitment of Grb2 and SHIP1 by the ITT-like motif of TIGIT suppresses granule polarization and cytotoxicity of NK cells. Cell Death Differ. 2013, 20, 456–464. [Google Scholar] [CrossRef]
- Peterson, M.E.; Long, E.O. Inhibitory receptor signaling via tyrosine phosphorylation of the adaptor Crk. Immunity 2008, 29, 578–588. [Google Scholar] [CrossRef] [Green Version]
- Liu, D.; Peterson, M.E.; Long, E.O. The adaptor protein Crk controls activation and inhibition of natural killer cells. Immunity 2012, 36, 600–611. [Google Scholar] [CrossRef] [Green Version]
- Ganesan, S.; Luu, T.T.; Chambers, B.J.; Meinke, S.; Brodin, P.; Vivier, E.; Wetzel, D.M.; Koleske, A.; Kadri, N.; Höglund, P.; et al. The Abl-1 kinase is dispensable for NK cell inhibitory signalling and is not involved in murine NK cell education. Scand. J. Immunol. 2017, 86, 135–142. [Google Scholar] [CrossRef]
- Holcomb, M.; Rufini, A.; Barila, D.; Klemke, R.L. Deregulation of proteasome function induces Abl-mediated cell death by uncoupling p130CAS and c-CrkII. J. Biol. Chem. 2006, 281, 2430–2440. [Google Scholar] [CrossRef] [Green Version]
- Kobashigawa, Y.; Tomitaka, A.; Kumeta, H.; Noda, N.N.; Yamaguchi, M.; Inagaki, F. Autoinhibition and phosphorylation-induced activation mechanisms of human cancer and autoimmune disease-related E3 protein Cbl-b. Proc. Natl. Acad. Sci. USA 2011, 108, 20579–20584. [Google Scholar] [CrossRef] [Green Version]
- Watzl, C.; Long, E.O. Signal transduction during activation and inhibition of natural killer cells. Curr. Protoc. Immunol. 2010, 11, 9–11. [Google Scholar] [CrossRef] [Green Version]
- Seidel, U.J.; Schlegel, P.; Lang, P. Natural killer cell mediated antibody-dependent cellular cytotoxicity in tumor immunotherapy with therapeutic antibodies. Front. Immunol. 2013, 4, 76. [Google Scholar] [CrossRef] [Green Version]
- Kruse, P.H.; Matta, J.; Ugolini, S.; Vivier, E. Natural cytotoxicity receptors and their ligands. Immunol. Cell Biol. 2014, 9, 221–229. [Google Scholar] [CrossRef]
- Gutierrez-Franco, J.; Hernandez-Gutierrez, R.; Bueno-Topete, M.R.; Haramati, J.; Navarro-Hernandez, R.-E.; Escarra-Senmarti, M.; Vega-Magaña, N.; Del Toro-Arreola, A.; Pereira-Suarez, A.L.; Del Toro-Arreola, S. Characterization of B7H6, an endogenous ligand for the NK cell activating receptor NKp30, reveals the identity of two different soluble isoforms during normal human pregnancy. Immunobiology 2018, 223, 57–63. [Google Scholar] [CrossRef]
- Bjørnsen, E.G.; Thiruchelvam-Kyle, L.; Hoelsbrekken, S.E.; Henden, C.; Saether, P.C.; Boysen, P.; Daws, M.R.; Dissen, E. B7H6 is a functional ligand for NKp30 in rat and cattle and determines NKp30 reactivity toward human cancer cell lines. Eur. J. Immunol. 2019, 49, 54–65. [Google Scholar] [CrossRef] [Green Version]
- Rajagopalan, S.; Long, E.O. Found.: A cellular activating ligand for NKp44. Blood 2013, 122, 2921–2922. [Google Scholar] [CrossRef] [Green Version]
- Vieillard, V.; Baychelier, F.; Debre, P. NKp44L: A new tool for fighting cancer. Oncoimmunology 2014, 3, 27988. [Google Scholar] [CrossRef]
- Bauer, S.; Groh, V.; Wu, J.; Steinle, A.; Phillips, J.H.; Lanier, L.L.; Spies, T. Activation of NK cells and T cells by NKG2D, a receptor for stress-inducible MICA. Science 1999, 285, 727–729. [Google Scholar] [CrossRef]
- Diefenbach, A.; Jamieson, A.; Liu, S.D.; Shastri, N.; Raulet, D.H. Ligands for the murine NKG2D receptor: Expression by tumor cells and activation of NK cells and macrophages. Nat. Immunol. 2000, 1, 119–126. [Google Scholar] [CrossRef]
- Krmpotic, A.; Hasan, M.; Loewendorf, A.; Saulig, T.; Halenius, A.; Lenac, T.; Polić, B.; Bubic, I.; Kriegeskorte, A.; Pugel, E.P.; et al. NK cell activation through the NKG2D ligand MULT-1 is selectively prevented by the glycoprotein encoded by mouse cytomegalovirus gene m145. J. Exp. Med. 2005, 201, 211–220. [Google Scholar] [CrossRef] [Green Version]
- Cosman, D.; Müllberg, J.; Sutherland, C.L.; Chin, W.; Armitage, R.; Fanslow, W.; Kubin, M.; Chalupny, N. ULBPs, novel MHC class I-related molecules, bind to CMV glycoprotein UL16 and stimulate NK cytotoxicity through the NKG2D receptor. Immunity 2001, 14, 123–133. [Google Scholar] [CrossRef]
- Latchman, Y.; McKay, P.F.; Reiser, H. Identification of the 2B4 molecule as a counter-receptor for CD48. J. Immunol. 1998, 161, 5809–5812. [Google Scholar]
- Brown, M.H.; Boles, K.; Van Der Merwe, P.A.; Kumar, V.; Mathew, P.A.; Barclay, A.N. 2B4, the natural killer and T cell immunoglobulin superfamily surface protein, is a ligand for CD48. J. Exp. Med. 1998, 188, 2083–2090. [Google Scholar] [CrossRef]
- Anderson, P.; Caligiuri, M.; O’Brien, C.; Manley, T.; Ritz, J.; Schlossman, S.F. Fc gamma receptor type III (CD16) is included in the zeta NK receptor complex expressed by human natural killer cells. Proc. Natl. Acad. Sci. USA 1990, 87, 2274–2278. [Google Scholar] [CrossRef] [Green Version]
- Walzer, T.; Bléry, M.; Chaix, J.; Fuseri, N.; Chasson, L.; Robbins, S.H.; Jaeger, S.; André, P.; Gauthier, L.; Daniel, L.; et al. Identification, activation, and selective in vivo ablation of mouse NK cells via NKp46. Proc. Natl. Acad. Sci. USA 2007, 104, 3384–3389. [Google Scholar] [CrossRef] [Green Version]
- Ghirlando, R.; Keown, M.B.; Mackay, G.; Lewis, M.S.; Unkeless, J.C.; Gould, H.J. Stoichiometry and thermodynamics of the interaction between the Fc fragment of human IgG1 and its low-affinity receptor Fc gamma RIII. Biochemistry 1995, 34, 13320–13327. [Google Scholar] [CrossRef]
- Ravetch, J.V.; Perussia, B. Alternative membrane forms of Fc gamma RIII(CD16) on human natural killer cells and neutrophils. Cell type-specific expression of two genes that differ in single nucleotide substitutions. J. Exp. Med. 1989, 170, 481–497. [Google Scholar] [CrossRef]
- Lanier, L.L.; Yu, G.; Phillips, J.H. Co-association of CD3 zeta with a receptor (CD16) for IgG Fc on human natural killer cells. Nature 1989, 342, 803–805. [Google Scholar] [CrossRef]
- Hibbs, M.; Selvaraj, P.; Carpén, O.; Springer, T.; Küster, H.; Jouvin, M.; Kinet, J. Mechanisms for regulating expression of membrane isoforms of Fc gamma RIII (CD16). Science 1989, 246, 1608–1611. [Google Scholar] [CrossRef]
- Kurosaki, T.; Gander, I.; Ravetch, J.V. A subunit common to an IgG Fc receptor and the T-cell receptor mediates assembly through different interactions. Proc. Natl. Acad. Sci. USA 1991, 88, 3837–3841. [Google Scholar] [CrossRef] [Green Version]
- Romee, R.; Foley, B.; Lenvik, T.; Wang, Y.; Zhang, B.; Ankarlo, D.; Luo, X.; Cooley, S.; Verneris, M.; Walcheck, B.; et al. NK cell CD16 surface expression and function is regulated by a disintegrin and metalloprotease-17 (ADAM17). Blood 2013, 121, 3599–3608. [Google Scholar] [CrossRef]
- Peruzzi, G.; Femnou, L.; Gil-Krzewska, A.; Borrego, F.; Weck, J.; Krzewski, K.; Coligan, J.E. Membrane-type 6 matrix metalloproteinase regulates the activation-induced downmodulation of CD16 in human primary NK cells. J. Immunol. 2013, 191, 1883–1894. [Google Scholar] [CrossRef] [Green Version]
- Srpan, K.; Ambrose, A.; Karampatzakis, A.; Saeed, M.; Cartwright, A.N.; Guldevall, K.; Matos, G.D.S.C.D.; Önfelt, B.; Davis, D.M. Shedding of CD16 disassembles the NK cell immune synapse and boosts serial engagement of target cells. J. Cell Biol. 2018, 217, 3267–3283. [Google Scholar] [CrossRef] [Green Version]
- Baxter, D. Active and passive immunization for cancer. Hum. Vaccin Immunother. 2014, 10, 2123–2129. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.; Huang, X.; Ye, J.; Pan, P.; Cao, Q.; Yang, B.; Li, Z.; Su, M.; Huang, C.; Gu, J. Immunoglobulin G is present in a wide variety of soft tissue tumors and correlates well with proliferation markers and tumor grades. Cancer 2010, 116, 1953–1963. [Google Scholar] [CrossRef]
- Qiu, Y.; Korteweg, C.; Chen, Z.; Li, J.; Luo, J.; Huang, G.; Gu, J. Immunoglobulin G expression and its colocalization with complement proteins in papillary thyroid cancer. Mod. Pathol. 2012, 25, 36–45. [Google Scholar] [CrossRef]
- Rich, B.S.; Honeyman, J.N.; Darcy, D.G.; Smith, P.T.; Williams, A.R.; Lim, I.I.P.; Johnson, L.K.; Gonen, M.; Simon, J.S.; Laquaglia, M.P.; et al. Endogenous antibodies for tumor detection. Sci. Rep. 2014, 4, 5088. [Google Scholar] [CrossRef]
- Brostjan, C.; Sobanov, Y.; Glienke, J.; Hayer, S.; Lehrach, H.; Francis, F.; Hofer, E. The NKG2 natural killer cell receptor family: Comparative analysis of promoter sequences. Genes. Immun. 2000, 1, 504–508. [Google Scholar] [CrossRef] [Green Version]
- Carapito, R.; Bahram, S. Genetics, genomics, and evolutionary biology of NKG2D ligands. Immunol. Rev. 2015, 267, 88–116. [Google Scholar] [CrossRef]
- Diefenbach, A.; Tomasello, E.; Lucas, M.; Jamieson, A.; Hsia, J.K.; Vivier, E.; Raulet, D.H. Selective associations with signaling proteins determine stimulatory versus costimulatory activity of NKG2D. Nat. Immunol. 2002, 3, 1142–1149. [Google Scholar] [CrossRef]
- Garrity, D.; Call, M.; Feng, J.; Wucherpfennig, K.W. The activating NKG2D receptor assembles in the membrane with two signaling dimers into a hexameric structure. Proc. Natl. Acad. Sci. USA 2005, 102, 7641–7646. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gilfillan, S.; Ho, E.L.; Cella, M.; Yokoyama, W.M.; Colonna, M. NKG2D recruits two distinct adapters to trigger NK cell activation and costimulation. Nat. Immunol. 2002, 3, 1150–1155. [Google Scholar] [CrossRef] [PubMed]
- Rosen, D.B.; Araki, M.; Hamerman, J.A.; Chen, T.; Yamamura, T.; Lanier, L.L. A Structural basis for the association of DAP12 with mouse, but not human, NKG2D. J. Immunol. 2004, 173, 2470–2478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, S.; Shoelson, S.E.; Chaudhuri, M.; Gish, G.; Pawson, T.; Haser, W.G.; King, F.; Roberts, T.; Ratnofsky, S.; Lechleider, R.J.; et al. SH2 domains recognize specific phosphopeptide sequences. Cell 1993, 72, 767–778. [Google Scholar] [CrossRef]
- Lanier, L.L.; Corliss, B.C.; Wu, J.; Leong, C.; Phillips, J.H. Immunoreceptor DAP12 bearing a tyrosine-based activation motif is involved in activating NK cells. Nature 1998, 391, 703–707. [Google Scholar] [CrossRef]
- McGilvray, R.W.; Eagle, R.A.; Watson, N.F.; Al-Attar, A.; Ball, G.; Jafferji, I.; Trowsdale, J.; Durrant, L. NKG2D ligand expression in human colorectal cancer reveals associations with prognosis and evidence for immunoediting. Clin. Cancer Res. 2009, 15, 6993–7002. [Google Scholar] [CrossRef] [Green Version]
- Haberthur, K.; Brennan, K.; Hoglund, V.; Balcaitis, S.; Chinn, H.; Davis, A.; Kreuser, S.; Winter, C.; Leary, S.E.; Deutsch, G.; et al. NKG2D ligand expression in pediatric brain tumors. Cancer Biol. Ther. 2016, 17, 1253–1265. [Google Scholar] [CrossRef] [Green Version]
- Hilpert, J.; Grosse-Hovest, L.; Grünebach, F.; Buechele, C.; Nuebling, T.; Raum, T.; Steinle, A.; Salih, H.R.; Tamassia, N.; Bazzoni, F.; et al. Comprehensive analysis of NKG2D ligand expression and release in leukemia: Implications for NKG2D-mediated NK cell responses. J. Immunol. 2012, 189, 1360–1371. [Google Scholar] [CrossRef] [Green Version]
- Vyas, M.; Reinartz, S.; Hoffmann, N.; Reiners, K.S.; Lieber, S.; Jansen, J.M.; Wagner, U.; Müller, R.; Von Strandmann, E.P. Soluble NKG2D ligands in the ovarian cancer microenvironment are associated with an adverse clinical outcome and decreased memory effector T cells independent of NKG2D downregulation. Oncoimmunology 2017, 6, 1339854. [Google Scholar] [CrossRef] [Green Version]
- Cai, X.; Caballero-Benitez, A.; Gewe, M.M.; Jenkins, I.; Drescher, C.W.; Strong, R.K.; Spies, T.; Groh, V. Control of tumor initiation by NKG2D naturally expressed on ovarian cancer cells. Neoplasia 2017, 19, 471–482. [Google Scholar] [CrossRef]
- Groh, V.; Wu, J.; Yee, C.; Spies, T. Tumour-derived soluble MIC ligands impair expression of NKG2D and T-cell activation. Nature 2002, 419, 734–738. [Google Scholar] [CrossRef]
- Waldhauer, I.; Steinle, A. Proteolytic release of soluble UL16-binding protein 2 from tumor cells. Cancer Res. 2006, 66, 2520–2526. [Google Scholar] [CrossRef] [Green Version]
- Kraetzel, K.; Stoelcker, B.; Eissner, G.; Multhoff, G.; Pfeifer, M.; Holler, E.; Schulz, C. NKG2D-dependent effector function of bronchial epithelium-activated alloreactive T-cells. Eur. Respir. J. 2008, 32, 563–570. [Google Scholar] [CrossRef] [Green Version]
- Hue, S.; Mention, J.-J.; Monteiro, R.C.; Zhang, S.; Cellier, C.; Schmitz, J.; Verkarre, V.; Fodil, N.; Bahram, S.; Cerf-Bensussan, N.; et al. A direct role for NKG2D/MICA interaction in villous atrophy during celiac disease. Immunity 2004, 21, 367–377. [Google Scholar] [CrossRef] [Green Version]
- Weil, S.; Memmer, S.; Lechner, A.; Huppert, V.; Giannattasio, A.; Becker, T.; Müller-Runte, A.; Lampe, K.; Beutner, D.; Quaas, A.; et al. Natural killer group 2D ligand depletion reconstitutes natural killer cell immunosurveillance of head and neck squamous cell carcinoma. Front. Immunol. 2017, 8, 387. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Larrocha, P.S.-L.; Zhang, B.; Wainwright, D.; Dhar, P.; Wu, J.D. Antibody targeting tumor-derived soluble NKG2D ligand sMIC provides dual co-stimulation of CD8 T cells and enables sMIC(+) tumors respond to PD1/PD-L1 blockade therapy. J. Immunother. Cancer. 2019, 7, 223. [Google Scholar] [CrossRef] [Green Version]
- Sivori, S.; Vitale, M.; Morelli, L.; Sanseverino, L.; Augugliaro, R.; Bottino, C.; Moretta, L.; Moretta, A. P46, a novel natural killer cell-specific surface molecule that mediates cell activation. J. Exp. Med. 1997, 186, 1129–1136. [Google Scholar] [CrossRef] [Green Version]
- Pende, D.; Parolini, S.; Pessino, A.; Sivori, S.; Augugliaro, R.; Morelli, L.; Marcenaro, E.; Accame, L.; Malaspina, A.; Biassoni, R.; et al. Identification and molecular characterization of NKp30, a novel triggering receptor involved in natural cytotoxicity mediated by human natural killer cells. J. Exp. Med. 1999, 190, 1505–1516. [Google Scholar] [CrossRef]
- Vitale, M.; Bottino, C.; Sivori, S.; Sanseverino, L.; Castriconi, R.; Marcenaro, E.; Augugliaro, R.; Moretta, L.; Moretta, A. NKp44, a novel triggering surface molecule specifically expressed by activated natural killer cells, is involved in non-major histocompatibility complex-restricted tumor cell lysis. J. Exp. Med. 1998, 187, 2065–2072. [Google Scholar] [CrossRef] [PubMed]
- Sivori, S.; Pende, D.; Bottino, C.; Marcenaro, E.; Pessino, A.; Biassoni, R.; Moretta, L.; Moretta, A. NKp46 is the major triggering receptor involved in the natural cytotoxicity of fresh or cultured human NK cells. Correlation between surface density of NKp46 and natural cytotoxicity against autologous, allogeneic or xenogeneic target cells. Eur. J. Immunol. 1999, 29, 1656–1666. [Google Scholar] [CrossRef]
- Glasner, A.; Ghadially, H.; Gur, C.; Stanietsky, N.; Tsukerman, P.; Enk, J.; Mandelboim, O.; Inngjerdingen, M.; Kveberg, L.; Vaage, J.T. Recognition and prevention of tumor metastasis by the NK receptor NKp46/NCR1. J. Immunol. 2012, 188, 2509–2515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foster, C.E.; Colonna, M.; Sun, P.D. Crystal structure of the human natural killer (NK) cell activating receptor NKp46 reveals structural relationship to other leukocyte receptor complex immunoreceptors. J. Biol. Chem. 2003, 278, 46081–46086. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaron-Mendelson, M.; Yossef, R.; Appel, M.; Zilka, A.; Hadad, U.; Afergan, F.; Rosental, B.; Engel, S.; Nedvetzki, S.; Braiman, A.; et al. Dimerization of NKp46 receptor is essential for NKp46-mediated lysis: Characterization of the dimerization site by epitope mapping. J. Immunol. 2012, 188, 6165–6174. [Google Scholar] [CrossRef] [PubMed]
- Hadad, U.; Thauland, T.J.; Martinez, O.M.; Butte, M.J.; Porgador, A.; Krams, S.M. NKp46 Clusters at the immune synapse and regulates NK cell polarization. Front. Immunol. 2015, 6, 495. [Google Scholar] [CrossRef] [Green Version]
- Glasner, A.; Levi, A.; Enk, J.; Isaacson, B.; Viukov, S.; Orlanski, S.; Scope, A.; Neuman, T.; Enk, C.D.; Hanna, J.; et al. NKp46 Receptor-mediated interferon-gamma production by natural killer cells increases fibronectin 1 to alter tumor architecture and control metastasis. Immunity 2018, 48, 107–119. [Google Scholar] [CrossRef]
- Neville, M.J.; Campbell, R.D. A new member of the Ig superfamily and a V-ATPase G subunit are among the predicted products of novel genes close to the TNF locus in the human MHC. J. Immunol. 1999, 162, 4745–4754. [Google Scholar]
- Rusakiewicz, S.; Perier, A.; Semeraro, M.; Pitt, J.M.; Von Strandmann, E.P.; Reiners, K.S.; Aspeslagh, S.; Pipéroglou, C.; Vély, F.; Ivagnes, A.; et al. NKp30 isoforms and NKp30 ligands are predictive biomarkers of response to imatinib mesylate in metastatic GIST patients. Oncoimmunology 2017, 6, 1137418. [Google Scholar] [CrossRef]
- Cantoni, C.; Bottino, C.; Vitale, M.; Pessino, A.; Augugliaro, R.; Malaspina, A.; Parolini, S.; Moretta, L.; Moretta, A.; Biassoni, R. NKp44, a triggering receptor involved in tumor cell lysis by activated human natural killer cells, is a novel member of the immunoglobulin superfamily. J. Exp. Med. 1999, 189, 787–796. [Google Scholar] [CrossRef]
- Campbell, K.S.; Yusa, S.-I.; Kikuchi-Maki, A.; Catina, T.L. NKp44 triggers NK cell activation through DAP12 association that is not influenced by a putative cytoplasmic inhibitory sequence. J. Immunol. 2004, 172, 899–906. [Google Scholar] [CrossRef] [Green Version]
- Byrd, A.; Hoffmann, S.C.; Jarahian, M.; Momburg, F.; Watzl, C. Expression analysis of the ligands for the Natural Killer cell receptors NKp30 and NKp44. PLoS ONE 2007, 2, 1339. [Google Scholar] [CrossRef]
- Cantoni, C.; Ponassi, M.; Biassoni, R.; Conte, R.; Spallarossa, A.; Moretta, A.; Moretta, L.; Bolognesi, M.; Bordo, D. The three-dimensional structure of the human NK cell receptor NKp44, a triggering partner in natural cytotoxicity. Structure 2003, 11, 725–734. [Google Scholar] [CrossRef] [Green Version]
- Chan, C.J.; Martinet, L.; Gilfillan, S.; Souza-Fonseca-Guimaraes, F.; Chow, M.T.; Town, L.; Ritchie, D.S.; Colonna, M.; Andrews, D.; Smyth, M.J. The receptors CD96 and CD226 oppose each other in the regulation of natural killer cell functions. Nat. Immunol. 2014, 15, 431–438. [Google Scholar] [CrossRef] [PubMed]
- Guillerey, C.; De Andrade, L.F.; Vuckovic, S.; Miles, K.; Ngiow, S.F.; Yong, M.C.; Teng, M.W.; Colonna, M.; Ritchie, D.S.; Chesi, M.; et al. Immunosurveillance and therapy of multiple myeloma are CD226 dependent. J. Clin. Investig. 2015, 125, 2077–2089. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tahara-Hanaoka, S.; Shibuya, K.; Kai, H.; Miyamoto, A.; Morikawa, Y.; Ohkochi, N.; Honda, S.-I.; Shibuya, A. Tumor rejection by the poliovirus receptor family ligands of the DNAM-1 (CD226) receptor. Blood 2006, 107, 1491–1496. [Google Scholar] [CrossRef] [Green Version]
- Iguchi-Manaka, A.; Kai, H.; Yamashita, Y.; Shibata, K.; Tahara-Hanaoka, S.; Honda, S.-I.; Yasui, T.; Kikutani, H.; Shibuya, K.; Shibuya, A. Accelerated tumor growth in mice deficient in DNAM-1 receptor. J. Exp. Med. 2008, 205, 2959–2964. [Google Scholar] [CrossRef] [Green Version]
- Smyth, M.J.; Crowe, N.Y.; Godfrey, D.I. NK cells and NKT cells collaborate in host protection from methylcholanthrene-induced fibrosarcoma. Int. Immunol. 2001, 13, 459–463. [Google Scholar] [CrossRef] [Green Version]
- Chan, C.J.; Andrews, D.; McLaughlin, N.M.; Yagita, H.; Gilfillan, S.; Colonna, M.; Smyth, M.J. DNAM-1/CD155 interactions promote cytokine and NK cell-mediated suppression of poorly immunogenic melanoma metastases. J. Immunol. 2010, 184, 902–911. [Google Scholar] [CrossRef]
- Gilfillan, S.; Chan, C.J.; Cella, M.; Haynes, N.M.; Rapaport, A.S.; Boles, K.S.; Andrews, D.; Smyth, M.J.; Colonna, M. DNAM-1 promotes activation of cytotoxic lymphocytes by nonprofessional antigen-presenting cells and tumors. J. Exp. Med. 2008, 205, 2965–2973. [Google Scholar] [CrossRef] [Green Version]
- Masson, D.; Jarry, A.; Baury, B.; Blanchardie, P.; Laboisse, C.; Lustenberger, P.; Denis, M. Overexpression of the CD155 gene in human colorectal carcinoma. Gut 2001, 49, 236–240. [Google Scholar] [CrossRef] [Green Version]
- Nakai, R.; Maniwa, Y.; Tanaka, Y.; Nishio, W.; Yoshimura, M.; Okita, Y.; Ohbayashi, C.; Satoh, N.; Ogita, H.; Takai, Y.; et al. Overexpression of Necl-5 correlates with unfavorable prognosis in patients with lung adenocarcinoma. Cancer Sci. 2010, 101, 1326–1330. [Google Scholar] [CrossRef]
- Bevelacqua, V.; Bevelacqua, Y.; Candido, S.; Skarmoutsou, E.; Amoroso, A.; Guarneri, C.; Strazzanti, A.; Gangemi, P.; Mazzarino, M.C.; D’Amico, F.; et al. Nectin like-5 overexpression correlates with the malignant phenotype in cutaneous melanoma. Oncotarget 2012, 3, 882–892. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishiwada, S.; Sho, M.; Yasuda, S.; Shimada, K.; Yamato, I.; Akahori, T.; Kinoshita, S.; Nagai, M.; Konishi, N.; Nakajima, Y. Clinical significance of CD155 expression in human pancreatic cancer. Anticancer Res. 2015, 35, 2287–2297. [Google Scholar] [PubMed]
- Sloan, K.E.; Eustace, B.K.; Stewart, J.K.; Zehetmeier, C.; Torella, C.; Simeone, M.E.; Roy, J.; Unger, C.; Louis, D.N.; Ilag, L.L.; et al. D155/PVR plays a key role in cell motility during tumor cell invasion and migration. BMC Cancer 2004, 4, 73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lakshmikanth, T.; Burke, S.; Ali, T.H.; Kimpfler, S.; Ursini, F.; Ruggeri, L.; Capanni, M.; Umansky, V.; Paschen, A.; Sucker, A.; et al. NCRs and DNAM-1 mediate NK cell recognition and lysis of human and mouse melanoma cell lines in vitro and in vivo. J. Clin. Investig. 2009, 119, 1251–1263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castriconi, R.; Griffin, K.J.; Kirschner, L.S.; Matyakhina, L.; Stergiopoulos, S.; Robinson-White, A.; Lenherr, S.; Weinberg, F.D.; Claflin, E.; Meoli, E.; et al. Natural killer cell-mediated killing of freshly isolated neuroblastoma cells: Critical role of DNAX accessory molecule-1-poliovirus receptor interaction. Cancer Res. 2004, 64, 9180–9184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bottino, C.; Castriconi, R.; Pende, D.; Rivera, P.; Nanni, M.; Carnemolla, B.; Cantoni, C.; Grassi, J.; Marcenaro, S.; Reymond, N.; et al. Identification of PVR (CD155) and Nectin-2 (CD112) as cell surface ligands for the human DNAM-1 (CD226) activating molecule. J. Exp. Med. 2003, 198, 557–567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Andrade, L.F.; Smyth, M.J.; Martinet, L. DNAM-1 control of natural killer cells functions through nectin and nectin-like proteins. Immunol. Cell Biol. 2014, 92, 237–244. [Google Scholar] [CrossRef]
- Bottino, C.; Falco, M.; Parolini, S.; Marcenaro, E.; Augugliaro, R.; Sivori, S.; Landi, E.; Biassoni, R.; Notarangelo, L.D.; Moretta, L.; et al. NTB-A [correction of GNTB-A], a novel SH2D1A-associated surface molecule contributing to the inability of natural killer cells to kill Epstein-Barr virus-infected B cells in X-linked lymphoproliferative disease. J. Exp. Med. 2001, 194, 235–246. [Google Scholar] [CrossRef]
- Garni-Wagner, B.A.; Purohit, A.A.; Mathew, P.; Bennett, M.; Kumar, V. A novel function-associated molecule related to non-MHC-restricted cytotoxicity mediated by activated natural killer cells and T cells. J. Immunol. 1993, 151, 60–70. [Google Scholar]
- Claus, M.; Wingert, S.; Watzl, C. Modulation of natural killer cell functions by interactions between 2B4 and CD48 in cis and in trans. Open Biol. 2016, 6. [Google Scholar] [CrossRef] [Green Version]
- Altvater, B.; Landmeier, S.; Pscherer, S.; Temme, J.; Schweer, K.; Kailayangiri, S.; Campana, D.; Juergens, H.; Pule, M.A.; Rossig, C. 2B4 (CD244) signaling by recombinant antigen-specific chimeric receptors costimulates natural killer cell activation to leukemia and neuroblastoma cells. Clin. Cancer Res. 2009, 15, 4857–4866. [Google Scholar] [CrossRef] [Green Version]
- Cruz-Munoz, M.-E.; Dong, Z.; Shi, X.; Zhang, S.; Veillette, A. Influence of CRACC, a SLAM family receptor coupled to the adaptor EAT-2, on natural killer cell function. Nat. Immunol. 2009, 10, 297–305. [Google Scholar] [CrossRef]
- Dong, Z.; Cruz-Munoz, M.-E.; Zhong, M.-C.; Chen, R.; Latour, S.; Veillette, A. Essential function for SAP family adaptors in the surveillance of hematopoietic cells by natural killer cells. Nat. Immunol. 2009, 10, 973–980. [Google Scholar] [CrossRef]
- Jakubowiak, A.J.; Benson, D.M.; Bensinger, W.; Siegel, D.S.; Zimmerman, T.M.; Mohrbacher, A.; Richardson, P.G.; Afar, D.E.; Singhal, A.K.; Anderson, K.C. Phase I trial of anti-CS1 monoclonal antibody elotuzumab in combination with bortezomib in the treatment of relapsed/refractory multiple myeloma. J. Clin. Oncol. 2012, 30, 1960–1965. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lonial, S.; Vij, R.; Harousseau, J.-L.; Facon, T.; Moreau, P.; Mazumder, A.; Kaufman, J.L.; Leleu, X.; Tsao, L.C.; Westland, C.; et al. Elotuzumab in combination with lenalidomide and low-dose dexamethasone in relapsed or refractory multiple myeloma. J. Clin. Oncol. 2012, 30, 1953–1959. [Google Scholar] [CrossRef] [PubMed]
- Collins, S.M.; Bakan, C.E.; Swartzel, G.D.; Hofmeister, C.C.; Efebera, Y.A.; Kwon, H.; Starling, G.C.; Ciarlariello, D.; Bhaskar, S.; Briercheck, E.L.; et al. Elotuzumab directly enhances NK cell cytotoxicity against myeloma via CS1 ligation: Evidence for augmented NK cell function complementing ADCC. Cancer Immunol. Immunother. 2013, 62, 1841–1849. [Google Scholar] [CrossRef] [Green Version]
- Ernst, D.; Williams, B.A.; Wang, X.-H.; Yoon, N.; Kim, K.-P.; Chiu, J.; Luo, Z.J.; Hermans, K.G.; Krueger, J.; Keating, A. Humanized anti-CD123 antibody facilitates NK cell antibody-dependent cell-mediated cytotoxicity (ADCC) of Hodgkin lymphoma targets via ARF6/PLD-1. Blood Cancer, J. 2019, 9, 6. [Google Scholar] [CrossRef] [PubMed]
- Xiao, L.; Cen, D.; Gan, H.; Sun, Y.; Huang, N.; Xiong, H.; Jin, Q.; Su, L.; Liu, X.; Wang, K.; et al. Adoptive transfer of NKG2D CAR mRNA-engineered natural killer cells in colorectal cancer patients. Mol. Ther. 2019, 27, 1114–1125. [Google Scholar] [CrossRef] [Green Version]
- Deng, X.; Gao, F.; Li, N.; Li, Q.; Zhou, Y.; Yang, T.; Cai, Z.; Du, P.; Chen, F.; Cai, J. Antitumor activity of NKG2D CAR-T cells against human colorectal cancer cells in vitro and in vivo. Am. J. Cancer Res. 2019, 9, 945–958. [Google Scholar]
- Fernández, L.; Fernández, A.; Mirones, I.; Escudero, A.; Cardoso, L.; Vela, M.; Lanzarot, D.; De Paz, R.; Leivas, A.; Gallardo, M.; et al. GMP-compliant manufacturing of NKG2D CAR memory T cells using CliniMACS prodigy. Front. Immunol. 2019, 10, 2361. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.; Cherwinski, H.; Spies, T.; Phillips, J.H.; Lanier, L.L. DAP10 and DAP12 form distinct, but functionally cooperative, receptor complexes in natural killer cells. J. Exp. Med. 2000, 192, 1059–1068. [Google Scholar] [CrossRef]
- Brumbaugh, K.M.; Binstadt, B.; Billadeau, D.D.; Schoon, R.A.; Dick, C.J.; Ten, R.M.; Leibson, P.J. Functional role for Syk tyrosine kinase in natural killer cell-mediated natural cytotoxicity. J. Exp. Med. 1997, 186, 1965–1974. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, K.; Zhong, B.; Gilvary, D.L.; Corliss, B.C.; Vivier, E.; Hong-Geller, E.; Wei, S.; Djeu, J.Y. Syk regulation of phosphoinositide 3-kinase-dependent NK cell function. J. Immunol. 2002, 168, 3155–3164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Billadeau, D.D.; Brumbaugh, K.M.; Dick, C.J.; Schoon, R.A.; Bustelo, X.R.; Leibson, P.J. The Vav-Rac1 pathway in cytotoxic lymphocytes regulates the generation of cell-mediated killing. J. Exp. Med. 1998, 188, 549–559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ting, A.T.; Karnitz, L.M.A.; Schoon, R.; Abraham, R.T.; Leibson, P.J. Fc gamma receptor activation induces the tyrosine phosphorylation of both phospholipase C (PLC)-gamma 1 and PLC-gamma 2 in natural killer cells. J. Exp. Med. 1992, 176, 1751–1755. [Google Scholar] [CrossRef] [Green Version]
- Micucci, F.; Capuano, C.; Marchetti, E.; Piccoli, M.; Frati, L.; Santoni, A.; Galandrini, R. PI5KI-dependent signals are critical regulators of the cytolytic secretory pathway. Blood 2008, 111, 4165–4172. [Google Scholar] [CrossRef] [PubMed]
- Galandrini, R.; Micucci, F.; Tassi, I.; Cifone, M.G.; Cinque, B.; Piccoli, M.; Frati, L.; Santoni, A. Arf6: A new player in FcgammaRIIIA lymphocyte-mediated cytotoxicity. Blood 2005, 106, 577–583. [Google Scholar] [CrossRef] [Green Version]
- Bryceson, Y.T.; March, M.; Ljunggren, H.-G.; Long, E.O. Synergy among receptors on resting NK cells for the activation of natural cytotoxicity and cytokine secretion. Blood 2006, 107, 159–166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matalon, O.; Fried, S.; Ben-Shmuel, A.; Pauker, M.H.; Joseph, N.; Keizer, D.; Piterburg, M.; Barda-Saad, M. Dephosphorylation of the adaptor LAT and phospholipase C-gamma by SHP-1 inhibits natural killer cell cytotoxicity. Sci. Signal 2016, 9, 54. [Google Scholar] [CrossRef]
- Wu, J.; Song, Y.; Bakker, A.B.H.; Bauer, S.; Spies, T.; Lanier, L.L.; Phillips, J.H. An activating immunoreceptor complex formed by NKG2D and DAP10. Science 1999, 285, 730–732. [Google Scholar] [CrossRef]
- Chang, C.; Dietrich, J.; Harpur, A.G.A.; Lindquist, J.; Haude, A.; Loke, Y.W.; King, A.; Colonna, M.; Trowsdale, J.; Wilson, M.J. Cutting edge: KAP10, a novel transmembrane adapter protein genetically linked to DAP12 but with unique signaling properties. J. Immunol. 1999, 163, 4651–4654. [Google Scholar]
- Upshaw, J.L.; Arneson, L.N.A.; Schoon, R.; Dick, C.J.; Billadeau, D.D.; Leibson, P.J. NKG2D-mediated signaling requires a DAP10-bound Grb2-Vav1 intermediate and phosphatidylinositol-3-kinase in human natural killer cells. Nat. Immunol. 2006, 7, 524–532. [Google Scholar] [CrossRef]
- Billadeau, D.D.; Upshaw, J.L.A.; Schoon, R.; Dick, C.J.; Leibson, P.J. NKG2D-DAP10 triggers human NK cell-mediated killing via a Syk-independent regulatory pathway. Nat. Immunol. 2003, 4, 557–564. [Google Scholar] [CrossRef] [PubMed]
- Zompi, S.; Hamerman, J.A.; Ogasawara, K.; Schweighoffer, E.; Tybulewicz, V.L.; Di Santo, J.P.; Lanier, L.L.; Colucci, F. NKG2D triggers cytotoxicity in mouse NK cells lacking DAP12 or Syk family kinases. Nat. Immunol. 2003, 4, 565–572. [Google Scholar] [CrossRef]
- Graham, D.B.; Cella, M.; Giurisato, E.; Fujikawa, K.; Miletic, A.V.; Kloeppel, T.; Brim, K.; Takai, T.; Shaw, A.S.; Colonna, M.; et al. Vav1 controls DAP10-mediated natural cytotoxicity by regulating actin and microtubule dynamics. J. Immunol. 2006, 177, 2349–2355. [Google Scholar] [CrossRef] [PubMed]
- Shibuya, K.; Lanier, L.L.; Phillips, J.H.; Ochs, H.D.; Shimizu, K.; Nakayama, E.; Nakauchi, H.; Shibuya, A. Physical and functional association of LFA-1 with DNAM-1 adhesion molecule. Immunity 1999, 11, 615–623. [Google Scholar] [CrossRef] [Green Version]
- Sidorenko, S.P.; Clark, E.A. The dual-function CD150 receptor subfamily: The viral attraction. Nat. Immunol. 2003, 4, 19–24. [Google Scholar] [CrossRef]
- Dong, Z.; Davidson, M.; Quintero, L.A.P.; Kurosaki, T.; Swat, W.; Veillette, A. The adaptor SAP controls NK cell activation by regulating the enzymes Vav-1 and SHIP-1 and by enhancing conjugates with target cells. Immunity 2012, 36, 974–985. [Google Scholar] [CrossRef] [Green Version]
- Watzl, C.; Stebbins, C.C.; Long, E.O. NK cell inhibitory receptors prevent tyrosine phosphorylation of the activation receptor 2B4 (CD244). J. Immunol. 2000, 165, 3545–3548. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Quintero, L.A.; Roncagalli, R.; Guo, H.; Latour, S.; Davidson, D.; Veillette, A. EAT-2, a SAP-like adaptor, controls NK cell activation through phospholipase Cgamma, Ca++, and Erk, leading to granule polarization. J. Exp. Med. 2014, 211, 727–742. [Google Scholar] [CrossRef] [Green Version]
- Saborit-Villarroya, I.; Barriocanal, Águeda, M.; Oliver-Vila, I.; Engel, P.; Sayós, J.; Martin, M. The adaptor 3BP2 activates CD244-mediated cytotoxicity in PKC- and SAP-dependent mechanisms. Mol. Immunol. 2008, 45, 3446–3453. [Google Scholar] [CrossRef]
- Saborit-Villarroya, I.; Del Valle, J.M.; Romero, X.; Esplugues, E.; Lauzurica, P.; Engel, P.; Martin, M. The adaptor protein 3BP2 binds human CD244 and links this receptor to Vav signaling, ERK activation, and NK cell killing. J. Immunol. 2005, 17, 4226–4235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jevremovic, D.; Billadeau, D.D.; Schoon, R.A.; Dick, C.J.; Leibson, P.J. Regulation of NK cell-mediated cytotoxicity by the adaptor protein 3BP2. J. Immunol. 2001, 166, 7219–7228. [Google Scholar] [CrossRef] [Green Version]
- Valdez, P.A.; Wang, H.; Seshasayee, D.; Campagne, M.V.L.; Gurney, A.; Lee, W.P.; Grewal, I.S. NTB-A, a new activating receptor in T cells that regulates autoimmune disease. J. Biol. Chem. 2004, 279, 18662–18669. [Google Scholar] [CrossRef] [Green Version]
- Eissmann, P.; Watzl, C. Molecular analysis of NTB-A signaling: A role for EAT-2 in NTB-A-mediated activation of human NK cells. J. Immunol. 2006, 177, 3170–3177. [Google Scholar] [CrossRef] [Green Version]
- Tassi, I.; Colonna, M. The cytotoxicity receptor CRACC (CS-1) recruits EAT-2 and activates the PI3K and phospholipase cgamma signaling pathways in human NK cells. J. Immunol. 2005, 175, 7996–8002. [Google Scholar] [CrossRef] [PubMed]
- Bouchon, A.; Cella, M.; Grierson, H.L.; Cohen, J.I.; Colonna, M. Cutting Edge: Activation of NK cell-mediated cytotoxicity by a SAP-independent receptor of the CD2 family. J. Immunol. 2001, 167, 5517–5521. [Google Scholar] [CrossRef] [Green Version]
- Eissmann, P.; Beauchamp, L.; Wooters, J.; Tilton, J.C.; Long, E.O.; Watzl, C. Molecular basis for positive and negative signaling by the natural killer cell receptor 2B4 (CD244). Blood 2005, 105, 4722–4729. [Google Scholar] [CrossRef] [Green Version]
- Tangye, S.G.; Lazetic, S.; Woollatt, E.; Sutherland, G.R.; Lanier, L.L.; Phillips, J.H. Cutting edge: Human 2B4, an activating NK cell receptor, recruits the protein tyrosine phosphatase SHP-2 and the adaptor signaling protein SAP. J. Immunol. 1999, 162, 6981–6985. [Google Scholar]
- Coffey, A.J.; Brooksbank, R.A.; Brandau, O.; Oohashi, T.; Howell, G.R.; Bye, J.M.; Cahn, A.P.; Durham, J.; Heath, P.; Wray, P.; et al. Host response to EBV infection in X-linked lymphoproliferative disease results from mutations in an SH2-domain encoding gene. Nat. Genet. 1998, 20, 129–135. [Google Scholar] [CrossRef]
- Nichols, K.E.; Harkin, D.P.; Levitz, S.; Krainer, M.; Kolquist, K.A.; Genovese, C.; Bernard, A.; Ferguson, M.; Zuo, L.; Snyder, E.; et al. Inactivating mutations in an SH2 domain-encoding gene in X-linked lymphoproliferative syndrome. Proc. Natl. Acad. Sci. USA 1998, 95, 13765–13770. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsu, H.-T.; Mace, E.M.; Carisey, A.F.; Viswanath, D.I.; Christakou, A.E.; Wiklund, M.; Önfelt, B.; Orange, J.S. NK cells converge lytic granules to promote cytotoxicity and prevent bystander killing. J. Cell Biol. 2016, 215, 875–889. [Google Scholar] [CrossRef]
- Li, C.; Ge, B.; Nicotra, M.L.; Stern, J.N.; Kopcow, H.D.; Chen, X.; Strominger, J.L. JNK MAP kinase activation is required for MTOC and granule polarization in NKG2D-mediated NK cell cytotoxicity. Proc. Natl. Acad. Sci. USA 2008, 105, 3017–3022. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mentlik, A.N.; Sanborn, K.B.; Holzbaur, E.L.; Orange, J.S. Rapid lytic granule convergence to the MTOC in natural killer cells is dependent on dynein but not cytolytic commitment. Mol. Biol. Cell. 2010, 21, 2241–2256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bryceson, Y.T.; March, M.; Ljunggren, H.-G.; Long, E.O. Activation, coactivation, and costimulation of resting human natural killer cells. Immunol. Rev. 2006, 214, 73–91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rezvani, K. Adoptive cell therapy using engineered natural killer cells. Bone Marrow Transplant 2019, 54, 785–788. [Google Scholar] [CrossRef]
- Lupo, K.B.; Matosevic, S. Natural killer cells as allogeneic effectors in adoptive cancer immunotherapy. Cancers 2019, 11, 769. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Immunological synapse (IS) of Natural Killer (NK) cell and target cell. (A) NK cells engage other cells via integrins and adhesion molecules, which create the immunological synapse (IS)—the subsequent process between engagement and killing or tolerance can be broken down into four steps. (B) First, filamentous actin (F-actin) is recruited to the IS. Inside-out signaling reinforces IS interactions and activating and inhibitory surface receptors cluster at the IS. (C) Second, NK lytic granules move along microtubules by dynein-dynactin motor proteins toward the microtubule-organizing center (MTOC). (D) Third, the polarized lytic granules and MTOC travel in an ATP-dependent manner through the actin mesh via myosin IIA to dock at the IS, and finally, the lytic granules fuse with the membrane and release the lytic contents into the target cells, a process also known as degranulation. The NK cell then detaches and moves on to the next target.
Figure 1.
Immunological synapse (IS) of Natural Killer (NK) cell and target cell. (A) NK cells engage other cells via integrins and adhesion molecules, which create the immunological synapse (IS)—the subsequent process between engagement and killing or tolerance can be broken down into four steps. (B) First, filamentous actin (F-actin) is recruited to the IS. Inside-out signaling reinforces IS interactions and activating and inhibitory surface receptors cluster at the IS. (C) Second, NK lytic granules move along microtubules by dynein-dynactin motor proteins toward the microtubule-organizing center (MTOC). (D) Third, the polarized lytic granules and MTOC travel in an ATP-dependent manner through the actin mesh via myosin IIA to dock at the IS, and finally, the lytic granules fuse with the membrane and release the lytic contents into the target cells, a process also known as degranulation. The NK cell then detaches and moves on to the next target.

Figure 2.
NK cell surface receptors involved in tumor recognition. NK cells express a myriad of inhibitory and activating receptors designed to recognize healthy or aberrant (non-healthy) cells. (A) Inhibitory receptors dampen activating NK cell signals via cytoplasmic tyrosine motifs in their cytoplasmic tails, regulating NK cell effector function. (B) In contrast, most activating receptors signal through cytoplasmic adaptor proteins. Although many of the receptors are expressed by both mouse and human NK cells (shared), some are exclusive.
Figure 2.
NK cell surface receptors involved in tumor recognition. NK cells express a myriad of inhibitory and activating receptors designed to recognize healthy or aberrant (non-healthy) cells. (A) Inhibitory receptors dampen activating NK cell signals via cytoplasmic tyrosine motifs in their cytoplasmic tails, regulating NK cell effector function. (B) In contrast, most activating receptors signal through cytoplasmic adaptor proteins. Although many of the receptors are expressed by both mouse and human NK cells (shared), some are exclusive.

Figure 3.
Inhibitory and activating NK cell signaling. NK cell effector function is collectively determined by the strongest activating or inhibitory signals. (A) Signaling downstream of inhibitory receptors is initiated by ligand engagement, followed by tyrosine phosphorylation of the signaling motif by Src-family kinases (SFK), Fyn or Lck. Once phosphorylated, there are three known inhibitory pathways: (1) Recruitment of SHP-1, SHP-2 or SHIP, which dephosphorylate Vav1; (2) Association with c-Abl kinase, which phosphorylates Crk disassociating it from its active complex and; (3) Phosphorylation and activation of Cbl-b by the TAM receptors. Cbl-b in turn ubiquitylates activating signaling intermediates such as LAT1 for degradation. (B) Signaling downstream of activating receptors is similarly transduced by tyrosine containing motifs that are phosphorylated by SFK. Various signaling intermediates such as Grb2, VAV1, or PI3K are then recruited, which induce cytotoxicity and cytokine release.
Figure 3.
Inhibitory and activating NK cell signaling. NK cell effector function is collectively determined by the strongest activating or inhibitory signals. (A) Signaling downstream of inhibitory receptors is initiated by ligand engagement, followed by tyrosine phosphorylation of the signaling motif by Src-family kinases (SFK), Fyn or Lck. Once phosphorylated, there are three known inhibitory pathways: (1) Recruitment of SHP-1, SHP-2 or SHIP, which dephosphorylate Vav1; (2) Association with c-Abl kinase, which phosphorylates Crk disassociating it from its active complex and; (3) Phosphorylation and activation of Cbl-b by the TAM receptors. Cbl-b in turn ubiquitylates activating signaling intermediates such as LAT1 for degradation. (B) Signaling downstream of activating receptors is similarly transduced by tyrosine containing motifs that are phosphorylated by SFK. Various signaling intermediates such as Grb2, VAV1, or PI3K are then recruited, which induce cytotoxicity and cytokine release.


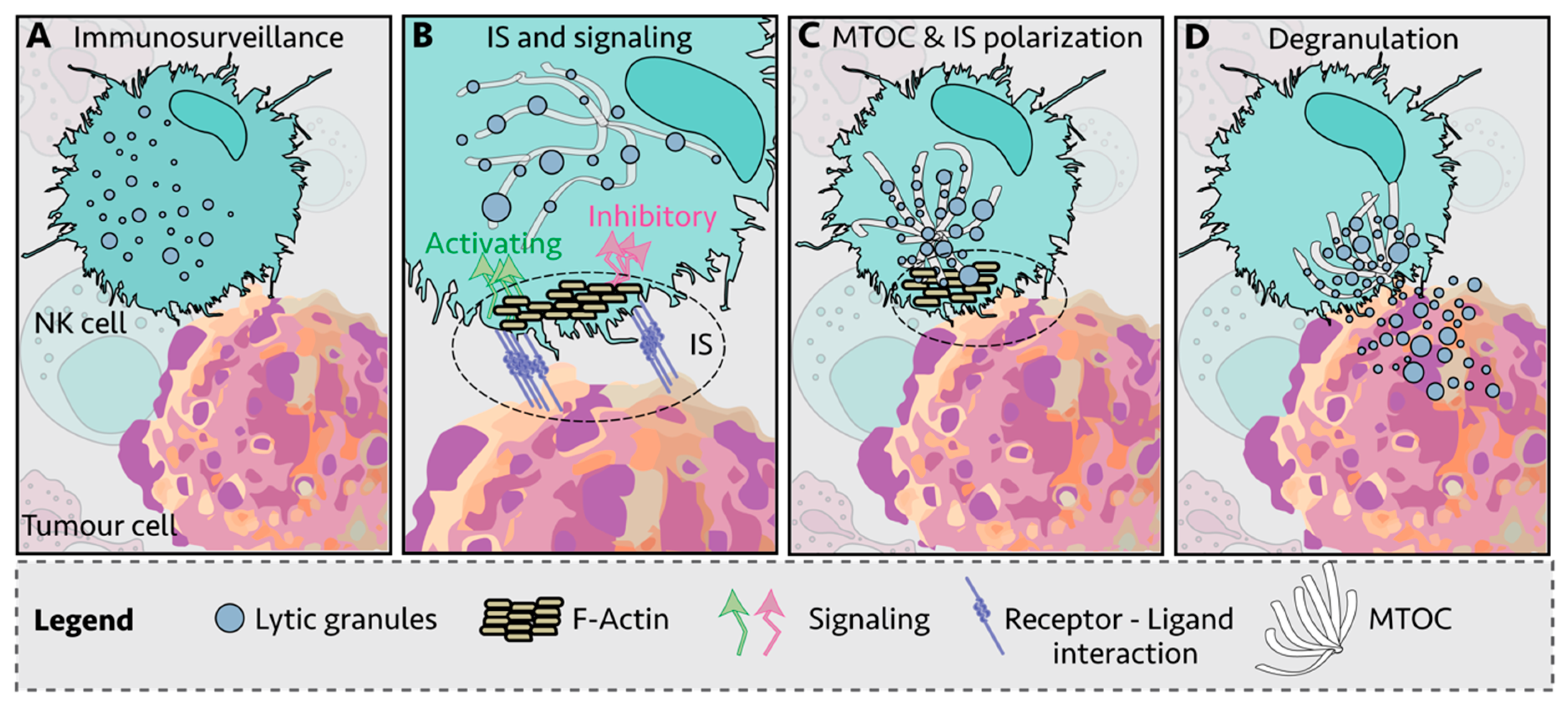{kind=link}
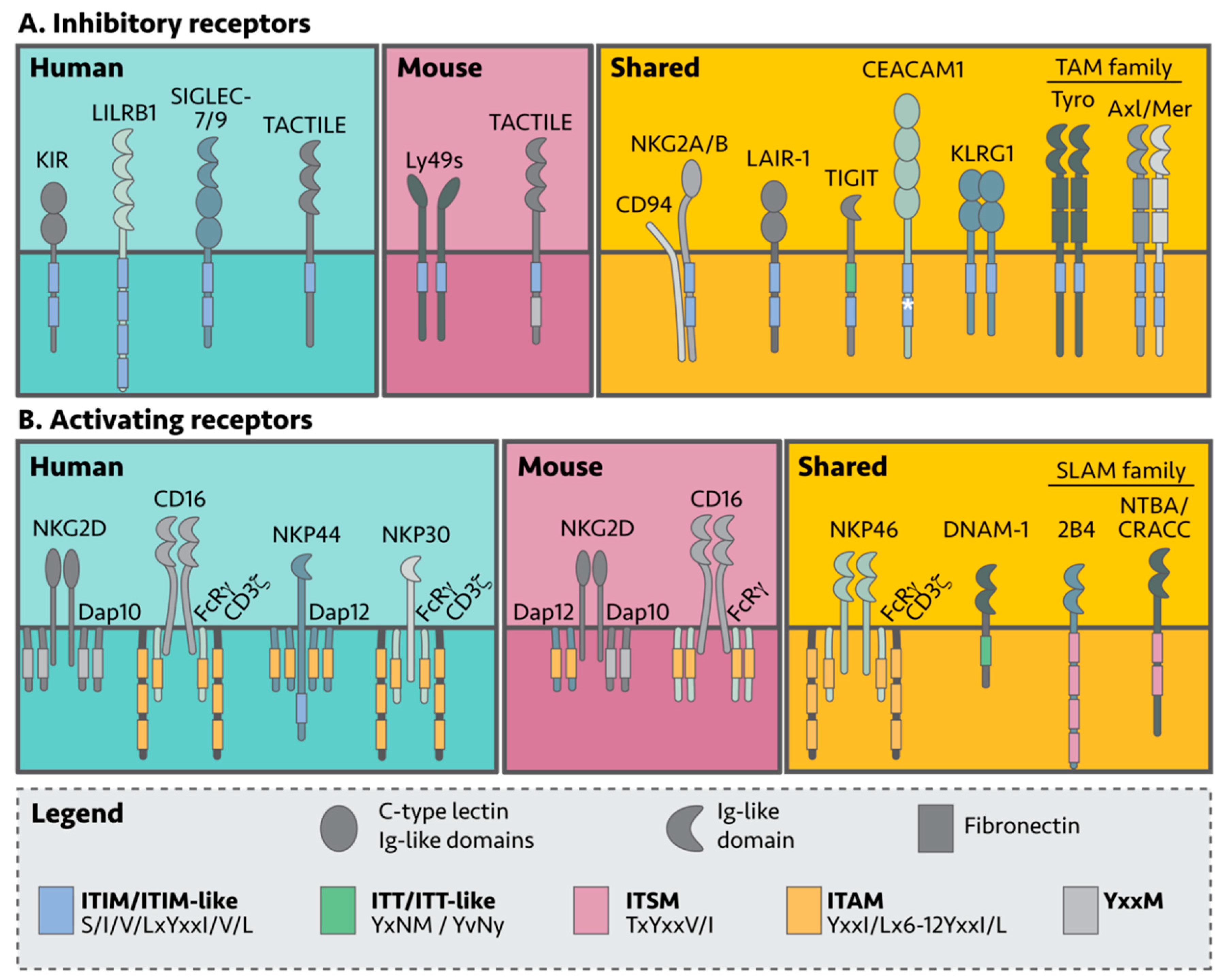{kind=link}
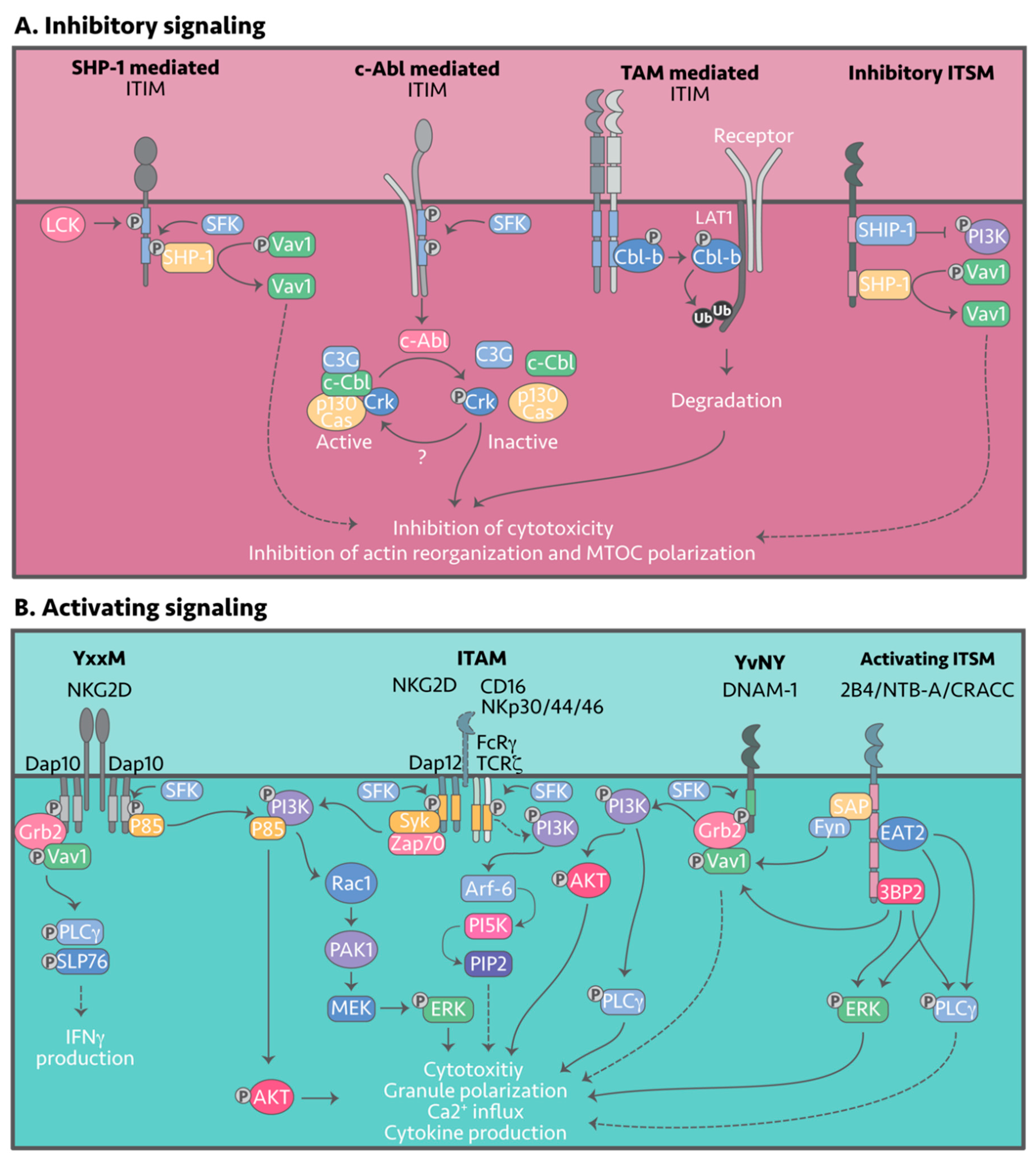{kind=link}
Table 1.
Consensus sequence of signaling motifs.
| Signaling Motifs | Consensus Sequence |
|---|---|
| ITIM | S/I/V/LxYxxI/V/L |
| ITT/ITT-Like | YxNM/YvNy |
| ITSM | TxYxxV/I |
| ITAM | YxxI/Lx6-12YxxI/L |
| Human Receptor | Classical MHC Ligands |
| KIR2DL1 | HLA-C group 2 molecules (Asn77 and Lys80) [45] |
| KIR2DL2 | HLA-B and HLA-C group 1 molecules (Ser77 and Asn80) [45] |
| KIR2DL3 | HLA-C group 1 molecules (Ser77 and Asn80) [45] and some HLA-CGroup 2 and HLA-B (weaker affinity than 2DL2) [46] |
| KIR2DL5(A+B) | unknown |
| KIR3DL1 | HLA-A and B with the Bw4 epitope, amino acid positions 77–83 [45] |
| KIR3DL2 | Some HLA-A allotypes [45] |
| KIR3DL3 | Unknown |
| LILRB1 | HLA-C [48,49] |
| Human receptor | Non-classical MHC ligands |
| CD94/NKG2A/B | HLA-E [59,60] |
| LILRB1 | HLA-E and HLA-G [61] and UL18 (MHC viral homologue) [62,63] |
| Human receptor | Non-MHC-I ligands |
| SIGLEG-7/9 | α2,3- and α2,6-linked sialylated proteins [64,65,66] |
| NKRP1A | LLT1 (CLEC2D) [67,68] |
| Mouse | Classical MHC ligands |
| Ly49A | H2-Dd [11,69,70,71,72,73,74], H2-Dk [74], H2-Ld, H2-Kb, H2-Kb and H2-Dp [69] |
| Ly49B | Unknown |
| Ly49C | H2-Kb [74,75], H2-Kd [74], H2-Kk, H2-Db, H2-Dd [76] |
| Ly49F | H2-Dd [76] |
| Ly49G | H2-Db [69] |
| Ly49I | H2-Kb [75] and H2-Kd [74] |
| Human receptor | Non-classical MHC ligands |
| Ly49A | H2-M3 [77] |
| Ly49C | H2-Q10 [78] |
| CD94/NKG2A/B | Qa-1 [59,60] |
| Human receptor | Non-MHC-I ligands |
| NKRP1B | Clr-b (Clec2d) [79] |
| gp49B1 | integrin αVβ3 [80] |
| Ly49E | urokinase plasminogen activator (uPA) [81] |
| Shared receptors | Non-MHC-I ligands |
| TIGIT | poliovirus receptor (CD155, PVR) and nectin-2 (CD112, PVRL2) [82,83] |
| TACTILE | poliovirus receptor (CD155, PVR) and nectin-2 (CD112, PVRL2) [82,83] |
| CEACAM1 | CEACAM1 [84] |
| LAIR-1 | Collagen [85] |
| KLRG1 | E- and N-Cadherin [86,87] |
| TAMs | Gas6 and Pros1 [88,89] |
Table 3.
Inhibitory NK cell receptors and their signaling motifs.
| Human | Form | Signaling Motif 1 | pY 2 Position | Reference | |
| KIR2DL1 | Monomer | 2× ITIM | VTYTQL | Y302 | [44] |
| IVYTEL | Y332 | ||||
| KIR2DL2 | Monomer | 2× ITIM | VTYTQL | Y302 | [44] |
| IVYAEL | Y332 | ||||
| KIR2DL3 | Monomer | 2× ITIM | VTYAQL | Y303 | [44] |
| IVYTEL | Y333 | ||||
| KIR2DL5 | Monomer | 2× ITIM | VTYAQL | Y298 | [44] |
| TMYMEL | Y228 | ||||
| KIR3DL1 | Monomer | 2× ITIM | VTYAQL | Y398 | [44] |
| ILYTEL | Y428 | ||||
| KIR3DL2 | Monomer | 2× ITIM | VTYAQL | Y398 | [44] |
| SVYTEL | Y428 | ||||
| KIR3DL3 | Homodimer | 1× ITIM | VTYAQL | Y381 | [44,139] |
| LILRB1/ILT2 | Monomer | 4× ITIM | NLYAAV | Y533 | [50,51,140] |
| VTYAEV | Y562 | ||||
| VTYAQL | Y614 | ||||
| SIYATL | Y644 | ||||
| SIGLEC-7/9 | Monomer | 1× ITIM | IQYAPL/LQYASL | Y437/Y433 | [131,132] |
| 1× ITIM | NEYSEI/TEYSEI | Y460/Y456 | |||
| NKRP1A | Homodimer | 1× ITIM-like | AIYAEL | Y7 | [141] |
| Mouse | |||||
| Ly49A | Homodimer | 1× ITIM | VTYSMV | Y8 | [93,142] |
| Ly49B | Homodimer | 1× ITIM | VTYTTL | Y8 | [93,142] |
| Ly49C, E-I | Homodimer | 1× ITIM | VTYSTL | Y8 | [93,142] |
| NKRP1B | Homodimer | ITIM | LVYADL | Y8 | [141] |
| Shared | |||||
| iNKG2A/B | Heterodimer with CD94 | 2× ITIM | (h) VIYSDL | Y8 | [95,96] |
| (m) VTYAEL | |||||
| 1× ITIM | (h) EITYAEL | Y40 | [95,96] | ||
| 1× ITIM-like | (m) IIYSDF | ||||
| TIGIT | Monomer | 1× ITT-like | YFN | (h) Y225 | [82,83] |
| (m) Y230 | |||||
| 1× ITIM | LSYRSL | (h) Y231 | |||
| (m) Y236 | |||||
| TACTILE | Monomer | 1× ITIM | IKYTCI | (h) Y566 | [105] |
| (m) Y583 | |||||
| 1× YxxM | (h) YHEM | (h) Y579 | |||
| CEACAM1 | Homomer Homodimer Oligomer | (h) 2× ITIM | VTYSTL | Y459 | [84,143,144] |
| IIYSEV | Y486 | ||||
| (m) 1× ITIM | VAYTVL | Y454 | |||
| (m) 1× ITSM | TVYSEV | Y481 | |||
| LAIR-1 | Monomer | 2× ITIM | (h) VTYAQL | Y251 | [145] |
| (h) ITYAAV | Y281 | ||||
| (m) VTYIQL | Y228 | ||||
| (m) STYAAI | Y257 | ||||
| KLGR1 | Homodimer | 1× ITIM | (h) VIYSML | Y7 | [146,147] |
| (m) SIYSTL | |||||
| Shared | Form | Signaling Motif 1 | pY 2 Position | Reference | |
| Tyro | Homodimer Heterodimer | 1× ITIM-like | IYNYL | (h) Y742 (m) Y744 | [148] |
| Axl | Homodimer Heterodimer | 1× ITIM | LLYSRL | (h) Y634 (m) Y628 | [148] |
| 1× ITIM-like | IYDYL | (h) Y759/761 (m) Y753/755 | |||
| Mer | Homodimer Heterodimer | 1× ITIM | LLYSRL | (h) Y685 (m) Y680 | [148] |
| 1× ITIM-like | MYDYL | (h) Y810/Y812 (m) Y805/807 | |||
1 m = mouse, h = human; 2 Phosphorylated tyrosine (pY).
Table 4.
Activating NK receptors paired to cellular ligands and signaling adaptors.
| Human | Ligands | Co-receptor |
|---|---|---|
| NKp30 | B7H6 [184,185] | CD3ζ and FCRγ |
| NKp44 | NKp44L (unusual isoform of MLL5) [186,187] | DAP12 |
| Shared | ||
| NKG2D | MICA/B, ULBP1-6 [188,189] (humans) | DAP10 |
| Rae1B, MULT1, H60 [190,191] (mice) | DAP10 and DAP12 | |
| NKp46 | Unknown | CD3ζ and FCRγ |
| DNAM-1 | CD155 and CD112 [82,83] | - |
| 2B4 | CD48 (SLAMF2) [192,193]. | - |
| NTB-A | NTB-A (SLAMF6, CD319 and Ly108 in mice) [192,193]. | - |
| CRACC | CRACC (SLAMF7, CD352) [192,193]. | - |
| CD16 | IgG [194,195] | CD3ζ and FCRγ |
Table 5.
Activating signaling motifs.
| Adaptor Proteins | Signaling Motif | Motif Sequence | pY 1,2 Position |
| DAP10 | YxxM | YINM | (h) Y86 |
| (m) Y71 | |||
| DAP12 | ITAM | YQELQGQRSDVYSDL | (h) Y91 & Y102 |
| ITAM | YQELQGQRPEVYSDL | (m) Y92 & Y103 | |
| FCR | ITAM | YTGLSTRNQETYETL | Y65 & Y76 |
| CD3 | 3x ITAM | YNELNLGRREEYDVL | Y72 & Y83 |
| YNALQKDKMAEAYSEI | Y111 & Y123 | ||
| YQGLSTATKDTYDAL | Y142 & Y153 | ||
| Receptor | Signaling Motif | Motif Sequence | pY1 Position |
| NKp44 | ITIM-like | EILYHTVA | (h) Y258 |
| DNAM-1 | ITT-like | YvNY | (h) Y322 & Y325 |
| (m) Y319 & Y322 | |||
| 2B4 | ITSM 1 | TIYEDV | (h) Y271 |
| TIYEYV | (m) Y266 & Y268 | ||
| ITSM 2 | TIYSMI | (h) Y297 | |
| TMYSMI | (m) Y325 | ||
| ITSM 3 | TLYSLI | (h) Y317 | |
| TVYSVV | (m) Y343 | ||
| ITSM 4 | T IYEVI | (h) Y342 | |
| TVYEEV | (m) Y369 | ||
| NTB-A | ITSM 1 | TVYASV | (h) Y285 |
| TVYAQV | (m) Y295 | ||
| ITSM 2 | TIYSTI | (h) Y309 | |
| TIYS IV | (m) Y319 | ||
| CRACC | ITSM-like | TEYDTI | (h) Y284 |
| ADYDTI | (m) Y282 | ||
| ITSM | TVYSTV | (h) Y304 | |
| TFYSTV | (m) Y302 |
1 m = mouse, h = human; 2 Phosphorylated tyrosine (pY).
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Meza Guzman, L.G.; Keating, N.; Nicholson, S.E. Natural Killer Cells: Tumor Surveillance and Signaling. Cancers 2020, 12, 952. https://doi.org/10.3390/cancers12040952
AMA Style
Meza Guzman LG, Keating N, Nicholson SE. Natural Killer Cells: Tumor Surveillance and Signaling. Cancers. 2020; 12(4):952. https://doi.org/10.3390/cancers12040952
Chicago/Turabian StyleMeza Guzman, Lizeth G., Narelle Keating, and Sandra E. Nicholson. 2020. "Natural Killer Cells: Tumor Surveillance and Signaling" Cancers 12, no. 4: 952. https://doi.org/10.3390/cancers12040952
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.