LncRNA DLG2-AS1 as a Novel Biomarker in Lung Adenocarcinoma

 , , , , , and
, , , , , and
Abstract
:1. Introduction
2. Results
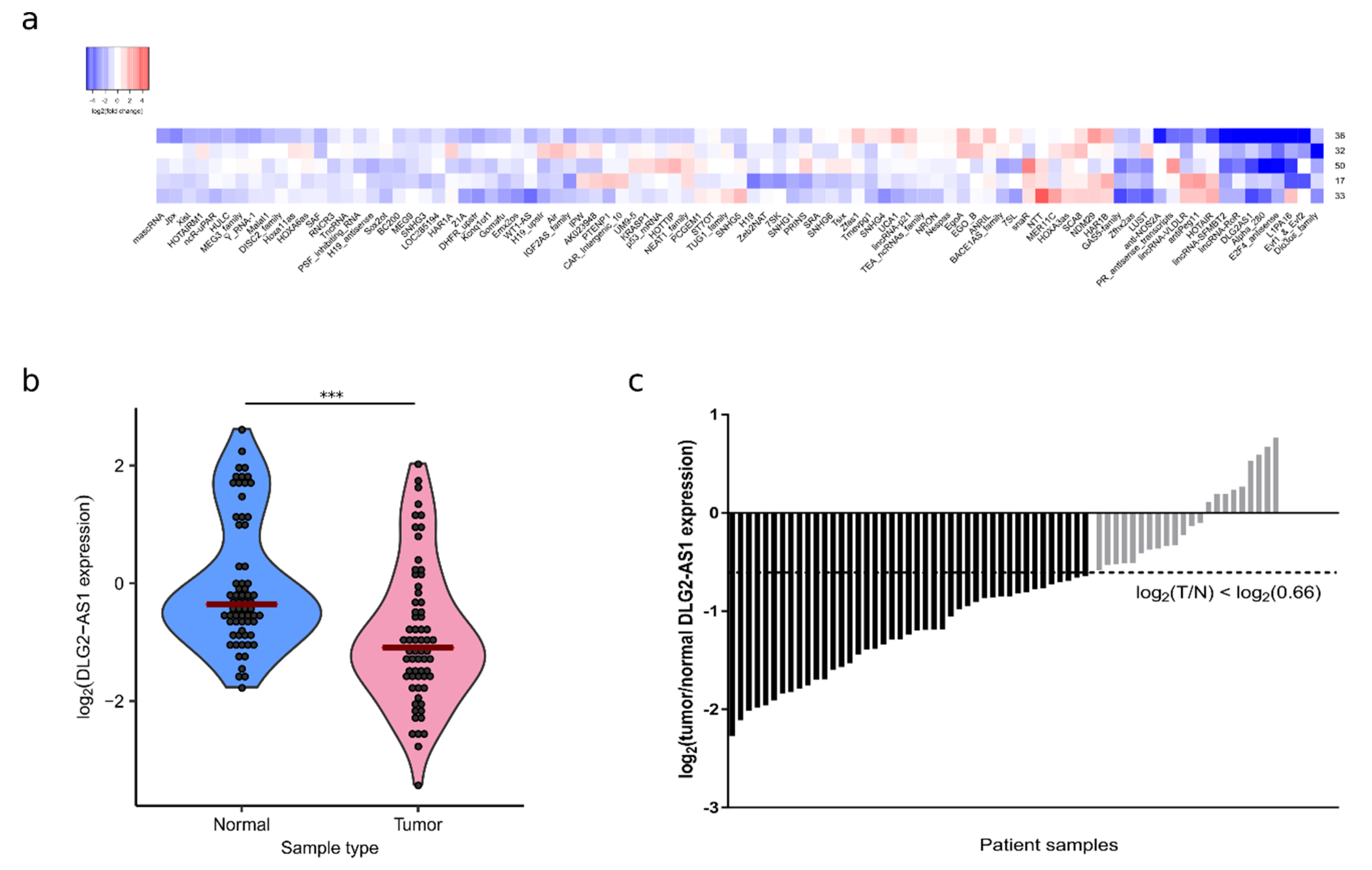
2.1. DLG2-AS1 Expression Is Downregulated in Lung Adenocarcinoma
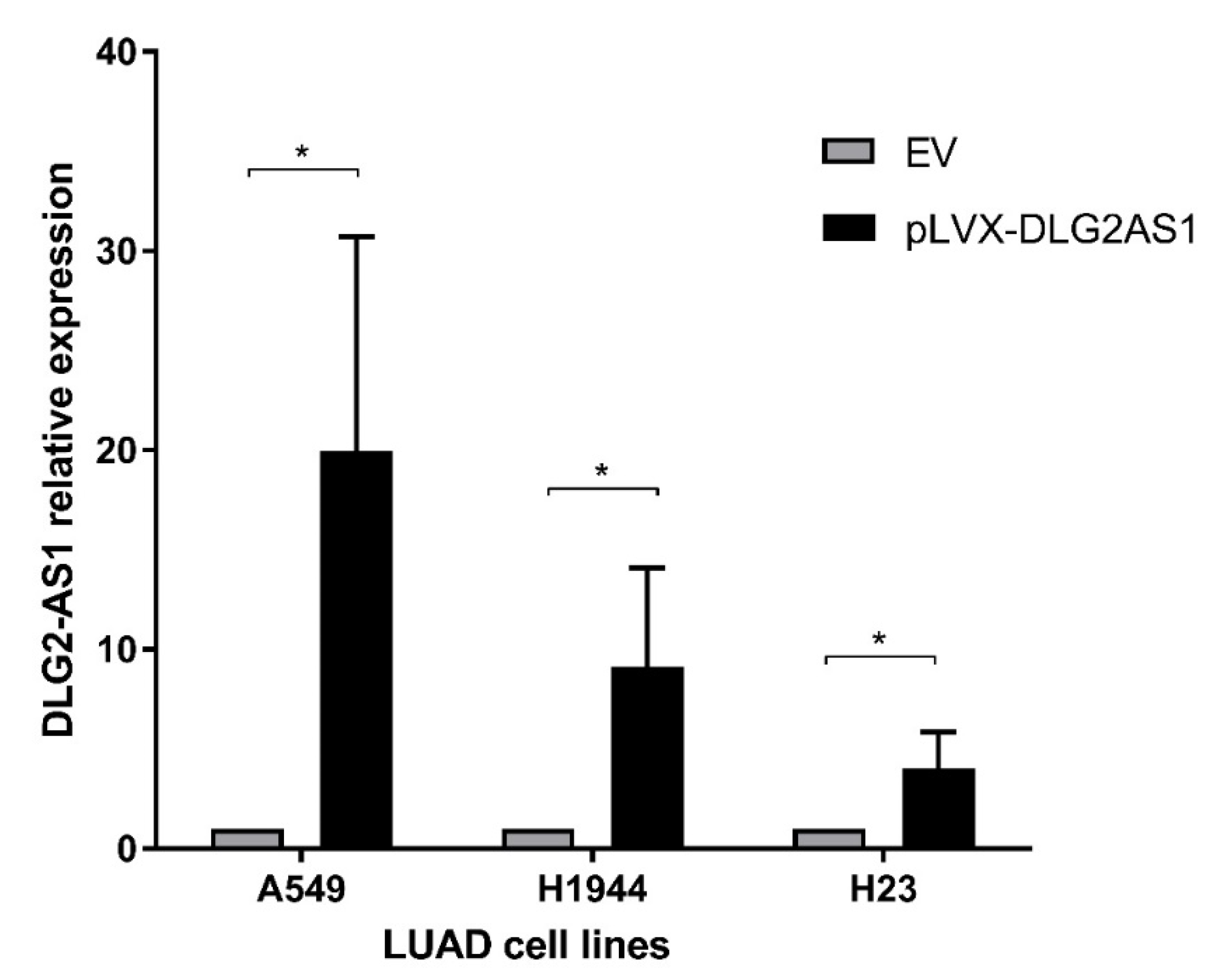
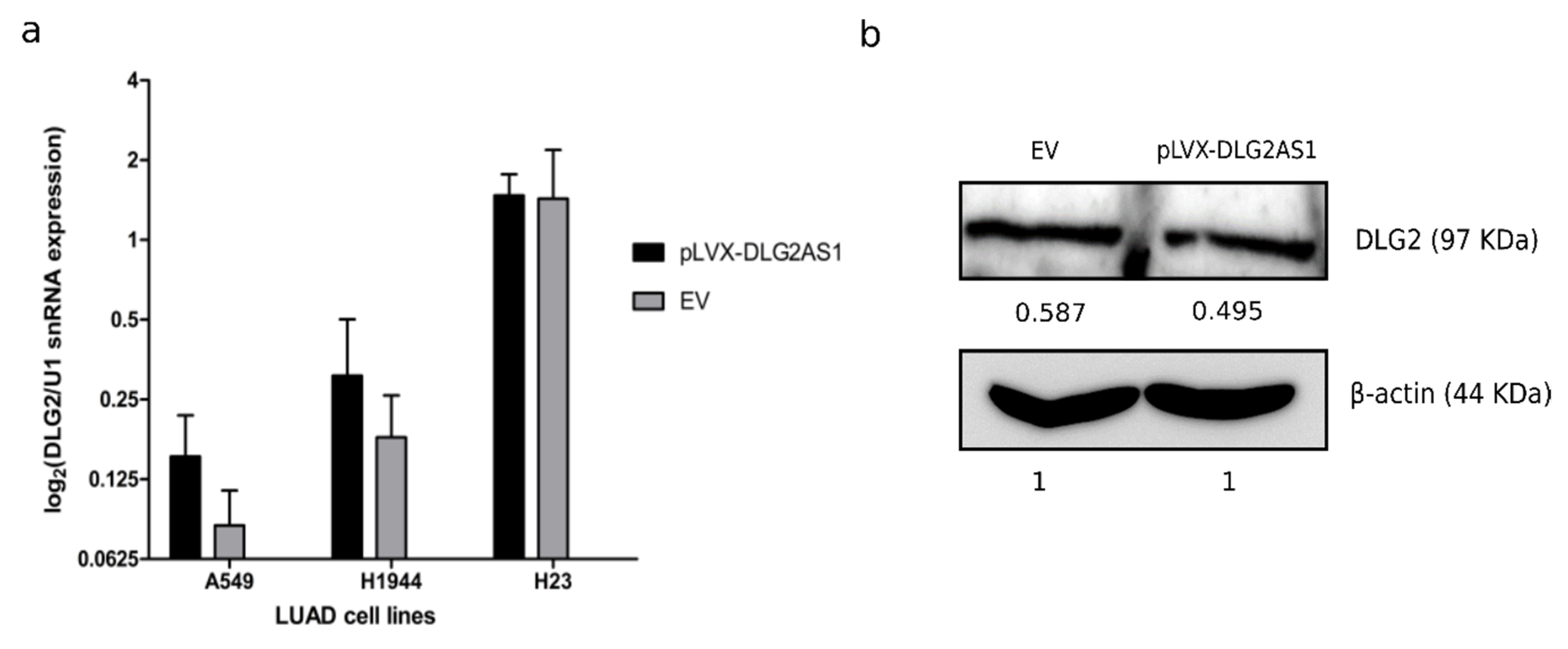
2.2. DLG2-AS1 Is Not a Cis-regulator of DLG2 Expression
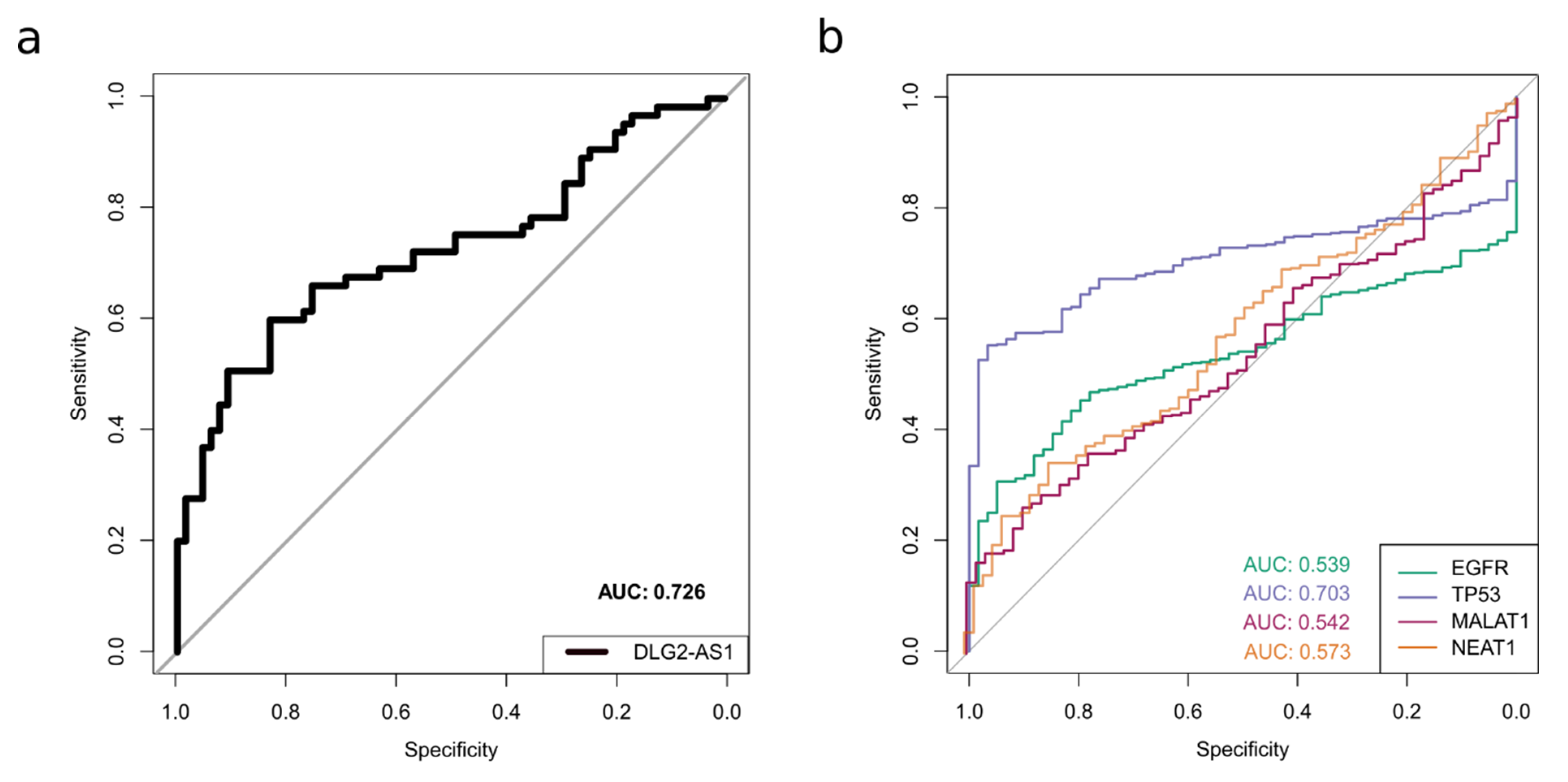
2.3. LncRNA DLG2-AS1 Expression Shows Potential as a Lung Adenocarcinoma Biomarker
3. Discussion
4. Materials and Methods
4.1. Sample Collection and Ethics Approval
4.2. LncRNA Profiling and Candidate Selection
4.3. Cell Culture
4.4. Cloning of DLG2-AS1 Vector and Transfection
4.5. DLG2 Protein Quantification
4.6. Analysis of TCGA-LUAD Data
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Oligonucleotide Name | 5′ ➔ 3′ Sequence |
|---|---|
| DLG2-AS1 Fw | ATCCGGATGTGAGGTTATAAT |
| DLG2-AS1 Rv | AATCCAGATCCCAAGACTTC |
| DLG2 Fw | GGGAGATACCATGATCACG |
| DLG2 Rv | CCACAAATTATGCAGTCGAGTTTCCC |
| U1 snRNA Fw | GAAGACCTCATTCTTTCCTATG |
| U1 snRNA Rv | CGGCTTCTATAAACTTGTGC |
| AAAA-EcoRI-DLG2-AS1 | AAAAGAATTCTTAACTTGTTAATCA |
| TTTT-XbaI-DLG2-AS1 | TTTTTCTAGACCAGATGGTCAGTGA |


References
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics. CA Cancer J. Clin. 2020, 70, 7–30. [Google Scholar] [CrossRef] [PubMed]
- Torre, L.A.; Siegel, R.L.; Jemal, A. Lung Cancer Statistics. Adv. Exp. Med. Biol. 2016, 893, 1–19. [Google Scholar] [CrossRef] [PubMed]
- ENCODE Project Consortium. An integrated encyclopedia of DNA elements in the human genome. Nature 2012, 489, 57–74. [Google Scholar] [CrossRef] [PubMed]
- Do, H.; Kim, W. Roles of Oncogenic Long Non-coding RNAs in Cancer Development. Genom. Inform. 2018, 16, e18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhat, S.A.; Ahmad, S.M.; Mumtaz, P.T.; Malik, A.A.; Dar, M.A.; Urwat, U.; Shah, R.A.; Ganai, N.A. Long non-coding RNAs: Mechanism of action and functional utility. Noncoding RNA Res. 2016, 1, 43–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanly, D.J.; Esteller, M.; Berdasco, M. Interplay between long non-coding RNAs and epigenetic machinery: Emerging targets in cancer? Philos. Trans. R. Soc. B Biol. Sci. 2018, 373, 20170074. [Google Scholar] [CrossRef]
- Perkel, J.M. Visiting “noncodarnia”. Biotechniques 2013, 54, 301, 303–304. [Google Scholar] [CrossRef]
- Peinado, P.; Herrera, A.; Baliñas, C.; Martín-Padrón, J.; Boyero-Corral, L.; Cuadros, M.; Coira, I.F.; Rodríguez-Lara, M.I.; Reyes-Zurita, F.; Rufino-Palomares, E.E.; et al. Long Non-Coding RNAs as Cancer Biomarkers. In Cancer and Noncoding RNAs, 1st ed.; Elsevier: Amsterdam, The Netherlands, 2008; pp. 95–114. [Google Scholar]
- Kornienko, A.E.; Guenzl, P.M.; Barlow, D.P.; Pauler, F.M. Gene regulation by the act of long non-coding RNA transcription. BMC Biol. 2013, 11, 59. [Google Scholar] [CrossRef] [Green Version]
- Loewen, G.; Jayawickramarajah, J.; Zhuo, Y.; Shan, B. Functions of IncRNA HOTAIR in lung cancer. J. Hematol. Oncol. 2014, 7, 90. [Google Scholar] [CrossRef] [Green Version]
- Ji, P.; Diederichs, S.; Wang, W.; Böing, S.; Metzger, R.; Schneider, P.M.; Tidow, N.; Brandt, B.; Buerger, H.; Bulk, E.; et al. MALAT-1, a novel noncoding RNA, and thymosin β4 predict metastasis and survival in early-stage non-small cell lung cancer. Oncogene 2003, 22, 8031–8041. [Google Scholar] [CrossRef] [Green Version]
- Gutschner, T.; Hämmerle, M.; Eißmann, M.; Hsu, J.; Kim, Y.; Hung, G.; Revenko, A.; Arun, G.; Stentrup, M.; Groß, M.; et al. The Noncoding RNA MALAT1 Is a Critical Regulator of the Metastasis Phenotype of Lung Cancer Cells. Cancer Res. 2013, 73, 1180–1189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, R.A.; Shah, N.; Wang, K.C.; Kim, J.; Horlings, H.M.; Wong, D.J.; Tsai, M.C.; Hung, T.; Argani, P.; Rinn, J.L.; et al. Long non-coding RNA HOTAIR reprograms chromatin state to promote cancer metastasis. Nature 2010, 464, 1071–1076. [Google Scholar] [CrossRef] [PubMed]
- Zhai, N.; Xia, Y.; Yin, R.; Liu, J.; Gao, F. A negative regulation loop of long noncoding RNA HOTAIR and p53 in non-small-cell lung cancer. Onco Targets Ther. 2016, 9, 5713–5720. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, Q.; Hann, S.S. HOTAIR: An Oncogenic Long Non-Coding RNA in Human Cancer. Cell. Physiol. Biochem. 2018, 47, 893–913. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Jin, Y.; Ren, O.; Ma, X.; Wang, B.; Wang, Y. Downregulation of the long noncoding RNA TUSC7 promotes NSCLC cell proliferation and correlates with poor prognosis. Am. J. Transl. Res. 2016, 8, 680–687. [Google Scholar] [PubMed]
- Zhou, H.; Chen, A.; Shen, J.; Zhang, X.; Hou, M.; Li, J.; Chen, J.; Zou, H.; Zhang, Y.; Deng, Q.; et al. Long non-coding RNA LOC285194 functions as a tumor suppressor by targeting p53 in non-small cell lung cancer. Oncol. Rep. 2019, 41, 15–26. [Google Scholar] [CrossRef] [Green Version]
- Lu, T.; Wang, Y.; Chen, D.; Liu, J.; Jiao, W. Potential clinical application of lncRNAs in non-small cell lung cancer. Onco Targets Ther. 2018, 11, 8045–8052. [Google Scholar] [CrossRef] [Green Version]
- Villegas, V.E.; Zaphiropoulos, P.G. Neighboring gene regulation by antisense long non-coding RNAs. Int. J. Mol. Sci. 2015, 16, 3251. [Google Scholar] [CrossRef] [Green Version]
- Zucchelli, S.; Fasolo, F.; Russo, R.; Cimatti, L.; Patrucco, L.; Takahashi, H.; Jones, M.H.; Santoro, C.; Sblattero, D.; Cotella, D.; et al. SINEUPs are modular antisense long non-coding RNAs that increase synthesis of target proteins in cells. Front. Cell. Neurosci. 2015, 9, 174. [Google Scholar] [CrossRef] [Green Version]
- Wu, C.H.; Hwang, M.J. Risk stratification for lung adenocarcinoma on EGFR and TP53 mutation status, chemotherapy, and PD-L1 immunotherapy. Cancer Med. 2019, 8, 5850–5861. [Google Scholar] [CrossRef] [Green Version]
- Molina, J.R.; Yang, P.; Cassivi, S.D.; Schild, S.E.; Adjei, A.A. Non-small cell lung cancer: Epidemiology, risk factors, treatment, and survivorship. Mayo Clin. Proc. 2008, 83, 584–594. [Google Scholar] [CrossRef]
- Lee, G.L.; Dobi, A.; Srivastava, S. Prostate cancer: Diagnostic performance of the PCA3 urine test. Nat. Rev. Urol. 2011, 8, 123–124. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.; Cho, K.O.; Rothschild, A.; Sheng, M. Heteromultimerization and NMDA receptor-clustering activity of Chapsyn-110, a member of the PSD-95 family of proteins. Neuron 1996, 17, 103–113. [Google Scholar] [CrossRef] [Green Version]
- Mazoyer, S.; Gayther, S.A.; Nagai, M.A.; Smith, S.A.; Dunning, A.; van Rensburg, E.J.; Albertsen, H.; White, R.; Ponder, B.A. A gene (DLG2) located at 17q12-q21 encodes a new homologue of the Drosophila tumor suppressor dIg-A. Genomics 1995, 28, 25–31. [Google Scholar] [CrossRef] [PubMed]
- Zubakov, D.; Stupar, Z.; Kovacs, G. Differential expression of a new isoform of DLG2 in renal oncocytoma. BMC Cancer 2006, 6, 106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Polesskaya, O.O.; Haroutunian, V.; Davis, K.L.; Hernandez, I.; Sokolov, B.P. Novel putative nonprotein-coding RNA gene from 11q14 displays decreased expression in brains of patients with schizophrenia. J. Neurosci. Res. 2003, 74, 111–122. [Google Scholar] [CrossRef]
- Somarowthu, S.; Legiewicz, M.; Chillón, I.; Marcia, M.; Liu, F.; Pyle, A.M. HOTAIR forms an intricate and modular secondary structure. Mol. Cell. 2015, 58, 353–361. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.; Schmidt, B.F.; Bruchez, M.P.; McManus, C.J. Structural analyses of NEAT1 lncRNAs suggest long-range RNA interactions that may contribute to paraspeckle architecture. Nucleic Acids Res. 2018, 46, 3742–3752. [Google Scholar] [CrossRef]
- Uroda, T.; Anastasakou, E.; Rossi, A.; Teulon, J.M.; Pellequer, J.L.; Annibale, P.; Pessey, O.; Inga, A.; Chillón, I.; Marcia, M. Conserved Pseudoknots in IncRNA MEG3 Are Essential for Stimulation of the p53 Pathway. Mol. Cell. 2019, 75, 982–995. [Google Scholar] [CrossRef] [Green Version]
- Xue, Z.; Hennelly, S.; Doyle, B.; Gulati, A.A.; Novikova, I.V.; Sanbonmatsu, K.Y.; Boyer, L.A. A G-rich motif in the IncRNA Braveheart interacts with a zinc finger transcription factor to specify the cardiovascular lineage. Mol. Cell. 2016, 64, 37–50. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.N.; Thiel, B.C.; Mrozowich, T.; Hennelly, S.P.; Hofacker, I.L.; Patel, T.R.; Sanbonmatsu, K.Y. Zinc-finger protein CNBP alters the 3-D structure of IncRNA Braveheart in solution. Nat. Commun. 2020, 11, 148. [Google Scholar] [CrossRef] [PubMed]


© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arenas, A.M.; Cuadros, M.; Andrades, A.; García, D.J.; Coira, I.F.; Rodríguez, M.I.; Baliñas-Gavira, C.; Peinado, P.; Álvarez-Pérez, J.C.; Medina, P.P. LncRNA DLG2-AS1 as a Novel Biomarker in Lung Adenocarcinoma. Cancers 2020, 12, 2080. https://doi.org/10.3390/cancers12082080
Arenas AM, Cuadros M, Andrades A, García DJ, Coira IF, Rodríguez MI, Baliñas-Gavira C, Peinado P, Álvarez-Pérez JC, Medina PP. LncRNA DLG2-AS1 as a Novel Biomarker in Lung Adenocarcinoma. Cancers. 2020; 12(8):2080. https://doi.org/10.3390/cancers12082080
Chicago/Turabian StyleArenas, Alberto M., Marta Cuadros, Alvaro Andrades, Daniel J. García, Isabel F. Coira, María Isabel Rodríguez, Carlos Baliñas-Gavira, Paola Peinado, Juan Carlos Álvarez-Pérez, and Pedro P. Medina. 2020. "LncRNA DLG2-AS1 as a Novel Biomarker in Lung Adenocarcinoma" Cancers 12, no. 8: 2080. https://doi.org/10.3390/cancers12082080