Inflammatory Proteins HMGA2 and PRTN3 as Drivers of Vulvar Squamous Cell Carcinoma Progression

 , , and
, , and
Abstract
:Simple Summary
Abstract
1. Introduction
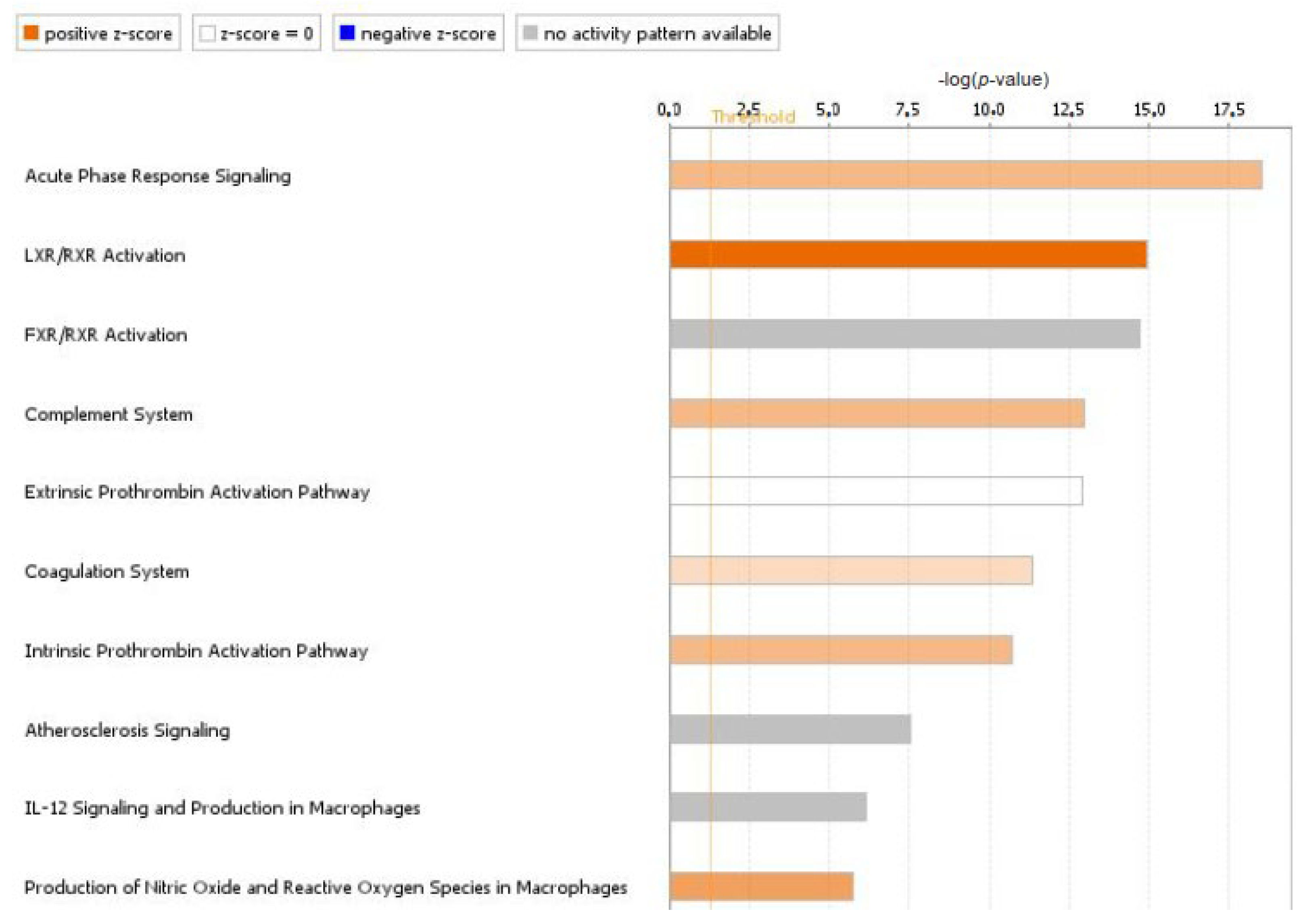
2. Results
2.1. Identification of Differentially Expressed Proteins Using iTRAQ
2.2. Verification by Parallel Reaction Monitoring
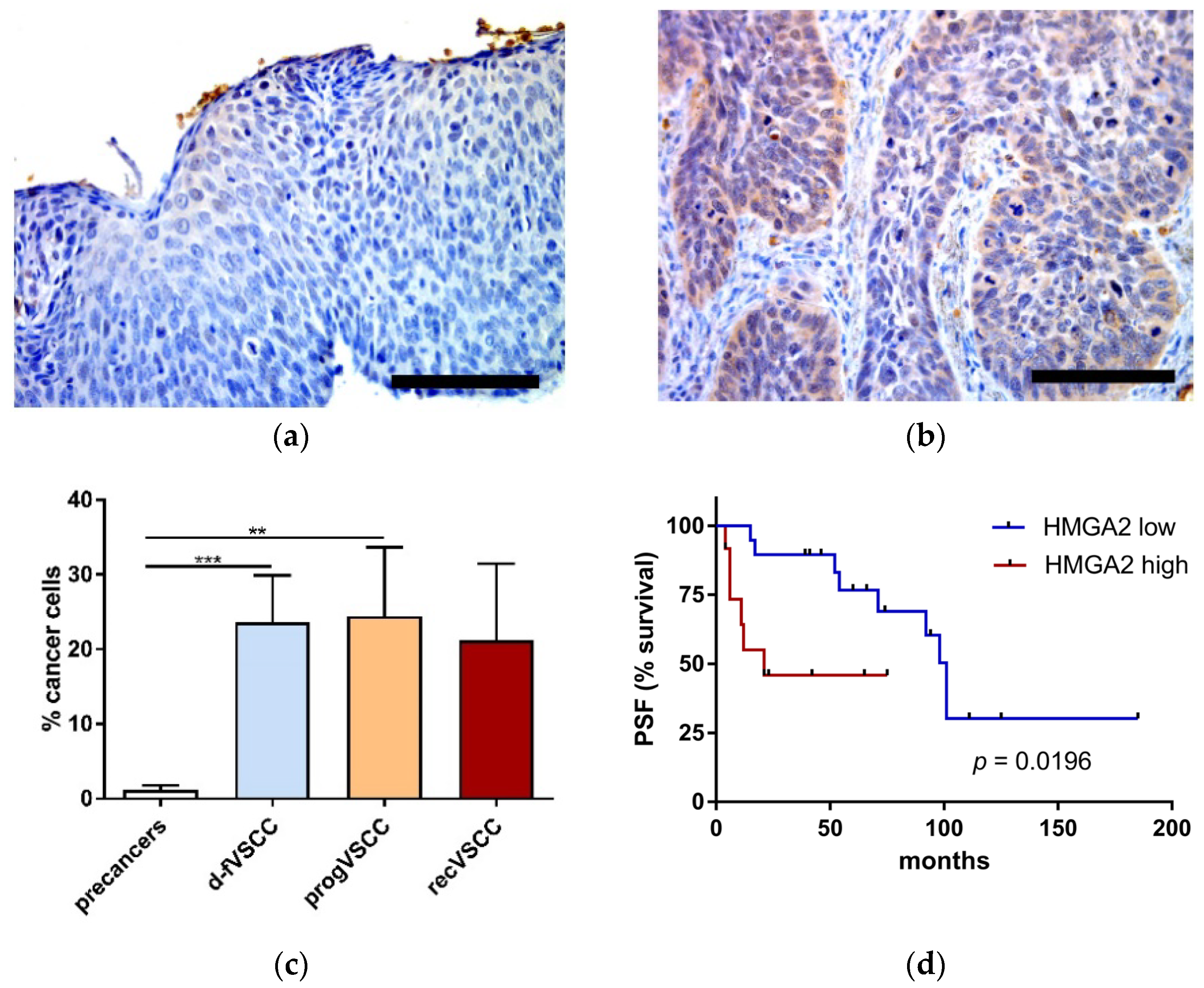
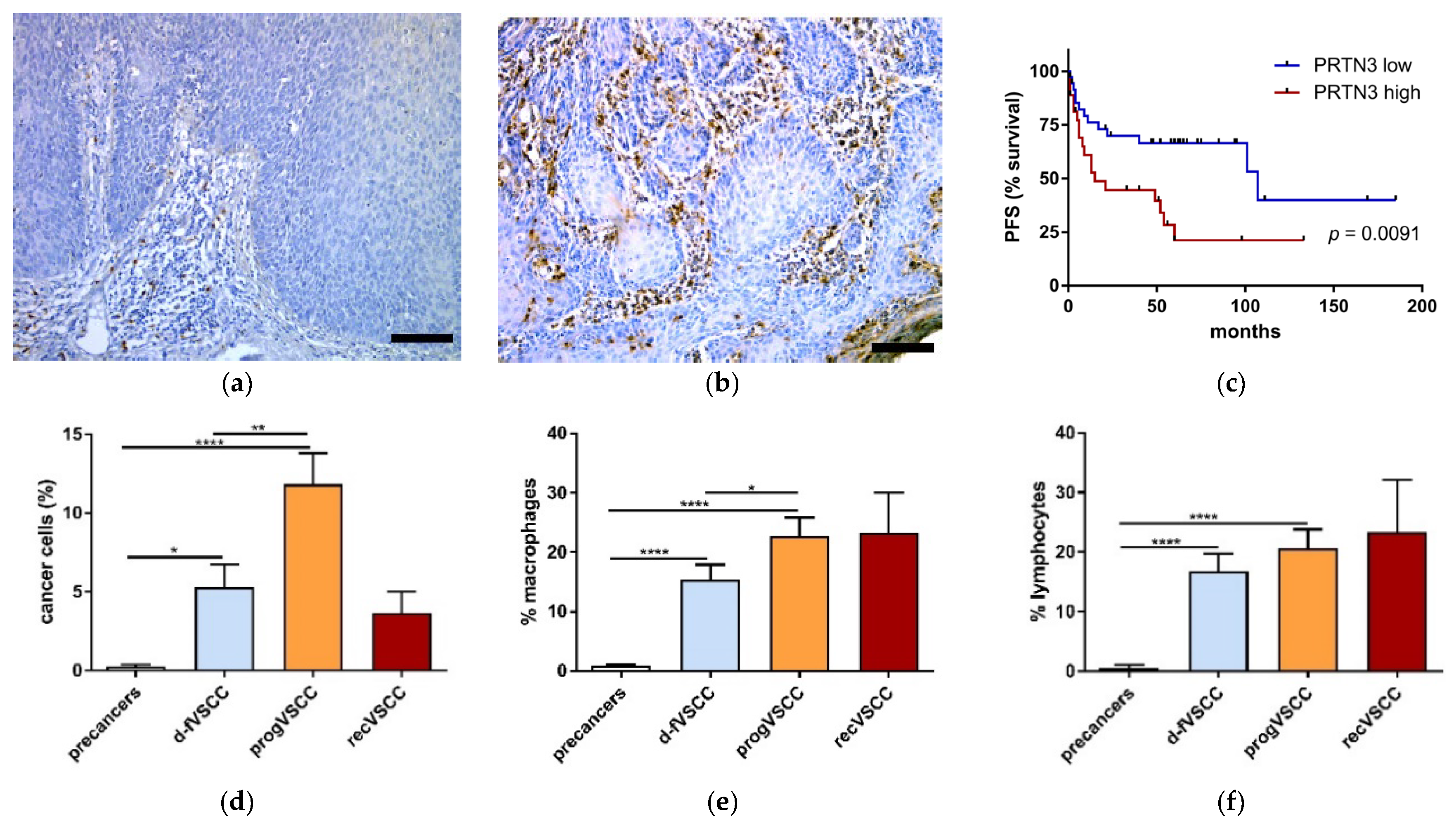
2.3. Immunohistochemistry
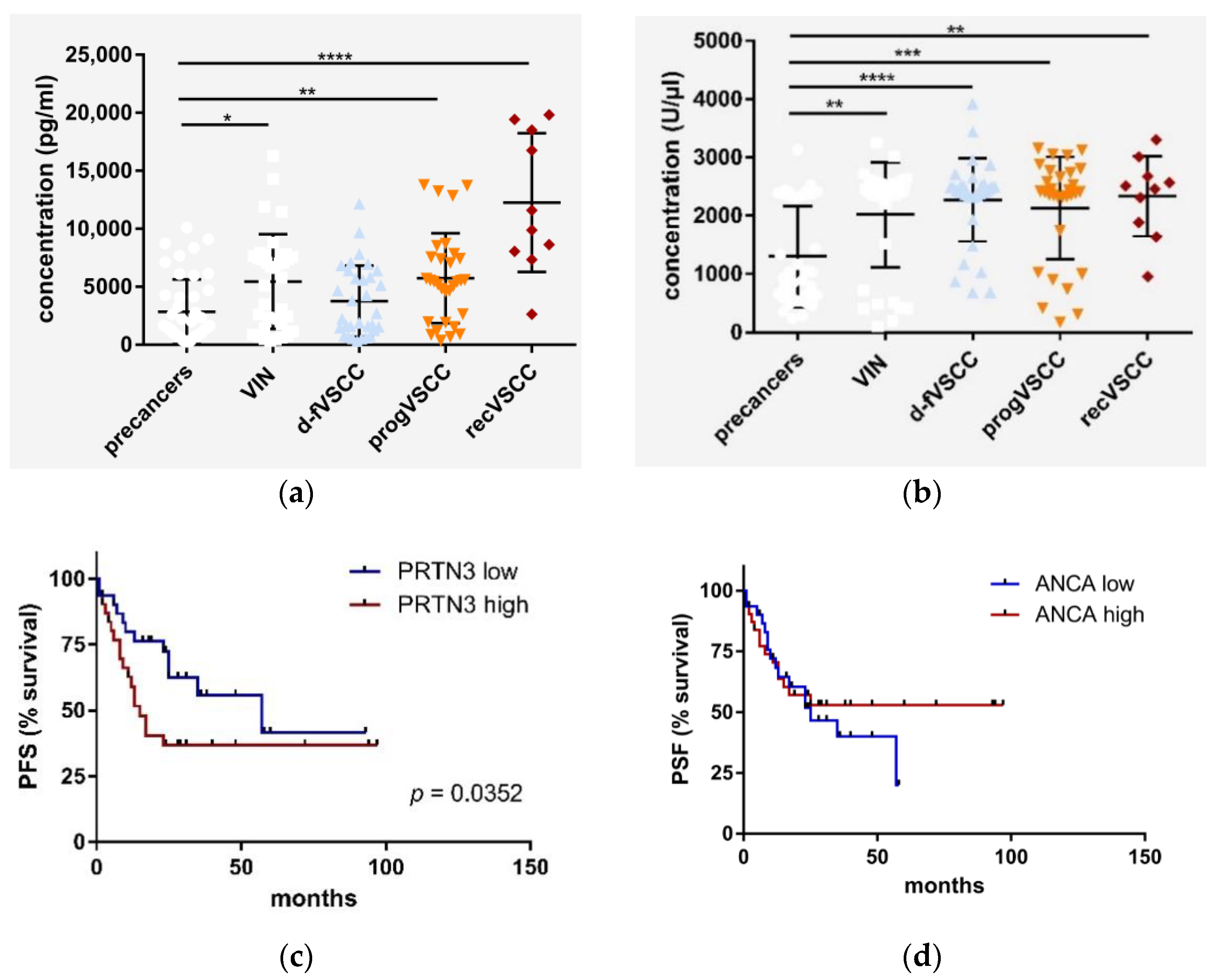
2.4. PRTN3 Screening in VSCC Patients’ Blood
2.5. The Relevance of HPV Status of VSCC Tumors
3. Discussion
4. Patients and Methods
4.1. Patients and Samples
4.2. Methods
4.2.1. High Risk HPV Genotyping
4.2.2. Isobaric Tags for Relative and Absolute Quantitation (iTRAQ)
4.2.3. Protein Verification by Parallel Reaction Monitoring (PRM)
4.2.4. Histopathology and Immunohistochemistry
4.2.5. Enzyme-Linked Immunosorbent Assays (ELISA)
4.2.6. Let-7c microRNA Quantification in Plasma
4.2.7. Vulvovaginal Microorganism Detection Using PCR
4.2.8. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [Green Version]
- Cancer Stat Facts: Vulvar Cancer. Available online: https://seer.cancer.gov/statfacts/html/vulva.html (accessed on 5 December 2020).
- The European Society of Gynaecologic Oncology (ESGO) Vulvar Cancer Guidelines. Available online: https://guidelines.esgo.org/media/2016/08/ESGO-Vulvar-cancer-Complete-report-fxd2.pdf (accessed on 5 December 2020).
- Gill, B.S.; Bernard, M.E.; Lin, J.F.; Balasubramani, G.K.; Rajagopalan, M.S.; Sukumvanich, P.; Krivak, T.C.; Olawaiye, A.B.; Kelley, J.L.; Beriwal, S. Impact of adjuvant chemotherapy with radiation for node-positive vulvar cancer: A National Cancer Data Base (NCDB) analysis. Gynecol. Oncol. 2015, 137, 365–372. [Google Scholar] [CrossRef]
- Gadducci, A.; Aletti, G.D. Locally advanced squamous cell carcinoma of the vulva: A challenging question for gynecologic oncologists. Gynecol. Oncol. 2020, 158, 208–217. [Google Scholar] [CrossRef]
- Cohen, P.A.; Anderson, L.; Eva, L.; Scurry, J. Clinical and molecular classification of vulvar squamous pre-cancers. Int. J. Gynecol. Cancer 2019, 29, 821–828. [Google Scholar] [CrossRef] [PubMed]
- Horne, Z.D.; Dohopolski, M.J.; Pradhan, D.; Bhargava, R.; Edwards, R.P.; Kelley, J.L.; Comerci, J.T.; Olawaiye, A.B.; Courtney-Brooks, M.B.; Bockmeier, M.M.; et al. Human papillomavirus infection mediates response and outcome of vulvar squamous cell carcinomas treated with radiation therapy. Gynecol. Oncol. 2018, 151, 96–101. [Google Scholar] [CrossRef] [PubMed]
- Clancy, A.A.; Spaans, J.N.; Weberpals, J.I. The forgotten woman’s cancer: Vulvar squamous cell carcinoma (VSCC) and a targeted approach to therapy. Ann. Oncol. 2016, 27, 1696–1705. [Google Scholar] [CrossRef] [PubMed]
- Zieba, S.; Chechlinska, M.; Kowalik, A.; Kowalewska, M. Genes, pathways and vulvar carcinoma—New insights from next-generation sequencing studies. Gynecol. Oncol. 2020, 158, 498–506. [Google Scholar] [CrossRef]
- Belczacka, I.; Latosinska, A.; Metzger, J.; Marx, D.; Vlahou, A.; Mischak, H.; Frantzi, M. Proteomics biomarkers for solid tumors: Current status and future prospects. Mass Spectrom Rev. 2019, 38, 49–78. [Google Scholar] [CrossRef]
- Balzeau, J.; Menezes, M.R.; Cao, S.; Hagan, J.P. The LIN28/let-7 Pathway in Cancer. Front. Genet. 2017, 8, 31. [Google Scholar] [CrossRef] [Green Version]
- Skold, S.; Rosberg, B.; Gullberg, U.; Olofsson, T. A secreted proform of neutrophil proteinase 3 regulates the proliferation of granulopoietic progenitor cells. Blood 1999, 93, 849–856. [Google Scholar] [CrossRef]
- von Vietinghoff, S.; Eulenberg, C.; Wellner, M.; Luft, F.C.; Kettritz, R. Neutrophil surface presentation of the anti-neutrophil cytoplasmic antibody-antigen proteinase 3 depends on N-terminal processing. Clin. Exp. Immunol. 2008, 152, 508–516. [Google Scholar] [CrossRef] [PubMed]
- Meyer-Hoffert, U.; Wiedow, O. Neutrophil serine proteases: Mediators of innate immune responses. Curr. Opin. Hematol. 2011, 18, 19–24. [Google Scholar] [CrossRef] [PubMed]
- Peng, X.; Ramstrom, S.; Kurz, T.; Grenegard, M.; Segelmark, M. The neutrophil serine protease PR3 induces shape change of platelets via the Rho/Rho kinase and Ca(2+) signaling pathways. Thromb. Res. 2014, 134, 418–425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mitra, S.; Sharma, M.K.; Kaur, I.; Khurana, R.; Modi, K.B.; Narang, R.; Mandal, A.; Dutta, S. Vulvar carcinoma: Dilemma, debates, and decisions. Cancer Manag. Res. 2018, 10, 61–68. [Google Scholar] [CrossRef] [Green Version]
- Sandberg, A.; Lindell, G.; Kallstrom, B.N.; Branca, R.M.; Danielsson, K.G.; Dahlberg, M.; Larson, B.; Forshed, J.; Lehtio, J. Tumor proteomics by multivariate analysis on individual pathway data for characterization of vulvar cancer phenotypes. Mol. Cell Proteomics 2012, 11, M112.016998. [Google Scholar] [CrossRef] [Green Version]
- Abdulrahman, Z.; Kortekaas, K.E.; De Vos Van Steenwijk, P.J.; Van Der Burg, S.H.; Van Poelgeest, M.I. The immune microenvironment in vulvar (pre)cancer: Review of literature and implications for immunotherapy. Expert Opin. Biol. Ther. 2018, 18, 1223–1233. [Google Scholar] [CrossRef]
- Robertson, E.S. Microbiome and Cancer; Humana Press: Cham, Switzerland, 2019. [Google Scholar] [CrossRef]
- Korkmaz, B.; Horwitz, M.S.; Jenne, D.E.; Gauthier, F. Neutrophil elastase, proteinase 3, and cathepsin G as therapeutic targets in human diseases. Pharmacol. Rev. 2010, 62, 726–759. [Google Scholar] [CrossRef] [Green Version]
- Ertas, I.E.; Gungorduk, K.; Akman, L.; Ozdemir, A.; Terek, M.C.; Ozsaran, A.; Sanci, M.; Dikmen, Y. Can preoperative neutrophil:lymphocyte and platelet:lymphocyte ratios be used as predictive markers for lymph node metastasis in squamous cell carcinoma of the vulva? Eur. J. Obstet. Gynecol. Reprod. Biol. 2013, 171, 138–142. [Google Scholar] [CrossRef]
- Gibbs, R.S. Microbiology of the female genital tract. Am. J. Obstet. Gynecol. 1987, 156, 491–495. [Google Scholar] [CrossRef]
- Singh, J.; Kalia, N.; Kaur, M. Recurrent Vulvovaginal Infections: Etiology, Diagnosis, Treatment and Management. In Infectious Diseases and Your Health.; Singh, P.P., Ed.; Springer: Singapore, 2018. [Google Scholar]
- Olejek, A.; Kozak-Darmas, I.; Kellas-Sleczka, S.; Nowak, L. The usefulness of bacteriological screening in wound healing aspect after vulvectomy. Menopause Rev. 2005, 4, 33–36. [Google Scholar]
- Laniewski, P.; Ilhan, Z.E.; Herbst-Kralovetz, M.M. The microbiome and gynaecological cancer development, prevention and therapy. Nat. Rev. Urol. 2020, 17, 232–250. [Google Scholar] [CrossRef] [PubMed]
- Cleynen, I.; Van de Ven, W.J. The HMGA proteins: A myriad of functions (Review). Int. J. Oncol. 2008, 32, 289–305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reeves, R.; Beckerbauer, L. HMGI/Y proteins: Flexible regulators of transcription and chromatin structure. Biochim. Biophys. Acta 2001, 1519, 13–29. [Google Scholar] [CrossRef]
- Fusco, A.; Fedele, M. Roles of HMGA proteins in cancer. Nat. Rev. Cancer 2007, 7, 899–910. [Google Scholar] [CrossRef]
- Agostini, A.; Panagopoulos, I.; Andersen, H.K.; Johannesen, L.E.; Davidson, B.; Trope, C.G.; Heim, S.; Micci, F. HMGA2 expression pattern and TERT mutations in tumors of the vulva. Oncol. Rep. 2015, 33, 2675–2680. [Google Scholar] [CrossRef] [Green Version]
- Clancy, D.M.; Henry, C.M.; Sullivan, G.P.; Martin, S.J. Neutrophil extracellular traps can serve as platforms for processing and activation of IL-1 family cytokines. FEBS J. 2017, 284, 1712–1725. [Google Scholar] [CrossRef] [Green Version]
- Soderberg, D.; Segelmark, M. Neutrophil Extracellular Traps in ANCA-Associated Vasculitis. Front. Immunol. 2016, 7, 256. [Google Scholar] [CrossRef]
- Crisford, H.; Sapey, E.; Stockley, R.A. Proteinase 3; a potential target in chronic obstructive pulmonary disease and other chronic inflammatory diseases. Respir. Res. 2018, 19, 180. [Google Scholar] [CrossRef]
- Pharos database entry. Available online: https://pharos.nih.gov/idg/ligands/sivelestat (accessed on 9 October 2019).
- Hayakawa, M.; Katabami, K.; Wada, T.; Sugano, M.; Hoshino, H.; Sawamura, A.; Gando, S. Sivelestat (selective neutrophil elastase inhibitor) improves the mortality rate of sepsis associated with both acute respiratory distress syndrome and disseminated intravascular coagulation patients. Shock 2010, 33, 14–18. [Google Scholar] [CrossRef] [Green Version]
- van den Berg, C.W.; Tambourgi, D.V.; Clark, H.W.; Hoong, S.J.; Spiller, O.B.; McGreal, E.P. Mechanism of neutrophil dysfunction: Neutrophil serine proteases cleave and inactivate the C5a receptor. J. Immunol. 2014, 192, 1787–1795. [Google Scholar] [CrossRef] [Green Version]
- Kam, A.Y.F.; Piryani, S.O.; McCall, C.M.; Park, H.S.; Rizzieri, D.A.; Doan, P.L. Targeting High Mobility Group Box-1 (HMGB1) Promotes Cell Death in Myelodysplastic Syndrome. Clin. Cancer Res. 2019, 25, 4155–4167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, H.; Li, H.; Chen, X.; Yang, Y.; Li, X.; Li, W.; Huang, C.; Meng, X.; Zhang, L.; Li, J. HMGA2, a driver of inflammation, is associated with hypermethylation in acute liver injury. Toxicol. Appl. Pharmacol. 2017, 328, 34–45. [Google Scholar] [CrossRef] [PubMed]
- Zieba, S.; Kowalik, A.; Zalewski, K.; Rusetska, N.; Goryca, K.; Piascik, A.; Misiek, M.; Bakula-Zalewska, E.; Kopczynski, J.; Kowalski, K.; et al. Somatic mutation profiling of vulvar cancer: Exploring therapeutic targets. Gynecol. Oncol. 2018, 150, 552–561. [Google Scholar] [CrossRef]
- WHO Classification of Tumours. Available online: https://tumourclassification.iarc.who.int/welcome/ (accessed on 8 December 2020).
- Zalewski, K.; Misiek, M.; Kowalik, A.; Bakula-Zalewska, E.; Kopczynski, J.; Zielinska, A.; Bidzinski, M.; Radziszewski, J.; Gozdz, S.; Kowalewska, M. Normalizers for microRNA quantification in plasma of patients with vulvar intraepithelial neoplasia lesions and vulvar carcinoma. Tumour Biol. 2017, 39, 1010428317717140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mayakonda, A.; Lin, D.C.; Assenov, Y.; Plass, C.; Koeffler, H.P. Maftools: Efficient and comprehensive analysis of somatic variants in cancer. Genome Res. 2018, 28, 1747–1756. [Google Scholar] [CrossRef] [Green Version]
- Malinowska, A.; Kistowski, M.; Bakun, M.; Rubel, T.; Tkaczyk, M.; Mierzejewska, J.; Dadlez, M. Diffprot-software for non-parametric statistical analysis of differential proteomics data. J. Proteomics 2012, 75, 4062–4073. [Google Scholar] [CrossRef]
- The Gene Ontology, C. The Gene Ontology Resource: 20 years and still GOing strong. Nucleic Acids Res. 2019, 47, D330–D338. [Google Scholar] [CrossRef] [Green Version]
- Kanehisa, M. Toward understanding the origin and evolution of cellular organisms. Protein Sci. 2019, 28, 1947–1951. [Google Scholar] [CrossRef]
- Falcon, S.; Gentleman, R. Using GOstats to test gene lists for GO term association. Bioinformatics 2007, 23, 257–258. [Google Scholar] [CrossRef] [Green Version]
- van Elsland, D.; Neefjes, J. Bacterial infections and cancer. EMBO Rep. 2018, 19. [Google Scholar] [CrossRef]
- Picardo, S.L.; Coburn, B.; Hansen, A.R. The microbiome and cancer for clinicians. Crit. Rev. Oncol. Hematol. 2019, 141, 1–12. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein | d-fVSCC vs. progVSCC | VSCC vs. Normal Vulvar Tissues | ||||||
|---|---|---|---|---|---|---|---|---|
| p-Value | adjp | FC | AUC | p-Value | adjp | FC | AUC | |
| HMGA2 | 0.0063 | 0.0824 | 14.4 | 0.73 | 0.0059 | 0.0095 | 45.5 | 0.76 |
| ANO1 | 0.0068 | 0.0824 | 3.37 | 0.73 | 0.6177 | 0.6312 | 1.80 | 0.55 |
| PRTN3 | 0.0117 | 0.0981 | 2.75 | 0.71 | <0.0001 | <0.0001 | 13.3 | 0.92 |
| UBE2C | 0.0070 | 0.0824 | 2.72 | 0.73 | 0.0001 | 0.0002 | 8.89 | 0.87 |
| KRT18 | 0.0220 | 0.0981 | 2.48 | 0.70 | <0.0001 | 0.0001 | 8.18 | 0.91 |
| S100A12 | 0.0160 | 0.0981 | 2.44 | 0.71 | <0.0001 | <0.0001 | 12.0 | 0.92 |
| PTX3 | 0.0359 | 0.1211 | 2.31 | 0.68 | 0.0535 | 0.0662 | 2.68 | 0.68 |
| RUNX1 | 0.0192 | 0.0981 | 2.06 | 0.70 | <0.0001 | 0.0001 | 6.73 | 0.90 |
| PADI2 | 0.0192 | 0.0981 | 1.89 | 0.70 | <0.0001 | <0.0001 | 3.72 | 0.88 |
| ISG15 | 0.0273 | 0.1069 | 1.88 | 0.69 | <0.0001 | <0.0001 | 35.4 | 0.94 |
| ABCC10 | 0.2133 | 0.3992 | 1.75 | 0.61 | 0.0068 | 0.0106 | 5.55 | 0.75 |
| IFIT3 | 0.0811 | 0.1907 | 1.74 | 0.65 | <0.0001 | <0.0001 | 16.5 | 0.91 |
| IDO1 | 0.8737 | 0.9126 | 1.68 | 0.52 | 0.0001 | 0.0001 | 17.4 | 0.88 |
| PTPMT1 | 0.2067 | 0.3992 | 1.57 | 0.61 | 0.0815 | 0.0957 | 1.72 | 0.67 |
| MX1 | 0.2550 | 0.4279 | 1.50 | 0.60 | <0.0001 | <0.0001 | 25.5 | 0.93 |
| KRTDAP | 0.6183 | 0.7647 | 0.88 | 0.54 | 0.0034 | 0.0057 | 0.49 | 0.77 |
| OGN | 0.0670 | 0.1852 | 0.81 | 0.66 | <0.0001 | <0.0001 | 0.20 | 0.91 |
| A2ML1 | 0.1499 | 0.3202 | 0.76 | 0.63 | 0.0329 | 0.0429 | 0.65 | 0.70 |
| EDEM2 | 0.2844 | 0.4435 | 0.74 | 0.59 | 0.1983 | 0.2273 | 1.85 | 0.62 |
| ABI3BP | 0.0230 | 0.0981 | 0.71 | 0.69 | <0.0001 | <0.0001 | 0.16 | 0.92 |
| PRELP | 0.0160 | 0.0981 | 0.70 | 0.71 | <0.0001 | <0.0001 | 0.26 | 0.93 |
| CSTA | 0.2925 | 0.4435 | 0.69 | 0.59 | 0.4850 | 0.5066 | 1.12 | 0.57 |
| SPRR3 | 0.0811 | 0.1907 | 0.62 | 0.65 | 0.3964 | 0.4234 | 2.07 | 0.58 |
| Characteristic | Stage of the Study | ||||
|---|---|---|---|---|---|
| iTRAQ Analysis | Verification by PRM | IHC Analysis | ELISA Serum Testing | ||
| Age–years | Median | 70.0 | 70.8 | 70.4 | 72.6 |
| Range | 45.3–93.9 | 45.3–94.2 | 37.1–93.9 | 41.8–87.9 | |
| FIGO stage | I | 19 | 46 | 54 | 35 |
| II | 2 | 5 | 3 | 4 | |
| III | 7 | 24 | 29 | 22 | |
| IV | 0 | 1 | 0 | 3 | |
| Histological grade | G1 | 12 | 23 | 34 | 17 |
| G2 | 9 | 28 | 37 | 28 | |
| G3 | 6 | 12 | 12 | 6 | |
| N/D | 1 | 13 | 6 | 13 | |
| hrHPV tumor status | positive | 13 | 38 | 36 | 31 |
| negative | 15 | 27 | 32 | 6 | |
| N/D | 0 | 11 | 18 | 27 | |
| Disease progression | d-fVSCC | 16 | 25 | 50 | 33 |
| progVSCC | 12 | 26 | 34 | 31 | |
| prosVSCC | 0 | 25 | 3 | 0 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fatalska, A.; Rusetska, N.; Bakuła-Zalewska, E.; Kowalik, A.; Zięba, S.; Wroblewska, A.; Zalewski, K.; Goryca, K.; Domański, D.; Kowalewska, M. Inflammatory Proteins HMGA2 and PRTN3 as Drivers of Vulvar Squamous Cell Carcinoma Progression. Cancers 2021, 13, 27. https://doi.org/10.3390/cancers13010027
Fatalska A, Rusetska N, Bakuła-Zalewska E, Kowalik A, Zięba S, Wroblewska A, Zalewski K, Goryca K, Domański D, Kowalewska M. Inflammatory Proteins HMGA2 and PRTN3 as Drivers of Vulvar Squamous Cell Carcinoma Progression. Cancers. 2021; 13(1):27. https://doi.org/10.3390/cancers13010027
Chicago/Turabian StyleFatalska, Agnieszka, Natalia Rusetska, Elwira Bakuła-Zalewska, Artur Kowalik, Sebastian Zięba, Agnieszka Wroblewska, Kamil Zalewski, Krzysztof Goryca, Dominik Domański, and Magdalena Kowalewska. 2021. "Inflammatory Proteins HMGA2 and PRTN3 as Drivers of Vulvar Squamous Cell Carcinoma Progression" Cancers 13, no. 1: 27. https://doi.org/10.3390/cancers13010027