IL-12 Family Cytokines in Cancer and Immunotherapy



1
Lineberger Comprehensive Cancer Center, The University of North Carolina at Chapel Hill School of Medicine, Chapel Hill, NC 27599, USA
2
Department of Genetics, The University of North Carolina at Chapel Hill School of Medicine, Chapel Hill, NC 27599, USA
*
Author to whom correspondence should be addressed.
Cancers 2021, 13(2), 167; https://doi.org/10.3390/cancers13020167
Submission received: 1 December 2020
/
Revised: 22 December 2020
/
Accepted: 29 December 2020
/
Published: 6 January 2021
(This article belongs to the Special Issue Challenges and Opportunities for Effective Cancer Immunotherapies)
{kind=link}
{kind=link}
Abstract
:Simple Summary
The IL-12 family cytokines play an important role in regulating the tumor immune contexture. Recent efforts geared towards the development of better immune therapeutic approaches have identified the need to overcome immune suppression and improve the quantity and quality of anti-tumor effector immune cells within the tumor milieu. In this review, we summarize the recent findings on IL-12 family cytokines in regulating anti-tumor immunity as well as the effectiveness and benefits of enhancing anti-tumor immunity in pre-clinical and clinical settings by targeting IL-12 family cytokines.
Abstract
The IL-12 family cytokines are a group of unique heterodimeric cytokines that include IL-12, IL-23, IL-27, IL-35 and, most recently, IL-39. Recent studies have solidified the importance of IL-12 cytokines in shaping innate and adaptive immune responses in cancer and identified multipronged roles for distinct IL-12 family members, ranging from effector to regulatory immune functions. These cytokines could serve as promising candidates for the development of immunomodulatory therapeutic approaches. Overall, IL-12 can be considered an effector cytokine and has been found to engage anti-tumor immunity by activating the effector Th1 response, which is required for the activation of cytotoxic T and NK cells and tumor clearance. IL-23 and IL-27 play dual roles in tumor immunity, as they can both activate effector immune responses and promote tumor growth by favoring immune suppression. IL-35 is a potent regulatory cytokine and plays a largely pro-tumorigenic role by inhibiting effector T cells. In this review, we summarize the recent findings on IL-12 family cytokines in the control of tumor growth with an emphasis primarily on immune regulation. We underscore the clinical implications for the use of these cytokines either in the setting of monotherapy or in combination with other conventional therapies for the more effective treatment of malignancies.
1. Introduction. IL-12 Family Cytokines: Composition, Signaling and Mechanism of Action
The IL-12 family cytokines are known to play essential roles in regulating innate and adaptive immune responses [1]. The functions of the IL-12 family cytokines have been widely studied in the settings of infection and auto-inflammatory diseases. The ability of these cytokines to modulate immune responses in cancer has been of significant interest. IL-12 family cytokines are typically secreted by innate immune cells but can also be secreted by adaptive immune cells depending on the disease and immune contexture. The members of this cytokine family are well known for shaping adaptive immune responses [2,3]. Due to their broad-spectrum roles in regulating immune responses, the IL-12 family cytokines are recognized as promising candidates for the modulation of anti-tumor immunity.
1.1. IL-12
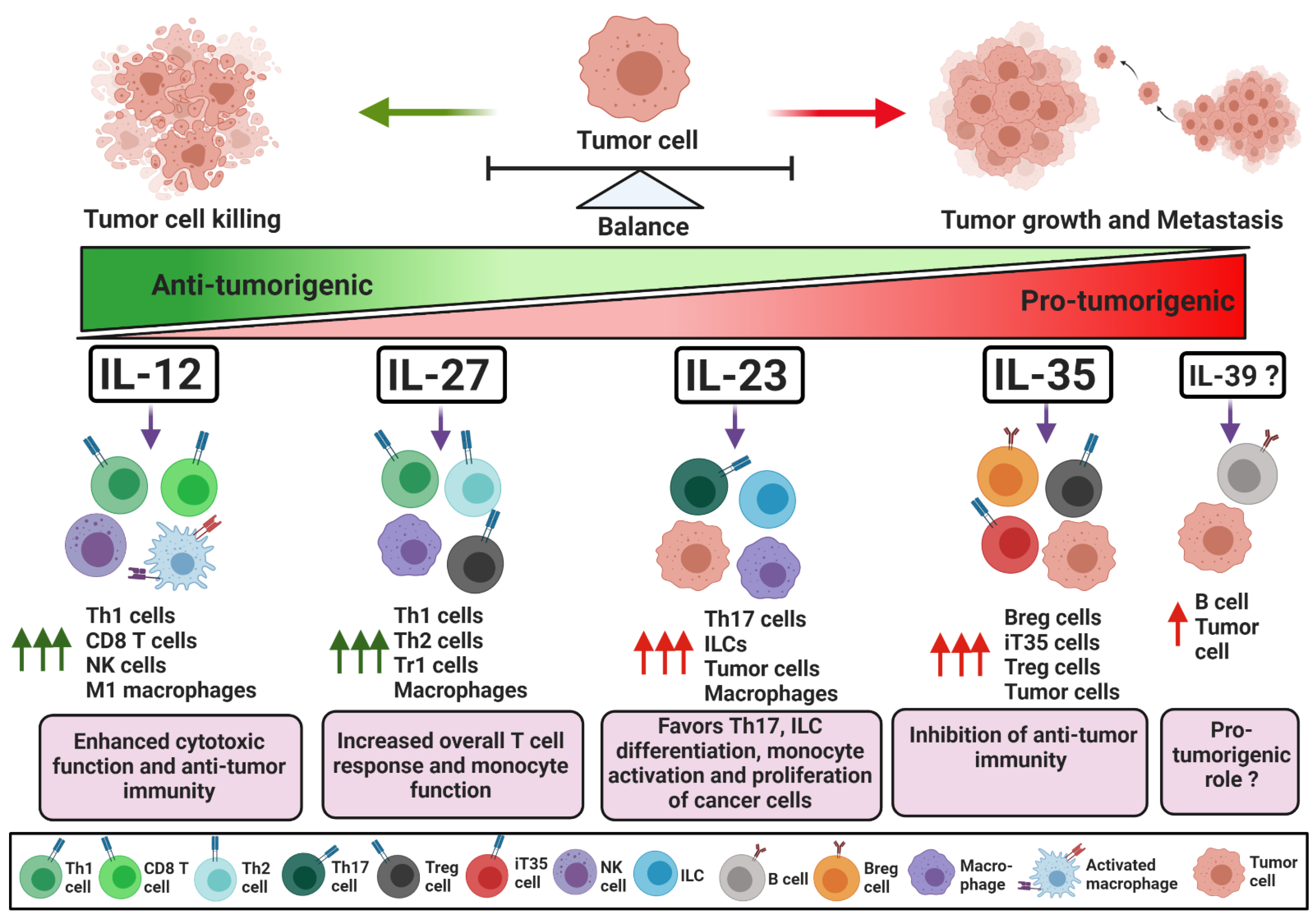
IL-12 is a heterodimeric cytokine composed of p40 and p35 subunits and is considered a largely pro-inflammatory cytokine (Figure 1). It is produced by antigen-presenting cells, such as dendritic cells and macrophages, and is crucial for the recruitment and effector functions of CD8+ T and NK cells [4]. Therefore, IL-12 is a major contributor to effective anti-tumor immune responses [5]. IL-12 signals through IL-12Rβ1 and IL-12Rβ2 receptors expressed on target cells, which allow downstream Jak2 and Tyk2 to promote the phosphorylation of and homo-dimerization of STAT4. The homodimer of pSTAT4 binds to its target genes and regulates gene expression [1]. In CD4+ T cells, STAT4 activation by IL-12 is required for the transcription of T-bet, a positive regulator of Th1 cell differentiation. T-bet enhances the expression of Th1-specific cytokines, chemokines, and Th1’s associated receptors. T-bet alone can positively regulate the expression of IFN-γ, while in combination with STAT4, it enhances transcription of CXCR3, IL-12Rβ1, CCL3 and CCL4 [6,7,8]. CCL3 and CCL4 are required for the intra-tumoral recruitment of cytotoxic NK cells and CD8+ T cells [9,10,11]. In the presence of IL-12, NK cells are activated, express CD69 and CD25, and can further proliferate in the tumor niche [12,13]. Activated Th1 and NK cells proliferate and infiltrate into the tumor, where Th1 cells support the effector functions of tumor-specific cytotoxic T cells [4,12,13]. The IFN-γ, granzyme, and perforin secreted by cytotoxic NK and CD8+ T cells can induce the apoptosis of cancer cells and control tumor growth. Moreover, IL-12 facilitates antigen presentation by upregulating MHCI on tumor cells, favoring polarization to M1 macrophages and attracting effector immune cells by enhancing the production of the chemokines CXCL9, CXCL10 and CXCL11 [5,14,15,16]. Additionally, T-bet and STAT4 act as negative regulators for RORγt and Foxp3, transcription factors responsible for Th17 and Treg generation, respectively, and limit their proliferation in the tumor microenvironment (TME) [17,18,19,20]. IL-12 can also neutralize signaling by negative regulatory receptors on CD8+ T cells. For example, IL-12 downregulates PD-1 and IFNγR2 expression on CD8+ T cells, protecting tumor infiltrating CD8+ T cells from IFN-γ-induced cell death [21]. The activation of anti-tumor immunity by anti-PD1 requires IL-12-mediated crosstalk between T cells and dendritic cells that enables CD8+ T cell-mediated tumor cell killing [22].
Besides its function in effector immune cells, IL-12 alters the plasticity of terminally differentiated Treg cells by converting Foxp3+ Treg cells to IFN-γ-producing Foxp3+ T cells. Treatment with IL-12 diminishes the level of IL-2, which is required for Treg cell survival and expansion [23]. IL-12 was shown to stimulate the IFN-γ-mediated inhibition of mouse Treg cell expansion. Mechanistically, IL-12-induced IFN-γ signaling causes cell cycle arrest in Treg cells and inhibits tumor-induced Treg cell proliferation. [23,24]. These studies demonstrate that IL-12 is not only required for the activation of effector anti-tumor immune responses but can also directly inhibit immune suppression. Thus, the use of IL-12 as a cancer immunotherapy could be beneficial in controlling tumor growth by activating anti-tumor cytotoxic immune responses. Overall, IL-12 targets and modulates T cells, NK cells and antigen-presenting cells (APCs) that regulate the fate of the anti-tumor immune response against the cancer cells.
1.2. IL-12 in Cancer Immunotherapy
Cytokine-based immunotherapy can be effective in the treatment of numerous malignancies. IL-12 can be considered a strong candidate for immunotherapy-based interventions, as it potentiates tumor-specific cytotoxic NK and CD8+ T cells that are largely responsible for tumor cell killing. However, the systemic administration of IL-12 is quite toxic; therefore, alternative methods of IL-12 delivery and/or the activation of T cells by IL-12 are needed [25,26]. To that extent, a recent report by Nguyen et al. provided comprehensive updates on the development and application of a localized delivery strategy for IL-12-based immunotherapy [27]. Wang et al. reported that systemic delivery via an oncolytic adenovirus encoding IL-12 lacking the signaling peptide reduced the toxic side effects and enhanced survival in mouse models of pancreatic cancer [26]. The nanoparticle-mediated delivery of IL-12 has also enhanced the cytotoxic activity against human hepatocellular carcinoma cells [28]. The in vitro nanoparticle-mediated delivery of IL-12 specifically into naïve CD8+ T cells favored their expansion and the activation of the effector phenotype [28]. Several studies have shown that IL-12 favors the survival and differentiation of naïve CD8+ T cells towards the effector phenotype. IL-12 acts as an anti-apoptotic factor for CD8+ T cells by the impediment of the activation-induced cell death of CD8+CD62Lhi naïve CD8+ T cells, increased T cell homing, and has shown sustainable anti-tumor activity against mouse models of melanoma [29]. These findings suggest that the priming of naïve CD8+ T cells with IL-12 prior to adoptive cell therapy could increase their effectiveness and anti-tumor activity. Similar preclinical studies have reported the synergistic use of IL-12 with adoptive T-cell-based immunotherapies. In a syngeneic mouse model, the administration of anti–VEGFR-2 chimeric antigen receptor (CAR) and IL-12–co-transduced T cells directly into the tumor site modified the immune suppressive environment by ablating systemic and intra-tumoral VEGFR-2+ myeloid-derived suppressor cells (MDSCs) [30]. Moreover, genetically engineered T cells, which expressed high levels of IL-12, were therapeutically effective against established murine B16 melanoma tumors, even in the absence of a tumor vaccine and IL-2 [31]. Although the survival of IL-12 engineered cells was low compared to endogenous T cells, they demonstrated improved functionality, were detected at a higher frequency in the melanoma, and maintained the activity of endogenous NK and CD8+ T cells. In clinical trials, IL-12 was shown to have anti-cancer activity against human glioma [32]. In this case, 31 high-grade glioma patients were treated with human IL-12 vector (Ad-RTS-hIL-12) in a multicenter phase 1 dose-escalation trial (NCT02026271), and showed evidence of increased IFN-γ and PD-1+ tumor-infiltrating lymphocytes [32]. These findings suggest that increases in the concentration of intra-tumoral IL-12 improve the efficacy of adoptive T cell therapies.
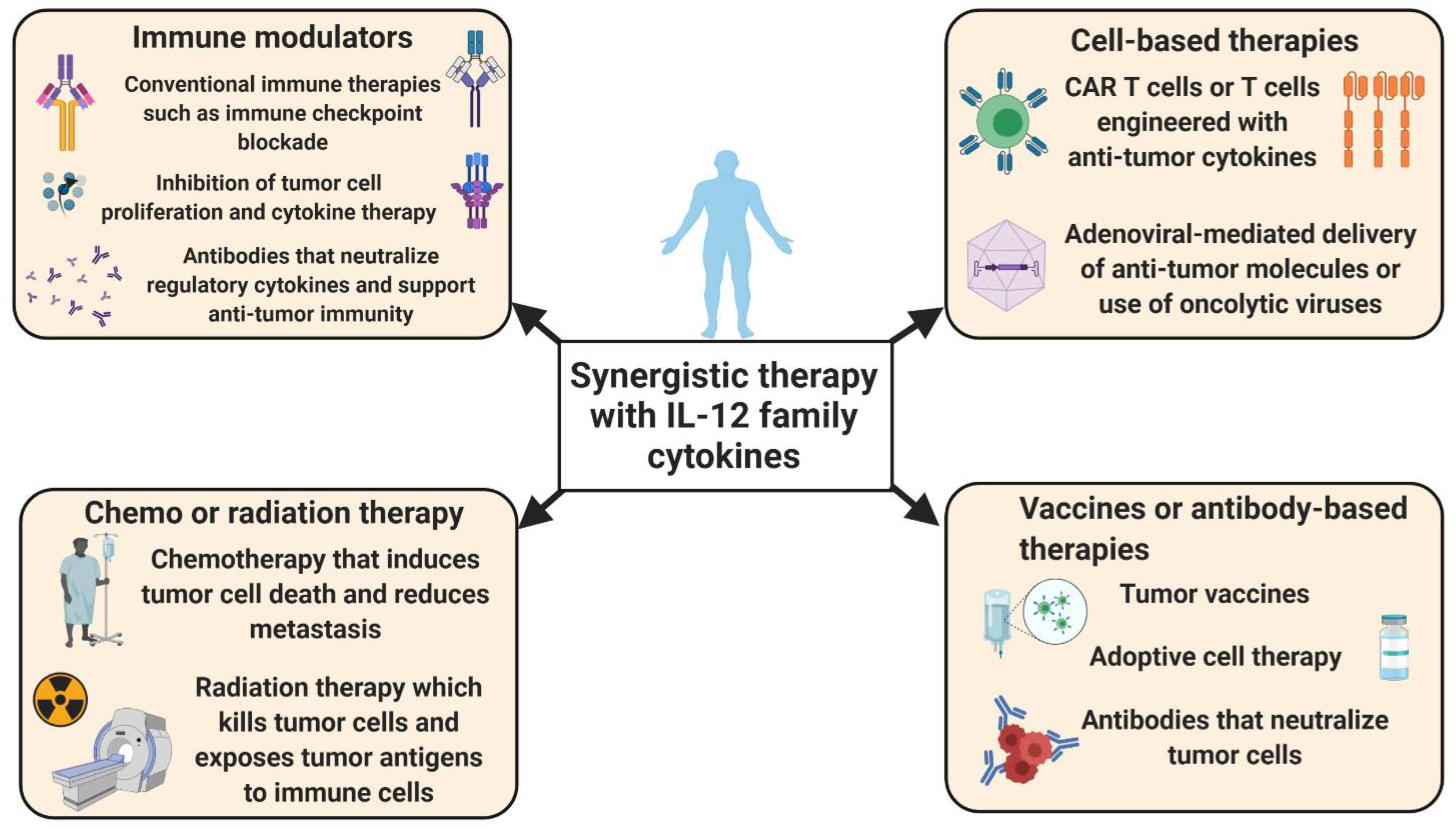
Another recent advance in potentiating T cell-based anti-tumor activity has been increasing the load of IL-12 in CAR T cells. In a recent pre-clinical mouse study, Kueberuwa et al. used CAR T cells expressing IL-12 and showed that modified CAR T cells were able to cure B cell lymphoma and improve the long-term survival rate [33]. In this setting, IL-12-engineered CAR T cells recruited host immune cells to elicit an anti-tumor immune response. In addition, IL-12-producing CAR T cells directed against ovarian cancer cells showed robust proliferation and secretion of IFN-γ, which resulted in increased survival in a mouse model [34]. Similar preclinical studies in a hepatocellular carcinoma model indicated that IL-12-expressing CAR T cells produced high levels of effector cytokines, accompanied by attenuated Treg cell infiltration and induced tumor cell lysis [35]. In summary, these observations reveal that the inducible expression of IL-12 improves the anti-tumor functions of CAR T cells and might provide a promising treatment strategy for cancer patients (Figure 2). Nevertheless, CAR T or TCR T cells expressing IL-12 resulted in severe, edema-like toxicity and increased serum levels of IFN-γ and TNF-α in mice with melanoma [36]. Additionally, IL-12 overexpressing CAR T cells can lead to cytokine release syndrome (CRS) and cause systemic inflammatory responses in patients treated with CAR T cells [37,38]. These severe reactions should be considered before the use of genetically engineered T cells, particularly IL-12-producing CAR T cells.
While conventional immune checkpoint blockade, such as anti-PD-L1, is commonly used for the treatment of many cancers, its efficacy is not universal, and combination therapies that augment T cell responses are needed. Hewitt et al. demonstrated that intra-tumoral IL-12 mRNA (MEDI1191) therapy was able to stimulate IL-12 production within the tumor milieu without toxic effects [39]. MEDI1191 is currently being assessed in a phase I trial in patients with solid tumors (NCT03946800). The combination of this approach with anti-PD-L1 enhanced anti-tumor immunity by promoting IFN-γ+ Th1 cell differentiation. Fallon et al. engineered the fusion of two molecules of murine IL-12 (NHS-muIL12) with a longer half-life than recombinant murine IL-12 [40]. The combination of NHS-muIL12 and anti-PD-L1 boosted T cell activation and effector function within the TME and augmented tumor regression in murine tumor models. Other immunotherapy combinations could also be used. For example, combined immune therapy with IL-12 and tumor necrosis factor-related apoptosis-inducing ligand (TRAIL) in humanized mouse models of hepatocellular carcinoma increased the infiltration of IFN-γ-producing NK cells and promoted the apoptosis of cancer cells. Additionally, in the presence of IL-12, TRAIL enhanced MHCI expression on antigen-presenting cells and downregulated the expression of intra-tumoral vascular endothelial growth factor (VEGF) and CD31 [41]. Another study showed that a combination of the immune-modulating protein aggregate magnesium–ammonium phospholinoleate–palmitoleate anhydride (P-MAPA) and human rIL-12 significantly reduced the migratory potential and invasion capacity by inducing the apoptosis of ovarian cancer cells [42]. These findings reveal that the use of IL-12 could substantially increase the effectiveness of cancer immunotherapy.
IL-12 may also have a beneficial role in synergizing with chemotherapy treatments. In patients with metastatic HER2+ cancers, a synergistic treatment of IL-12 with chemotherapy, such as trastuzumab, stimulated NK cell activity [43]. Similar synergistic approaches could be used with radiation therapy. Deplanque et al. showed that radiation-induced immune suppression in well-established tumors could be overcome by increases in IL-12-dependent Th1 responses in mouse models of colon cancer [44]. Recent reports showed that radiation therapy in combination with IL-12 induced clonal epitope-specific T cell expansion and infiltration, blocked tumor growth and the improved survival of animals with human rhabdomyosarcoma xenografts [45]. Human recombinant IL-12 was also shown to have a protective role in cancer patients treated with radiation therapy. In this setting, it diminished the complications that can arise from radiation therapy, such as severe myelosuppression or pancytopenia [46]. In murine pancreatic cancer, immunotherapy using IL-12+ microspheres in combination with stereotactic body radiation therapy induced intra-tumoral IFN-γ production, repolarized myeloid suppressors, promoted robust T cell activation and efficiently eliminated established liver metastases [47]. Overall, these observations reveal the importance of IL-12-based therapies in the initiation and stimulation of anti-tumor immune responses.
1.3. IL-23
IL-23 is a heterodimeric cytokine made up of p40 and p19 subunits. In cancer, IL-23 has been shown to have both pro- and anti-tumorigenic roles (Figure 1). Here, we discuss the dual role of IL-23 in modulating effector and/or regulatory immune responses in cancer.
1.3.1. IL-23 as a Suppressor of Anti-Tumor Immunity
The pro-tumorigenic role of IL-23 was first reported by Langowski et al., where it was observed that the genetic deletion or blockade of IL-23 in mice led to an increased infiltration of cytotoxic T cells with protective effects against cancer [48]. IL-23 can be secreted by dendritic cells, monocytes, neutrophils and innate lymphoid cells (ILCs) [49,50,51,52]. Upon stimulation, macrophages, dendritic cells and neutrophils have been shown to secrete IL-23 [53]. For example, IL-6, VEGF, CCL22 and/or PGE2 produced by tumor cells recruited tumor-associated macrophages, which in turn produced IL-23 and maintained suppressive Treg activity in the TME [54,55,56,57,58]. IL-23 produced by tumor-educated neutrophils activated downstream AKT and p38 pathways in mesenchymal stem cells and transformed those mesenchymal stem cells into cancer-associated fibroblasts [59]. CXCL5 could also induce the expression of IL-23 in neutrophils to enable the enhanced migration and invasion of gastric cancer cells [60]. Microbial products could also activate intra-tumoral myeloid cells, which in turn produced IL-23 and promoted colorectal neoplasms in mice [61]. Similar studies by Jin et al. reported that microbiota stimulated myeloid cells to induce IL-23 production in mouse models of lung cancer [62].
The role of IL-23 in regulating Th17 cell differentiation is widely studied and has been shown to play a critical role in Th17 cell expansion and the maintenance of the Th17 phenotype. IL-23 is required for the stabilization of RORγt, a transcription factor for Th17 cell generation, and facilitates the secretion of the effector cytokines IL-17 and IL-21 by Th17 cells [63,64]. IL-23 also stimulates IL-17 production from γδ T cells, which can support tumor growth [62,65,66,67]. IL-23 signals through IL-12Rβ1 and IL-23R in target cells and activates Jak2 and Tyk2, resulting in the phosphorylation of STAT3 and STAT4 [1]. IL-23R signaling in CD4+ T cells activates STAT3, which in turn stabilizes RORγt expression and enhances IL-17 gene transcription [63,64]. IL-23 or IL-23-induced IL-17 signaling also activates NFĸB, which regulates genes responsible for the recruitment of tumor-associated macrophages (TAMs) and MDSCs in the TME [68,69,70].
Thus, IL-23 is recognized as mediator of the Th17 response and plays an important role in shaping immune responses towards cancer. For example, CCL20 produced by tumor cells cooperated with stromal IL-23 to recruit IL-17-producing Th17 cells in the TME [71,72,73,74]. IL-17, in turn, promoted tumor angiogenesis by inducing angiogenic factors, such as VEGF and PGE2, and activated oncogenic STAT3 signaling, which was essential for the expression of pro-survival and pro-angiogenic genes [72,73,74].
IL-23 can also enhance inflammation in the TME and promote tumorigenesis independently of IL-17. The elevated expression of IL-23R on tumor cells enhanced tumor-associated inflammation and promoted the development and metastasis of cancer [75,76]. As one example, IL-23 induced the activation of STAT3 in lung cancer cells expressing IL-23R and stimulated their proliferation [77]. The expression of IL-12Rβ2 and IL-23R on laryngeal tumor cells mediated crosstalk between the cancer cells and tumor-infiltrating lymphocytes and affected the prognosis of laryngeal cancer patients [78]. Furthermore, the expression of IL-23R on B-acute lymphoblastic leukemia (B-ALL) cells in B-ALL patients was increased compared to normal B-lymphocytes and acted to upregulate miR15a and downregulate the pro-survival factor BCL2 [79]. These results indicate that, apart from regulating the Th17 response, IL-23 can also potentially alter the fate and function of cancer cells.
1.3.2. IL-23 as an Activator of Anti-Tumor Immunity
IL-23 is known to activate and maintain the expression of Th17-specific transcription factors, and these transcription factors can negatively regulate the immune-suppressive Treg cell response. IL-23 signaling can repress Treg differentiation and maintain the active transcription of IL-17, IL-21, IL-22 and IL-23R [63,64]. IL-23 can also regulate the function of diverse subsets of immune cells in the TME. For example, IL-23 upregulated the expression of IL-23R on type 3 innate lymphoid cells (ILC3), granulocytes and NK cells, which in turn induced their pro-inflammatory cytokine production and cytotoxic function [80,81,82]. In the presence of IL-23, intraepithelial cells secreted IL-22, which augmented their barrier function and maintained the CD4+ T cells’ effector phenotype [83,84,85]. On the other hand, activated fibroblasts secreted CCL5 and MCP-1, which recruited IL-23-producing macrophages [86,87]. IL-23+ macrophages expanded anti-tumorigenic Th17 cells that had a distinct phenotype known as Th1-like Th17 cells and promoted tumor-specific immune responses by secreting IFN-γ, CXCL9 and CXCL10 [86,87]. Thus, IL-23 and IL-17 have been shown to play multipronged roles in tumorigenesis by activating or inhibiting effector anti-tumor immunity. These findings demonstrate that IL-23 could modulate pro- as well as anti-tumor immune responses (Figure 1).
Overall, IL-23 uses different downstream signaling pathways in tumor and immune cells that regulate pro-inflammatory and anti-inflammatory pathways. Overall, context-dependent functions of IL-23 in cancer could drive the development of inhibitor or augmentative therapy axes, respectively.
1.4. IL-23 in Cancer Immunotherapy
IL-23 is an important player in inflammatory responses, and IL-23-producing and responding cells are highly abundant in the tumor milieu [88,89]. In fact, the abundance of IL-23 in tumors is a general feature of cancer [85,88,90]. The levels of IL-23 and IL-23R were found to be significantly higher in breast cancer tissues and positively correlated with the patient’s tumor size, TNM stage and metastasis [90]. In colorectal cancer, the levels of IL-23 in patients’ serum samples were found to gradually increase with tumor stage progression [91,92]. IL-23 could activate Th17, ILC3, granulocytes, NK cells and intra-epithelial lymphocytes, which exaggerated gut inflammation and promoted the growth of colon cancer [93]. In multiple myeloma patients, IL-23 induced IL-17 and RORC in the bone marrow microenvironment [94]. Similar studies reported that IL-23 could promote IL-17-mediated tumorigenesis by converting type 1 innate lymphoid cells (ILC1) into ILC3, resulting in poor prognosis for patients with lung carcinoma [95]. Moreover, the overexpression of IL-23 mRNA was observed in serum samples of breast cancer patients, where IL-23 played a pro-tumorigenic role by upregulating the expression of regulatory cytokines [96]. Liu et al. observed elevated levels of serum IL-23 from hepatocellular carcinoma patients, and this was associated with poor clinical outcomes [97]. They demonstrated that ILC3 cells were the main source of IL-23 in hepatocellular carcinoma, and these cells directly suppressed the CD8+ T cell response by enhancing apoptosis and preventing the proliferation of CD8+ T cells [97].
Prostate cancer patients can exhibit resistance to androgen-deprivation therapy, a condition known as castration-resistant prostate cancer (CRPC). Increased levels of IL-23 were also observed in blood and tumor samples of patients with CRPC [98]. The major source of IL-23 was shown to be MDSCs. In a mouse model of CRPC, the blockade of IL-23 improved sensitivity to androgen deprivation and synergized with conventional therapies to boost anti-tumor immunity [98]. Another study showed that elevated IL-23 was associated with poor survival in renal cell carcinoma patients [99]. Mechanistically, intra-tumoral IL-23+ macrophages augmented both Treg cell proliferation and IL-10/TGF-β secretion and suppressed the cytotoxic functionality of CD8+ T cells. The blockade of IL-23 promoted CD8+ T cell cytotoxicity and improved the overall survival of mice. Moreover, the blockade of IL-23 together with immune checkpoint blockade augmented therapeutic benefits [99]. Therefore, blockade of IL-23 in combination with immune checkpoint inhibitors can be a potential therapeutic approach in several cancer types (Figure 2). Conversely, reports from pancreatic cancer patients found that the levels of IL-23 decreased with tumor progression, and long-term survivors had increased IL-23 expression [100,101]. Similarly, IL-23 was shown to play a dual role in premalignant oral lesions to cancer in mice, where IL-23 was pro-inflammatory in premalignant oral lesions and inhibitory as the lesions progressed to cancer [102]. Thus, IL-23 may have distinct effects on anti-tumor immunity [103,104].
Sheng et al. showed that IL-23 can also enhance metastasis through the upregulation of anti-apoptotic factors in cancer cells [90]. In this case, the neutralization of IL-23p19 reduced cell proliferation and induced cell apoptosis by reducing the expression of BCL2 in breast cancer cell lines [90]. Besides its effects on the survival of tumor cells, IL-23 induced the expression of the proliferative marker Ki67 and endothelial marker CD31, promoting tumor cell proliferation, mammary tumor growth and pulmonary metastasis in mouse models of breast cancer [105]. Apart from the role of IL-23 in modulating the tumor cell cycle and growth, IL-23 exerts its effects on immune cells within the TME. IL-23 signaling stimulates the expression of regulatory genes, such as IL-10, TGF-β and VEGF, and increases the infiltration of immunosuppressive M2 macrophages and neutrophils that reduce the ability of effector CD4+ and CD8+ T cells to infiltrate tumors [105,106]. Zhang et al. showed that IL-23 is involved in the formation of immune-tolerant and pro-angiogenic TME through the induction of IL-17; this promoted the development of invasive prostate adenocarcinomas in mice [107]. These studies revealed the effects of IL-23 on the modulation of both immune cells and cancer cells. Thus, neutralizing IL-23 may augment tumor-specific immune responses and may control tumor cell proliferation and metastasis in certain cancer types.
Some preclinical studies have shown that IL-23 depletion combined with CAR T cells promotes tumor suppression, and neutralizing IL-23 can be effective particularly in combination with CAR T cell therapy [108]. The monoclonal antibody (mAB)-mediated depletion of IL-23 in mouse models of prostate cancer reinstated sensitivity to androgen deprivation therapy [98]. In one preclinical study, depleting IL-23mAB therapy in combination with CAR T cell therapy showed promising effects in the treatment of prostate cancer. The CAR T cells were generated by dual targeting using an IL-23-specific antibody and prostate-specific membrane antigen (PSMA)-specific mAB. IL-23mAB/PSMA CAR T cells had greater effectiveness than PSMA CAR only in the suppression of prostate cancer growth. Mechanistically, treatment with IL-23mAB/PSMA-CAR significantly enhanced CD45RO+CD8+ and CD127+CD4+ CAR T cells and indicated a potential role for IL-23 in the efficacy of CAR T cell therapy against prostate cancer [108,109]. By contrast, IL-23-expressing CARs were also shown to have anti-tumorigenic properties in preclinical studies. Ma et al. engineered the expression of the p40 subunit of IL-23 in T cells (p40-Td cells), and this enhanced the proliferation of activated T cells via autocrine signaling [110]. They also demonstrated in xenograft and syngeneic mouse models that p40-Td CAR T cells had enhanced anti-tumor capacity, with increases in granzyme B and decreases in PD-1 expression. Using neuroblastoma and pancreatic cancer models, they observed that p40-Td CAR T cells had greater efficacy in comparison to unmodified CAR T cells and exhibited diminished side effects in comparison to IL-18- or IL-15-producing CAR T cells [110]. Thus, the immune therapeutic strategies that target IL-23 may have use in the treatment of cancer.
1.5. IL-27
IL-27 is a heterodimeric cytokine consisting of EBi3 and p28 subunits [1]. IL-27 is generally involved in the differentiation and activation of different CD4+ T cell subsets (Figure 1). Antigen-presenting cells stimulated by tumor antigens and CD40 can secrete IL-27, which has a dual function in regulating immune responses against cancer [111].
1.5.1. IL-27 as a Suppressor of Anti-Tumor Immunity
IL-27 has been shown to have anti-inflammatory properties. For example, IL-27 activated STAT3 and stabilized Foxp3 expression and IL-10 transcription in T cells, leading to sustained immune suppression [112]. Such IL-10-producing T cells are known as Tr1 cells [112,113]. Tr1 cells are immunosuppressive and downregulate the cytotoxic activity of CD8+ T cells and NK cells, as well as preventing the production of TNF-α and IL-6 by monocytes, which are important for anti-tumor immunity [113,114]. Mechanistically, IL-27 signaling through STAT1 and STAT3 could activate the transcription factor c-Maf and induce the generation of Tr1 cells and IL-10 production by T cells [115,116,117,118]. IL-27 has also been shown to act as an anti-inflammatory cytokine by inhibiting Th2 and Th17 cell-specific immune responses and promoting the activation and proliferation of Treg cells [112,119,120]. Additionally, the activation of STAT1 and STAT3 by IL-27 can induce the expression of IL-1β, TNF-α and IL-18, which are important for the polarization of the immunosuppressive M2 phenotype [119,120]. In advanced non-small cell lung cancer (NSCLC) patients, IL-27 has also been shown to play a pro-tumorigenic role, where IL-27 induced tolerogenic dendritic cells that helped cancer cells to escape from immune surveillance [121].
1.5.2. IL-27 as an Activator of Anti-Tumor Immunity
On the other hand, IL-27 produced by dendritic cells, monocytes and macrophages following TLR activation may positively regulate Th1 differentiation through STAT1 dimerization. This subsequently leads to T-bet activation and Th1-specific gene expression [122,123,124]. IL-27 can also enhance the surface expression of MHCI and MHCII in monocytes as well as co-stimulatory CD80, CD86, and adhesion molecule CD54, overall promoting the pro-inflammatory activity of T cells [125]. IL-27 signals through IL-27R (WSX-1) and gp130 receptors on target cells [1]. IL-27R and gp130 receptors recruit Jak1 and Jak2, which in turn activate STAT1 and STAT3 [1]. IL-27 plays a critical role in the development of Th1 and IL-10-producing T cells. Th1 cells express T-bet and the effector cytokine IFN-γ through STAT1 dimerization induced by downstream IL-27 signaling [124,126]. In certain scenarios, the IL-27 receptor can also activate STAT4 and STAT5. For example, IL-27 signals through STAT1 and STAT4 to induce T-bet expression in CD4+ T cells, which results in the upregulation of intracellular IFN-γ [124]. At the same time, the activation of STAT5 enhances the proliferation of Th1 cells in the TME [122,127]. For example, signaling through WSX-1 and gp130 on CD8+ T cells activated STAT1 and STAT4 and resulted in the increased expression of granzyme, perforin and IFN-γ in a T-bet-dependent manner [128,129]. In NK cells, the activation of STAT1/STAT4 by IL-27 stimulated T-bet-dependent IFN-γ production and IL-27R expression [130,131]. These overall pro-inflammatory effects of IL-27 can stimulate anti-tumor immunity and contribute to tumor cell clearance [130,131,132]. Additionally, IL-27 may antagonize the production of IL-2, which is required for T cell activation, and regulated the intensity and duration of the effector CD4+ T cell response [133,134].
IL-27 signaling in innate and adaptive immune cells as well as in cancer cells regulates the expression of distinct key molecules important for the activation of immune responses in the tumor milieu. To this end, IL-27 was reported to enhance the expression of DNAM-1, NKG2D and CD69 on NK cells [131]. NK cells also secreted high amounts of perforin and granzyme B and showed enhanced cytotoxic activity against cancer cells [131]. Moreover, the synergistic action of IL-27 with IL-18 activated and increased NK cell proliferation, cytotoxicity and IFN-γ production [131]. Another important aspect of IL-27 in regulating the NK cell response against uterine endometrial cancer cell (UECC) was recently reported by Zhou et al. In this study, the production of IL-27 by uterine endometrial cancer cells in the presence of rapamycin upregulated the expression of WSX-1 and gp130 on NK cells and increased their cytotoxic activity against cancer cells in a xenograft mouse model [135]. IL-27 also reduced the proliferation of human ovarian cancer cells by activating STAT3 and inhibiting Akt signaling pathways [136]. In mouse models of colon carcinoma, IL-27 induced NK cell-mediated cytotoxicity and anti-tumor immunity by activating STAT3 and triggering the expression of perforins [137].
IL-27 can also potentiate anti-tumor immunity by supporting the differentiation and expansion of myeloid progenitor cells into anti-tumorigenic M1 macrophages. For example, IL-27 was shown to act on Sca-1+c-Kit+ cells in bone marrow and promoted their differentiation into M1 macrophages in preclinical mouse studies of B16F10 melanoma and MC38 colon adenocarcinoma [138]. IL-27 also enhanced the expression of inducible nitric oxide synthase, which sustained the anti-tumor activity of macrophages within the TME [138]. Similar studies in pancreatic cancer using co-cultures of human pancreatic cell lines and TAMs reported the role of IL-27 in restricting the differentiation of TAMs into immune suppressive M2 macrophages, which enhanced the effectiveness of gemcitabine [139]. Besides the role of IL-27 in regulating the differentiation of macrophages, IL-27 can also regulate several key surface molecules on cancer cells. Carbotti et al. showed that IL-27 triggered STAT1/STAT3 phosphorylation and upregulated the expression of surface MHC class I antigen on human small cell lung cancer cells, boosting the CD8+ T cell response [140]. In non-small cell lung cancer patients, IL-27 promoted the anti-tumor function of myeloid cells and suppressed epithelial-to-mesenchymal transition (EMT) in cancer cells [141]. In human prostate cancer cells, IL-27 upregulated TLR3 expression and enhanced TLR-mediated cell death [142]. A relationship between IL-27 genetic polymorphisms and cancer risk has been identified. The IL-27 2905T/G genotype is linked with a decreased susceptibility to and development of cervical cancer in patients [143]. IL-27 also hindered NSCLC and squamous cell carcinoma (SCC) growth in cooperation with granulocyte and macrophage-driven necrosis, CXCL3 production, and diminished EMT-related gene expression [141]. These results indicate that IL-27 has great potential and can be utilized in the development of novel immune-therapeutic approaches.
1.6. IL-27 in Cancer Immunotherapy
Given the ability of IL-27 to regulate both pro-tumorigenic and anti-tumorigenic responses, the choice of the augmentation or blockade of IL-27 should be carefully evaluated given the tumor type and the immune contexture. Recent efforts have made therapeutic use of recombinant IL-27, either alone or in combination with other therapies. Majumder et al. demonstrated that treatment with rIL-27 ameliorated angiogenesis and immunosuppression in mouse models of benzo(a)pyrene (BaP)-induced lung carcinogenesis [144]. Zhu et al. also used an IL-27-expressing recombinant adeno-associated virus (AAV-IL-27) for the treatment of mouse models of lung cancer [145]. Such treatment significantly reduced tumor growth and enhanced the T cell response by downregulating the Treg cell frequency in the peripheral blood and in the TME [145]. These approaches could supplement current methods of Treg depletion, as attempts for total Treg cell depletion in cancer can result in autoimmunity. Furthermore, AAV-IL-27 drastically increased the efficacy of immune checkpoint blockade and Granulocyte Macrophage Colony Stimulating Factor (GM-CSF) vaccine treatment with no significant adverse effects [145]. Similar studies showed the recombinant adeno-associated virus (rAAV)-mediated delivery of IL-27 reduced Treg frequency and increased the effectiveness of immunotherapeutic agents in different mouse tumor models [146]. In this case, the intra-tumoral injection of AAV-IL-27 robustly increased anti-tumor immunity against plasmacytoma J558 and B16F10 mouse tumors by inducing the expression of CXCR3 in T cells and increasing responsiveness to anti-PD-1 or T cell adoptive transfer therapy [146]. These observations indicate that the intra-tumoral administration of IL-27 in combination with immune checkpoint blockade or adoptive T cell therapy could be a suitable approach for cancer treatment.
With regards to tackling the potential pro-tumor activity of IL-27, several studies have shown that IL-27 can regulate the immunosuppression and exhaustion of T cells. For example, IL-27 could also induce indoleamine 2,3-dioxygenase (IDO) and PD-L1 expression on monocytes and tumor-associated macrophages and on human prostate, breast and lung cancer cells [147,148,149,150]. Moreover, IL-27 signaling in human ovarian cancer cells activated STAT1/STAT3 and induced the constitutive expression of IDO [149]. These alterations on monocytes and tumor cells potentiate escape from immune attack. Similar studies in human lymphoma cell lines showed that the enhancement of IL-27 expression by tumor cells triggered the surface expression of PD-L1 on tumor-associated macrophages [151]. Moreover, IL-27 signaling on tumor cells activated STAT1; upregulated the expression of PD-L1 and production of IL-10, TGF-β and IDO; and downregulated MHCI expression [111,152]. Finally, IL-27 signaling in NK and CD8+ T cells upregulated the cell surface expression of negative regulators, such as LAG3 and Tim3, contributing to the exhaustion of the effector phenotype [153,154]. In mouse models of melanoma, the inhibitory receptor Tim-3 was a crucial regulator of T cell dysfunction in tumorigenesis. IL-27 was shown to induce the expression of nuclear factor interleukin 3 regulated (NFIL3), which in association with T-bet, induced the expression of Tim-3 and the immunosuppressive cytokine IL-10 [153]. In this case, the blockade of IL-27 signaling resulted in reduced NFIL3 and Tim-3 expression, and decreased T cell exhaustion [153]. Similar studies reported pro-tumorigenic action of IL-27 favoring the expansion of Treg cells, where IL-27R−/− Treg cells were unable to suppress anti-tumor immunity in mouse models of melanoma [155]. Furthermore, IL-27 secreted by tumor-infiltrated neutrophils upregulated CD39 expression and enhanced the immune suppressive capacity of CD163+ macrophages [156]. Here, the depletion of IL-27 attenuated the expression of PD-L1 and IL-10 in macrophages isolated from ovarian cancer patients [156]. These studies indicate that the IL-27/STAT3 axis can be a potential target for immunotherapy.
1.7. IL-35
IL-35 is a recent addition to the IL-12 family of cytokines and is composed of the p35 and EBi3 subunits [1,3]. IL-35 is a potent immune suppressive cytokine produced mostly by regulatory B (Breg) cells, iT35 cells and Treg cells and can play an important role in the suppression of effector immune responses [157]. IL-35 has also recently been detected in macrophages, dendritic cells and tumor cells [158,159,160]. The role of IL-35 in tumor growth is becoming more prominent due to its ability to inhibit effector immune responses (Figure 1). IL-35 regulates the differentiation of CD4+ T cells and strongly favors regulatory T cell fate [161]. IL-35 is also thought to regulate the Th1 and Th17 lineage-specific transcription factors T-bet and RORγt as well as cytokines IFN-γ and IL-17 [162,163,164]. For example, IL-35 suppressed the effector functions of CD4+ cells and favored tumor growth by facilitating the exhaustion of CD8+ T cells [165,166]. Additionally, IL-35 restricted CD8+ T cell activation by suppressing the expression of the costimulatory surface molecule CD28, and Th1 cytokine production, and reduced cytolytic functions by repressing the expression of perforins [167,168]. Furthermore, IL-35 converted effector T cells into IL-35-producing T cells known as iT35 cells and activated IL-35 production and the proliferation of Treg cells [163,169,170]. A subset of mouse and human regulatory B cells can be a major source of IL-35 in certain malignant tumor types [167,171,172]. Additionally, IL-35 can enhance its own expression in B cells through IL-12Rβ2 and IL-27Rα and the activation of STAT1 and STAT3, resulting in the augmented formation of Breg cells [173,174]. Mechanistically, the loss of IL-35 hindered pancreatic tumor growth via increases in effector CD4+ T cells and CD8+ T cells within the TME [165]. A specialized subset of B cells marked by CD21hiCD1dhiCD5+ was identified as the main source of IL-35 in pancreatic cancer. The B cell-specific production of IL-35 promoted the expansion of Treg cells and blocked the anti-tumor activity of effector CD4+ T cells [169]. Furthermore, IL-35 directly acted on CD8+ T cells and suppressed their infiltration and effector function by downregulating the expression of IFN-γ and CXCR3 in a STAT3-dependent manner [169]. IL-35-producing B cells were also elevated in the peripheral blood of gastric cancer (GC) patients and were responsible for the accumulation of immune suppressive Treg cells, MDSCs, IL-10-producing B cells and CD14+ monocytes [170].
IL-35 signals through IL-12Rβ2 and gp130 receptors in T cells, which triggers Jak1 and Jak2 to activate STAT1 and STAT4 [157]. IL-35-mediated signaling via STAT1/STAT4 in conventional T cells can polarize them to IL-35-producing iT35 cells [171]. The expression of IL-35 by Foxp3+ Treg cells can reduce the proliferation of effector T cells and can lead to T cell exhaustion [171,172]. In this regard, Turnis at al. also demonstrated that IL-35+ Treg cells could enhance the surface expression of the negative regulators Tim-1, PD-1 and LAG3 on anti-tumor T cells [166]. A clinical study also showed that IL-35 produced by Treg cells promoted the growth of acute myeloid leukemia (AML) blasts in adult AML patients by limiting the activity of CD4+CD25− T effector cells and increasing cancer cell proliferation [175]. IL-35 was reported to limit anti-tumor immunity in NSCLC patients by suppressing Th1 and Th17 responses and cytotoxic genes in CD8+ T cells [176]. Apart from T cells, IL-35+ macrophages were able to upregulate IL-12Rβ2 expression on cancer cells and make them more responsive to IL-35, facilitating the activation of JAK2–STAT6 and GATA3, and increased metastasis in mouse models of breast and lung cancer [159]. Moreover, Zou et al. showed that IL-35 facilitated the polarization of neutrophils, leading to increases in tumor growth in mouse models of melanoma and hepatocellular carcinoma [177]. Furthermore, IL-35 promoted immune suppression by favoring M2 macrophage polarization [178,179]. IL-35 also impeded DC maturation by reducing the cell surface expression of MHCI and MHCII and co-stimulatory molecules CD80/CD86 [180,181,182]. Overall, these observations indicate that IL-35 has a strong immune suppressive effect in the TME and could play a major role in controlling anti-tumor immunity; therefore, targeting IL-35 in cancer may hold substantial therapeutic potential for the treatment of patients with cancer.
1.8. IL-35 in Cancer Immunotherapy
A detailed understanding of IL-35 in cancer is essential. This information may lead to the development of new immunotherapeutic agents for the treatment of cancer patients, and it can be used as a potential biomarker for disease progression. As an example, elevated levels of IL-35 have been associated with poor prognosis in many solid cancer types [183,184,185,186].
1.8.1. IL-35 in Inhibiting Tumor Growth
Although IL-35 plays a pro-tumorigenic role in several cancer models, such as melanoma, pancreatic cancer, NSCLCC, breast cancer, lymphoma and gastric cancer, among others, Zhang et al. reported that IL-35 inhibited the cell migration, proliferation, colony formation and invasion of human colon cancer cells by suppressing β-catenin [187]. In this instance, IL-35 suppressed cancer growth in vivo and sensitized colon cancer cells to chemotherapy. Thus, IL-35 in combination with chemotherapy could be a useful treatment for colon cancer. Similar studies showed that IL-35 plays an inhibitory role in human NSCLC and colon cancer cells by suppressing cell migration and colony formation [188,189]. In hepatocellular carcinoma (HCC) patients, IL-35 expression was significantly reduced in patients with advanced cancer as compared to early stages. The HCC patients with reduced IL-35 had increased tumor size, distant metastases and positive microvascular invasion. Furthermore, the overexpression of IL-35 upregulated MHCI and CD95 in HepG2 cancer cells and reduced cell migration, colony formation and invasion. These data indicate that IL-35 plays an anti-tumorigenic role in HCC by enhancing MHCI-specific anti-tumor immunity and by reducing cancer cell migration. [190].
1.8.2. IL-35 in Promoting Tumor Growth
However, most research studies have reported that increased levels of IL-35 can be correlated with the pathogenicity and progression of cancer. The levels of IL-35 in the serum samples of prostate cancer patients were highly elevated compared with healthy controls and correlated with disease progression [184,191]. Similar studies reported elevated levels of circulating IL-35 in breast cancer patients, which positively correlated with the expression of Ki67, p53 and EGFR on breast cancer cells [192]. Furthermore, high levels of IL-35 were shown to be associated with the poor survival of diffuse-large B-cell lymphoma (DLBCL) patients treated with chemotherapy [193]. Additionally, increased levels of IL-35 were observed in the bone marrow of acute myeloid leukemia (AML) patients and correlated positively with clinical stages [194]. IL-35 can also be secreted by cancer cells and was shown to have a pro-tumorigenic effect particularly in pancreatic cancer. To this end, IL-35 was associated with poor prognosis in pancreatic cancer patients and is also thought to promote the growth of pancreatic cancer cells by enabling metastasis and extravasation [195]. In this case, IL-35 signaling in cancer cells activated STAT1 via the gp130 receptor, triggered the expression of ICAM1, and increased endothelial adhesion and metastasis. The human pancreatic cancer cell production of IL-35 also favored the proliferation of tumor cells by promoting the expression of cyclin B, cyclin D, Cdk2 and Cdk4, and it inhibited apoptosis by increasing levels of Bcl2 and decreasing levels of TRAILR1 [196]. Jin et al. showed that elevated levels of IL-35 in pancreatic cancer patients correlated with increases in tumor size, TNM staging and lymph node metastasis [197]. Furthermore, IL-35 produced by human nasopharyngeal carcinoma, B cell lymphoma and melanoma tumor cells enhanced the infiltration of CD11b+Gr1+ myeloid cells and promoted angiogenesis [160]. These findings indicate that IL-35 may have a distinct role in cancer when produced by immune cells or cancer cells.
Accumulating evidence suggests that IL-35 blockade could synergize with immunotherapy to ameliorate tumor growth. Turnis et al. showed that anti-PD-1 therapy combined with the depletion of IL-35 and/or deletion of Treg cell-specific IL-35 expression significantly limited murine melanoma and colon cancer growth by promoting cytotoxic T cell proliferation, effector function and long-term memory formation [166]. Similarly, the loss of B cell-specific IL-35 expression or antibody-based IL-35 blockade synergized with anti-PD1 treatment in a CD8+ T cell-dependent manner to inhibit pancreatic cancer growth in preclinical models [169]. A similar approach of combining IL-35 blockade with the immune checkpoint inhibition of PD1 or PD-L1 was effective in NSCLC [185]. In this case, the neutralization of IL-35 hindered immunosuppressive macrophages and Treg cells while enhancing anti-tumor immunity. These findings identify the potential behind targeting IL-35-mediated immune suppression in the TME and highlight the importance of IL-35 as a therapeutic target in synergistic approaches with immune checkpoint therapy for the treatment of multiple cancer types.
1.9. IL-39
IL-39 is a newly proposed member of the IL-12 family. It is a heterodimeric cytokine consisting of the subunits p19 and EBi3 [198]. Most studies to date have only focused on cancer cell lines, and no concrete reports in animal models or human subjects are available yet. Thus, it is premature to designate IL-39 as an inflammatory or regulatory cytokine at this point in time. Wang et al. demonstrated that IL-39 could be secreted by LPS-stimulated B cells and could induce inflammation by activating STAT1/STAT3 in mouse models of lupus [198]. IL-39-producing B cells increased in mice with lupus, and the silencing of IL-39 led to reduced disease severity [199]. In this study, IL-39 induced the differentiation and expansion of CD11b+Gr1+ neutrophils in vivo. Furthermore, activated neutrophils could further potentiate the expression of B cell activation factor (BAFF) in GL7+ B cells, creating a positive feedback loop that results in the increased production of IL-39 [199]. IL-39 was also shown to have a pro-tumorigenic function in pancreatic cancer, where in the presence of IL-39, colonies of human pancreatic cancer cells significantly increased their proliferation rate [200]. This was accompanied by the reduced expression of p21 and TRAILR1. IL-39 is thought to signal via IL-23R and gp130 in target cells and to activate downstream STAT1 and STAT3 signaling [201,202]. Floss et al. engineered shuffled IL-12 family cytokine receptors, which are responsive to IL-39. The authors found that IL-39 may use two additional receptor combinations, IL-23R/IL-12Rβ2 and gp130/IL12Rβ1, in Ba/F3 cells [203]. These findings may highlight the flexibility of receptor usage by IL-39. More work needs to be done in order to understand the potential of targeting IL-39 in cancer immunotherapy.
2. Conclusions and Future Perspectives
IL-12 family cytokines play a critical role in the regulation of innate and adaptive immune responses. Their functions in the modulation of immune responses are well reported in autoimmunity and infectious diseases. These cytokines also play key roles in cancer initiation and progression. Tumor growth and spread have a direct relationship with host immune responses, and it is clear that IL-12 family cytokines can regulate tumor growth (Figure 1). Therefore, targeting or modifying the immune response against the tumor by harnessing the biological functionality of IL-12 family cytokines has recently gained lots of attention. The silent feature of various tumors is that they escape the host immune attack by favoring immune suppression within the TME. Cancer cells can secrete immune suppressive cytokines and chemokines and, in conjunction with regulatory immune cells, hinder the activity and proliferation of tumor-specific cytotoxic cells. Due to the “cold” nature of many cancers, therapies such as immune checkpoint blockade, adoptive T cell therapy, tumor vaccines, conventional chemotherapy and/or radiotherapy are frequently unable to manifest effective responses. Thus, strategies aiming to boost immune infiltration and functionality would be highly beneficial.
In this review, we summarized recent studies and advancements in the IL-12 field, namely, the roles and potential for the targeting of IL-12, IL-23, IL-27, IL-35 and IL-39 in tumorigenesis. The overarching objective is to make a tumor more susceptible to immune attack and become more responsive to conventional therapies. Targeting these cytokines may alter the tumor phenotype from immunologically “cold” to immunologically “hot”. As discussed above, these cytokines are secreted not only by immune cells but also by tumor cells. Therefore, therapies focusing on IL-12 family cytokines can block the tumor cell cycle, induce apoptosis and prevent tumor cell proliferation, together with facilitating effector immune responses against cancer cells. Synergistic therapies that focus on IL-12 family cytokines and immune checkpoint blockade, such as anti-PD1, neutralizing antibodies, adoptive T cell therapy and CAR T cell therapy have shown promising effects in preclinical models. Localized IL-12 delivery and synergistic therapy consisting of IL-12 with immune checkpoint inhibitors and adoptive cell transfer is under investigation in clinical trials [27]. Hu et al. demonstrated that the combination of IL-12 and doxorubicin could enhance the infiltration of cytotoxic T cells into large solid tumors in different human xenograft models [204]. The local expression of IL-12 was achieved by injecting IL-12 DNA and conducting in vivo electroporation. This treatment hampered Treg cell infiltration and increased the effector functions of tumor-infiltrated T cells [204,205]. This strategy is under investigation in clinical trials (NCT01579318, NCT00323206, NCT01502293 and NCT02345330) and also in combination with pembrolizumab (NCT02493361 and NCT03132675) [205]. Although IL-12 is an effector cytokine and recruits a variety of effector immune cells at the tumoral site, it is possible that it can induce inflammatory effects. Therefore, future studies, particularly studies involving clinical trials, should consider the inflammatory effects associated with IL-12 family cytokines while using them as immunotherapy for cancer patients.
Future studies should place more emphasis on developing appropriate targeting reagents suitable for use in human clinical trials. More preclinical studies need to be performed, especially on IL-39. Given the role of IL-12 family cytokines in modulating the immune cell as well as cancer cell phenotype and receptor expression pattern, downstream signaling pathways need to be studied in detail, as they may dictate downstream therapeutic strategies. Preclinical studies using genetic mouse models need to include head-to-head comparisons across the distinct subunits that comprise IL-12 cytokines in order to establish which particular IL-12 family cytokine(s) play a role in a specific cancer type. Although IL-12 family cytokines seem to be promising candidates for the treatment of different malignancies, their associated side-effects and roles in regulating the local versus systemic immune system still need to be elaborated in detail.
Funding
This work was supported in part by National Institutes of Health R37 CA230786 (Y.P.-G.), University Cancer Research Fund at the University of North Carolina at Chapel Hill, and a Concern Foundation Conquer Cancer Now Award (Y.P.-G.).
Acknowledgments
We thank W.J. Bell for help with editing the manuscript. Figures created using BioRender.com.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Wojno, E.D.T.; Hunter, C.A.; Stumhofer, J.S. The immunobiology of the interleukin-12 family: Room for discovery. Immunity 2019, 50, 851–870. [Google Scholar] [CrossRef] [PubMed]
- Mal, X.; Trinchieri, G. Regulation of interleukin-12 production in antigen-presenting cells. Dev. Funct. Myeloid Subsets 2001, 79, 55–92. [Google Scholar]
- Vignali, D.A.; Kuchroo, V.K. Il-12 family cytokines: Immunological playmakers. Nat. Immunol. 2012, 13, 722–728. [Google Scholar]
- Zundler, S.; Neurath, M.F. Interleukin-12: Functional activities and implications for disease. Cytokine Growth Factor Rev. 2015, 26, 559–568. [Google Scholar] [CrossRef]
- Tugues, S.; Burkhard, S.; Ohs, I.; Vrohlings, M.; Nussbaum, K.; Berg, J.V.; Kulig, P.; Becher, B. New insights into il-12-mediated tumor suppression. Cell Death Differ. 2015, 22, 237–246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thieu, V.T.; Yu, Q.; Chang, H.-C.; Yeh, N.; Nguyen, E.T.; Sehra, S.; Kaplan, M.H. Stat4 is required for t-bet to promote il-12-dependent th1 fate determination. Immunity 2008, 29, 679–690. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, C.L.; Schilling, M.M.; Cho, S.H.; Lee, K.; Wei, M.; Boothby, M. Stat4 and t-bet are required for the plasticity of ifn-γ expression across th2 ontogeny and influence changes in ifng promoter DNA methylation. J. Immunol. 2013, 191, 678–687. [Google Scholar] [CrossRef] [Green Version]
- Jenner, R.G.; Townsend, M.J.; Jackson, I.; Sun, K.; Bouwman, R.D.; Young, R.A.; Glimcher, L.H.; Lord, G.M. The transcription factors t-bet and gata-3 control alternative pathways of T-cell differentiation through a shared set of target genes. Proc. Natl. Acad. Sci. USA 2009, 106, 17876–17881. [Google Scholar]
- Vilgelm, A.E.; Richmond, A. Chemokines modulate immune surveillance in tumorigenesis, metastasis, and response to immunotherapy. Front. Immunol. 2019, 10, 333. [Google Scholar] [CrossRef] [Green Version]
- Allen, F.; Bobanga, I.D.; Rauhe, P.; Barkauskas, D.; Teich, N.; Tong, C.; Myers, J.; Huang, A.Y. Ccl3 augments tumor rejection and enhances cd8+ T cell infiltration through nk and cd103+ dendritic cell recruitment via ifnγ. Oncoimmunology 2018, 7, e1393598. [Google Scholar] [CrossRef] [Green Version]
- Allen, F.; Rauhe, P.; Askew, D.; Tong, A.A.; Nthale, J.; Eid, S.; Myers, J.T.; Tong, C.; Huang, A.Y. Ccl3 enhances antitumor immune priming in the lymph node via ifnγ with dependency on natural killer cells. Front. Immunol. 2017, 8, 1390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zwirner, N.W.; Ziblat, A. Regulation of nk cell activation and effector functions by the il-12 family of cytokines: The case of il-27. Front. Immunol. 2017, 8, 25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oka, N.; Markova, T.; Tsuzuki, K.; Li, W.; El-Darawish, Y.; Pencheva-Demireva, M.; Yamanishi, K.; Yamanishi, H.; Sakagami, M.; Tanaka, Y. Il-12 regulates the expansion, phenotype, and function of murine nk cells activated by il-15 and il-18. Cancer Immunol. Immunother. 2020, 69, 1699–1712. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Wu, B.; Ma, T.; Lin, Y.; Cheng, F.; Xiong, H.; Xie, C.; Liu, C.; Wang, Q.; Li, Z. Overexpression of il-12 reverses the phenotype and function of m2 macrophages to m1 macrophages. Int. J. Clin. Exp. Pathol. 2016, 9, 8963–8972. [Google Scholar]
- Suzuki, S.; Umezu, Y.; Saijo, Y.; Satoh, G.; Abe, Y.; Satoh, K.; Nukiwa, T. Exogenous recombinant human il-12 augments mhc class i antigen expression on human cancer cells in vitro. Tohoku J. Exp. Med. 1998, 185, 223–226. [Google Scholar] [CrossRef] [Green Version]
- Kerkar, S.P.; Goldszmid, R.S.; Muranski, P.; Chinnasamy, D.; Yu, Z.; Reger, R.N.; Leonardi, A.J.; Morgan, R.A.; Wang, E.; Marincola, F.M. Il-12 triggers a programmatic change in dysfunctional myeloid-derived cells within mouse tumors. J. Clin. Investig. 2011, 121, 4746–4757. [Google Scholar] [PubMed] [Green Version]
- Lazarevic, V.; Chen, X.; Shim, J.; Hwang, E.; Jang, E.; Bolm, A.; Oukka, M.; Kuchroo, V.; Glimcher, L. Transcription factor t-bet represses th17 differentiation by preventing runx1-mediated activation of the ror7t gene. Nat. Immunol. 2011, 12, 96–104. [Google Scholar] [CrossRef] [Green Version]
- Lin, Z.-W.; Wu, L.-X.; Xie, Y.; Ou, X.; Tian, P.-K.; Liu, X.-P.; Min, J.; Wang, J.; Chen, R.-F.; Chen, Y.-J. The expression levels of transcription factors t-bet, gata-3, rorγt and foxp3 in peripheral blood lymphocyte (pbl) of patients with liver cancer and their significance. Int. J. Med Sci. 2015, 12, 7–16. [Google Scholar] [CrossRef] [Green Version]
- Li, C.; Jiang, P.; Wei, S.; Xu, X.; Wang, J. Regulatory t cells in tumor microenvironment: New mechanisms, potential therapeutic strategies and future prospects. Mol. Cancer 2020, 19, 1–23. [Google Scholar]
- O’Malley, J.T.; Sehra, S.; Thieu, V.T.; Yu, Q.; Chang, H.C.; Stritesky, G.L.; Nguyen, E.T.; Mathur, A.N.; Levy, D.E.; Kaplan, M.H. Signal transducer and activator of transcription 4 limits the development of adaptive regulatory T cells. Immunology 2009, 127, 587–595. [Google Scholar]
- Lin, L.; Rayman, P.; Pavicic, P.G.; Tannenbaum, C.; Hamilton, T.; Montero, A.; Ko, J.; Gastman, B.; Finke, J.; Ernstoff, M. Ex vivo conditioning with il-12 protects tumor-infiltrating cd8+ T cells from negative regulation by local ifn-γ. Cancer Immunol. Immunother. 2019, 68, 395–405. [Google Scholar] [CrossRef] [PubMed]
- Garris, C.S.; Arlauckas, S.P.; Kohler, R.H.; Trefny, M.P.; Garren, S.; Piot, C.; Engblom, C.; Pfirschke, C.; Siwicki, M.; Gungabeesoon, J. Successful anti-pd-1 cancer immunotherapy requires T cell-dendritic cell crosstalk involving the cytokines ifn-γ and il-12. Immunity 2018, 49, 1148–1161.e7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, X.; Leonard, K.; Collins, L.I.; Cai, S.F.; Mayer, J.C.; Payton, J.E.; Walter, M.J.; Piwnica-Worms, D.; Schreiber, R.D.; Ley, T.J. Interleukin 12 stimulates ifn-γ–mediated inhibition of tumor-induced regulatory T-cell proliferation and enhances tumor clearance. Cancer Res. 2009, 69, 8700–8709. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, J.; Zhao, J.; Perlman, S. Differential effects of il-12 on tregs and non-treg T cells: Roles of ifn-γ, il-2 and il-2r. PLoS ONE 2012, 7, e46241. [Google Scholar] [CrossRef] [PubMed]
- Leonard, J.P.; Sherman, M.L.; Fisher, G.L.; Buchanan, L.J.; Larsen, G.; Atkins, M.B.; Sosman, J.A.; Dutcher, J.P.; Vogelzang, N.J.; Ryan, J.L. Effects of single-dose interleukin-12 exposure on interleukin-12–associated toxicity and interferon-γ production. Blood J. Am. Soc. Hematol. 1997, 90, 2541–2548. [Google Scholar]
- Wang, P.; Li, X.; Wang, J.; Gao, D.; Li, Y.; Li, H.; Chu, Y.; Zhang, Z.; Liu, H.; Jiang, G. Re-designing interleukin-12 to enhance its safety and potential as an anti-tumor immunotherapeutic agent. Nat. Commun. 2017, 8, 1–15. [Google Scholar]
- Nguyen, K.G.; Vrabel, M.R.; Mantooth, S.M.; Hopkins, J.J.; Wagner, E.S.; Gabaldon, T.A.; Zaharoff, D.A. Localized interleukin-12 for cancer immunotherapy. Front. Immunol. 2020, 11, 575597. [Google Scholar] [CrossRef]
- Li, J.; Lin, W.; Chen, H.; Xu, Z.; Ye, Y.; Chen, M. Dual-target il-12-containing nanoparticles enhance t cell functions for cancer immunotherapy. Cell. Immunol. 2020, 349, 104042. [Google Scholar] [CrossRef]
- Díaz-Montero, C.M.; el Naggar, S.; al Khami, A.; el Naggar, R.; Montero, A.J.; Cole, D.J.; Salem, M.L. Priming of naive cd8+ T cells in the presence of il-12 selectively enhances the survival of cd8+ cd62l hi cells and results in superior anti-tumor activity in a tolerogenic murine model. Cancer Immunol. Immunother. 2008, 57, 563–572. [Google Scholar] [CrossRef] [Green Version]
- Chinnasamy, D.; Yu, Z.; Kerkar, S.P.; Zhang, L.; Morgan, R.A.; Restifo, N.P.; Rosenberg, S.A. Local delivery of lnterleukin-12 using T cells targeting vegf receptor-2 eradicates multiple vascularized tumors in mice. Clin. Cancer Res. 2012, 18, 1672–1683. [Google Scholar] [CrossRef] [Green Version]
- Kerkar, S.P.; Muranski, P.; Kaiser, A.; Boni, A.; Sanchez-Perez, L.; Yu, Z.; Palmer, D.C.; Reger, R.N.; Borman, Z.A.; Zhang, L. Tumor-specific cd8+ t cells expressing interleukin-12 eradicate established cancers in lymphodepleted hosts. Cancer Res. 2010, 70, 6725–6734. [Google Scholar] [CrossRef] [Green Version]
- Chiocca, E.A.; John, S.Y.; Lukas, R.V.; Solomon, I.H.; Ligon, K.L.; Nakashima, H.; Triggs, D.A.; Reardon, D.A.; Wen, P.; Stopa, B.M. Regulatable interleukin-12 gene therapy in patients with recurrent high-grade glioma: Results of a phase 1 trial. Sci. Transl. Med. 2019, 11, eaaw5680. [Google Scholar] [CrossRef] [PubMed]
- Kueberuwa, G.; Kalaitsidou, M.; Cheadle, E.; Hawkins, R.E.; Gilham, D.E. Cd19 car T cells expressing il-12 eradicate lymphoma in fully lymphoreplete mice through induction of host immunity. Mol. Ther.-Oncolytics 2018, 8, 41–51. [Google Scholar] [CrossRef] [Green Version]
- Koneru, M.; Purdon, T.J.; Spriggs, D.; Koneru, S.; Brentjens, R.J. Il-12 secreting tumor-targeted chimeric antigen receptor T cells eradicate ovarian tumors in vivo. Oncoimmunology 2015, 4, e994446. [Google Scholar] [PubMed] [Green Version]
- Liu, Y.; Di, S.; Shi, B.; Zhang, H.; Wang, Y.; Wu, X.; Luo, H.; Wang, H.; Li, Z.; Jiang, H. Armored inducible expression of il-12 enhances antitumor activity of glypican-3–targeted chimeric antigen receptor–engineered T cells in hepatocellular carcinoma. J. Immunol. 2019, 203, 198–207. [Google Scholar] [CrossRef]
- Kunert, A.; Chmielewski, M.; Wijers, R.; Berrevoets, C.; Abken, H.; Debets, R. Intra-tumoral production of il18, but not il12, by tcr-engineered T cells is non-toxic and counteracts immune evasion of solid tumors. Oncoimmunology 2018, 7, e1378842. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brudno, J.N.; Kochenderfer, J.N. Toxicities of chimeric antigen receptor T cells: Recognition and management. Blood 2016, 127, 3321–3330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonifant, C.L.; Jackson, H.J.; Brentjens, R.J.; Curran, K.J. Toxicity and management in car T-cell therapy. Mol. Ther.-Oncolytics 2016, 3, 16011. [Google Scholar] [CrossRef] [PubMed]
- Hewitt, S.L.; Bailey, D.; Zielinski, J.; Apte, A.; Musenge, F.; Karp, R.; Burke, S.; Garcon, F.; Mishra, A.; Gurumurthy, S. Intratumoral interleukin-12 mrna therapy promotes th1 transformation of the tumor microenvironment. Clin. Cancer Res. 2020, 26, 6284–6298. [Google Scholar] [CrossRef]
- Fallon, J.K.; Vandeveer, A.J.; Schlom, J.; Greiner, J.W. Enhanced antitumor effects by combining an il-12/anti-DNA fusion protein with avelumab, an anti-pd-l1 antibody. Oncotarget 2017, 8, 20558–20571. [Google Scholar] [CrossRef] [Green Version]
- El-Shemi, A.G.; Ashshi, A.M.; Na, Y.; Li, Y.; Basalamah, M.; Al-Allaf, F.A.; Oh, E.; Jung, B.-K.; Chae-Ok, Y. Combined therapy with oncolytic adenoviruses encoding trail and il-12 genes markedly suppressed human hepatocellular carcinoma both in vitro and in an orthotopic transplanted mouse model. J. Exp. Clin. Cancer Res. 2016, 35, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lupi, L.A.; Delella, F.K.; Cucielo, M.S.; Romagnoli, G.G.; Kaneno, R.; Nunes, I.d.S.; Domeniconi, R.F.; Martinez, M.; Martinez, F.E.; Fávaro, W.J. P-mapa and interleukin-12 reduce cell migration/invasion and attenuate the toll-like receptor-mediated inflammatory response in ovarian cancer skov-3 cells: A preliminary study. Molecules 2020, 25, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bekaii-Saab, T.S.; Roda, J.M.; Guenterberg, K.D.; Ramaswamy, B.; Young, D.C.; Ferketich, A.K.; Lamb, T.A.; Grever, M.R.; Shapiro, C.L.; Carson, W.E. A phase i trial of paclitaxel and trastuzumab in combination with interleukin-12 in patients with her2/neu-expressing malignancies. Mol. Cancer Ther. 2009, 8, 2983–2991. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deplanque, G.; Shabafrouz, K.; Obeid, M. Can local radiotherapy and il-12 synergise to overcome the immunosuppressive tumor microenvironment and allow “in situ tumor vaccination”? Cancer Immunol. Immunother. 2017, 66, 833–840. [Google Scholar] [CrossRef]
- Eckert, F.; Jelas, I.; Oehme, M.; Huber, S.M.; Sonntag, K.; Welker, C.; Gillies, S.D.; Strittmatter, W.; Zips, D.; Handgretinger, R. Tumor-targeted il-12 combined with local irradiation leads to systemic tumor control via abscopal effects in vivo. Oncoimmunology 2017, 6, e1323161. [Google Scholar] [CrossRef] [Green Version]
- Guo, N.; Wang, W.-Q.; Gong, X.-J.; Gao, L.; Yang, L.-R.; Yu, W.-N.; Shen, H.-Y.; Wan, L.-Q.; Jia, X.-F.; Wang, Y.-S. Study of recombinant human interleukin-12 for treatment of complications after radiotherapy for tumor patients. World J. Clin. Oncol. 2017, 8, 158–167. [Google Scholar] [CrossRef]
- Mills, B.N.; Connolly, K.A.; Ye, J.; Murphy, J.D.; Uccello, T.P.; Han, B.J.; Zhao, T.; Drage, M.G.; Murthy, A.; Qiu, H. Stereotactic body radiation and interleukin-12 combination therapy eradicates pancreatic tumors by repolarizing the immune microenvironment. Cell Rep. 2019, 29, 406–421.e5. [Google Scholar] [CrossRef] [Green Version]
- Langowski, J.L.; Zhang, X.; Wu, L.; Mattson, J.D.; Chen, T.; Smith, K.; Basham, B.; McClanahan, T.; Kastelein, R.A.; Oft, M. Il-23 promotes tumour incidence and growth. Nature 2006, 442, 461–465. [Google Scholar] [CrossRef]
- Kastelein, R.A.; Hunter, C.A.; Cua, D.J. Discovery and biology of il-23 and il-27: Related but functionally distinct regulators of inflammation. Annu. Rev. Immunol. 2007, 25, 221–242. [Google Scholar] [CrossRef] [Green Version]
- Ignacio, A.; Breda, C.N.S.; Camara, N.O.S. Innate lymphoid cells in tissue homeostasis and diseases. World J. Hepatol. 2017, 9, 979–989. [Google Scholar] [CrossRef]
- Tian, Z.; van Velkinburgh, J.C.; Wu, Y.; Ni, B. Innate lymphoid cells involve in tumorigenesis. Int. J. Cancer 2016, 138, 22–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamassia, N.; Arruda-Silva, F.; Wright, H.L.; Moots, R.J.; Gardiman, E.; Bianchetto-Aguilera, F.; Gasperini, S.; Capone, M.; Maggi, L.; Annunziato, F. Human neutrophils activated via tlr8 promote th17 polarization through il-23. J. Leukoc. Biol. 2019, 105, 1155–1165. [Google Scholar] [CrossRef] [PubMed]
- Cauli, A.; Piga, M.; Floris, A.; Mathieu, A. Current Perspective on the Role of the Interleukin-23/Interleukin-17 Axis in Inflammation and Disease (Chronic Arthritis and Psoriasis). ImmunoTargets Ther. 2015, 4, 185–190. Available online: https://www.dovepress.com/front_end/cr_data/cache/pdf/download_1606621601_5fc319a1519b0/ITT-62870-current-perspectives-on-the-role-of-the-interleukin-23-inter_100115.pdf (accessed on 30 November 2020). [CrossRef] [PubMed] [Green Version]
- Wang, K.; Karin, M. The il-23 to il-17 cascade inflammation-related cancers. Clin. Exp. Rheumatol. 2015, 33, S87–S90. [Google Scholar]
- Qian, X.; Gu, L.; Ning, H.; Zhang, Y.; Hsueh, E.C.; Fu, M.; Hu, X.; Wei, L.; Hoft, D.F.; Liu, J. Increased th17 cells in the tumor microenvironment is mediated by il-23 via tumor-secreted prostaglandin e2. J. Immunol. 2013, 190, 5894–5902. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Li, D.; Shi, Q.; Huang, X.; Ju, X. Umbilical cord blood-derived helios-positive regulatory t cells promote angiogenesis in acute lymphoblastic leukemia in mice via ccl22 and the vegfa-vegfr2 pathway. Mol. Med. Rep. 2019, 19, 4195–4204. [Google Scholar] [CrossRef] [Green Version]
- Martinenaite, E.; Ahmad, S.M.; Hansen, M.; Met, Ö.; Westergaard, M.W.; Larsen, S.K.; Klausen, T.W.; Donia, M.; Svane, I.M.; Andersen, M.H. Ccl22-specific T cells: Modulating the immunosuppressive tumor microenvironment. Oncoimmunology 2016, 5, e1238541. [Google Scholar] [CrossRef] [Green Version]
- Wiedemann, G.M.; Röhrle, N.; Makeschin, M.-C.; Fesseler, J.; Endres, S.; Mayr, D.; Anz, D. Peritumoural ccl1 and ccl22 expressing cells in hepatocellular carcinomas shape the tumour immune infiltrate. Pathology 2019, 51, 586–592. [Google Scholar] [CrossRef]
- Zhang, J.; Ji, C.; Li, W.; Mao, Z.; Shi, Y.; Shi, H.; Ji, R.; Qian, H.; Xu, W.; Zhang, X. Tumor-educated neutrophils activate mesenchymal stem cells to promote gastric cancer growth and metastasis. Front. Cell Dev. Biol. 2020, 8, 788. [Google Scholar] [CrossRef]
- Mao, Z.; Zhang, J.; Shi, Y.; Li, W.; Shi, H.; Ji, R.; Mao, F.; Qian, H.; Xu, W.; Zhang, X. Cxcl5 promotes gastric cancer metastasis by inducing epithelial-mesenchymal transition and activating neutrophils. Oncogenesis 2020, 9, 1–14. [Google Scholar]
- Grivennikov, S.I.; Wang, K.; Mucida, D.; Stewart, C.A.; Schnabl, B.; Jauch, D.; Taniguchi, K.; Yu, G.-Y.; Österreicher, C.H.; Hung, K.E. Adenoma-linked barrier defects and microbial products drive il-23/il-17-mediated tumour growth. Nature 2012, 491, 254–258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, C.; Lagoudas, G.K.; Zhao, C.; Bullman, S.; Bhutkar, A.; Hu, B.; Ameh, S.; Sandel, D.; Liang, X.S.; Mazzilli, S. Commensal microbiota promote lung cancer development via γδ T cells. Cell 2019, 176, 998–1013.e16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaffen, S.L.; Jain, R.; Garg, A.V.; Cua, D.J. The il-23–il-17 immune axis: From mechanisms to therapeutic testing. Nat. Rev. Immunol. 2014, 14, 585–600. [Google Scholar] [CrossRef] [PubMed]
- Iwakura, Y.; Ishigame, H. The il-23/il-17 axis in inflammation. J. Clin. Investig. 2006, 116, 1218–1222. [Google Scholar] [CrossRef] [Green Version]
- Ma, S.; Cheng, Q.; Cai, Y.; Gong, H.; Wu, Y.; Yu, X.; Shi, L.; Wu, D.; Dong, C.; Liu, H. Il-17a produced by γδ T cells promotes tumor growth in hepatocellular carcinoma. Cancer Res. 2014, 74, 1969–1982. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.S.; Tato, C.M.; Joyce-Shaikh, B.; Gulen, M.F.; Cayatte, C.; Chen, Y.; Blumenschein, W.M.; Judo, M.; Ayanoglu, G.; McClanahan, T.K. Interleukin-23-independent il-17 production regulates intestinal epithelial permeability. Immunity 2015, 43, 727–738. [Google Scholar] [CrossRef] [Green Version]
- Kimura, Y.; Nagai, N.; Tsunekawa, N.; Sato-Matsushita, M.; Yoshimoto, T.; Cua, D.J.; Iwakura, Y.; Yagita, H.; Okada, F.; Tahara, H. Il-17a-producing cd 30+ vδ1 T cells drive inflammation-induced cancer progression. Cancer Sci. 2016, 107, 1206–1214. [Google Scholar] [CrossRef]
- Pastor-Fernández, G.; Mariblanca, I.R.; Navarro, M.N. Decoding il-23 signaling cascade for new therapeutic opportunities. Cells 2020, 9, 2044. [Google Scholar] [CrossRef]
- Tang, L.; Wang, K. Chronic inflammation in skin malignancies. J. Mol. Signal. 2016, 11, 2. [Google Scholar]
- Wang, K.; Kim, M.K.; di Caro, G.; Wong, J.; Shalapour, S.; Wan, J.; Zhang, W.; Zhong, Z.; Sanchez-Lopez, E.; Wu, L.-W. Interleukin-17 receptor a signaling in transformed enterocytes promotes early colorectal tumorigenesis. Immunity 2014, 41, 1052–1063. [Google Scholar] [CrossRef] [Green Version]
- Kirshberg, S.; Izhar, U.; Amir, G.; Demma, J.; Vernea, F.; Beider, K.; Shlomai, Z.; Wald, H.; Zamir, G.; Shapira, O.M. Involvement of ccr6/ccl20/il-17 axis in nsclc disease progression. PLoS ONE 2011, 6, e24856. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Yang, T.; Liu, X.; Guo, J.n.; Xie, T.; Ding, Y.; Lin, M.; Yang, H. Il-17 promotes tumor angiogenesis through stat3 pathway mediated upregulation of vegf in gastric cancer. Tumor Biol. 2016, 37, 5493–5501. [Google Scholar] [CrossRef] [PubMed]
- Pan, B.; Shen, J.; Cao, J.; Zhou, Y.; Shang, L.; Jin, S.; Cao, S.; Che, D.; Liu, F.; Yu, Y. Interleukin-17 promotes angiogenesis by stimulating vegf production of cancer cells via the stat3/giv signaling pathway in non-small-cell lung cancer. Sci. Rep. 2015, 5, 16053. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Yi, T.; Kortylewski, M.; Pardoll, D.M.; Zeng, D.; Yu, H. Il-17 can promote tumor growth through an il-6–stat3 signaling pathway. J. Exp. Med. 2009, 206, 1457–1464. [Google Scholar] [PubMed] [Green Version]
- Liu, J.; Wang, L.; Wang, T.; Wang, J. Expression of il-23r and il-17 and the pathology and prognosis of urinary bladder carcinoma. Oncol. Lett. 2018, 16, 4325–4330. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.; Smyth, M.J.; Teng, M.W. Interleukin (il)-12 and il-23 and their conflicting roles in cancer. Cold Spring Harb. Perspect. Biol. 2018, 10, a028530. [Google Scholar] [CrossRef]
- Li, J.; Zhang, L.; Zhang, J.; Wei, Y.; Li, K.; Huang, L.; Zhang, S.; Gao, B.; Wang, X.; Lin, P. Interleukin 23 regulates proliferation of lung cancer cells in a concentration-dependent way in association with the interleukin-23 receptor. Carcinogenesis 2013, 34, 658–666. [Google Scholar]
- Tao, Y.; Tao, T.; Gross, N.; Peng, X.; Li, Y.; Huang, Z.; Liu, L.; Li, G.; Chen, X.; Yang, J. Combined effect of il-12rβ2 and il-23r expression on prognosis of patients with laryngeal cancer. Cell. Physiol. Biochem. 2018, 50, 1041–1054. [Google Scholar] [CrossRef]
- Cocco, C.; Canale, S.; Frasson, C.; di Carlo, E.; Ognio, E.; Ribatti, D.; Prigione, I.; Basso, G.; Airoldi, I. Interleukin-23 acts as antitumor agent on childhood b-acute lymphoblastic leukemia cells. Blood J. Am. Soc. Hematol. 2010, 116, 3887–3898. [Google Scholar] [CrossRef] [Green Version]
- Chen, F.; Cao, A.; Yao, S.; Evans-Marin, H.L.; Liu, H.; Wu, W.; Carlsen, E.D.; Dann, S.M.; Soong, L.; Sun, J. Mtor mediates il-23 induction of neutrophil il-17 and il-22 production. J. Immunol. 2016, 196, 4390–4399. [Google Scholar]
- Lee, P.W.; Smith, A.J.; Yang, Y.; Selhorst, A.J.; Liu, Y.; Racke, M.K.; Lovett-Racke, A.E. Il-23r–activated stat3/stat4 is essential for th1/th17-mediated cns autoimmunity. JCI Insight 2017, 2, e91663. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, R.; Hedl, M.; Abraham, C. Il23 group> il23r recycling and amplifies innate receptor-induced signalling and cytokines in human macrophages, and the ibd-protective il23r r381q variant modulates these outcomes. Gut 2020, 69, 264–273. [Google Scholar] [CrossRef] [PubMed]
- McCuaig, S.; Barras, D.; Mann, E.; Friedrich, M.; Bullers, S.J.; Janney, A.; Garner, L.C.; Domingo, E.; Koelzer, V.H.; Delorenzi, M. The interleukin 22 pathway interacts with mutant kras to promote poor prognosis in colon cancer. Clin. Cancer Res. 2020, 26, 4313–4325. [Google Scholar] [CrossRef] [PubMed]
- Aden, K.; Rehman, A.; Falk-Paulsen, M.; Secher, T.; Kuiper, J.; Tran, F.; Pfeuffer, S.; Sheibani-Tezerji, R.; Breuer, A.; Luzius, A. Epithelial il-23r signaling licenses protective il-22 responses in intestinal inflammation. Cell Rep. 2016, 16, 2208–2218. [Google Scholar] [CrossRef] [Green Version]
- Markota, A.; Endres, S.; Kobold, S. Targeting interleukin-22 for cancer therapy. Hum. Vaccines Immunother. 2018, 14, 2012–2015. [Google Scholar]
- Guéry, L.; Hugues, S. Th17 cell plasticity and functions in cancer immunity. BioMed Res. Int. 2015, 2015, 1–11. [Google Scholar]
- Bailey, S.R.; Nelson, M.H.; Himes, R.A.; Li, Z.; Mehrotra, S.; Paulos, C.M. Th17 cells in cancer: The ultimate identity crisis. Front. Immunol. 2014, 5, 276. [Google Scholar]
- Li, J.; Lau, G.; Chen, L.; Yuan, Y.-F.; Huang, J.; Luk, J.M.; Xie, D.; Guan, X.-Y. Interleukin 23 promotes hepatocellular carcinoma metastasis via nf-kappa b induced matrix metalloproteinase 9 expression. PLoS ONE 2012, 7, e46264. [Google Scholar] [CrossRef] [Green Version]
- Chang, H.-H.; Young, S.H.; Sinnett-Smith, J.; Chou, C.E.N.; Moro, A.; Hertzer, K.M.; Hines, O.J.; Rozengurt, E.; Eibl, G. Prostaglandin e2 activates the mtorc1 pathway through an ep4/camp/pka-and ep1/ca2+-mediated mechanism in the human pancreatic carcinoma cell line panc-1. Am. J. Physiol.-Cell Physiol. 2015, 309, C639–C649. [Google Scholar] [CrossRef] [Green Version]
- Sheng, S.; Zhang, J.; Ai, J.; Hao, X.; Luan, R. Aberrant expression of il-23/il-23r in patients with breast cancer and its clinical significance. Mol. Med. Rep. 2018, 17, 4639–4644. [Google Scholar]
- Elessawi, D.F.; Alkady, M.M.; Ibrahim, I.M. Diagnostic and prognostic value of serum il-23 in colorectal cancer. Arab J. Gastroenterol. 2019, 20, 65–68. [Google Scholar] [CrossRef] [PubMed]
- Stanilov, N.; Miteva, L.; Jovchev, J.; Cirovski, G.; Stanilova, S. The prognostic value of preoperative serum levels of il-12p40 and il-23 for survival of patients with colorectal cancer. Apmis 2014, 122, 1223–1229. [Google Scholar] [CrossRef] [PubMed]
- Neurath, M.F. Il-23 in inflammatory bowel diseases and colon cancer. Cytokine Growth Factor Rev. 2019, 45, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Hmid, A.B.; Selmi, O.; Rekik, R.; Lamari, H.; Zamali, I.; Ladeb, S.; Safra, I.; Othman, T.B.; Romdhane, N.B.; Ahmed, M.B. Rorc overexpression as a sign of th17 lymphocytes accumulation in multiple myeloma bone marrow. Cytokine 2020, 134, 155210. [Google Scholar] [CrossRef]
- Koh, J.; Kim, H.Y.; Lee, Y.; Park, I.K.; Kang, C.H.; Kim, Y.T.; Kim, J.-E.; Choi, M.; Lee, W.-W.; Jeon, Y.K. Il23-producing human lung cancer cells promote tumor growth via conversion of innate lymphoid cell 1 (ilc1) into ilc3. Clin. Cancer Res. 2019, 25, 4026–4037. [Google Scholar] [CrossRef] [Green Version]
- Khodadadi, A.; Razmkhah, M.; Eskandari, A.-R.; Hosseini, A.; Habibagahi, M.; Ghaderi, A.; Jaberipour, M. Il-23/il-27 ratio in peripheral blood of patients with breast cancer. Iran. J. Med. Sci. 2014, 39, 350–356. [Google Scholar]
- Liu, Y.; Song, Y.; Lin, D.; Lei, L.; Mei, Y.; Jin, Z.; Gong, H.; Zhu, Y.; Hu, B.; Zhang, Y. Ncr−group 3 innate lymphoid cells orchestrate il-23/il-17 axis to promote hepatocellular carcinoma development. EBioMedicine 2019, 41, 333–344. [Google Scholar] [CrossRef] [Green Version]
- Calcinotto, A.; Spataro, C.; Zagato, E.; di Mitri, D.; Gil, V.; Crespo, M.; de Bernardis, G.; Losa, M.; Mirenda, M.; Pasquini, E. Il-23 secreted by myeloid cells drives castration-resistant prostate cancer. Nature 2018, 559, 363–369. [Google Scholar]
- Fu, Q.; Xu, L.; Wang, Y.; Jiang, Q.; Liu, Z.; Zhang, J.; Zhou, Q.; Zeng, H.; Tong, S.; Wang, T. Tumor-associated macrophage-derived interleukin-23 interlinks kidney cancer glutamine addiction with immune evasion. Eur. Urol. 2019, 75, 752–763. [Google Scholar]
- Błogowski, W.; Deskur, A.; Budkowska, M.; Sałata, D.; Madej-Michniewicz, A.; Dąbkowski, K.; Dołęgowska, B.; Starzyńska, T. Selected cytokines in patients with pancreatic cancer: A preliminary report. PLoS ONE 2014, 9, e97613. [Google Scholar] [CrossRef] [Green Version]
- Hussain, S.M.; Reed, L.F.; Krasnick, B.A.; Miranda-Carboni, G.; Fields, R.C.; Bi, Y.; Elahi, A.; Ajidahun, A.; Dickson, P.V.; Deneve, J.L. Il23 and tgf-ss diminish macrophage associated metastasis in pancreatic carcinoma. Sci. Rep. 2018, 8, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caughron, B.; Yang, Y.; Young, M.R.I. Role of il-23 signaling in the progression of premalignant oral lesions to cancer. PLoS ONE 2018, 13, e0196034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baird, A.-M.; Leonard, J.; Naicker, K.M.; Kilmartin, L.; O’Byrne, K.J.; Gray, S.G. Il-23 is pro-proliferative, epigenetically regulated and modulated by chemotherapy in non-small cell lung cancer. Lung Cancer 2013, 79, 83–90. [Google Scholar] [CrossRef] [PubMed]
- Fukuda, M.; Ehara, M.; Suzuki, S.; Ohmori, Y.; Sakashita, H. Il-23 promotes growth and proliferation in human squamous cell carcinoma of the oral cavity. Int. J. Oncol. 2010, 36, 1355–1365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nie, W.; Yu, T.; Sang, Y.; Gao, X. Tumor-promoting effect of il-23 in mammary cancer mediated by infiltration of m2 macrophages and neutrophils in tumor microenvironment. Biochem. Biophys. Res. Commun. 2017, 482, 1400–1406. [Google Scholar] [CrossRef] [PubMed]
- Kortlever, R.M.; Sodir, N.M.; Wilson, C.H.; Burkhart, D.L.; Pellegrinet, L.; Swigart, L.B.; Littlewood, T.D.; Evan, G.I. Myc cooperates with ras by programming inflammation and immune suppression. Cell 2017, 171, 1301–1315.e14. [Google Scholar] [CrossRef]
- Zhang, Q.; Liu, S.; Zhang, Q.; Xiong, Z.; Wang, A.R.; Myers, L.; Melamed, J.; Tang, W.W.; You, Z. Interleukin-17 promotes development of castration-resistant prostate cancer potentially through creating an immunotolerant and pro-angiogenic tumor microenvironment. Prostate 2014, 74, 869–879. [Google Scholar] [CrossRef] [Green Version]
- Wang, D.; Shao, Y.; Zhang, X.; Lu, G.; Liu, B. Il-23 and psma-targeted duo-car t cells in prostate cancer eradication in a preclinical model. J. Transl. Med. 2020, 18, 1–10. [Google Scholar] [CrossRef]
- Testa, U.; Castelli, G.; Pelosi, E. Cellular and molecular mechanisms underlying prostate cancer development: Therapeutic implications. Medicines 2019, 6, 82. [Google Scholar] [CrossRef] [Green Version]
- Ma, X.; Shou, P.; Smith, C.; Chen, Y.; Du, H.; Sun, C.; Kren, N.P.; Michaud, D.; Ahn, S.; Vincent, B. Interleukin-23 engineering improves car t cell function in solid tumors. Nat. Biotechnol. 2020, 38, 448–459. [Google Scholar] [CrossRef]
- Fabbi, M.; Carbotti, G.; Ferrini, S. Dual roles of il-27 in cancer biology and immunotherapy. Mediat. Inflamm. 2017, 2017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pot, C.; Apetoh, L.; Awasthi, A.; Kuchroo, V.K. Induction of regulatory tr1 cells and inhibition of th17 cells by il-27. Semin. Immunol. 2011, 23, 438–445. [Google Scholar] [CrossRef] [Green Version]
- Pot, C.; Apetoh, L.; Awasthi, A.; Kuchroo, V.K. Molecular pathways in the induction of interleukin-27-driven regulatory type 1 cells. J. Interf. Cytokine Res. 2010, 30, 381–388. [Google Scholar] [CrossRef] [PubMed]
- Murugaiyan, G.; Saha, B. Il-27 in tumor immunity and immunotherapy. Trends Mol. Med. 2013, 19, 108–116. [Google Scholar] [CrossRef] [PubMed]
- Murugaiyan, G.; Mittal, A.; Lopez-Diego, R.; Maier, L.M.; Anderson, D.E.; Weiner, H.L. Il-27 is a key regulator of il-10 and il-17 production by human cd4+ T cells. J. Immunol. 2009, 183, 2435–2443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qi, J.; Zhang, Z.; Tang, X.; Li, W.; Chen, W.; Yao, G. Il-27 regulated cd4+ il-10+ T cells in experimental sjögren syndrome. Front. Immunol. 2020, 11, 1699. [Google Scholar] [CrossRef]
- Pot, C.; Jin, H.; Awasthi, A.; Liu, S.M.; Lai, C.-Y.; Madan, R.; Sharpe, A.H.; Karp, C.L.; Miaw, S.-C.; Ho, I.-C. Cutting edge: Il-27 induces the transcription factor c-maf, cytokine il-21, and the costimulatory receptor icos that coordinately act together to promote differentiation of il-10-producing tr1 cells. J. Immunol. 2009, 183, 797–801. [Google Scholar] [CrossRef] [Green Version]
- Chang, K.-K.; Liu, L.-B.; Jin, L.-P.; Zhang, B.; Mei, J.; Li, H.; Wei, C.-Y.; Zhou, W.-J.; Zhu, X.-Y.; Shao, J. Il-27 triggers il-10 production in th17 cells via a c-maf/ror γ t/blimp-1 signal to promote the progression of endometriosis. Cell Death Dis. 2017, 8, e2666. [Google Scholar] [CrossRef] [Green Version]
- Guzzo, C.; Mat, N.F.C.; Gee, K. Interleukin-27 induces a stat1/3-and nf-κb-dependent proinflammatory cytokine profile in human monocytes. J. Biol. Chem. 2010, 285, 24404–24411. [Google Scholar] [CrossRef] [Green Version]
- Wang, N.; Liang, H.; Zen, K. Molecular mechanisms that influence the macrophage m1–m2 polarization balance. Front. Immunol. 2014, 5, 614. [Google Scholar] [CrossRef] [Green Version]
- Lu, Y.; Xu, W.; Gu, Y.; Chang, X.; Wei, G.; Rong, Z.; Qin, L.; Chen, X.; Zhou, F. Non-small cell lung cancer cells modulate the development of human cd1c+ conventional dendritic cell subsets mediated by cd103 and cd205. Front. Immunol. 2019, 10, 2829. [Google Scholar] [CrossRef] [PubMed]
- Iwasaki, Y.; Fujio, K.; Okamura, T.; Yamamoto, K. Interleukin-27 in t cell immunity. Int. J. Mol. Sci. 2015, 16, 2851–2863. [Google Scholar] [CrossRef] [PubMed]
- Owaki, T.; Asakawa, M.; Morishima, N.; Hata, K.; Fukai, F.; Matsui, M.; Mizuguchi, J.; Yoshimoto, T. A role for il-27 in early regulation of th1 differentiation. J. Immunol. 2005, 175, 2191–2200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamiya, S.; Owaki, T.; Morishima, N.; Fukai, F.; Mizuguchi, J.; Yoshimoto, T. An indispensable role for stat1 in il-27-induced t-bet expression but not proliferation of naive cd4+ T cells. J. Immunol. 2004, 173, 3871–3877. [Google Scholar] [CrossRef] [Green Version]
- Feng, X.M.; Liu, N.; Yang, S.G.; Hu, L.Y.; Chen, X.L.; Fang, Z.H.; Ren, Q.; Lu, S.H.; Liu, B.; Han, Z.C. Regulation of the class ii and class i mhc pathways in human thp-1 monocytic cells by interleukin-27. Biochem. Biophys. Res. Commun. 2008, 367, 553–559. [Google Scholar] [CrossRef]
- Torrado, E.; Fountain, J.; Pearl, J.; Cooper, A. Il-27 sustains t-bet expression and promotes the development of terminally differentiated cd4 T cells during tuberculosis (59.5). Am. Assoc. Immnol. 2012, 188 (Suppl. 1), 59.5. [Google Scholar]
- Hirahara, K.; Onodera, A.; Villarino, A.V.; Bonelli, M.; Sciumè, G.; Laurence, A.; Sun, H.-W.; Brooks, S.R.; Vahedi, G.; Shih, H.-Y. Asymmetric action of stat transcription factors drives transcriptional outputs and cytokine specificity. Immunity 2015, 42, 877–889. [Google Scholar] [CrossRef] [Green Version]
- Schneider, R.; Yaneva, T.; Beauseigle, D.; El-Khoury, L.; Arbour, N. Il-27 increases the proliferation and effector functions of human naive cd8+ t lymphocytes and promotes their development into tc1 cells. Eur. J. Immunol. 2011, 41, 47–59. [Google Scholar] [CrossRef]
- Morishima, N.; Mizoguchi, I.; Okumura, M.; Chiba, Y.; Xu, M.; Shimizu, M.; Matsui, M.; Mizuguchi, J.; Yoshimoto, T. A pivotal role for interleukin-27 in cd8+ T cell functions and generation of cytotoxic t lymphocytes. J. Biomed. Biotechnol. 2010, 2010, 1–10. [Google Scholar] [CrossRef]
- Gotthardt, D.; Trifinopoulos, J.; Sexl, V.; Putz, E.M. Jak/stat cytokine signaling at the crossroad of nk cell development and maturation. Front. Immunol. 2019, 10, 2590. [Google Scholar] [CrossRef] [Green Version]
- Choi, Y.H.; Lim, E.J.; Kim, S.W.; Moon, Y.W.; Park, K.S.; An, H.-J. Il-27 enhances il-15/il-18-mediated activation of human natural killer cells. J. Immunother. Cancer 2019, 7, 168. [Google Scholar] [CrossRef] [PubMed]
- Hisada, M.; Kamiya, S.; Fujita, K.; Belladonna, M.L.; Aoki, T.; Koyanagi, Y.; Mizuguchi, J.; Yoshimoto, T. Potent antitumor activity of interleukin-27. Cancer Res. 2004, 64, 1152–1156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Villarino, A.V.; Stumhofer, J.S.; Saris, C.J.; Kastelein, R.A.; de Sauvage, F.J.; Hunter, C.A. Il-27 limits il-2 production during th1 differentiation. J. Immunol. 2006, 176, 237–247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Owaki, T.; Asakawa, M.; Kamiya, S.; Takeda, K.; Fukai, F.; Mizuguchi, J.; Yoshimoto, T. Il-27 suppresses cd28-medicated il-2 production through suppressor of cytokine signaling 3. J. Immunol. 2006, 176, 2773–2780. [Google Scholar] [CrossRef] [Green Version]
- Zhou, W.-J.; Chang, K.-K.; Wu, K.; Yang, H.-L.; Mei, J.; Xie, F.; Li, D.-J.; Li, M.-Q. Rapamycin synergizes with cisplatin in antiendometrial cancer activation by improving il-27–stimulated cytotoxicity of nk cells. Neoplasia 2018, 20, 69–79. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Zhou, B.; Zhang, K.; Song, Y.; Zhang, L.; Xi, M. Il-27 suppresses skov3 cells proliferation by enhancing stat3 and inhibiting the akt signal pathway. Mol. Immunol. 2016, 78, 155–163. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Sato, A.; Shimozato, O.; Shingyoji, M.; Tada, Y.; Tatsumi, K.; Shimada, H.; Hiroshima, K.; Tagawa, M. Administration of DNA encoding the interleukin-27 gene augments antitumour responses through non-adaptive immunity. Scand. J. Immunol. 2015, 82, 320–327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiba, Y.; Mizoguchi, I.; Furusawa, J.; Hasegawa, H.; Ohashi, M.; Xu, M.; Owaki, T.; Yoshimoto, T. Interleukin-27 exerts its antitumor effects by promoting differentiation of hematopoietic stem cells to m1 macrophages. Cancer Res. 2018, 78, 182–194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yao, L.; Wang, M.; Niu, Z.; Liu, Q.; Gao, X.; Zhou, L.; Liao, Q.; Zhao, Y. Interleukin-27 inhibits malignant behaviors of pancreatic cancer cells by targeting m2 polarized tumor associated macrophages. Cytokine 2017, 89, 194–200. [Google Scholar] [CrossRef] [PubMed]
- Carbotti, G.; Nikpoor, A.R.; Vacca, P.; Gangemi, R.; Giordano, C.; Campelli, F.; Ferrini, S.; Fabbi, M. Il-27 mediates hla class i up-regulation, which can be inhibited by the il-6 pathway, in hla-deficient small cell lung cancer cells. J. Exp. Clin. Cancer Res. 2017, 36, 140. [Google Scholar] [CrossRef] [Green Version]
- Airoldi, I.; Tupone, M.G.; Esposito, S.; Russo, M.V.; Barbarito, G.; Cipollone, G.; di Carlo, E. Interleukin-27 re-educates intratumoral myeloid cells and down-regulates stemness genes in non-small cell lung cancer. Oncotarget 2015, 6, 3694–3708. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kourko, O.; Smyth, R.; Cino, D.; Seaver, K.; Petes, C.; Eo, S.Y.; Basta, S.; Gee, K. Poly (i: C)-mediated death of human prostate cancer cell lines is induced by interleukin-27 treatment. J. Interf. Cytokine Res. 2019, 39, 483–494. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.-Q.; Zhao, W.-H.; Zhao, X.-X.; Zhang, J.; Nan, K.-J. Association between il-27 2905t/g genotypes and the risk and survival of cervical cancer: A case-control study. Biomarkers 2016, 21, 272–275. [Google Scholar] [CrossRef] [PubMed]
- Majumder, D.; Debnath, R.; Maiti, D. Il-27 along with il-28b ameliorates the pulmonary redox impairment, inflammation and immunosuppression in benzo (a) pyrene induced lung cancer bearing mice. Life Sci. 2020, 260, 118384. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Liu, J.-Q.; Shi, M.; Cheng, X.; Ding, M.; Zhang, J.C.; Davis, J.P.; Varikuti, S.; Satoskar, A.R.; Lu, L. Il-27 gene therapy induces depletion of tregs and enhances the efficacy of cancer immunotherapy. JCI Insight 2018, 3. [Google Scholar] [CrossRef] [Green Version]
- Hu, A.; Ding, M.; Zhu, J.; Liu, J.-Q.; Pan, X.; Ghoshal, K.; Bai, X.-F. Intra-tumoral delivery of il-27 using adeno-associated virus stimulates anti-tumor immunity and enhances the efficacy of immunotherapy. Front. Cell Dev. Biol. 2020, 8, 210. [Google Scholar] [CrossRef]
- Yan, H.; Viswanadhapalli, S.; Chupp, D.; Fernandez, M.; Wu, S.; Wang, J.; Moroney, J.; Taylor, J.; Im, J.; Rivera, C. B cell-produced il-27 up-regulates pd-l1 expression in the tumor microenvironment to promote breast cancer development. AACR 2019. [Google Scholar] [CrossRef]
- Chen, S.; Crabill, G.A.; Pritchard, T.S.; McMiller, T.L.; Wei, P.; Pardoll, D.M.; Pan, F.; Topalian, S.L. Mechanisms regulating pd-l1 expression on tumor and immune cells. J. Immunother. Cancer 2019, 7, 1–12. [Google Scholar] [CrossRef]
- Carbotti, G.; Barisione, G.; Airoldi, I.; Mezzanzanica, D.; Bagnoli, M.; Ferrero, S.; Petretto, A.; Fabbi, M.; Ferrini, S. Il-27 induces the expression of ido and pd-l1 in human cancer cells. Oncotarget 2015, 6, 43267–43280. [Google Scholar] [CrossRef] [Green Version]
- Rolvering, C.; Zimmer, A.D.; Ginolhac, A.; Margue, C.; Kirchmeyer, M.; Servais, F.; Hermanns, H.M.; Hergovits, S.; Nazarov, P.V.; Nicot, N. The pd-l1-and il6-mediated dampening of the il27/stat1 anticancer responses are prevented by α-pd-l1 or α-il6 antibodies. J. Leukoc. Biol. 2018, 104, 969–985. [Google Scholar] [CrossRef]
- Horlad, H.; Ma, C.; Yano, H.; Pan, C.; Ohnishi, K.; Fujiwara, Y.; Endo, S.; Kikukawa, Y.; Okuno, Y.; Matsuoka, M. An il-27/stat3 axis induces expression of programmed cell death 1 ligands (pd-l1/2) on infiltrating macrophages in lymphoma. Cancer Sci. 2016, 107, 1696–1704. [Google Scholar] [CrossRef] [PubMed]
- Meissl, K.; Macho-Maschler, S.; Müller, M.; Strobl, B. The good and the bad faces of stat1 in solid tumours. Cytokine 2017, 89, 12–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, C.; Sakuishi, K.; Xiao, S.; Sun, Z.; Zaghouani, S.; Gu, G.; Wang, C.; Tan, D.J.; Wu, C.; Rangachari, M. An il-27/nfil3 signalling axis drives tim-3 and il-10 expression and t-cell dysfunction. Nat. Commun. 2015, 6, 1–12. [Google Scholar]
- Anderson, A.C.; Joller, N.; Kuchroo, V.K. Lag-3, tim-3, and tigit: Co-inhibitory receptors with specialized functions in immune regulation. Immunity 2016, 44, 989–1004. [Google Scholar] [CrossRef] [Green Version]
- Park, Y.-J.; Ryu, H.; Choi, G.; Kim, B.-S.; Hwang, E.S.; Kim, H.S.; Chung, Y. Il-27 confers a protumorigenic activity of regulatory t cells via cd39. Proc. Natl. Acad. Sci. USA 2019, 116, 3106–3111. [Google Scholar] [CrossRef] [Green Version]
- d’Almeida, S.M.; Kauffenstein, G.; Roy, C.; Basset, L.; Papargyris, L.; Henrion, D.; Catros, V.; Ifrah, N.; Descamps, P.; Croue, A. The ecto-atpdase cd39 is involved in the acquisition of the immunoregulatory phenotype by m-csf-macrophages and ovarian cancer tumor-associated macrophages: Regulatory role of il-27. Oncoimmunology 2016, 5, e1178025. [Google Scholar] [CrossRef] [Green Version]
- Pylayeva-Gupta, Y. Molecular pathways: Interleukin-35 in autoimmunity and cancer. Clin. Cancer Res. 2016, 22, 4973–4978. [Google Scholar] [CrossRef] [Green Version]
- Hao, S.; Chen, X.; Wang, F.; Shao, Q.; Liu, J.; Zhao, H.; Yuan, C.; Ren, H.; Mao, H. Breast cancer cell–derived il-35 promotes tumor progression via induction of il-35-producing induced regulatory T cells. Carcinogenesis 2018, 39, 1488–1496. [Google Scholar] [CrossRef] [Green Version]
- Lee, C.-C.; Lin, J.-C.; Hwang, W.-L.; Kuo, Y.-J.; Chen, H.-K.; Tai, S.-K.; Lin, C.-C.; Yang, M.-H. Macrophage-secreted interleukin-35 regulates cancer cell plasticity to facilitate metastatic colonization. Nat. Commun. 2018, 9, 1–18. [Google Scholar]
- Wang, Z.; Liu, J.-Q.; Liu, Z.; Shen, R.; Zhang, G.; Xu, J.; Basu, S.; Feng, Y.; Bai, X.-F. Tumor-derived il-35 promotes tumor growth by enhancing myeloid cell accumulation and angiogenesis. J. Immunol. 2013, 190, 2415–2423. [Google Scholar] [CrossRef] [Green Version]
- Collison, L.W.; Workman, C.J.; Kuo, T.T.; Boyd, K.; Wang, Y.; Vignali, K.M.; Cross, R.; Sehy, D.; Blumberg, R.S.; Vignali, D.A. The inhibitory cytokine il-35 contributes to regulatory T-cell function. Nature 2007, 450, 566–569. [Google Scholar] [CrossRef] [PubMed]
- Niedbala, W.; Wei, X.q.; Cai, B.; Hueber, A.J.; Leung, B.P.; McInnes, I.B.; Liew, F.Y. Il-35 is a novel cytokine with therapeutic effects against collagen-induced arthritis through the expansion of regulatory t cells and suppression of th17 cells. Eur. J. Immunol. 2007, 37, 3021–3029. [Google Scholar] [CrossRef] [PubMed]
- Wirtz, S.; Billmeier, U.; Mchedlidze, T.; Blumberg, R.S.; Neurath, M.F. Interleukin-35 mediates mucosal immune responses that protect against t-cell–dependent colitis. Gastroenterology 2011, 141, 1875–1886. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.-Q.; Liu, Z.; Zhang, X.; Shi, Y.; Talebian, F.; Carl, J.W.; Yu, C.; Shi, F.-D.; Whitacre, C.C.; Trgovcich, J. Increased th17 and regulatory T cell responses in ebv-induced gene 3-deficient mice lead to marginally enhanced development of autoimmune encephalomyelitis. J. Immunol. 2012, 188, 3099–3106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mirlekar, B.; Michaud, D.; Searcy, R.; Greene, K.; Pylayeva-Gupta, Y. Il35 hinders endogenous antitumor t-cell immunity and responsiveness to immunotherapy in pancreatic cancer. Cancer Immunol. Res. 2018, 6, 1014–1024. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turnis, M.E.; Sawant, D.V.; Szymczak-Workman, A.L.; Andrews, L.P.; Delgoffe, G.M.; Yano, H.; Beres, A.J.; Vogel, P.; Workman, C.J.; Vignali, D.A. Interleukin-35 limits anti-tumor immunity. Immunity 2016, 44, 316–329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, H.; Zhang, T.; Yan, M.-X.; Wu, W. Il-35 inhibits cd8 (+) T cells activity by suppressing expression of costimulatory molecule cd28 and th1 cytokine production. Transl. Cancer Res. 2019, 8, 1319–1325. [Google Scholar] [CrossRef]
- Yang, L.; Shao, X.; Jia, S.; Zhang, Q.; Jin, Z. Interleukin-35 dampens cd8+ T cells activity in patients with non-viral hepatitis-related hepatocellular carcinoma. Front. Immunol. 2019, 10, 1032. [Google Scholar] [CrossRef] [Green Version]
- Mirlekar, B.; Michaud, D.; Lee, S.J.; Kren, N.P.; Harris, C.; Greene, K.; Goldman, E.C.; Gupta, G.P.; Fields, R.C.; Hawkins, W.G. B cell–derived il35 drives stat3-dependent cd8+ T-cell exclusion in pancreatic cancer. Cancer Immunol. Res. 2020, 8, 292–308. [Google Scholar] [CrossRef] [Green Version]
- Wang, K.; Liu, J.; Li, J. Il-35-producing b cells in gastric cancer patients. Medicine 2018, 97, e0710. [Google Scholar] [CrossRef]
- Collison, L.W.; Delgoffe, G.M.; Guy, C.S.; Vignali, K.M.; Chaturvedi, V.; Fairweather, D.; Satoskar, A.R.; Garcia, K.C.; Hunter, C.A.; Drake, C.G. The composition and signaling of the il-35 receptor are unconventional. Nat. Immunol. 2012, 13, 290–299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collison, L.W.; Chaturvedi, V.; Henderson, A.L.; Giacomin, P.R.; Guy, C.; Bankoti, J.; Finkelstein, D.; Forbes, K.; Workman, C.J.; Brown, S.A. Il-35-mediated induction of a potent regulatory t cell population. Nat. Immunol. 2010, 11, 1093–1101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dambuza, I.M.; He, C.; Choi, J.K.; Yu, C.-R.; Wang, R.; Mattapallil, M.J.; Wingfield, P.T.; Caspi, R.R.; Egwuagu, C.E. Il-12p35 induces expansion of il-10 and il-35-expressing regulatory B cells and ameliorates autoimmune disease. Nat. Commun. 2017, 8, 1–12. [Google Scholar]
- Wang, R.-X.; Yu, C.-R.; Dambuza, I.M.; Mahdi, R.M.; Dolinska, M.B.; Sergeev, Y.V.; Wingfield, P.T.; Kim, S.-H.; Egwuagu, C.E. Interleukin-35 induces regulatory B cells that suppress autoimmune disease. Nat. Med. 2014, 20, 633–641. [Google Scholar] [CrossRef] [PubMed]
- Tao, Q.; Pan, Y.; Wang, Y.; Wang, H.; Xiong, S.; Li, Q.; Wang, J.; Tao, L.; Wang, Z.; Wu, F. Regulatory T cells-derived il-35 promotes the growth of adult acute myeloid leukemia blasts. Int. J. Cancer 2015, 137, 2384–2393. [Google Scholar] [CrossRef]
- Wang, H.-M.; Zhang, X.-H.; Feng, M.-M.; Qiao, Y.-J.; Ye, L.-Q.; Chen, J.; Fan, F.-F.; Guo, L.-L. Interleukin-35 suppresses the antitumor activity of T cells in patients with non-small cell lung cancer. Cell. Physiol. Biochem. 2018, 47, 2407–2419. [Google Scholar] [CrossRef]
- Zou, J.-M.; Qin, J.; Li, Y.-C.; Wang, Y.; Li, D.; Shu, Y.; Luo, C.; Wang, S.-S.; Chi, G.; Guo, F. Il-35 induces n2 phenotype of neutrophils to promote tumor growth. Oncotarget 2017, 8, 33501–33514. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Lin, Y.; Li, C.; Zhang, X.; Cheng, L.; Dai, L.; Wang, Y.; Wang, F.; Shi, G.; Li, Y. Il-35 decelerates the inflammatory process by regulating inflammatory cytokine secretion and m1/m2 macrophage ratio in psoriasis. J. Immunol. 2016, 197, 2131–2144. [Google Scholar] [CrossRef]
- Ye, J.; Huang, Y.; Que, B.; Chang, C.; Liu, W.; Hu, H.; Liu, L.; Shi, Y.; Wang, Y.; Wang, M. Interleukin-12p35 knock out aggravates doxorubicin-induced cardiac injury and dysfunction by aggravating the inflammatory response, oxidative stress, apoptosis and autophagy in mice. EBioMedicine 2018, 35, 29–39. [Google Scholar] [CrossRef] [Green Version]
- Haller, S.; Duval, A.; Migliorini, R.; Stevanin, M.; Mack, V.; Acha-Orbea, H. Interleukin-35-producing cd8α+ dendritic cells acquire a tolerogenic state and regulate t cell function. Front. Immunol. 2017, 8, 98. [Google Scholar] [CrossRef] [Green Version]
- Dixon, K.O.; van der Kooij, S.W.; Vignali, D.A.; van Kooten, C. Human tolerogenic dendritic cells produce il-35 in the absence of other il-12 family members. Eur. J. Immunol. 2015, 45, 1736–1747. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Sun, Y.; Zheng, Y.; Zhang, M.; Jin, X.; Kang, K.; Wang, Y.; Li, S.; Zhang, H.; Zhao, Q. Administration of interleukin-35-conditioned autologous tolerogenic dendritic cells prolong allograft survival after heart transplantation. Cell. Physiol. Biochem. 2018, 49, 1221–1237. [Google Scholar] [CrossRef] [PubMed]
- Xue, W.; Yan, D.; Kan, Q. Interleukin-35 as an emerging player in tumor microenvironment. J. Cancer 2019, 10, 2074–2082. [Google Scholar] [CrossRef] [Green Version]
- Zhu, J.; Yang, X.; Wang, Y.; Zhang, H.; Guo, Z. Interleukin-35 is associated with the tumorigenesis and progression of prostate cancer. Oncol. Lett. 2019, 17, 5094–5102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heim, L.; Kachler, K.; Siegmund, R.; Trufa, D.I.; Mittler, S.; Geppert, C.-I.; Friedrich, J.; Rieker, R.J.; Sirbu, H.; Finotto, S. Increased expression of the immunosuppressive interleukin-35 in patients with non-small cell lung cancer. Br. J. Cancer 2019, 120, 903–912. [Google Scholar] [CrossRef] [Green Version]
- Ma, Y.; Chen, L.; Xie, G.; Zhou, Y.; Yue, C.; Yuan, X.; Zheng, Y.; Wang, W.; Deng, L.; Shen, L. Elevated level of interleukin-35 in colorectal cancer induces conversion of t cells into itr35 by activating stat1/stat3. Oncotarget 2016, 7, 73003–73015. [Google Scholar] [CrossRef]
- Zhang, J.; Mao, T.; Wang, S.; Wang, D.; Niu, Z.; Sun, Z.; Zhang, J. Interleukin-35 expression is associated with colon cancer progression. Oncotarget 2017, 8, 71563–71573. [Google Scholar] [CrossRef] [Green Version]
- Sun, M.; Zheng, X.; Meng, Q.; Dong, Y.; Zhang, G.; Rao, D.; An, X.; Yang, Z.; Pan, L.; Zhang, S. Interleukin-35 expression in non-small cell lung cancer is associated with tumor progression. Cell. Physiol. Biochem. 2018, 51, 1839–1851. [Google Scholar] [CrossRef]
- Jiang, Q.; Ma, L.; Li, R.; Sun, J. Colon cancer-induced interleukin-35 inhibits beta-catenin-mediated pro-oncogenic activity. Oncotarget 2018, 9, 11989–11998. [Google Scholar] [CrossRef]
- Long, J.; Guo, H.; Cui, S.; Zhang, H.; Liu, X.; Li, D.; Han, Z.; Xi, L.; Kou, W.; Xu, J. Il-35 expression in hepatocellular carcinoma cells is associated with tumor progression. Oncotarget 2016, 7, 45678–45686. [Google Scholar] [CrossRef] [Green Version]
- Chatrabnous, N.; Ghaderi, A.; Ariafar, A.; Razeghinia, M.S.; Nemati, M.; Jafarzadeh, A. Serum concentration of interleukin-35 and its association with tumor stages and foxp3 gene polymorphism in patients with prostate cancer. Cytokine 2019, 113, 221–227. [Google Scholar] [CrossRef]
- Chen, G.; Liang, Y.; Guan, X.; Chen, H.; Liu, Q.; Lin, B.; Chen, C.; Huang, M.; Chen, J.; Wu, W. Circulating low il-23: Il-35 cytokine ratio promotes progression associated with poor prognosisin breast cancer. Am. J. Transl. Res. 2016, 8, 2255–2264. [Google Scholar]
- Larousserie, F.; Kebe, D.; Huynh, T.; Audebourg, A.; Tamburini, J.; Terris, B.; Devergne, O. Evidence for il-35 expression in diffuse large b-cell lymphoma and impact on the patient’s prognosis. Front. Oncol. 2019, 9, 563. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Tao, Q.; Wang, H.; Wang, Z.; Wu, F.; Pan, Y.; Tao, L.; Xiong, S.; Wang, Y.; Zhai, Z. Elevated il-35 in bone marrow of the patients with acute myeloid leukemia. Hum. Immunol. 2015, 76, 681–686. [Google Scholar] [CrossRef]
- Huang, C.; Li, N.; Li, Z.; Chang, A.; Chen, Y.; Zhao, T.; Li, Y.; Wang, X.; Zhang, W.; Wang, Z. Tumour-derived interleukin 35 promotes pancreatic ductal adenocarcinoma cell extravasation and metastasis by inducing icam1 expression. Nat. Commun. 2017, 8, 1–15. [Google Scholar]
- Nicholl, M.B.; Ledgewood, C.L.; Chen, X.; Bai, Q.; Qin, C.; Cook, K.M.; Herrick, E.J.; Diaz-Arias, A.; Moore, B.J.; Fang, Y. Il-35 promotes pancreas cancer growth through enhancement of proliferation and inhibition of apoptosis: Evidence for a role as an autocrine growth factor. Cytokine 2014, 70, 126–133. [Google Scholar] [CrossRef]
- Jin, P.; Ren, H.; Sun, W.; Xin, W.; Zhang, H.; Hao, J. Circulating il-35 in pancreatic ductal adenocarcinoma patients. Hum. Immunol. 2014, 75, 29–33. [Google Scholar] [CrossRef]
- Wang, X.; Wei, Y.; Xiao, H.; Liu, X.; Zhang, Y.; Han, G.; Chen, G.; Hou, C.; Ma, N.; Shen, B. A novel il-23p19/ebi3 (il-39) cytokine mediates inflammation in lupus-like mice. Eur. J. Immunol. 2016, 46, 1343–1350. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Liu, X.; Zhang, Y.; Wang, Z.; Zhu, G.; Han, G.; Chen, G.; Hou, C.; Wang, T.; Ma, N. Interleukin (il)-39 [il-23p19/epstein–barr virus-induced 3 (ebi3)] induces differentiation/expansion of neutrophils in lupus-prone mice. Clin. Exp. Immunol. 2016, 186, 144–156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manning, A.A.; Zhao, L.; Zhu, Z.; Xiao, H.; Redington, C.G.; Ding, V.A.; Stewart-Hester, T.; Bai, Q.; Dunlap, J.; Wakefield, M.R. Il-39 acts as a friend to pancreatic cancer. Med. Oncol. 2019, 36, 22. [Google Scholar] [CrossRef] [Green Version]
- Hawkes, J.E.; Yan, B.Y.; Chan, T.C.; Krueger, J.G. Discovery of the il-23/il-17 signaling pathway and the treatment of psoriasis. J. Immunol. 2018, 201, 1605–1613. [Google Scholar] [CrossRef]
- Hasegawa, H.; Mizoguchi, I.; Chiba, Y.; Ohashi, M.; Xu, M.; Yoshimoto, T. Expanding diversity in molecular structures and functions of the il-6/il-12 heterodimeric cytokine family. Front. Immunol. 2016, 7, 479. [Google Scholar] [CrossRef] [Green Version]
- Floss, D.; Schönberg, M.; Franke, M.; Horstmeier, F.; Engelowski, E.; Schneider, A.; Rosenfeldt, E.; Scheller, J. Il-6/il-12 cytokine receptor shuffling of extra-and intracellular domains reveals canonical stat activation via synthetic il-35 and il-39 signaling. Sci. Rep. 2017, 7, 1–13. [Google Scholar]
- Hu, J.; Sun, C.; Bernatchez, C.; Xia, X.; Hwu, P.; Dotti, G.; Li, S. T-cell homing therapy for reducing regulatory T cells and preserving effector t-cell function in large solid tumors. Clin. Cancer Res. 2018, 24, 2920–2934. [Google Scholar] [CrossRef] [Green Version]
- Berraondo, P.; Etxeberria, I.; Ponz-Sarvise, M.; Melero, I. Revisiting interleukin-12 as a cancer immunotherapy agent. Clin. Cancer Res. 2018, 24, 2716–2718. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Role of IL-12 family cytokines in maintaining a balance between effector and regulatory immune responses in tumorigenesis. IL-12 activates an effector immune response against tumor cells by promoting both M1 macrophage polarization and IFN-γ- production by Th1 cells, which in turn, stimulate anti-tumor cytotoxic CD8+ and NK cells. IL-27 and IL-23 have dual effects on immune cells in cancer. IL-27 and IL-23 can induce an overall T cell-mediated immune response and also modulate immune suppressive macrophages. Furthermore, IL-23 can stimulate the proliferation and growth of tumor cells. Conversely, IL-35 is a strong immune suppressive cytokine; it induces regulatory B and T cell activation and proliferation that subverts anti-tumor immunity and stimulates tumor growth and metastasis. IL-39 was recently shown to be secreted by B cells and may increase cancer cell proliferation. ↑ Arrow indicates increase in respective cell type activity; Tr1—T regulatory type 1; ILC—innate lymphoid cell; Breg—regulatory B cell; Treg—regulatory T cell; iT35—IL35-inducible regulatory T cell.
Figure 1.
Role of IL-12 family cytokines in maintaining a balance between effector and regulatory immune responses in tumorigenesis. IL-12 activates an effector immune response against tumor cells by promoting both M1 macrophage polarization and IFN-γ- production by Th1 cells, which in turn, stimulate anti-tumor cytotoxic CD8+ and NK cells. IL-27 and IL-23 have dual effects on immune cells in cancer. IL-27 and IL-23 can induce an overall T cell-mediated immune response and also modulate immune suppressive macrophages. Furthermore, IL-23 can stimulate the proliferation and growth of tumor cells. Conversely, IL-35 is a strong immune suppressive cytokine; it induces regulatory B and T cell activation and proliferation that subverts anti-tumor immunity and stimulates tumor growth and metastasis. IL-39 was recently shown to be secreted by B cells and may increase cancer cell proliferation. ↑ Arrow indicates increase in respective cell type activity; Tr1—T regulatory type 1; ILC—innate lymphoid cell; Breg—regulatory B cell; Treg—regulatory T cell; iT35—IL35-inducible regulatory T cell.

Figure 2.
The therapeutic modulation of IL-12 family cytokines may enhance the efficacy of conventional therapy. Recent studies indicate that the therapies targeting (either upregulating or downregulating) the IL-12 family cytokines in combination with other standard therapies may increase treatment effectiveness. Context-dependent functions of IL-12 cytokines in cancer could drive the development of inhibitor or augmentative therapy axes, respectively. Drugs or antibodies targeting IL-12 family cytokines may help to restrain immune suppression within the tumor microenvironment (TME) and allow for the infiltration and proliferation of anti-tumor immune cells. Additionally, the targeted delivery of these cytokines with the help of adenovirus or chimeric antigen receptor (CAR) T cells may enhance cytotoxicity and tumor cell clearance. Such approaches could make tumor cells more sensitive to radiation, chemotherapy and immune checkpoint blockade therapy.
Figure 2.
The therapeutic modulation of IL-12 family cytokines may enhance the efficacy of conventional therapy. Recent studies indicate that the therapies targeting (either upregulating or downregulating) the IL-12 family cytokines in combination with other standard therapies may increase treatment effectiveness. Context-dependent functions of IL-12 cytokines in cancer could drive the development of inhibitor or augmentative therapy axes, respectively. Drugs or antibodies targeting IL-12 family cytokines may help to restrain immune suppression within the tumor microenvironment (TME) and allow for the infiltration and proliferation of anti-tumor immune cells. Additionally, the targeted delivery of these cytokines with the help of adenovirus or chimeric antigen receptor (CAR) T cells may enhance cytotoxicity and tumor cell clearance. Such approaches could make tumor cells more sensitive to radiation, chemotherapy and immune checkpoint blockade therapy.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Mirlekar, B.; Pylayeva-Gupta, Y. IL-12 Family Cytokines in Cancer and Immunotherapy. Cancers 2021, 13, 167. https://doi.org/10.3390/cancers13020167
AMA Style
Mirlekar B, Pylayeva-Gupta Y. IL-12 Family Cytokines in Cancer and Immunotherapy. Cancers. 2021; 13(2):167. https://doi.org/10.3390/cancers13020167
Chicago/Turabian StyleMirlekar, Bhalchandra, and Yuliya Pylayeva-Gupta. 2021. "IL-12 Family Cytokines in Cancer and Immunotherapy" Cancers 13, no. 2: 167. https://doi.org/10.3390/cancers13020167
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.