
Polymorphisms within Autophagy-Related Genes Influence the Risk of Developing Colorectal Cancer: A Meta-Analysis of Four Large Cohorts

 ,
,  , , , , , , , , , add
Show full author list
, , , , , , , , , add
Show full author list
Abstract
:Simple Summary
Abstract
1. Introduction
2. Material and Methods
2.1. Study Populations
2.2. Gene and SNP Selection, Association Analysis, and Meta-Analysis
2.3. Genotyping of Imputed SNPs in the CORSA and Czech Republic CCS Cohorts
2.4. Functional Association of the Autophagy-Related Variants with Immune Responses
2.5. Correlation between Autophagy-Related SNPs and Serum Steroid Hormone Levels
2.6. Correlation of Autophagy SNPs and Blood Cell Counts and Serum/Plasmatic Proteomic Profile
2.7. Impact of Autophagy-Related Variants on the Autophagy Flux
2.8. In Silico Functional and eQTL Analysis
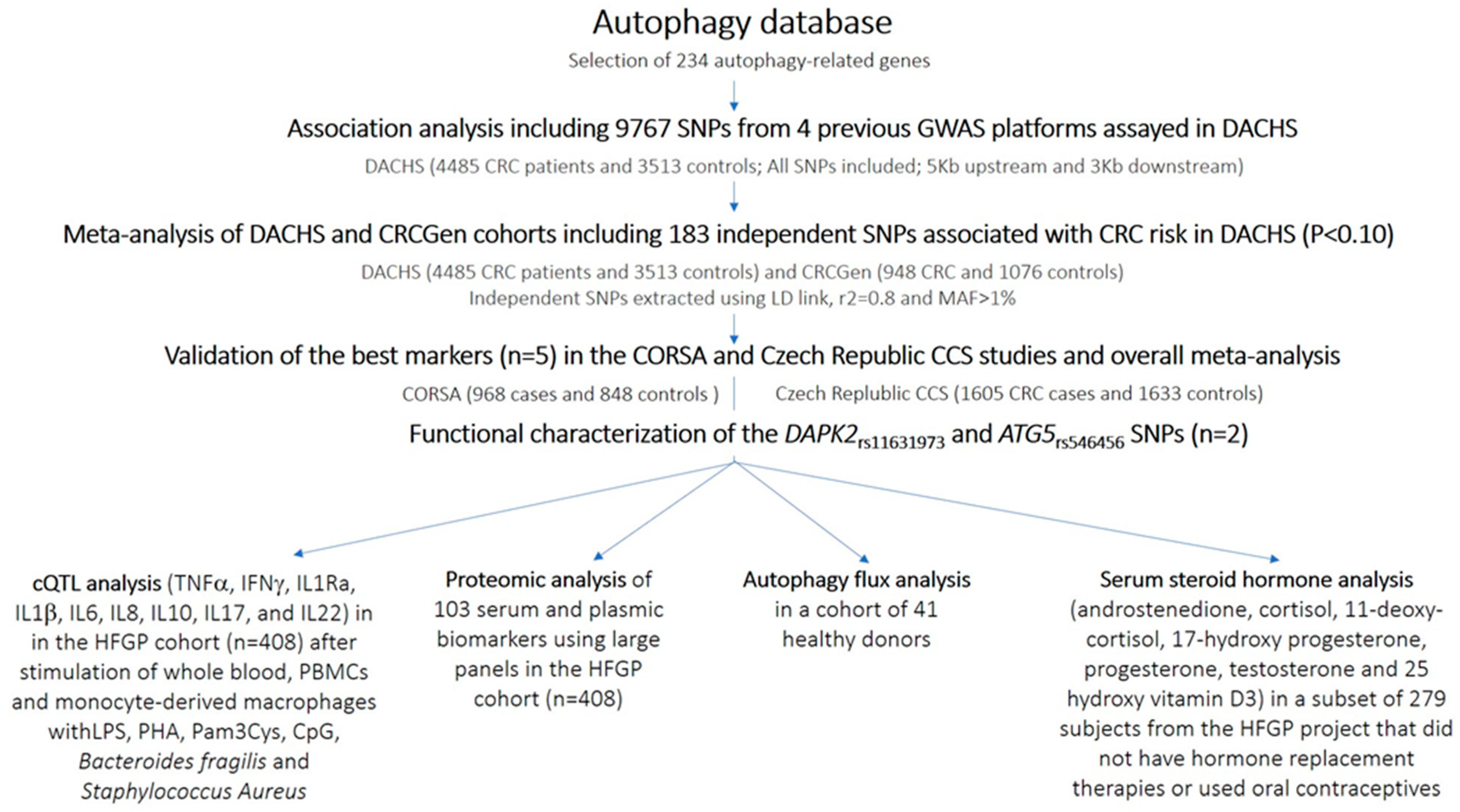
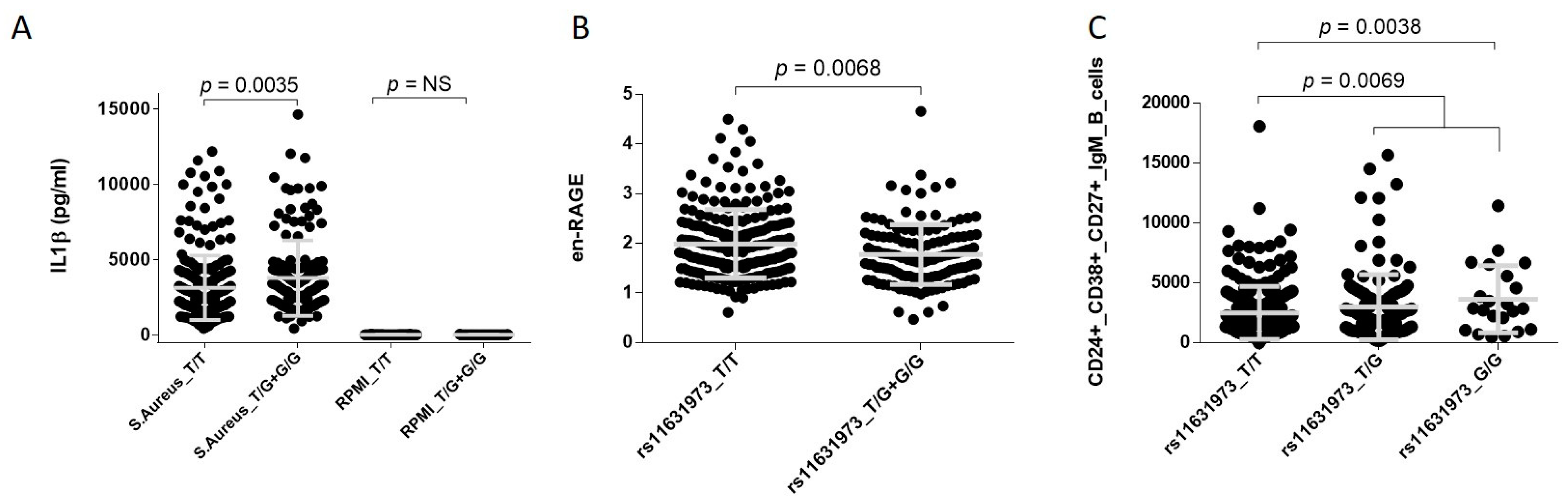
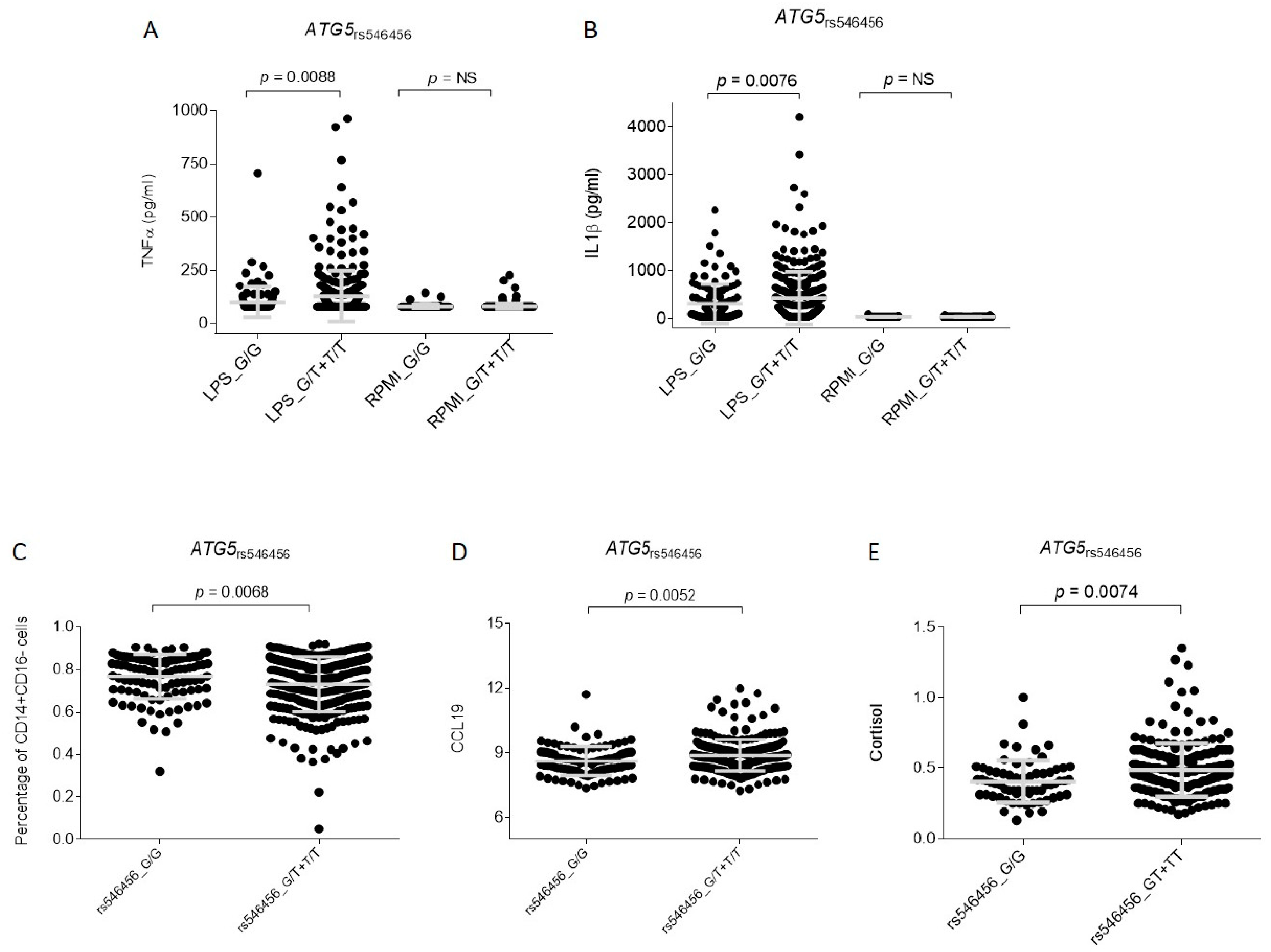
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Siegel, R.; Naishadham, D.; Jemal, A. Cancer statistics, 2013. CA Cancer J. Clin. 2013, 63, 11–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Punt, C.J.; Tol, J. More is less—combining targeted therapies in metastatic colorectal cancer. Nat. Rev. Clin. Oncol. 2009, 6, 731–733. [Google Scholar] [CrossRef]
- Huyghe, J.R.; Bien, S.A.; Harrison, T.A.; Kang, H.M.; Chen, S.; Schmit, S.L.; Conti, D.V.; Qu, C.; Jeon, J.; Edlund, C.K.; et al. Discovery of common and rare genetic risk variants for colorectal cancer. Nat. Genet. 2019, 51, 76–87. [Google Scholar] [CrossRef]
- Song, M.; Chan, A.T.; Sun, J. Influence of the Gut Microbiome, Diet, and Environment on Risk of Colorectal Cancer. Gastroenterology 2020, 158, 322–340. [Google Scholar] [CrossRef] [PubMed]
- Burada, F.; Nicoli, E.R.; Ciurea, M.E.; Uscatu, D.C.; Ioana, M.; Gheonea, D.I. Autophagy in colorectal cancer: An important switch from physiology to pathology. World J. Gastrointest. Oncol. 2015, 7, 271–284. [Google Scholar] [CrossRef] [PubMed]
- Degenhardt, K.; Mathew, R.; Beaudoin, B.; Bray, K.; Anderson, D.; Chen, G.; Mukherjee, C.; Shi, Y.; Gelinas, C.; Fan, Y.; et al. Autophagy promotes tumor cell survival and restricts necrosis, inflammation, and tumorigenesis. Cancer Cell 2006, 10, 51–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, J.M.; Huang, S.; Wu, T.T.; Foster, N.R.; Sinicrope, F.A. Prognostic impact of Beclin 1, p62/sequestosome 1 and LC3 protein expression in colon carcinomas from patients receiving 5-fluorouracil as adjuvant chemotherapy. Cancer Biol. Ther. 2013, 14, 100–107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coppola, D.; Khalil, F.; Eschrich, S.A.; Boulware, D.; Yeatman, T.; Wang, H.G. Down-regulation of Bax-interacting factor-1 in colorectal adenocarcinoma. Cancer 2008, 113, 2665–2670. [Google Scholar] [CrossRef]
- Yang, H.Z.; Ma, Y.; Zhou, Y.; Xu, L.M.; Chen, X.J.; Ding, W.B.; Zou, H.B. Autophagy contributes to the enrichment and survival of colorectal cancer stem cells under oxaliplatin treatment. Cancer Lett. 2015, 361, 128–136. [Google Scholar] [CrossRef]
- Patel, S.; Hurez, V.; Nawrocki, S.T.; Goros, M.; Michalek, J.; Sarantopoulos, J.; Curiel, T.; Mahalingam, D. Vorinostat and hydroxychloroquine improve immunity and inhibit autophagy in metastatic colorectal cancer. Oncotarget 2016. [Google Scholar] [CrossRef] [Green Version]
- Galluzzi, L.; Pietrocola, F.; Bravo-San Pedro, J.M.; Amaravadi, R.K.; Baehrecke, E.H.; Cecconi, F.; Codogno, P.; Debnath, J.; Gewirtz, D.A.; Karantza, V.; et al. Autophagy in malignant transformation and cancer progression. EMBO J. 2015, 34, 856–880. [Google Scholar] [CrossRef] [PubMed]
- Matsuzawa-Ishimoto, Y.; Hwang, S.; Cadwell, K. Autophagy and Inflammation. Annu. Rev. Immunol. 2018, 36, 73–101. [Google Scholar] [CrossRef] [PubMed]
- Deretic, V.; Saitoh, T.; Akira, S. Autophagy in infection, inflammation and immunity. Nat. Rev. Immunol. 2013, 13, 722–737. [Google Scholar] [CrossRef] [PubMed]
- Ngabire, D.; Kim, G.D. Autophagy and Inflammatory Response in the Tumor Microenvironment. Int. J. Mol. Sci. 2017, 18, 2016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nicoli, E.R.; Dumitrescu, T.; Uscatu, C.D.; Popescu, F.D.; Streata, I.; Sosoi, S.S.; Ivanov, P.; Dumitrescu, A.; Barbalan, A.; Lungulescu, D.; et al. Determination of autophagy gene ATG16L1 polymorphism in human colorectal cancer. Rom. J. Morphol. Embryol. Rev. Roum. Morphol. Embryol. 2014, 55, 57–62. [Google Scholar]
- Zeng, C.; Matsuda, K.; Jia, W.H.; Chang, J.; Kweon, S.S.; Xiang, Y.B.; Shin, A.; Jee, S.H.; Kim, D.H.; Zhang, B.; et al. Identification of Susceptibility Loci and Genes for Colorectal Cancer Risk. Gastroenterology 2016, 150, 1633–1645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grimm, W.A.; Messer, J.S.; Murphy, S.F.; Nero, T.; Lodolce, J.P.; Weber, C.R.; Logsdon, M.F.; Bartulis, S.; Sylvester, B.E.; Springer, A.; et al. The Thr300Ala variant in ATG16L1 is associated with improved survival in human colorectal cancer and enhanced production of type I interferon. Gut 2016, 65, 456–464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, Y.; Galluzzi, L.; Zitvogel, L.; Kroemer, G. Autophagy and cellular immune responses. Immunity 2013, 39, 211–227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernandez-Rozadilla, C.; Cazier, J.B.; Tomlinson, I.P.; Carvajal-Carmona, L.G.; Palles, C.; Lamas, M.J.; Baiget, M.; Lopez-Fernandez, L.A.; Brea-Fernandez, A.; Abuli, A.; et al. A colorectal cancer genome-wide association study in a Spanish cohort identifies two variants associated with colorectal cancer risk at 1p33 and 8p12. BMC Genom. 2013, 14, 55. [Google Scholar] [CrossRef] [Green Version]
- Guo, F.; Weigl, K.; Carr, P.R.; Heisser, T.; Jansen, L.; Knebel, P.; Chang-Claude, J.; Hoffmeister, M.; Brenner, H. Use of Polygenic Risk Scores to Select Screening Intervals After Negative Findings From Colonoscopy. Clin. Gastroenterol. Hepatol. Off. Clin. Pract. J. Am. Gastroenterol. Assoc. 2020. [Google Scholar] [CrossRef] [PubMed]
- Schmit, S.L.; Edlund, C.K.; Schumacher, F.R.; Gong, J.; Harrison, T.A.; Huyghe, J.R.; Qu, C.; Melas, M.; Van Den Berg, D.J.; Wang, H.; et al. Novel Common Genetic Susceptibility Loci for Colorectal Cancer. J. Natl. Cancer Inst. 2019, 111, 146–157. [Google Scholar] [CrossRef] [PubMed]
- Hofer, P.; Hagmann, M.; Brezina, S.; Dolejsi, E.; Mach, K.; Leeb, G.; Baierl, A.; Buch, S.; Sutterluty-Fall, H.; Karner-Hanusch, J.; et al. Bayesian and frequentist analysis of an Austrian genome-wide association study of colorectal cancer and advanced adenomas. Oncotarget 2017, 8, 98623–98634. [Google Scholar] [CrossRef] [PubMed]
- Peters, U.; Jiao, S.; Schumacher, F.R.; Hutter, C.M.; Aragaki, A.K.; Baron, J.A.; Berndt, S.I.; Bezieau, S.; Brenner, H.; Butterbach, K.; et al. Identification of Genetic Susceptibility Loci for Colorectal Tumors in a Genome-Wide Meta-analysis. Gastroenterology 2013, 144, 799–807.e724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schumacher, F.R.; Schmit, S.L.; Jiao, S.; Edlund, C.K.; Wang, H.; Zhang, B.; Hsu, L.; Huang, S.C.; Fischer, C.P.; Harju, J.F.; et al. Genome-wide association study of colorectal cancer identifies six new susceptibility loci. Nat. Commun. 2015, 6, 7138. [Google Scholar] [CrossRef] [Green Version]
- Weigl, K.; Chang-Claude, J.; Knebel, P.; Hsu, L.; Hoffmeister, M.; Brenner, H. Strongly enhanced colorectal cancer risk stratification by combining family history and genetic risk score. Clin. Epidemiol. 2018, 10, 143–152. [Google Scholar] [CrossRef] [Green Version]
- Law, P.J.; Timofeeva, M.; Fernandez-Rozadilla, C.; Broderick, P.; Studd, J.; Fernandez-Tajes, J.; Farrington, S.; Svinti, V.; Palles, C.; Orlando, G.; et al. Association analyses identify 31 new risk loci for colorectal cancer susceptibility. Nat. Commun. 2019, 10, 2154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanchez-Maldonado, J.M.; Martinez-Bueno, M.; Canhao, H.; Ter Horst, R.; Munoz-Pena, S.; Moniz-Diez, A.; Rodriguez-Ramos, A.; Escudero, A.; Sorensen, S.B.; Hetland, M.L.; et al. NFKB2 polymorphisms associate with the risk of developing rheumatoid arthritis and response to TNF inhibitors: Results from the REPAIR consortium. Sci. Rep. 2020, 10, 4316. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Oosting, M.; Smeekens, S.P.; Jaeger, M.; Aguirre-Gamboa, R.; Le, K.T.T.; Deelen, P.; Ricano-Ponce, I.; Schoffelen, T.; Jansen, A.F.M.; et al. A Functional Genomics Approach to Understand Variation in Cytokine Production in Humans. Cell 2016, 167, 1099–1110.e1014. [Google Scholar] [CrossRef] [Green Version]
- Aguirre-Gamboa, R.; Joosten, I.; Urbano, P.C.M.; van der Molen, R.G.; van Rijssen, E.; van Cranenbroek, B.; Oosting, M.; Smeekens, S.; Jaeger, M.; Zorro, M.; et al. Differential Effects of Environmental and Genetic Factors on T and B Cell Immune Traits. Cell Rep. 2016, 17, 2474–2487. [Google Scholar] [CrossRef] [Green Version]
- Orru, V.; Steri, M.; Sole, G.; Sidore, C.; Virdis, F.; Dei, M.; Lai, S.; Zoledziewska, M.; Busonero, F.; Mulas, A.; et al. Genetic variants regulating immune cell levels in health and disease. Cell 2013, 155, 242–256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Westra, H.J.; Peters, M.J.; Esko, T.; Yaghootkar, H.; Schurmann, C.; Kettunen, J.; Christiansen, M.W.; Fairfax, B.P.; Schramm, K.; Powell, J.E.; et al. Systematic identification of trans eQTLs as putative drivers of known disease associations. Nat. Genet. 2013, 45, 1238–1243. [Google Scholar] [CrossRef] [Green Version]
- Ber, Y.; Shiloh, R.; Gilad, Y.; Degani, N.; Bialik, S.; Kimchi, A. DAPK2 is a novel regulator of mTORC1 activity and autophagy. Cell Death Differ. 2015, 22, 465–475. [Google Scholar] [CrossRef] [Green Version]
- Geering, B.; Stoeckle, C.; Rozman, S.; Oberson, K.; Benarafa, C.; Simon, H.U. DAPK2 positively regulates motility of neutrophils and eosinophils in response to intermediary chemoattractants. J. Leukoc. Biol. 2014, 95, 293–303. [Google Scholar] [CrossRef] [PubMed]
- Geering, B.; Zokouri, Z.; Hurlemann, S.; Gerrits, B.; Auslander, D.; Britschgi, A.; Tschan, M.P.; Simon, H.U.; Fussenegger, M. Identification of Novel Death-Associated Protein Kinase 2 Interaction Partners by Proteomic Screening Coupled with Bimolecular Fluorescence Complementation. Mol. Cell. Biol. 2016, 36, 132–143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rizzi, M.; Tschan, M.P.; Britschgi, C.; Britschgi, A.; Hugli, B.; Grob, T.J.; Leupin, N.; Mueller, B.U.; Simon, H.U.; Ziemiecki, A.; et al. The death-associated protein kinase 2 is up-regulated during normal myeloid differentiation and enhances neutrophil maturation in myeloid leukemic cells. J. Leukoc. Biol. 2007, 81, 1599–1608. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Humbert, M.; Federzoni, E.A.; Britschgi, A.; Schlafli, A.M.; Valk, P.J.; Kaufmann, T.; Haferlach, T.; Behre, G.; Simon, H.U.; Torbett, B.E.; et al. The tumor suppressor gene DAPK2 is induced by the myeloid transcription factors PU.1 and C/EBPalpha during granulocytic differentiation but repressed by PML-RARalpha in APL. J. Leukoc. Biol. 2014, 95, 83–93. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Liu, L.; Sun, Y.; Xiang, J.; Zhou, D.; Wang, L.; Xu, H.; Yang, X.; Du, N.; Zhang, M.; et al. MicroRNA-520g promotes epithelial ovarian cancer progression and chemoresistance via DAPK2 repression. Oncotarget 2016, 7, 26516–26534. [Google Scholar] [CrossRef] [Green Version]
- Furuta, G.T.; Nieuwenhuis, E.E.; Karhausen, J.; Gleich, G.; Blumberg, R.S.; Lee, J.J.; Ackerman, S.J. Eosinophils alter colonic epithelial barrier function: Role for major basic protein. Am. J. Physiol. Gastrointest. Liver Physiol. 2005, 289, G890–G897. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McLoed, A.G.; Sherrill, T.P.; Cheng, D.S.; Han, W.; Saxon, J.A.; Gleaves, L.A.; Wu, P.; Polosukhin, V.V.; Karin, M.; Yull, F.E.; et al. Neutrophil-Derived IL-1beta Impairs the Efficacy of NF-kappaB Inhibitors against Lung Cancer. Cell Rep. 2016, 16, 120–132. [Google Scholar] [CrossRef] [Green Version]
- Butt, J.; Epplein, M. Helicobacter pylori and colorectal cancer-A bacterium going abroad? PLoS Pathog. 2019, 15, e1007861. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Correa, P.; Houghton, J. Carcinogenesis of Helicobacter pylori. Gastroenterology 2007, 133, 659–672. [Google Scholar] [CrossRef] [PubMed]
- Shimabukuro-Vornhagen, A.; Schlosser, H.A.; Gryschok, L.; Malcher, J.; Wennhold, K.; Garcia-Marquez, M.; Herbold, T.; Neuhaus, L.S.; Becker, H.J.; Fiedler, A.; et al. Characterization of tumor-associated B-cell subsets in patients with colorectal cancer. Oncotarget 2014, 5, 4651–4664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Wang, Y.; Lu, Y.; Zhang, Q.; Qu, X. Heterozygous deletion of ATG5 in Apc(Min/+) mice promotes intestinal adenoma growth and enhances the antitumor efficacy of interferon-gamma. Cancer Biol. Ther. 2015, 16, 383–391. [Google Scholar] [CrossRef] [Green Version]
- Lauzier, A.; Normandeau-Guimond, J.; Vaillancourt-Lavigueur, V.; Boivin, V.; Charbonneau, M.; Rivard, N.; Scott, M.S.; Dubois, C.M.; Jean, S. Colorectal cancer cells respond differentially to autophagy inhibition in vivo. Sci. Rep. 2019, 9, 11316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Qi, J.; Yu, J.; Chen, H.; Zou, Z.; Lin, X.; Guo, L. Overexpression of Rictor protein in colorectal cancer is correlated with tumor progression and prognosis. Oncol. Lett. 2017, 14, 6198–6202. [Google Scholar] [CrossRef] [Green Version]
- Iula, L.; Keitelman, I.A.; Sabbione, F.; Fuentes, F.; Guzman, M.; Galletti, J.G.; Gerber, P.P.; Ostrowski, M.; Geffner, J.R.; Jancic, C.C.; et al. Autophagy Mediates Interleukin-1beta Secretion in Human Neutrophils. Front. Immunol. 2018, 9, 269. [Google Scholar] [CrossRef] [Green Version]
- Xu, Z.; Zhu, C.; Chen, C.; Zong, Y.; Feng, H.; Liu, D.; Feng, W.; Zhao, J.; Lu, A. CCL19 suppresses angiogenesis through promoting miR-206 and inhibiting Met/ERK/Elk-1/HIF-1alpha/VEGF-A pathway in colorectal cancer. Cell Death Dis. 2018, 9, 974. [Google Scholar] [CrossRef] [Green Version]
- Saitoh, T.; Akira, S. Regulation of inflammasomes by autophagy. J. Allergy Clin. Immunol. 2016, 138, 28–36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pliyev, B.K.; Menshikov, M. Differential effects of the autophagy inhibitors 3-methyladenine and chloroquine on spontaneous and TNF-alpha-induced neutrophil apoptosis. Apoptosis 2012, 17, 1050–1065. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Zhang, Y.; He, Z.; Yin, K.; Li, B.; Zhang, L.; Xu, Z. Chronic stress promotes gastric cancer progression and metastasis: An essential role for ADRB2. Cell Death Dis. 2019, 10, 788. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moreno-Smith, M.; Lutgendorf, S.K.; Sood, A.K. Impact of stress on cancer metastasis. Future Oncol. 2010, 6, 1863–1881. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swertz, M.A.; Dijkstra, M.; Adamusiak, T.; van der Velde, J.K.; Kanterakis, A.; Roos, E.T.; Lops, J.; Thorisson, G.A.; Arends, D.; Byelas, G.; et al. The MOLGENIS toolkit: Rapid prototyping of biosoftware at the push of a button. BMC Bioinform. 2010, 11 (Suppl. 12), S12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilkinson, M.D.; Dumontier, M.; Aalbersberg, I.J.; Appleton, G.; Axton, M.; Baak, A.; Blomberg, N.; Boiten, J.W.; da Santos, L.B.S.; Bourne, P.E.; et al. Addendum: The FAIR Guiding Principles for scientific data management and stewardship. Sci. Data 2019, 6, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilkinson, M.D.; Dumontier, M.; Aalbersberg, I.J.; Appleton, G.; Axton, M.; Baak, A.; Blomberg, N.; Boiten, J.W.; da Santos, L.B.S.; Bourne, P.E.; et al. The FAIR Guiding Principles for scientific data management and stewardship. Sci. Data 2016, 3, 160018. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Gene | Variant_dbSNP | Risk Allele | DACHS Cohort (n = 7998) | CRCGen Cohort (n = 2024) | Meta-Analysis (n = 10,022) | pHet | |||
|---|---|---|---|---|---|---|---|---|---|
| OR (95% CI) a | p-Value | OR (95% CI) a | p-Value | OR (95% CI) a | p-Value | ||||
| NRG3 | rs11196336 | C | 1.18 (1.10–1.28) | 0.00002 | 1.09 (0.93–1.28) | 0.3125 | 1.17 (1.09–1.25) | 0.0000185 | 0.354 |
| DAPK2 | rs11631973 | G | 1.15 (1.06–1.24) | 0.00060 | 1.17 (1.00–1.36) | 0.0563 | 1.15 (1.07–1.23) | 0.0000799 | 0.846 |
| EGFR | rs2075108 | A | 1.15 (1.03–1.28) | 0.01308 | 1.39 (1.11–1.73) | 0.0035 | 1.19 (1.08–1.31) | 0.000457 | 0.134 |
| TP73 | rs4648553 | A | 1.10 (1.02–1.18) | 0.01103 | 1.21 (1.05–1.39) | 0.0104 | 1.12 (1.05–1.20) | 0.000590 | 0.254 |
| LOC100128105 | rs6565506 | C | 1.09 (1.01–1.17) | 0.02129 | 1.21 (1.04–1.41) | 0.0122 | 1.11 (1.04–1.19) | 0.001597 | 0.214 |
| ATG5 | rs546456 | T | 1.09 (1.03–1.16) | 0.00574 | 1.10 (0.97–1.25) | 0.1324 | 1.09 (1.03–1.16) | 0.001700 | 0.918 |
| Gene_Variant | Risk Allele | DACHS Cohort (n = 7998) | CRCGen Cohort (n = 2024) | CORSA (n = 1816) | Czech Republic CCS (n = 3238) | OR (95% CI) a | p-Value | pCorr | pHet | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| OR (95% CI) a | p-Value | OR (95% CI) a | p-Value | OR (95% CI) a | p-Value | OR (95% CI) a | p-Value | ||||||
| DAPK2rs11631973 | G | 1.15 (1.06–1.24) | 0.0006 | 1.17 (1.00–1.36) | 0.056 | 1.12 (0.96–1.30) | 0.14 | 1.07 (0.96–1.20) | 0.25 | 1.13 (1.07–1.19) | 2.19 × 10−5 | 0.0041 | 0.760 |
| DAPK2rs11635284 | G | 1.13 (1.05–1.22) | 0.001 | 1.19 (1.02–1.39) | 0.032 | 1.12 (0.96–1.30) | 0.15 | 1.06 (0.95–1.19) | 0.28 | 1.12 (1.06–1.18) | 4.77 × 10−5 | 0.0087 | 0.679 |
| ATG5rs546456 | T | 1.09 (1.03–1.16) | 0.006 | 1.10 (0.97–1.25) | 0.132 | 1.07 (0.94–1.22) | 0.33 | 1.06 (0.95–1.18) | 0.28 | 1.08 (1.04–1.14) | 0.00062 | 0.11 | 0.958 |
| ATG5rs490010 | G | 1.08 (1.01–1.15) | 0.020 | 1.09 (0.96–1.23) | 0.181 | 1.09 (0.96–1.25) | 0.18 | 1.06 (0.95–1.19) | 0.30 | 1.08 (1.03–1.13) | 0.002 | NS | 0.985 |
| NRG3rs11196336 | C | 1.18 (1.10–1.28) | 0.00002 | 1.09 (0.93–1.28) | 0.313 | 1.11 (0.95–1.30) | 0.19 | 0.94 (0.82–1.08) | 0.25 | 1.11 (1.05–1.18) | 0.00025 | 0.045 | 0.041 |
| EGFRrs2075108 | A | 1.15 (1.03–1.28) | 0.013 | 1.39 (1.11–1.73) | 0.0035 | 0.86 (0.68–1.08) | 0.20 | 0.89 (0.70–1.14) | 0.36 | 1.10 (1.01–1.20) | 0.025 | NS | 0.007 |
| TP73rs4648553 | A | 1.10 (1.02–1.18) | 0.011 | 1.21 (1.05–1.39) | 0.010 | 0.87 (0.75–1.01) | 0.075 | 1.11 (0.96–1.28) * | 0.17 | 1.08 (1.02–1.14) | 0.0039 | NS | 0.013 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sainz, J.; García-Verdejo, F.J.; Martínez-Bueno, M.; Kumar, A.; Sánchez-Maldonado, J.M.; Díez-Villanueva, A.; Vodičková, L.; Vymetálková, V.; Martin Sánchez, V.; Da Silva Filho, M.I.; et al. Polymorphisms within Autophagy-Related Genes Influence the Risk of Developing Colorectal Cancer: A Meta-Analysis of Four Large Cohorts. Cancers 2021, 13, 1258. https://doi.org/10.3390/cancers13061258
Sainz J, García-Verdejo FJ, Martínez-Bueno M, Kumar A, Sánchez-Maldonado JM, Díez-Villanueva A, Vodičková L, Vymetálková V, Martin Sánchez V, Da Silva Filho MI, et al. Polymorphisms within Autophagy-Related Genes Influence the Risk of Developing Colorectal Cancer: A Meta-Analysis of Four Large Cohorts. Cancers. 2021; 13(6):1258. https://doi.org/10.3390/cancers13061258
Chicago/Turabian StyleSainz, Juan, Francisco José García-Verdejo, Manuel Martínez-Bueno, Abhishek Kumar, José Manuel Sánchez-Maldonado, Anna Díez-Villanueva, Ludmila Vodičková, Veronika Vymetálková, Vicente Martin Sánchez, Miguel Inacio Da Silva Filho, and et al. 2021. "Polymorphisms within Autophagy-Related Genes Influence the Risk of Developing Colorectal Cancer: A Meta-Analysis of Four Large Cohorts" Cancers 13, no. 6: 1258. https://doi.org/10.3390/cancers13061258