
Functional Genetic Variants in ATG10 Are Associated with Acute Myeloid Leukemia

 , , , , ,
, , , , ,  and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. DNA Extraction, SNP Selection Criteria and Genotyping
2.3. Association Studies
2.4. mRNA Expression Analysis by qRT-PCR
2.5. Protein Expression and Immunoblot Analysis
2.6. Peripheral Blood Mononuclear Cells (PBMC) Isolation
2.7. Autophagy Flux Analysis
2.8. Statistical Analysis
3. Results
3.1. Demographic Characterization of the Cohorts
3.2. Linkage Analysis of ATG10 SNPs
3.3. Associations of Genetic Variants on ATG10 with AML
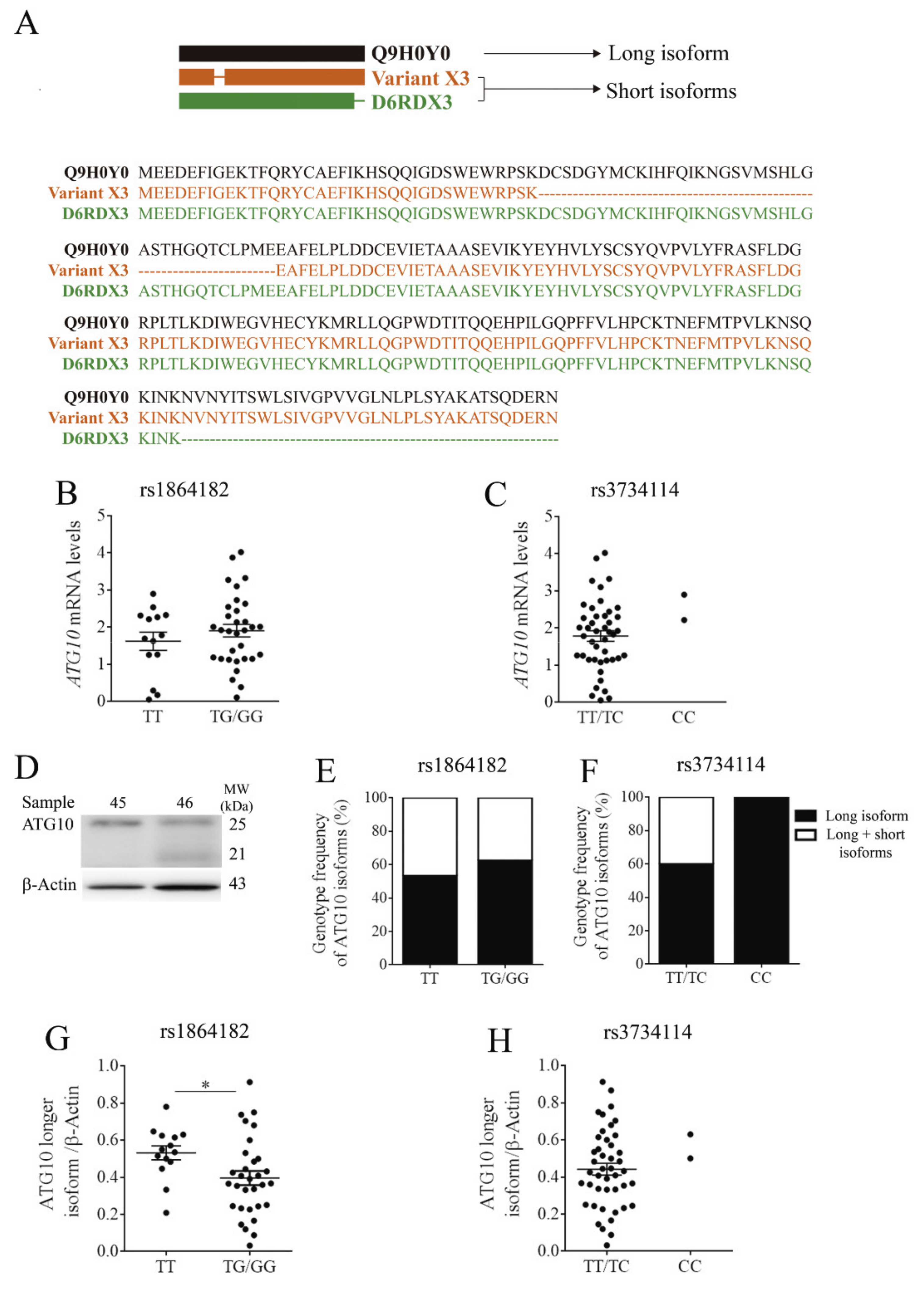
3.4. Impact of ATG10rs1864182 and ATG10rs3734114 in mRNA and Protein Levels
3.5. Functional Effects of ATG10rs1864182 and ATG10rs3734114 on Autophagy
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Estey, E.; Dohner, H. Acute myeloid leukaemia. Lancet 2006, 368, 1894–1907. [Google Scholar] [CrossRef]
- Klco, J.M.; Mullighan, C.G. Advances in germline predisposition to acute leukaemias and myeloid neoplasms. Nat. Rev. Cancer 2020. [Google Scholar] [CrossRef]
- Steensma, D.P. Clinical consequences of clonal hematopoiesis of indeterminate potential. Blood Adv. 2018, 2, 3404–3410. [Google Scholar] [CrossRef]
- Chen, D.P.; Chang, S.W.; Wang, P.N.; Hus, F.P.; Tseng, C.P. Association between single nucleotide polymorphisms within HLA region and disease relapse for patients with hematopoietic stem cell transplantation. Sci. Rep. 2019, 9, 13731. [Google Scholar] [CrossRef] [Green Version]
- Illmer, T.; Schuler, U.S.; Thiede, C.; Schwarz, U.I.; Kim, R.B.; Gotthard, S.; Freund, D.; Schakel, U.; Ehninger, G.; Schaich, M. MDR1 gene polymorphisms affect therapy outcome in acute myeloid leukemia patients. Cancer Res. 2002, 62, 4955–4962. [Google Scholar]
- Kim, D.H.; Park, J.Y.; Sohn, S.K.; Lee, N.Y.; Baek, J.H.; Jeon, S.B.; Kim, J.G.; Suh, J.S.; Do, Y.R.; Lee, K.B. Multidrug resistance-1 gene polymorphisms associated with treatment outcomes in de novo acute myeloid leukemia. Int. J. Cancer 2006, 118, 2195–2201. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.Y.; Shi, Z.Z.; Zhang, S.J.; Zhu, Y.M.; Gu, B.W.; Li, G.; Bai, X.T.; Gao, X.D.; Hu, J.; Jin, W.; et al. Association between single nucleotide polymorphisms in deoxycytidine kinase and treatment response among acute myeloid leukaemia patients. Pharmacogenetics 2004, 14, 759–768. [Google Scholar] [CrossRef] [PubMed]
- Xu, P.; Wang, X.; Xu, Y.; Chen, B.; Ouyang, J. The rs9909659G/A polymorphisms of the STAT3 gene provide prognostic information in acute myeloid leukemia. Transl. Cancer Res. 2016, 5, 448–457. [Google Scholar] [CrossRef]
- Damm, F.; Heuser, M.; Morgan, M.; Yun, H.; Grosshennig, A.; Gohring, G.; Schlegelberger, B.; Dohner, K.; Ottmann, O.; Lubbert, M.; et al. Single nucleotide polymorphism in the mutational hotspot of WT1 predicts a favorable outcome in patients with cytogenetically normal acute myeloid leukemia. J. Clin. Oncol. 2010, 28, 578–585. [Google Scholar] [CrossRef] [PubMed]
- AbdElMaksoud, S.S.; ElGamal, R.A.E.; Pessar, S.A.; Salem, D.D.E.; Abdelsamee, H.F.; Agamy, H.S. Prognostic implications of IDH1rs11554137 and IDH2R140Q SNPs mutations in cytogenetically normal acute myeloid leukemia. Egypt. J. Med. Hum. Genet. 2019, 20. [Google Scholar] [CrossRef] [Green Version]
- Ho, P.A.; Kopecky, K.J.; Alonzo, T.A.; Gerbing, R.B.; Miller, K.L.; Kuhn, J.; Zeng, R.; Ries, R.E.; Raimondi, S.C.; Hirsch, B.A.; et al. Prognostic implications of the IDH1 synonymous SNP rs11554137 in pediatric and adult AML: A report from the Children’s Oncology Group and SWOG. Blood 2011, 118, 4561–4566. [Google Scholar] [CrossRef]
- Castro, I.; Sampaio-Marques, B.; Ludovico, P. Targeting Metabolic Reprogramming in Acute Myeloid Leukemia. Cells 2019, 8, 967. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schnittger, S.; Kohl, T.M.; Leopold, N.; Schoch, C.; Wichmann, H.E.; Kern, W.; Lohse, P.; Hiddemann, W.; Haferlach, T.; Spiekermann, K. D324N single-nucleotide polymorphism in the FLT3 gene is associated with higher risk of myeloid leukemias. Genes Chromosomes Cancer 2006, 45, 332–337. [Google Scholar] [CrossRef] [PubMed]
- Cingeetham, A.; Vuree, S.; Dunna, N.R.; Gorre, M.; Nanchari, S.R.; Edathara, P.M.; Meka, P.; Annamaneni, S.; Digumarthi, R.; Sinha, S.; et al. Influence of BCL2-938C>A and BAX-248G>A promoter polymorphisms in the development of AML: Case-control study from South India. Tumour Biol. J. Int. Soc. Oncodev. Biol. Med. 2015, 36, 7967–7976. [Google Scholar] [CrossRef] [PubMed]
- Cingeetham, A.; Vuree, S.; Dunna, N.R.; Gorre, M.; Nanchari, S.R.; Edathara, P.M.; Mekkaw, P.; Annamaneni, S.; Digumarthi, R.R.; Sinha, S.; et al. Association of caspase9 promoter polymorphisms with the susceptibility of AML in south Indian subjects. Tumour Biol. J. Int. Soc. Oncodev. Biol. Med. 2014, 35, 8813–8822. [Google Scholar] [CrossRef]
- Mizushima, N. Autophagy: Process and function. Genes Dev. 2007, 21, 2861–2873. [Google Scholar] [CrossRef] [Green Version]
- Yun, C.W.; Lee, S.H. The Roles of Autophagy in Cancer. Int. J. Mol. Sci. 2018, 19, 3466. [Google Scholar] [CrossRef] [Green Version]
- Du, W.; Xu, A.; Huang, Y.; Cao, J.; Zhu, H.; Yang, B.; Shao, X.; He, Q.; Ying, M. The role of autophagy in targeted therapy for acute myeloid leukemia. Autophagy 2020, 1–15. [Google Scholar] [CrossRef]
- Chen, X.X.; Li, Z.P.; Zhu, J.H.; Xia, H.T.; Zhou, H. Systematic Analysis of Autophagy-Related Signature Uncovers Prognostic Predictor for Acute Myeloid Leukemia. DNA Cell Biol. 2020, 39, 1595–1605. [Google Scholar] [CrossRef]
- Phillips, A.R.; Suttangkakul, A.; Vierstra, R.D. The ATG12-conjugating enzyme ATG10 Is essential for autophagic vesicle formation in Arabidopsis thaliana. Genetics 2008, 178, 1339–1353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.C.; Zhang, M.Q.; Zhang, J.P. Opposite Effects of Two Human ATG10 Isoforms on Replication of a HCV Sub-genomic Replicon Are Mediated via Regulating Autophagy Flux in Zebrafish. Front. Cell Infect. Microbiol. 2018, 8, 109. [Google Scholar] [CrossRef]
- Zhao, Q.; Hu, Z.Y.; Zhang, J.P.; Jiang, J.D.; Ma, Y.Y.; Li, J.R.; Peng, Z.G.; Chen, J.H. Dual Roles of Two Isoforms of Autophagy-related Gene ATG10 in HCV-Subgenomic replicon Mediated Autophagy Flux and Innate Immunity. Sci. Rep. 2017, 7, 11250. [Google Scholar] [CrossRef]
- Jiang, P.; Mizushima, N. Autophagy and human diseases. Cell Res. 2014, 24, 69–79. [Google Scholar] [CrossRef] [Green Version]
- Xie, K.; Liang, C.; Li, Q.; Yan, C.; Wang, C.; Gu, Y.; Zhu, M.; Du, F.; Wang, H.; Dai, J.; et al. Role of ATG10 expression quantitative trait loci in non-small cell lung cancer survival. Int. J. Cancer 2016, 139, 1564–1573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salvatori, B.; Iosue, I.; Djodji Damas, N.; Mangiavacchi, A.; Chiaretti, S.; Messina, M.; Padula, F.; Guarini, A.; Bozzoni, I.; Fazi, F.; et al. Critical Role of c-Myc in Acute Myeloid Leukemia Involving Direct Regulation of miR-26a and Histone Methyltransferase EZH2. Genes Cancer 2011, 2, 585–592. [Google Scholar] [CrossRef] [Green Version]
- Zhu, F.; Huang, R.; Li, J.; Liao, X.; Huang, Y.; Lai, Y. Identification of Key Genes and Pathways Associated with RUNX1 Mutations in Acute Myeloid Leukemia Using Bioinformatics Analysis. Med. Sci. Monit. 2018, 24, 7100–7108. [Google Scholar] [CrossRef] [PubMed]
- Buffen, K.; Oosting, M.; Quintin, J.; Ng, A.; Kleinnijenhuis, J.; Kumar, V.; van de Vosse, E.; Wijmenga, C.; van Crevel, R.; Oosterwijk, E.; et al. Autophagy controls BCG-induced trained immunity and the response to intravesical BCG therapy for bladder cancer. PLoS Pathog. 2014, 10, e1004485. [Google Scholar] [CrossRef]
- Li, Q.X.; Zhou, X.; Huang, T.T.; Tang, Y.; Liu, B.; Peng, P.; Sun, L.; Wang, Y.H.; Yuan, X.L. The Thr300Ala variant of ATG16L1 is associated with decreased risk of brain metastasis in patients with non-small cell lung cancer. Autophagy 2017, 13, 1053–1063. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Plantinga, T.S.; van de Vosse, E.; Huijbers, A.; Netea, M.G.; Joosten, L.A.; Smit, J.W.; Netea-Maier, R.T. Role of genetic variants of autophagy genes in susceptibility for non-medullary thyroid cancer and patients outcome. PLoS ONE 2014, 9, e94086. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, Y.; Han, H.; Jin, Y.; Zhou, X.; Yi, M.; Tang, Y.; Li, Q. Implications of the autophagy core gene variations on brain metastasis risk in non-small cell lung cancer treated with EGFR-TKI. Oncol. Transl. Med. 2020, 5, 185–192. [Google Scholar]
- Sanchez-Maldonado, J.M.; Campa, D.; Springer, J.; Badiola, J.; Niazi, Y.; Moniz-Diez, A.; Hernandez-Mohedo, F.; Gonzalez-Sierra, P.; Ter Horst, R.; Macauda, A.; et al. Host immune genetic variations influence the risk of developing acute myeloid leukaemia: Results from the NuCLEAR consortium. Blood Cancer J. 2020, 10, 75. [Google Scholar] [CrossRef]
- Clarke, G.M.; Anderson, C.A.; Pettersson, F.H.; Cardon, L.R.; Morris, A.P.; Zondervan, K.T. Basic statistical analysis in genetic case-control studies. Nat. Protoc. 2011, 6, 121–133. [Google Scholar] [CrossRef] [Green Version]
- Bustin, S.A.; Benes, V.; Garson, J.A.; Hellemans, J.; Huggett, J.; Kubista, M.; Mueller, R.; Nolan, T.; Pfaffl, M.W.; Shipley, G.L.; et al. The MIQE guidelines: Minimum information for publication of quantitative real-time PCR experiments. Clin. Chem. 2009, 55, 611–622. [Google Scholar] [CrossRef] [Green Version]
- Klionsky, D.J.; Abdelmohsen, K.; Abe, A.; Abedin, M.J.; Abeliovich, H.; Acevedo Arozena, A.; Adachi, H.; Adams, C.M.; Adams, P.D.; Adeli, K.; et al. Guidelines for the use and interpretation of assays for monitoring autophagy (3rd edition). Autophagy 2016, 12, 1–222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Juliusson, G.; Antunovic, P.; Derolf, A.; Lehmann, S.; Mollgard, L.; Stockelberg, D.; Tidefelt, U.; Wahlin, A.; Hoglund, M. Age and acute myeloid leukemia: Real world data on decision to treat and outcomes from the Swedish Acute Leukemia Registry. Blood 2009, 113, 4179–4187. [Google Scholar] [CrossRef] [Green Version]
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2016. Ca A Cancer J. Clin. 2016, 66, 7–30. [Google Scholar] [CrossRef] [Green Version]
- De-Morgan, A.; Meggendorfer, M.; Haferlach, C.; Shlush, L. Male predominance in AML is associated with specific preleukemic mutations. Leukemia 2020. [Google Scholar] [CrossRef] [PubMed]
- Flanagan, M.D.; Whitehall, S.K.; Morgan, B.A. An Atg10-like E2 enzyme is essential for cell cycle progression but not autophagy in Schizosaccharomyces pombe. Cell Cycle 2013, 12, 271–277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levine, B.; Packer, M.; Codogno, P. Development of autophagy inducers in clinical medicine. J. Clin. Investig. 2015, 125, 14–24. [Google Scholar] [CrossRef] [Green Version]
- Liu, W.J.; Ye, L.; Huang, W.F.; Guo, L.J.; Xu, Z.G.; Wu, H.L.; Yang, C.; Liu, H.F. p62 links the autophagy pathway and the ubiqutin-proteasome system upon ubiquitinated protein degradation. Cell. Mol. Biol. Lett. 2016, 21, 29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin-Maestro, P.; Gargini, R.; A Sproul, A.; Garcia, E.; Anton, L.C.; Noggle, S.; Arancio, O.; Avila, J.; Garcia-Escudero, V. Mitophagy Failure in Fibroblasts and iPSC-Derived Neurons of Alzheimer’s Disease-Associated Presenilin 1 Mutation. Front. Mol. Neurosci. 2017, 10, 291. [Google Scholar] [CrossRef] [Green Version]
- Hara, T.; Nakamura, K.; Matsui, M.; Yamamoto, A.; Nakahara, Y.; Suzuki-Migishima, R.; Yokoyama, M.; Mishima, K.; Saito, I.; Okano, H.; et al. Suppression of basal autophagy in neural cells causes neurodegenerative disease in mice. Nature 2006, 441, 885–889. [Google Scholar] [CrossRef]
- Chen, R.H.; Chen, Y.H.; Huang, T.Y. Ubiquitin-mediated regulation of autophagy. J. Biomed. Sci. 2019, 26, 80. [Google Scholar] [CrossRef]
- Tan, J.M.M.; Wong, E.S.P.; Dawson, V.L.; Dawson, T.; Lim, K.-L. Lysine 63-linked polyubiquitin potentially partners with p62 to promote the clearance of protein inclusions by autophagy. Autophagy 2008, 4, 251–253. [Google Scholar] [CrossRef] [Green Version]
- Erichsen, H.C.; Chanock, S.J. SNPs in cancer research and treatment. Br. J. Cancer 2004, 90, 747–751. [Google Scholar] [CrossRef] [Green Version]
- Shastry, B.S. SNPs in disease gene mapping, medicinal drug development and evolution. J. Hum. Genet. 2007, 52, 871–880. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qin, Z.; Xue, J.; He, Y.; Ma, H.; Jin, G.; Chen, J.; Hu, Z.; Liu, X.; Shen, H. Potentially functional polymorphisms in ATG10 are associated with risk of breast cancer in a Chinese population. Gene 2013, 527, 491–495. [Google Scholar] [CrossRef] [PubMed]
- White, K.A.; Luo, L.; Thompson, T.A.; Torres, S.; Hu, C.A.; Thomas, N.E.; Lilyquist, J.; Anton-Culver, H.; Gruber, S.B.; From, L.; et al. Variants in autophagy-related genes and clinical characteristics in melanoma: A population-based study. Cancer Med. 2016, 5, 3336–3345. [Google Scholar] [CrossRef] [Green Version]
- Yuan, J.; Zhang, N.; Yin, L.; Zhu, H.; Zhang, L.; Zhou, L.; Yang, M. Clinical Implications of the Autophagy Core Gene Variations in Advanced Lung Adenocarcinoma Treated with Gefitinib. Sci. Rep. 2017, 7, 17814. [Google Scholar] [CrossRef] [Green Version]
- Fernandez-Mateos, J.; Seijas-Tamayo, R.; Klain, J.C.A.; Borgonon, M.P.; Perez-Ruiz, E.; Mesia, R.; Del Barco, E.; Coloma, C.S.; Dominguez, A.R.; Daroqui, J.C.; et al. Analysis of autophagy gene polymorphisms in Spanish patients with head and neck squamous cell carcinoma. Sci. Rep. 2017, 7, 6887. [Google Scholar] [CrossRef] [Green Version]
- Consortium, G.T. The GTEx Consortium atlas of genetic regulatory effects across human tissues. Science 2020, 369, 1318–1330. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.P.; Niu, Y.N.; Yuan, N.; Zhang, A.H.; Chao, D.; Xu, Q.P.; Wang, L.J.; Zhang, X.G.; Zhao, W.L.; Zhao, Y.; et al. Role of autophagy in acute myeloid leukemia therapy. Chin. J. Cancer 2013, 32, 130–135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Evangelisti, C.; Evangelisti, C.; Chiarini, F.; Lonetti, A.; Buontempo, F.; Neri, L.M.; McCubrey, J.A.; Martelli, A.M. Autophagy in acute leukemias: A double-edged sword with important therapeutic implications. Biochim. Biophys. Acta 2015, 1853, 14–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amaravadi, R.K.; Kimmelman, A.C.; Debnath, J. Targeting Autophagy in Cancer: Recent Advances and Future Directions. Cancer Discov. 2019, 9, 1167–1181. [Google Scholar] [CrossRef] [Green Version]
- Fernandes, A.; Azevedo, M.M.; Pereira, O.; Sampaio-Marques, B.; Paiva, A.; Correia-Neves, M.; Castro, I.; Ludovico, P. Proteolytic systems and AMP-activated protein kinase are critical targets of acute myeloid leukemia therapeutic approaches. Oncotarget 2015, 6, 31428–31440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Callaway, E. ‘It will change everything’: DeepMind’s AI makes gigantic leap in solving protein structures. Nature 2020, 588, 203–204. [Google Scholar] [CrossRef]
- Galluzzi, L.; Green, D.R. Autophagy-Independent Functions of the Autophagy Machinery. Cell 2019, 177, 1682–1699. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Cohort 1—Spanish Multicenter | ||
|---|---|---|
| Healthy Donors | 356 | |
| Gender | 166 male | |
| 187 female | ||
| 3 NA | ||
| Age | 56 ± 6 years mean age | |
| Acute Myeloid Leukemia | 309 | |
| Gender | 167 male | |
| 133 female | ||
| 9 NA | ||
| Age | 58 ± 17 years mean age | |
| Cohort 2—Portuguese Donors | ||
| Healthy Donors | 46 | |
| Gender | 12 male | |
| 34 female | ||
| Age | 39 ± 14 years mean age | |
| Genotyped SNPs | rs1864182 | rs1864183 | rs3734114 |
|---|---|---|---|
| Chromosome | 5 | 5 | 5 |
| Chromosome Position | 82253421 | 82253397 | 82058570 |
| Major Allele | T | C | T |
| Base change | T > G | C > T | T > C |
| Minor Allele Frequency (MAF) in the controls | 0.508 | 0.429 | 0.202 |
| p value for HWE test in our controls | 0.821 | 0.302 | 0.365 |
| SNPs | Genotypes | Donors No. (%) | χ2 (pad) | LR Dominant | LR Recessive | |
|---|---|---|---|---|---|---|
| Control | AML | OR (CI) | OR (CI) | |||
| rs1864182 | TT | 77 (25) | 87 (35) | 0.174 | 0.58 (0.42–0.80) p = 0.001; pad = 0.006 | 0.70 (0.47–1.04) p = 0.077; pad = 0.462 |
| TG | 154 (49) | 110 (44) | ||||
| GG | 81 (26) | 53 (21) | ||||
| rs1864183 | CC | 109 (34) | 77 (29) | 1.506 | 1.29 (0.94–1.78) p = 0.113; pad = 0.678 | 1.31 (0.88–1.94) p = 0.180; pad = 1.080 |
| CT | 146 (46) | 126 (47) | ||||
| TT | 63 (20) | 65 (24) | ||||
| rs3734114 | TT | 204 (66) | 168 (68) | 0.012 | 0.86 (0.61–1.22) p = 0.414; pad = 2.484 | 2.70 (1.36–5.34) p = 0.004; pad = 0.024 |
| TC | 92 (30) | 53 (21) | ||||
| CC | 14 (4) | 28 (11) | ||||
| SNP | Genotype | Counts (%) |
|---|---|---|
| rs1864182 | TT | 14 (30.4) |
| TG | 18 (39.1) | |
| GG | 14 (30.4) | |
| rs374114 | TT | 26 (56.5) |
| TC | 18 (39.1) | |
| CC | 2 (4.3) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Castro, I.; Sampaio-Marques, B.; C. Areias, A.; Sousa, H.; Fernandes, Â.; Sanchez-Maldonado, J.M.; Cunha, C.; Carvalho, A.; Sainz, J.; Ludovico, P. Functional Genetic Variants in ATG10 Are Associated with Acute Myeloid Leukemia. Cancers 2021, 13, 1344. https://doi.org/10.3390/cancers13061344
Castro I, Sampaio-Marques B, C. Areias A, Sousa H, Fernandes Â, Sanchez-Maldonado JM, Cunha C, Carvalho A, Sainz J, Ludovico P. Functional Genetic Variants in ATG10 Are Associated with Acute Myeloid Leukemia. Cancers. 2021; 13(6):1344. https://doi.org/10.3390/cancers13061344
Chicago/Turabian StyleCastro, Isabel, Belém Sampaio-Marques, Anabela C. Areias, Hugo Sousa, Ângela Fernandes, José Manuel Sanchez-Maldonado, Cristina Cunha, Agostinho Carvalho, Juan Sainz, and Paula Ludovico. 2021. "Functional Genetic Variants in ATG10 Are Associated with Acute Myeloid Leukemia" Cancers 13, no. 6: 1344. https://doi.org/10.3390/cancers13061344