
BOLD Coupling between Lesioned and Healthy Brain Is Associated with Glioma Patients’ Recovery

 , , ,
, , ,  , , , and
, , , and 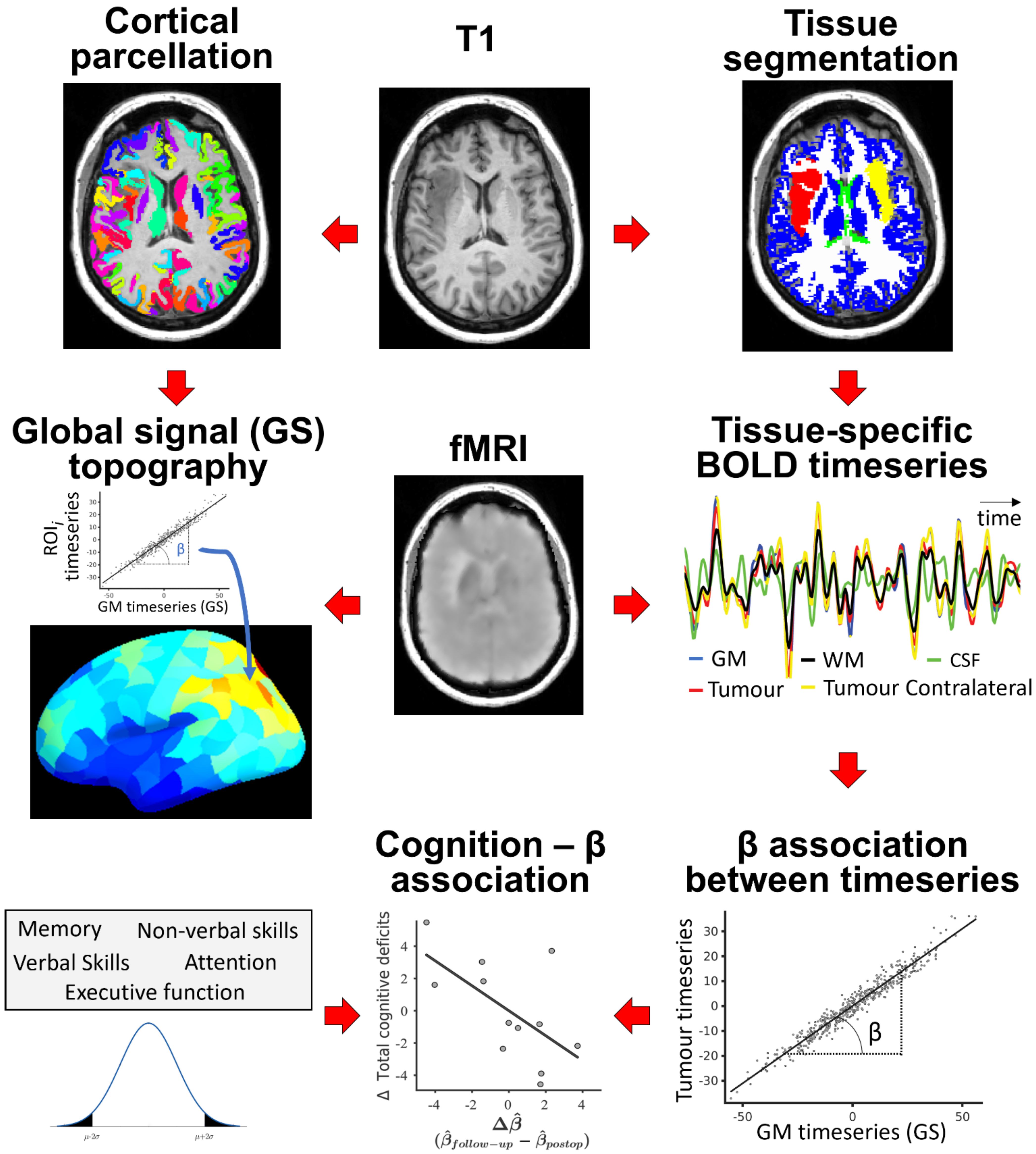{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample
2.2. MRI Data Acquisition and Pre-Processing
2.3. Lesion Masking, Image Co-Registration, Parcellation and Time-Series Extraction
2.4. Neuropsychological Assessment
2.5. BOLD Signal Extraction from Tissue Compartments and Analysis
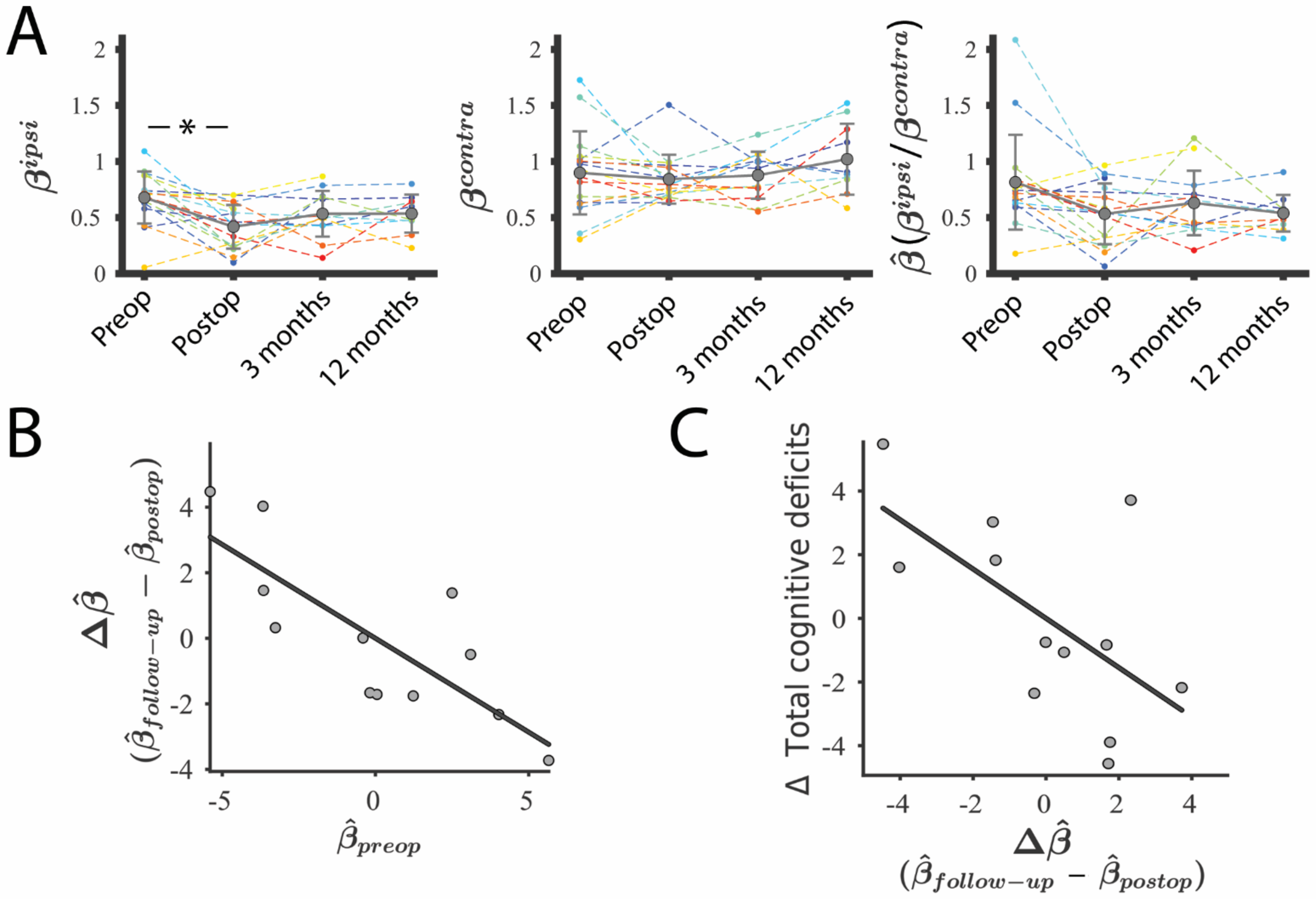
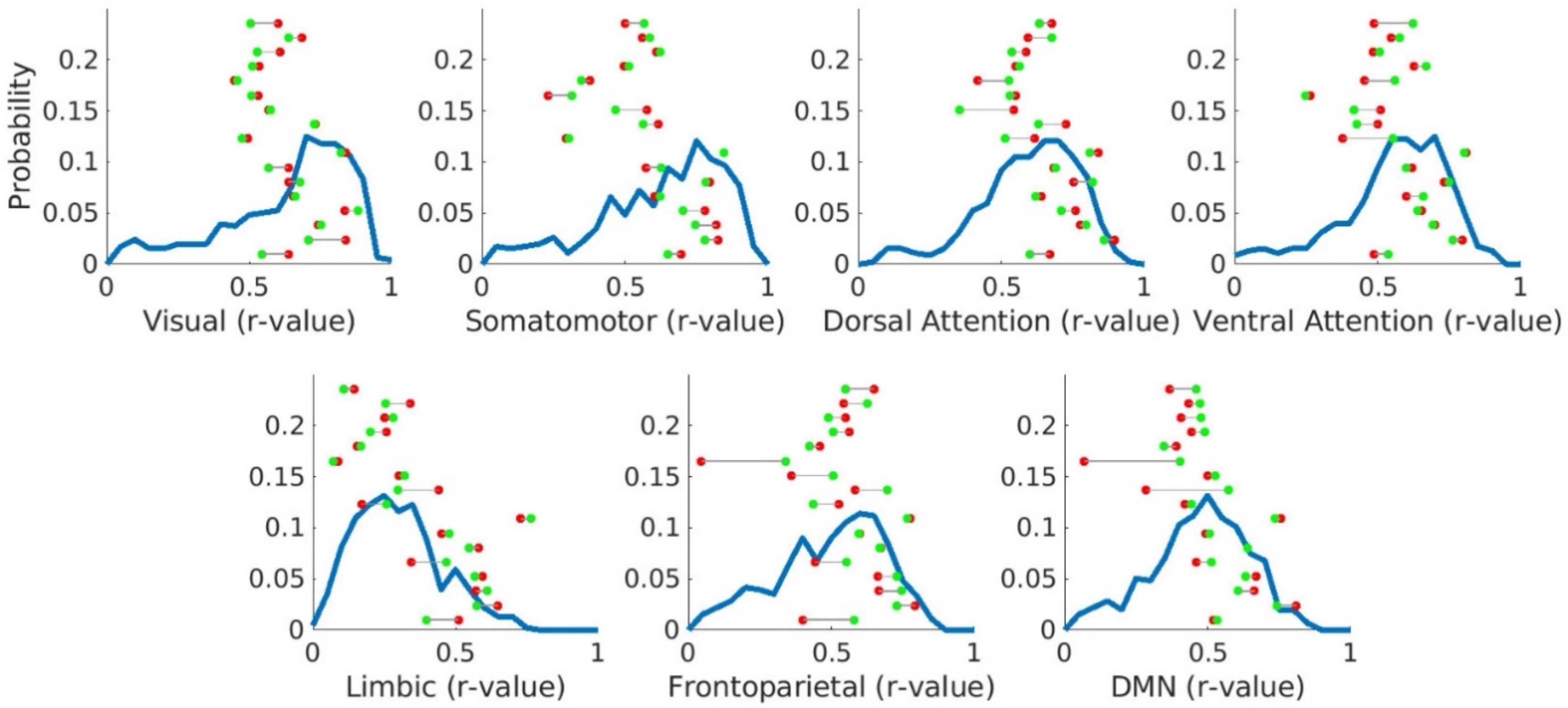
3. Results
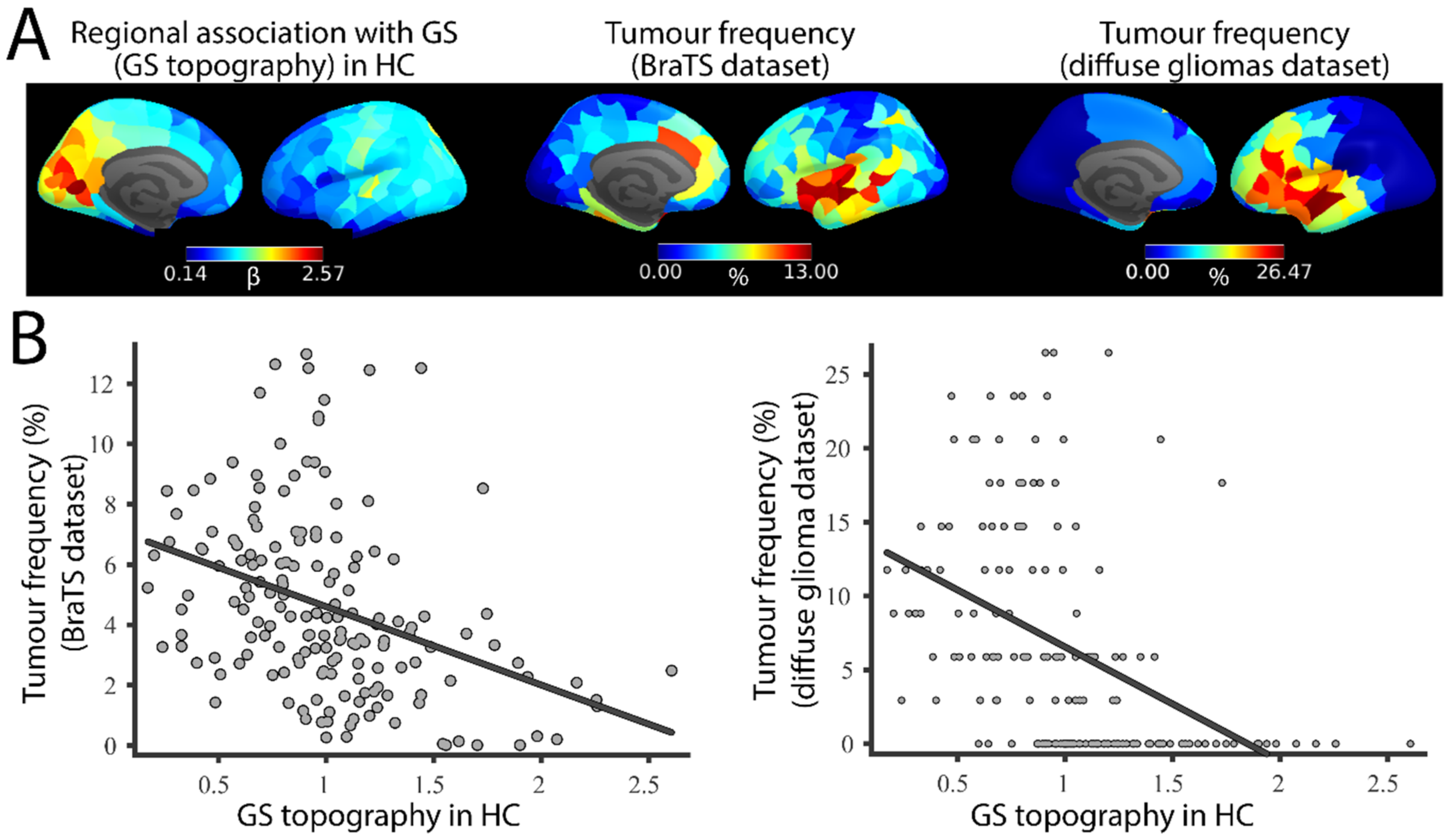
3.1. GS Topography Is Associated with Tumour Occurrence
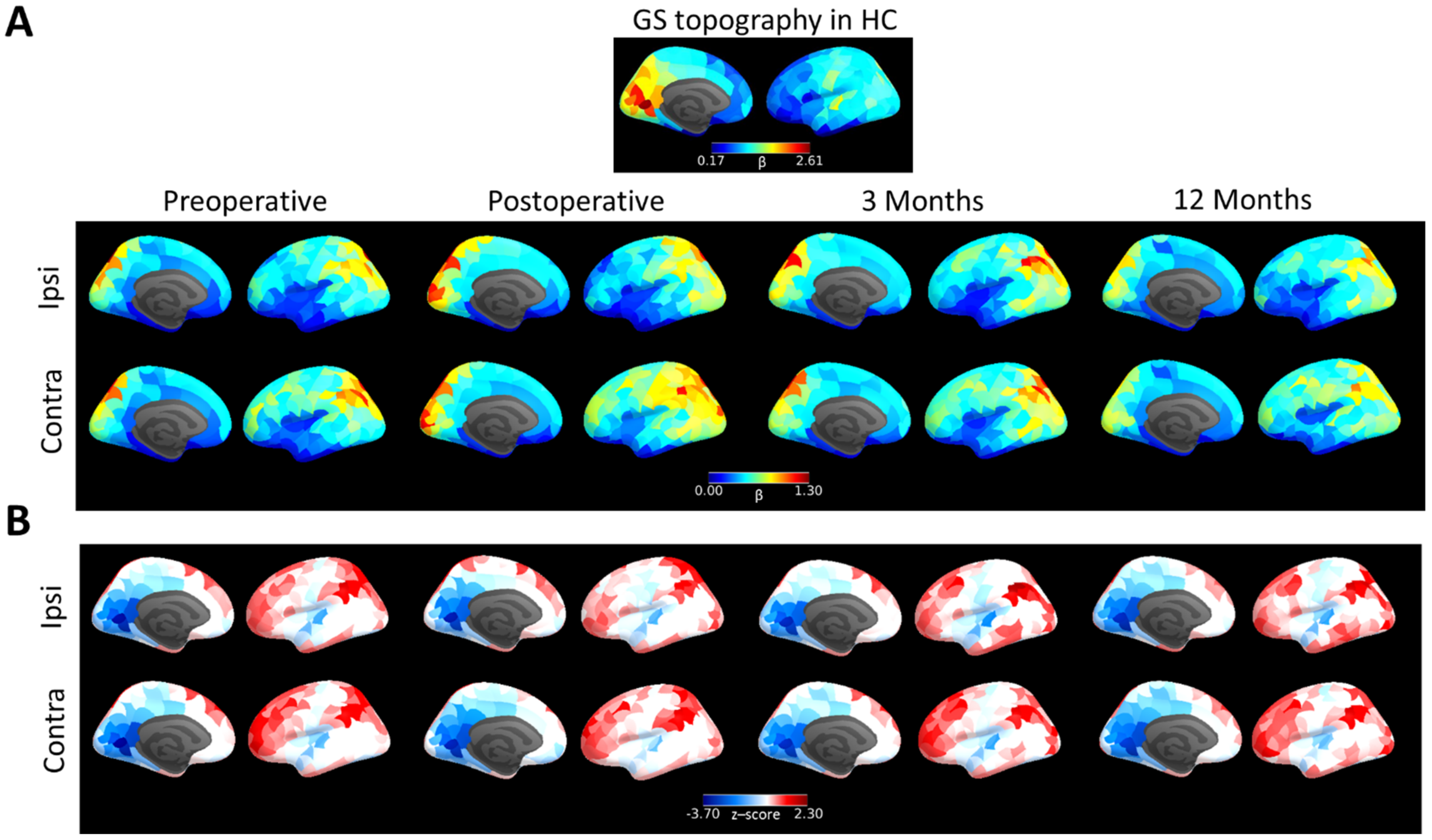
3.2. Brain Tumour Patients Have a Long-Term Alteration of GS Topography
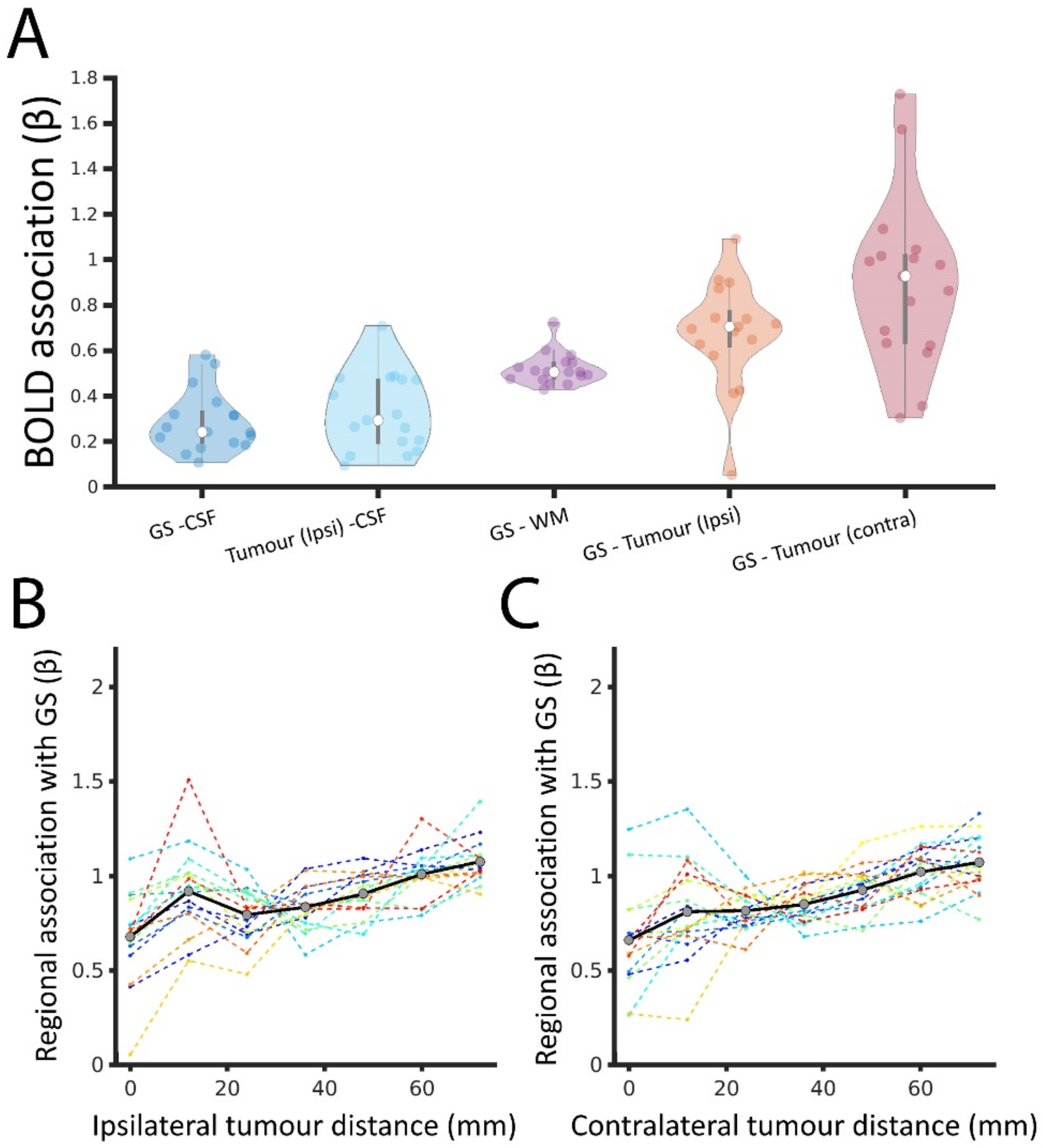
3.3. Tumour BOLD Time Series Is Coupled with GS
3.4. Lesion–GS Coupling Is Preserved during Recovery and Is Associated with Cognition
3.5. Canonical Resting-State Networks Are Preserved in Brain Tumour Patients
4. Discussion
Limitations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Brown, P.D.; Maurer, M.J.; Rummans, T.A.; Pollock, B.E.; Ballman, K.; Sloan, J.A.; Boeve, B.F.; Arusell, R.M.; Clark, M.M.; Buckner, J.C. A Prospective Study of Quality of Life in Adults with Newly Diagnosed High-grade Gliomas: The Impact of the Extent of Resection on Quality of Life and Survival. Neurosurgery 2005, 57, 495–504. [Google Scholar] [CrossRef]
- Taphoorn, M.J.; Klein, M. Cognitive deficits in adult patients with brain tumours. Lancet Neurol. 2004, 3, 159–168. [Google Scholar] [CrossRef]
- Scheibel, R.; Meyers, C.; Levin, V. Cognitive dysfunction following surgery for intracerebral glioma: Influence of histopathology, lesion location, and treatment. J. Neuro-Oncol. 1996, 30, 61–69. [Google Scholar] [CrossRef] [PubMed]
- Dwan, T.M.; Ownsworth, T.; Chambers, S.; Walker, D.G.; Shum, D.H.K. Neuropsychological Assessment of Individuals with Brain Tumor: Comparison of Approaches Used in the Classification of Impairment. Front. Oncol. 2015, 5, 56. [Google Scholar] [CrossRef] [Green Version]
- Mauer, M.; Stupp, R.; Taphoorn, M.J.B.; Coens, C.; Osoba, D.; Marosi, C.; Wong, R.; de Witte, O.; Cairncross, J.G.; Efficace, F.; et al. The prognostic value of health-related quality-of-life data in predicting survival in glioblastoma cancer patients: Results from an international randomised phase III EORTC Brain Tumour and Radiation Oncology Groups, and NCIC Clinical Trials Group study. Br. J. Cancer 2007, 97, 302–307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.-G.; Ogawa, S. Biophysical and Physiological Origins of Blood Oxygenation Level-Dependent fMRI Signals. Br. J. Pharmacol. 2012, 32, 1188–1206. [Google Scholar] [CrossRef] [PubMed]
- Zhu, D.C.; Tarumi, T.; Khan, M.A.; Zhang, R. Vascular Coupling in Resting-State FMRI: Evidence from Multiple Modalities. Br. J. Pharmacol. 2015, 35, 1910–1920. [Google Scholar] [CrossRef] [Green Version]
- Power, J.D.; Plitt, M.; Laumann, T.O.; Martin, A. Sources and implications of whole-brain fMRI signals in humans. NeuroImage 2016, 146, 609–625. [Google Scholar] [CrossRef] [Green Version]
- Chang, C.; Glover, G.H. Relationship between respiration, end-tidal CO2, and BOLD signals in resting-state fMRI. NeuroImage 2009, 47, 1381–1393. [Google Scholar] [CrossRef] [Green Version]
- Liu, T.T.; Nalci, A.; Falahpour, M. The global signal in fMRI: Nuisance or Information? Neuroimage 2017, 150, 213–229. [Google Scholar] [CrossRef] [Green Version]
- Scholvinck, M.L.; Maier, A.; Ye, F.Q.; Duyn, J.H.; Leopold, D.A. Neural basis of global resting-state fMRI activity. Proc. Natl. Acad. Sci. USA 2010, 107, 10238–10243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wen, H.; Liu, Z. Broadband Electrophysiological Dynamics Contribute to Global Resting-State fMRI Signal. J. Neurosci. 2016, 36, 6030–6040. [Google Scholar] [CrossRef] [Green Version]
- Wong, C.W.; Olafsson, V.; Tal, O.; Liu, T.T. The amplitude of the resting-state fMRI global signal is related to EEG vigilance measures. NeuroImage 2013, 83, 983–990. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wong, C.W.; Olafsson, V.; Tal, O.; Liu, T.T. Anti-correlated networks, global signal regression, and the effects of caffeine in resting-state functional MRI. NeuroImage 2012, 63, 356–364. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Bolt, T.; Bzdok, D.; Nomi, J.S.; Yeo, B.T.T.; Spreng, R.N.; Uddin, L.Q. Topography and behavioral relevance of the global signal in the human brain. Sci. Rep. 2019, 9, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Stevens, M.T.R.; Clarke, D.B.; Stroink, G.; Beyea, S.D.; D’Arcy, R.C. Improving fMRI reliability in presurgical mapping for brain tumours. J. Neurol. Neurosurg. Psychiatry 2015, 87, 267–274. [Google Scholar] [CrossRef]
- Tyndall, A.J.; Reinhardt, J.; Tronnier, V.; Mariani, L.; Stippich, C. Presurgical motor, somatosensory and language fMRI: Technical feasibility and limitations in 491 patients over 13 years. Eur. Radiol. 2016, 27, 267–278. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, S.; Sair, H.I.; Gujar, S.; Pillai, J.J. Language Mapping with fMRI: Current Standards and Reproducibility. Top Magn. Reason. Imaging 2019, 28, 225–233. [Google Scholar] [CrossRef]
- Chamberland, M.; Bernier, M.; Fortin, D.; Whittingstall, K.; Descoteaux, M. 3D interactive tractography-informed resting-state fMRI connectivity. Front. Neurosci. 2015, 9, 257. [Google Scholar] [CrossRef] [Green Version]
- Huang, H.; Lu, J.; Wu, J.; Ding, Z.; Chen, S.; Duan, L.; Cui, J.; Chen, F.; Kang, D.; Qi, L.; et al. Tumor Tissue Detection using Blood-Oxygen-Level-Dependent Functional MRI based on Independent Component Analysis. Sci. Rep. 2018, 8, 1223. [Google Scholar] [CrossRef]
- Hart, M.; Romero-Garcia, R.; Price, S.J.; Suckling, J. Global Effects of Focal Brain Tumors on Functional Complexity and Network Robustness: A Prospective Cohort Study. Neurosurgery 2018, 84, 1201–1213. [Google Scholar] [CrossRef] [Green Version]
- Schouwenaars, I.T.; de Dreu, M.J.; Rutten, G.-J.M.; Ramsey, N.F.; Jansma, J.M. A functional MRI study of presurgical cognitive deficits in glioma patients. Neuro-Oncol. Pract. 2020, 8, 81–90. [Google Scholar] [CrossRef] [PubMed]
- Yetkin, F.; Mendelsohn, D. Hypoxia imaging in brain tumors. Neuroimaging Clin. N. Am. 2002, 12, 537–552. [Google Scholar] [CrossRef]
- Orukari, I.; Siegel, J.S.; Warrington, N.M.; Baxter, G.; Bauer, A.Q.; Shimony, J.S.; Rubin, J.B.; Culver, J.P. Altered hemodynamics contribute to local but not remote functional connectivity disruption due to glioma growth. Br. J. Pharmacol. 2018, 40, 100–115. [Google Scholar] [CrossRef]
- Daniel, A.G.S.; Park, K.Y.; Roland, J.; Dierker, D.; Gross, J.; Humphries, J.B.; Hacker, C.D.; Snyder, A.Z.; Shimony, J.S.; Leuthardt, E.C. Functional connectivity within glioblastoma impacts overall survival. Neuro-Oncology 2020, 23, 412–421. [Google Scholar] [CrossRef] [PubMed]
- Erez, Y.; Assem, M.; Coelho, P.; Romero-Garcia, R.; Owen, M.; McDonald, A.; Woodberry, E.; Morris, R.C.; Price, S.J. Intraoperative mapping of executive function using electrocorticography for patients with low-grade gliomas. Acta. Neurochir. 2020, 163, 1299–1309. [Google Scholar] [CrossRef]
- Shafto, M.A.; Can, C.; Tyler, L.K.; Dixon, M.; Taylor, J.R.; Rowe, J.B.; Cusack, R.; Calder, A.J.; Marslen-Wilson, W.D.; Duncan, J.; et al. The Cambridge Centre for Ageing and Neuroscience (Cam-CAN) study protocol: A cross-sectional, lifespan, multidisciplinary examination of healthy cognitive ageing. BMC Neurol. 2014, 14, 1–25. [Google Scholar] [CrossRef] [Green Version]
- Bethlehem, R.A.; Paquola, C.; Seidlitz, J.; Ronan, L.; Bernhardt, B.; Cam-CAN Consortium; Tsvetanov, K.A. Dispersion of functional gradients across the adult lifespan. NeuroImage 2020, 222, 117299. [Google Scholar] [CrossRef]
- Mandal, A.S.; Romero-Garcia, R.; Hart, M.G.; Suckling, J. Genetic, cellular, and connectomic characterization of the brain regions commonly plagued by glioma. Brain 2020, 143, 3294–3307. [Google Scholar] [CrossRef] [PubMed]
- Salimi-Khorshidi, G.; Douaud, G.; Beckmann, C.F.; Glasser, M.F.; Griffanti, L.; Smith, S. Automatic denoising of functional MRI data: Combining independent component analysis and hierarchical fusion of classifiers. NeuroImage 2014, 90, 449–468. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romero-Garcia, R.; Erez, Y.; Oliver, G.; Owen, M.; Merali, S.; Poologaindran, A.; Morris, R.C.; Price, S.J.; Santarius, T.; Suckling, J.; et al. Practical Application of Networks in Neurosurgery: Combined 3-Dimensional Printing, Neuronavigation, and Preoperative Surgical Planning. World Neurosurg. 2020, 137, e126–e137. [Google Scholar] [CrossRef]
- Bullmore, E.; Fadili, J.; Maxim, V.; Şendur, L.; Whitcher, B.; Suckling, J.; Brammer, M.; Breakspear, M. Wavelets and functional magnetic resonance imaging of the human brain. NeuroImage 2004, 23, S234–S249. [Google Scholar] [CrossRef] [Green Version]
- Lommers, E.; Simon, J.; Reuter, G.; Delrue, G.; Dive, D.; Degueldre, C.; Balteau, E.; Phillips, C.; Maquet, P. Multiparameter MRI quantification of microstructural tissue alterations in multiple sclerosis. NeuroImage Clin. 2019, 23, 101879. [Google Scholar] [CrossRef]
- Romero-Garcia, R.; Atienza, M.; Clemmensen, L.; Cantero, J.L. Effects of network resolution on topological properties of human neocortex. NeuroImage 2012, 59, 3522–3532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yeo, B.T.; Krienen, F.M.; Sepulcre, J.; Sabuncu, M.; Lashkari, D.; Hollinshead, M.; Roffman, J.; Smoller, J.W.; Zöllei, L.; Polimeni, J.; et al. The organization of the human cerebral cortex estimated by intrinsic functional connectivity. J. Neurophysiol. 2011, 106, 1125–1165. [Google Scholar] [CrossRef] [PubMed]
- Meyers, J.E.; Zellinger, M.M.; Kockler, T.; Wagner, M.; Miller, R.M. A Validated Seven-Subtest Short Form for the WAIS-IV. Appl. Neuropsychol. Adult 2013, 20, 249–256. [Google Scholar] [CrossRef] [PubMed]
- Quental, N.B.M.; Brucki, S.; Bueno, O.F.A. Visuospatial Function in Early Alzheimer’s Disease—The Use of the Visual Object and Space Perception (VOSP) Battery. PLoS ONE 2013, 8, e68398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vlaar, A.M.M.; Wade, D. The Adult Memory and Information Processing Battery (AMIPB) test of information-processing speed: A study of its reliability and feasibility in patients with multiple sclerosis. Clin. Rehabil. 2003, 17, 386–393. [Google Scholar] [CrossRef]
- Lawson, R.A.; Yarnall, A.J.; Duncan, G.W.; Khoo, T.K.; Breen, D.P.; Barker, R.A.; Collerton, D.; Taylor, J.-P.; Burn, D.J. Severity of mild cognitive impairment in early Parkinson’s disease contributes to poorer quality of life. Park. Relat. Disord. 2014, 20, 1071–1075. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schinka, J.A.; Loewenstein, D.A.; Raj, A.; Schoenberg, M.; Banko, J.L.; Potter, H.; Duara, R. Defining Mild Cognitive Impairment: Impact of Varying Decision Criteria on Neuropsychological Diagnostic Frequencies and Correlates. Am. J. Geriatr. Psychiatry 2010, 18, 684–691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Owen, M.; Romero-Garcia, R.; McDonald, A.; Woodberry, E.; Assem, M.; Coelho, P.; Morris, R.C.; Price, S.J.; Santarius, T.; Suckling, J.; et al. Assessment of neuropsychological function during early treatment of diffuse glioma. medRxiv 2020. [Google Scholar] [CrossRef]
- Váša, F.; Romero-Garcia, R.; Kitzbichler, M.G.; Seidlitz, J.; Whitaker, K.J.; Vaghi, M.M.; Kundu, P.; Patel, A.X.; Fonagy, P.; Dolan, R.J.; et al. Conservative and disruptive modes of adolescent change in human brain functional connectivity. Proc. Natl. Acad. Sci. USA 2020, 117, 3248–3253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gordon, E.M.; Laumann, T.O.; Adeyemo, B.; Huckins, J.; Kelley, W.M.; Petersen, S.E. Generation and Evaluation of a Cortical Area Parcellation from Resting-State Correlations. Cereb. Cortex 2014, 26, 288–303. [Google Scholar] [CrossRef]
- Fox, M.E.; King, T.Z. Functional Connectivity in Adult Brain Tumor Patients: A Systematic Review. Brain Connect. 2018, 8, 381–397. [Google Scholar] [CrossRef] [PubMed]
- Manan, H.A.; Franz, E.A.; Yahya, N. Functional connectivity changes in patients with brain tumours—A systematic review on resting state-fMRI. Neurol. Psychiatry Brain Res. 2020, 36, 73–82. [Google Scholar] [CrossRef]
- Krishna, S.; Choudhury, A.; Seo, K.; Ni, L.; Kakaizada, S.; Lee, A.; Aabedi, A.; Cao, C.; Sudharshan, R.; Egladyous, A.; et al. Glioblastoma remodeling of neural circuits in the human brain decreases survival. bioRxiv 2021. [Google Scholar] [CrossRef]
- Shulman, R.G.; Rothman, D.L. Interpreting functional imaging studies in terms of neurotransmitter cycling. Proc. Natl. Acad. Sci. USA 1998, 95, 11993–11998. [Google Scholar] [CrossRef] [Green Version]
- Turchi, J.; Chang, C.; Ye, F.Q.; Russ, B.; Yu, D.K.; Cortes, C.R.; Monosov, I.E.; Duyn, J.H.; Leopold, D.A. The Basal Forebrain Regulates Global Resting-State fMRI Fluctuations. Neuron 2018, 97, 940–952.e4. [Google Scholar] [CrossRef] [Green Version]
- Murphy, K.; Fox, M.D. Towards a consensus regarding global signal regression for resting state functional connectivity MRI. NeuroImage 2016, 154, 169–173. [Google Scholar] [CrossRef] [Green Version]
- Uddin, L.Q. Mixed Signals: On Separating Brain Signal from Noise. Trends Cogn. Sci. 2017, 21, 405–406. [Google Scholar] [CrossRef]
- Aguirre, G.; Zarahn, E.; D’Esposito, M. The Inferential Impact of Global Signal Covariates in Functional Neuroimaging Analyses. NeuroImage 1998, 8, 302–306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verhaak, R.G.; Hoadley, K.; Purdom, E.; Wang, V.; Qi, Y.; Wilkerson, M.D.; Miller, C.; Ding, L.; Golub, T.; Mesirov, J.P.; et al. Integrated Genomic Analysis Identifies Clinically Relevant Subtypes of Glioblastoma Characterized by Abnormalities in PDGFRA, IDH1, EGFR, and NF1. Cancer Cell 2010, 17, 98–110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, C.; Sage, J.C.; Miller, M.R.; Verhaak, R.G.; Hippenmeyer, S.; Vogel, H.; Foreman, O.; Bronson, R.T.; Nishiyama, A.; Luo, L.; et al. Mosaic Analysis with Double Markers Reveals Tumor Cell of Origin in Glioma. Cell 2011, 146, 209–221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seidlitz, J.; Nadig, A.; Liu, S.; Bethlehem, R.A.I.; Vértes, P.E.; Morgan, S.E.; Váša, F.; Romero-Garcia, R.; Lalonde, F.M.; Clasen, L.S. Transcriptomic and cellular decoding of regional brain vulnerability to neurogenetic disorders. Nat. Commun. 2020, 11, 3358. [Google Scholar] [CrossRef] [PubMed]
- Mandal, A.S.; Romero-Garcia, R.; Seidlitz, J.; Hart, M.G.; Alexander-Bloch, A.; Suckling, J. Lesion covariance networks reveal proposed origins and pathways of diffuse gliomas. medRxiv 2021. [Google Scholar] [CrossRef]
- Venkatesh, H.S.; Johung, T.B.; Caretti, V.; Noll, A.; Tang, Y.; Nagaraja, S.; Gibson, E.M.; Mount, C.; Polepalli, J.; Mitra, S.S.; et al. Neuronal Activity Promotes Glioma Growth through Neuroligin-3 Secretion. Cell 2015, 161, 803–816. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Venkatesh, H.S.; Morishita, W.; Geraghty, A.C.; Silverbush, D.; Gillespie, S.M.; Arzt, M.; Tam, L.T.; Espenel, C.; Ponnuswami, A.; Ni, L.; et al. Electrical and synaptic integration of glioma into neural circuits. Nature 2019, 573, 539–545. [Google Scholar] [CrossRef]
- Venkataramani, V.; Tanev, D.I.; Strahle, C.; Studier-Fischer, A.; Fankhauser, L.; Kessler, T.; Körber, C.; Kardorff, M.; Ratliff, M.; Xie, R.; et al. Glutamatergic synaptic input to glioma cells drives brain tumour progression. Nature 2019, 573, 532–538. [Google Scholar] [CrossRef]
- Monje, M. Synaptic Communication in Brain Cancer. Cancer Res. 2020, 80, 2979–2982. [Google Scholar] [CrossRef]
- Yang, G.J.; Murray, J.D.; Glasser, M.; Pearlson, G.D.; Krystal, J.H.; Schleifer, C.; Repovs, G.; Anticevic, A. Altered Global Signal Topography in Schizophrenia. Cereb. Cortex 2016, 27, 5156–5169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Liao, W.; Han, S.; Li, J.; Zhang, Y.; Zhao, J.; Chen, H. Altered dynamic global signal topography in antipsychotic-naive adolescents with early-onset schizophrenia. Schizophr. Res. 2019, 208, 308–316. [Google Scholar] [CrossRef]
- Han, S.; Wang, X.; He, Z.; Sheng, W.; Zou, Q.; Li, L.; Yang, Y.; Guo, X.; Fan, Y.-S.; Guo, J.; et al. Decreased static and increased dynamic global signal topography in major depressive disorder. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2019, 94, 109665. [Google Scholar] [CrossRef]
- Li, R.; Wang, H.; Wang, L.; Zhang, L.; Zou, T.; Wang, X.; Liao, W.; Zhang, Z.; Lu, G.; Chen, H. Shared and distinct global signal topography disturbances in subcortical and cortical networks in human epilepsy. Hum. Brain Mapp. 2020, 42, 412–426. [Google Scholar] [CrossRef]
- Lv, Y.; Margulies, D.S.; Craddock, R.C.; Long, X.; Winter, B.; Gierhake, D.; Endres, M.; Villringer, K.; Fiebach, J.; Villringer, A. Identifying the perfusion deficit in acute stroke with resting-state functional magnetic resonance imaging. Ann. Neurol. 2012, 73, 136–140. [Google Scholar] [CrossRef]
- Perry, A.; Wesseling, P. Histologic classification of gliomas. Handb. Clin. Neurol. 2016, 134, 71–95. [Google Scholar] [CrossRef]
- Huang, Q.; Zhang, R.; Hu, X.; Ding, S.; Qian, J.; Lei, T.; Cao, X.; Tao, L.; Qian, Z.; Liu, H. Disturbed Small-World Networks and Neurocognitive Function in Frontal Lobe Low-Grade Glioma Patients. PLoS ONE 2014, 9, e94095. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Ding, S.; Hu, X.; Yang, K.; Xiao, C.; Zou, Y.; Chen, Y.; Tao, L.; Liu, H.; Qian, Z. Reduced efficiency of functional brain network underlying intellectual decline in patients with low-grade glioma. Neurosci. Lett. 2013, 543, 27–31. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, M.R.; Johnson, S.P.; Ramasawmy, R.; Pedley, R.B.; Lythgoe, M.F.; Walker-Samuel, S. Decomposition of spontaneous fluctuations in tumour oxygenation using BOLD MRI and independent component analysis. Br. J. Cancer 2015, 113, 1168–1177. [Google Scholar] [CrossRef] [PubMed]
- Chow, D.S.; Horenstein, C.I.; Canoll, P.; Lignelli, A.; Hillman, E.M.C.; Filippi, C.G.; Grinband, J. Glioblastoma Induces Vascular Dysregulation in Nonenhancing Peritumoral Regions in Humans. Am. J. Roentgenol. 2016, 206, 1073–1081. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pillai, J.J.; Zacà, D. Comparison of BOLD Cerebrovascular Reactivity Mapping and DSC MR Perfusion Imaging for Prediction of Neurovascular Uncoupling Potential in Brain Tumors. Technol. Cancer Res. Treat. 2012, 11, 361–374. [Google Scholar] [CrossRef]
- Bartolomei, F.; Bosma, I.; Klein, M.; Baayen, J.C.; Reijneveld, J.C.; Postma, T.J.; Heimans, J.J.; van Dijk, B.W.; De Munck, J.; de Jongh, A.; et al. Disturbed functional connectivity in brain tumour patients: Evaluation by graph analysis of synchronization matrices. Clin. Neurophysiol. 2006, 117, 2039–2049. [Google Scholar] [CrossRef]
- Zimmermann, M.; Rössler, K.; Kaltenhäuser, M.; Grummich, P.; Brandner, N.; Buchfelder, M.; Dörfler, A.; Kölble, K.; Stadlbauer, A. Comparative fMRI and MEG localization of cortical sensorimotor function: Bimodal mapping supports motor area reorganization in glioma patients. PLoS ONE 2019, 14, e0213371. [Google Scholar] [CrossRef] [Green Version]
- Bowden, S.; Gill, B.; Englander, Z.; Horenstein, C.; Zanazzi, G.; Chang, P.; Samanamud, J.; Lignelli, A.; Bruce, J.; Canoll, P.; et al. Local Glioma Cells Are Associated with Vascular Dysregulation. Am. J. Neuroradiol. 2018, 39, 507–514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sagberg, L.M.; Drewes, C.; Jakola, A.; Solheim, O. Accuracy of operating neurosurgeons’ prediction of functional levels after intracranial tumor surgery. J. Neurosurg. 2017, 126, 1173–1180. [Google Scholar] [CrossRef] [Green Version]
- Duffau, H. New Philosophy, Clinical Pearls, and Methods for Intraoperative Cognition Mapping and Monitoring “à la carte” in Brain Tumor Patients. Neurosurgery 2021, 88, 919–930. [Google Scholar] [CrossRef] [PubMed]
- Desmurget, M.; Bonnetblanc, F.; Duffau, H. Contrasting acute and slow-growing lesions: A new door to brain plasticity. Brain 2006, 130, 898–914. [Google Scholar] [CrossRef] [Green Version]
- Wefel, J.S.; Noll, K.R.; Scheurer, M.E. Neurocognitive functioning and genetic variation in patients with primary brain tumours. Lancet Oncol. 2016, 17, e97–e108. [Google Scholar] [CrossRef] [Green Version]
- Romero-Garcia, R.; Suckling, J.; Owen, M.; Assem, M.; Sinha, R.; Coelho, P.; Woodberry, E.; Price, S.J.; Burke, A.; Santarius, T.; et al. Memory recovery is related to default mode network impairment and neurite density during brain tumours treatment. J. Neurosurg. 2020, 1–11. [Google Scholar] [CrossRef]
- Maesawa, S.; Bagarinao, E.; Fujii, M.; Futamura, M.; Motomura, K.; Watanabe, H.; Mori, D.; Sobue, G.; Wakabayashi, T. Evaluation of Resting State Networks in Patients with Gliomas: Connectivity Changes in the Unaffected Side and Its Relation to Cognitive Function. PLoS ONE 2015, 10, e0118072. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herbet, G.; Maheu, M.; Costi, E.; LaFargue, G.; Duffau, H. Mapping neuroplastic potential in brain-damaged patients. Brain 2016, 139, 829–844. [Google Scholar] [CrossRef] [PubMed]
- Kliemann, D.; Adolphs, R.; Tyszka, J.M.; Fischl, B.; Yeo, B.T.; Nair, R.; Dubois, J.; Paul, L.K. Intrinsic Functional Connectivity of the Brain in Adults with a Single Cerebral Hemisphere. Cell Rep. 2019, 29, 2398–2407.e4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holodny, A.I.; Shevzov-Zebrun, N.; Brennan, N.; Peck, K.K. Motor and Sensory Mapping. Neurosurg. Clin. N. Am. 2011, 22, 207–218. [Google Scholar] [CrossRef]
- Black, D.; Vachha, B.; Mian, A.; Faro, S.; Maheshwari, M.; Sair, H.; Petrella, J.; Pillai, J.; Welker, K. American Society of Functional Neuroradiology–Recommended fMRI Paradigm Algorithms for Presurgical Language Assessment. Am. J. Neuroradiol. 2017, 38, E65–E73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bressler, S.L.; Menon, V. Large-scale brain networks in cognition: Emerging methods and principles. Trends Cogn. Sci. 2010, 14, 277–290. [Google Scholar] [CrossRef]
- Gomez-Feria, J.; Narros, J.L.; Ciriza, G.G.; Roldan-Lora, F.; Schrader, I.M.; Martin-Rodríguez, J.F.; Mir, P. 3D Printing of Diffuse Low-Grade Gliomas Involving Eloquent Cortical Areas and Subcortical Functional Pathways: Technical Note. World Neurosurg. 2021, 147, 164–171.e4. [Google Scholar] [CrossRef]
- Hu, S.; Kang, H.; Baek, Y.; El Fakhri, G.; Kuang, A.; Choi, H.S. Real-Time Imaging of Brain Tumor for Image-Guided Surgery. Adv. Health Mater. 2018, 7, e1800066. [Google Scholar] [CrossRef] [PubMed]
- Mikhail, M.; Mithani, K.; Ibrahim, G.M. Presurgical and Intraoperative Augmented Reality in Neuro-Oncologic Surgery: Clinical Experiences and Limitations. World Neurosurg. 2019, 128, 268–276. [Google Scholar] [CrossRef]
- McCulloch, P.; Altman, D.G.; Campbell, W.B.; Flum, D.R.; Glasziou, P.; Marshall, J.C.; Nicholl, J. No surgical innovation without evaluation: The IDEAL recommendations. Lancet 2009, 374, 1105–1112. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Romero-Garcia, R.; Hart, M.G.; Bethlehem, R.A.I.; Mandal, A.; Assem, M.; Crespo-Facorro, B.; Gorriz, J.M.; Burke, G.A.A.; Price, S.J.; Santarius, T.; et al. BOLD Coupling between Lesioned and Healthy Brain Is Associated with Glioma Patients’ Recovery. Cancers 2021, 13, 5008. https://doi.org/10.3390/cancers13195008
Romero-Garcia R, Hart MG, Bethlehem RAI, Mandal A, Assem M, Crespo-Facorro B, Gorriz JM, Burke GAA, Price SJ, Santarius T, et al. BOLD Coupling between Lesioned and Healthy Brain Is Associated with Glioma Patients’ Recovery. Cancers. 2021; 13(19):5008. https://doi.org/10.3390/cancers13195008
Chicago/Turabian StyleRomero-Garcia, Rafael, Michael G. Hart, Richard A. I. Bethlehem, Ayan Mandal, Moataz Assem, Benedicto Crespo-Facorro, Juan Manuel Gorriz, G. A. Amos Burke, Stephen J. Price, Thomas Santarius, and et al. 2021. "BOLD Coupling between Lesioned and Healthy Brain Is Associated with Glioma Patients’ Recovery" Cancers 13, no. 19: 5008. https://doi.org/10.3390/cancers13195008