
Folic Acid-Appended Hydroxypropyl-β-Cyclodextrin Exhibits Potent Antitumor Activity in Chronic Myeloid Leukemia Cells via Autophagic Cell Death

 , and
, and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Reagents and Cell Lines
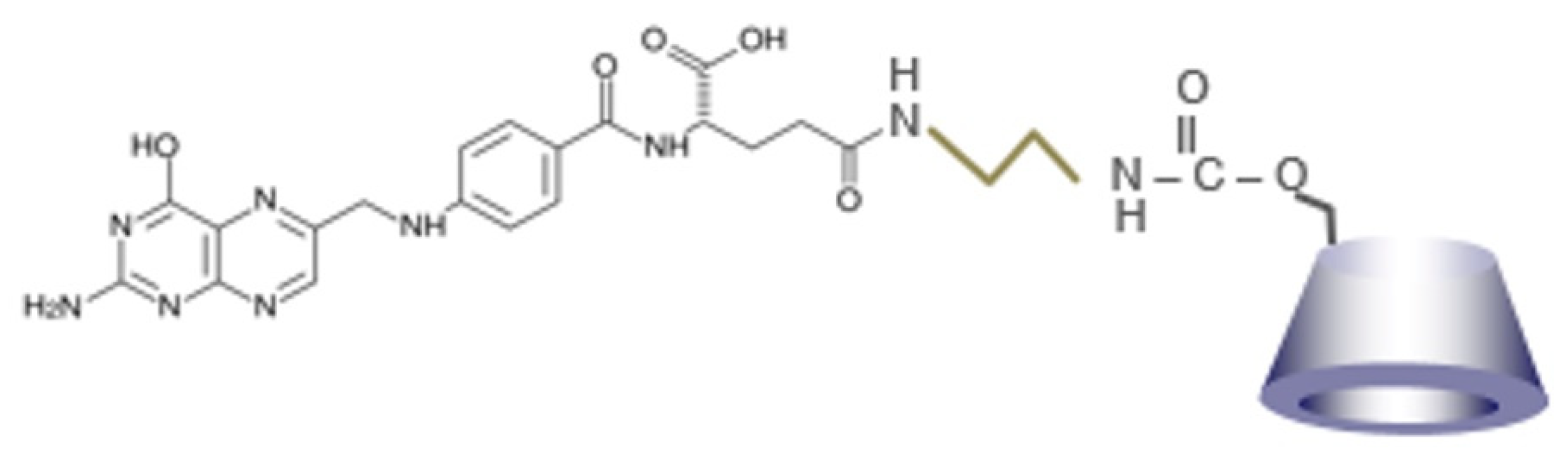
2.2. Preparation of FA-HP-β-CyD
2.3. Mice
2.4. Analysis of Cell Growth and Viability
2.5. In Vitro Antitumor Activity in the Presence of a Competitive Inhibitor of FR
2.6. Flow Cytometric Analysis
2.7. Western Blot Analysis
2.8. Cellular Association of FA-HP-β-CyD
2.9. Intracellular Distribution of TRITC-FA-HP-β-CyD
2.10. Activation of Caspases 3/7
2.11. Autophagosome Formation
2.12. Cell Culture Using Autophagy Inhibitors
2.13. Detection of Mitophagy
2.14. Intracellular Adenosine Triphosphate (ATP) Quantification
2.15. ROS Generation Assay
2.16. Statistical Analysis
3. Results
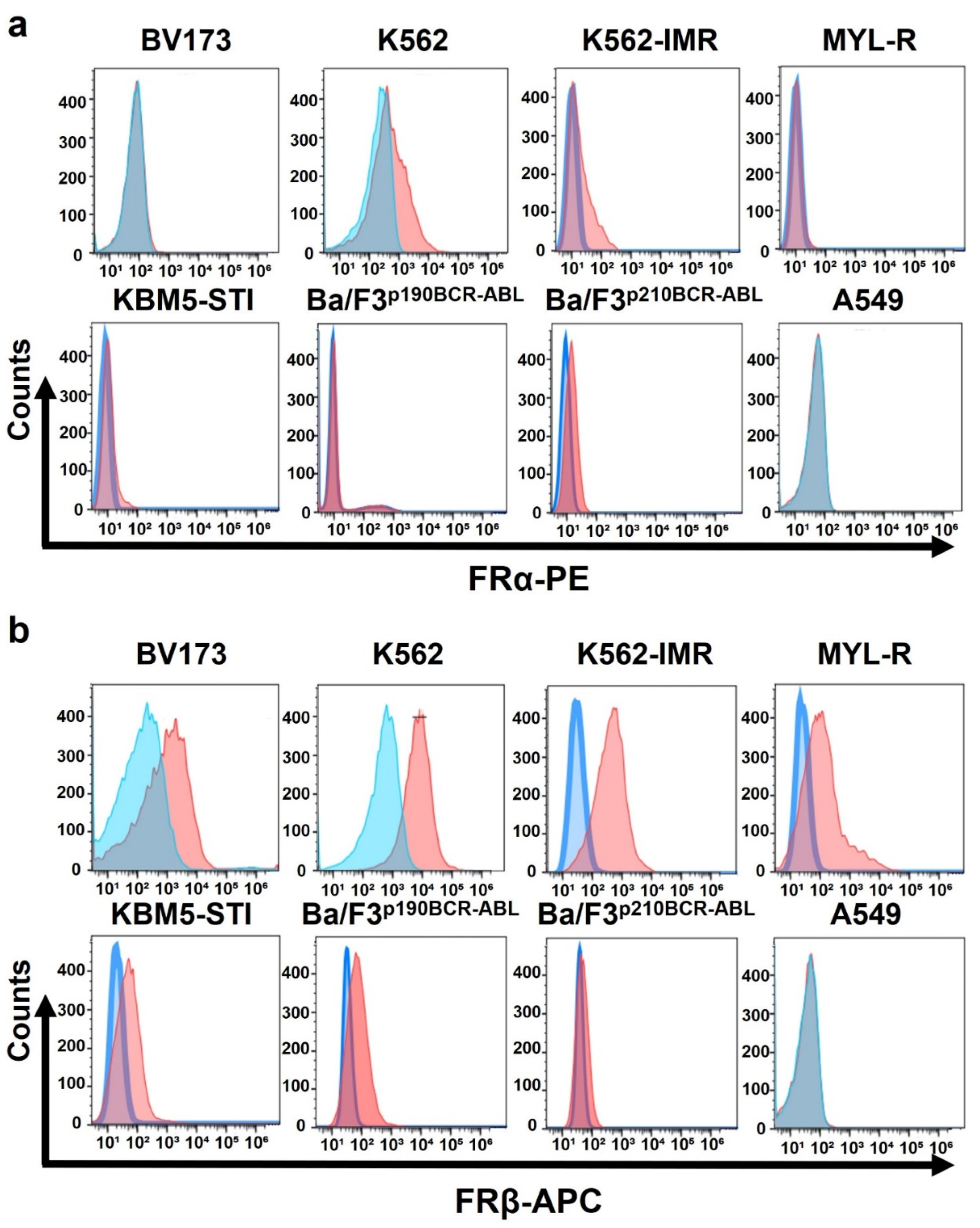
3.1. Leukemia Cell Lines Highly Express Folate Receptor β but Not Folate Receptor α
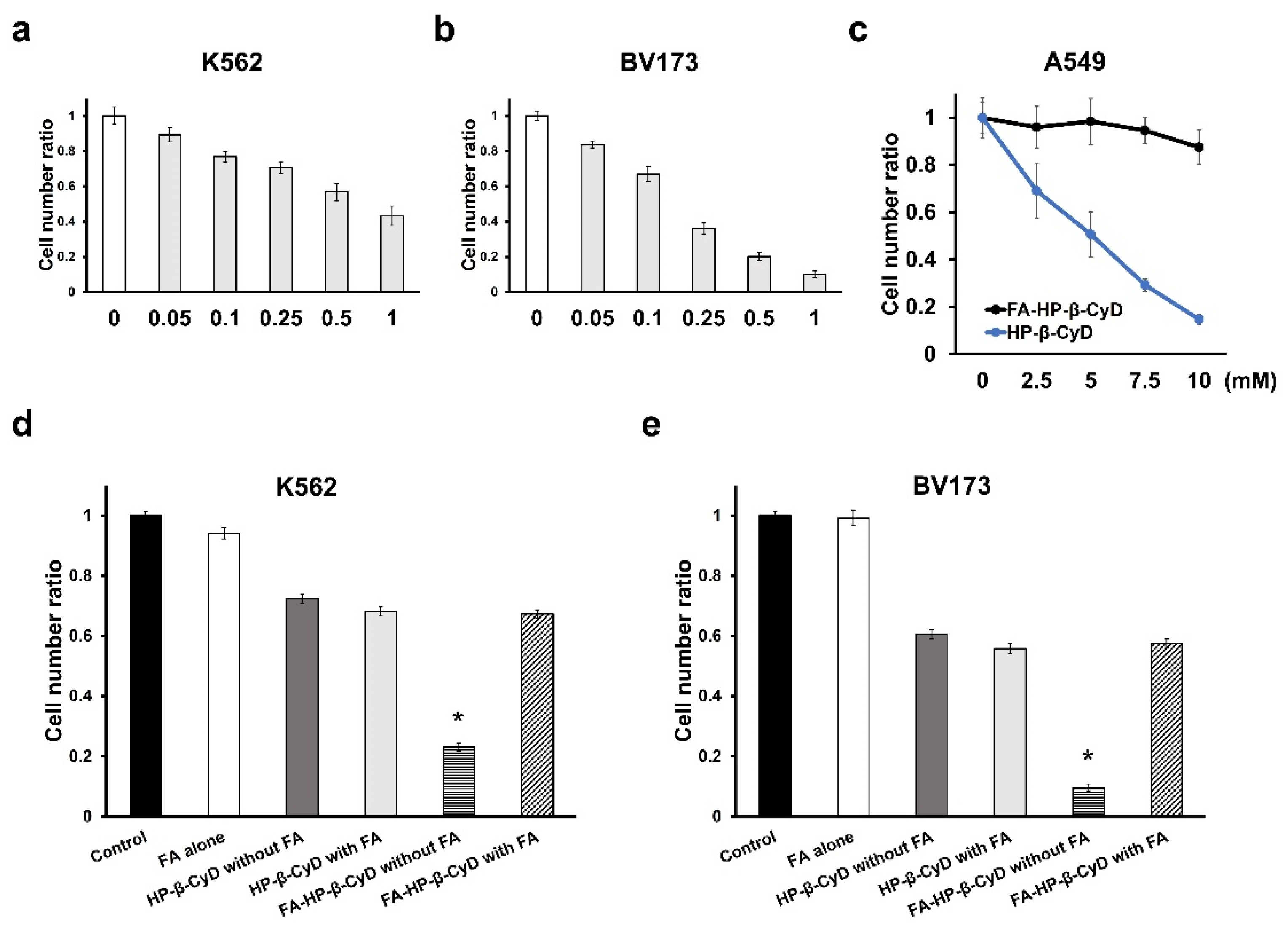
3.2. Anti-Leukemia Effect of FA-HP-β-CyD
3.3. FA-HP-β-CyD Inhibits the Growth of Leukemia Cell Lines More Potently than HP-β-CyD
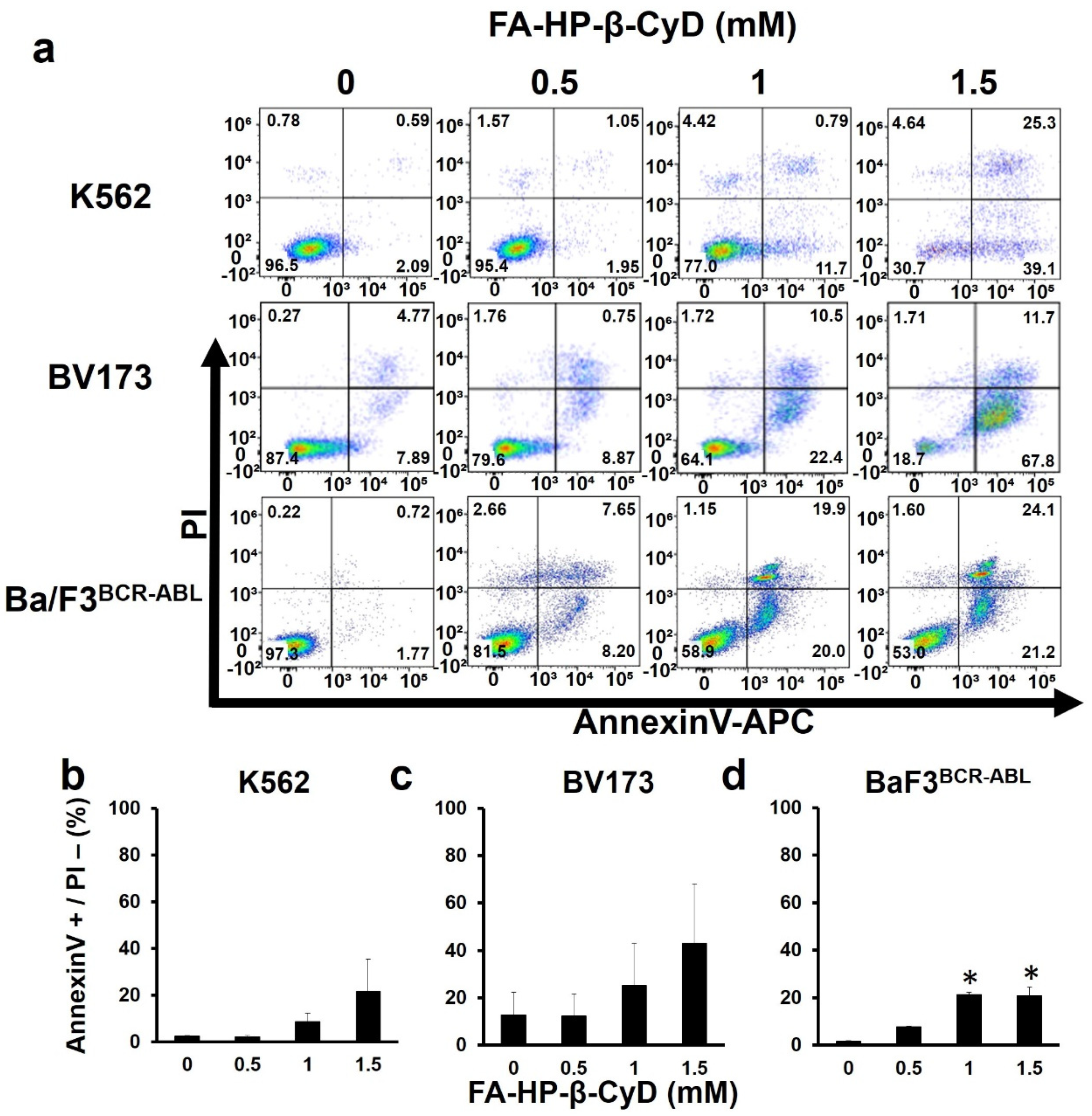
3.4. FA-HP-β-CyD Inhibits Cell Growth by Inducing Apoptosis
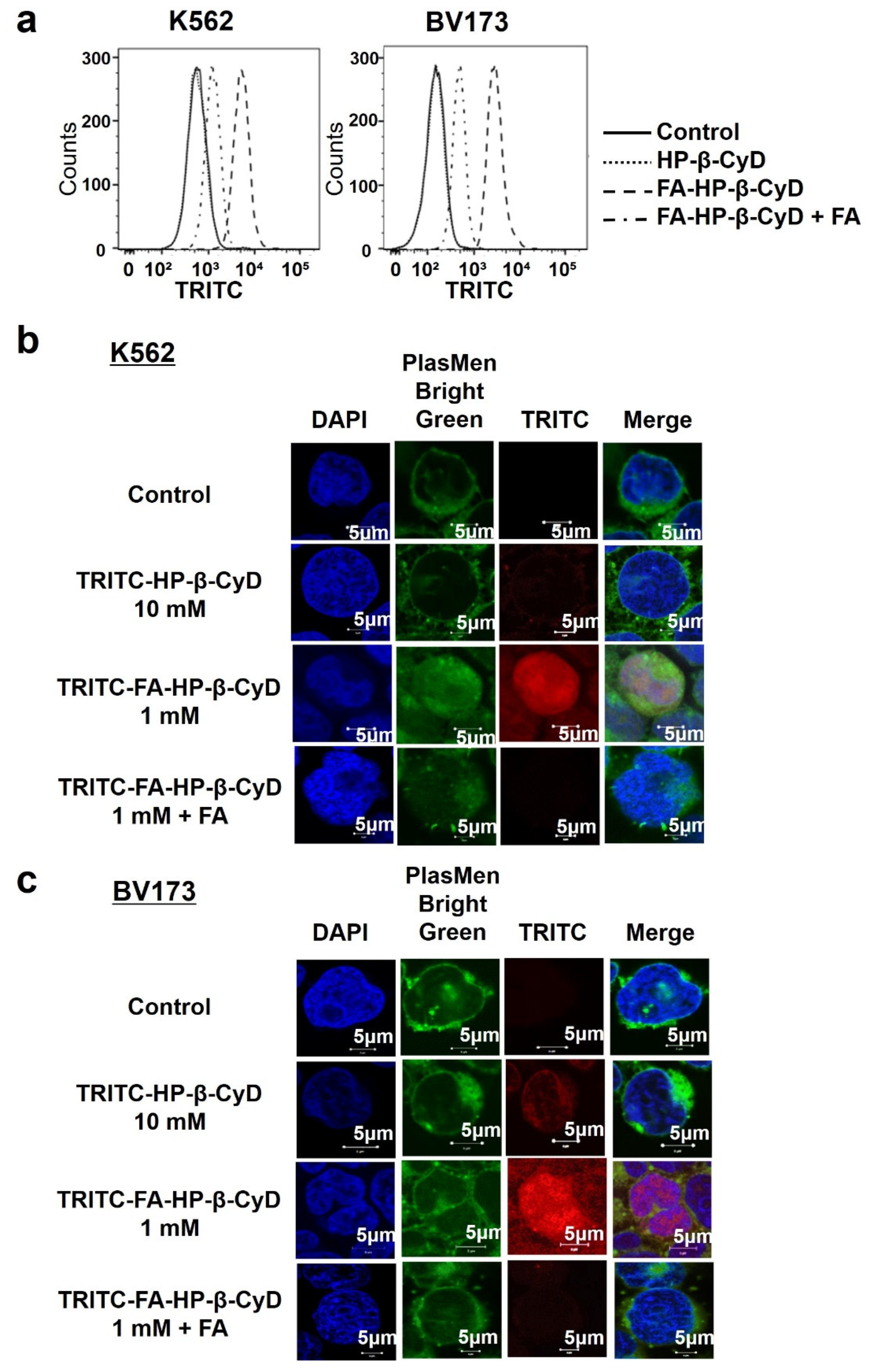
3.5. Cellular Uptake of FA-HP-β-CyD
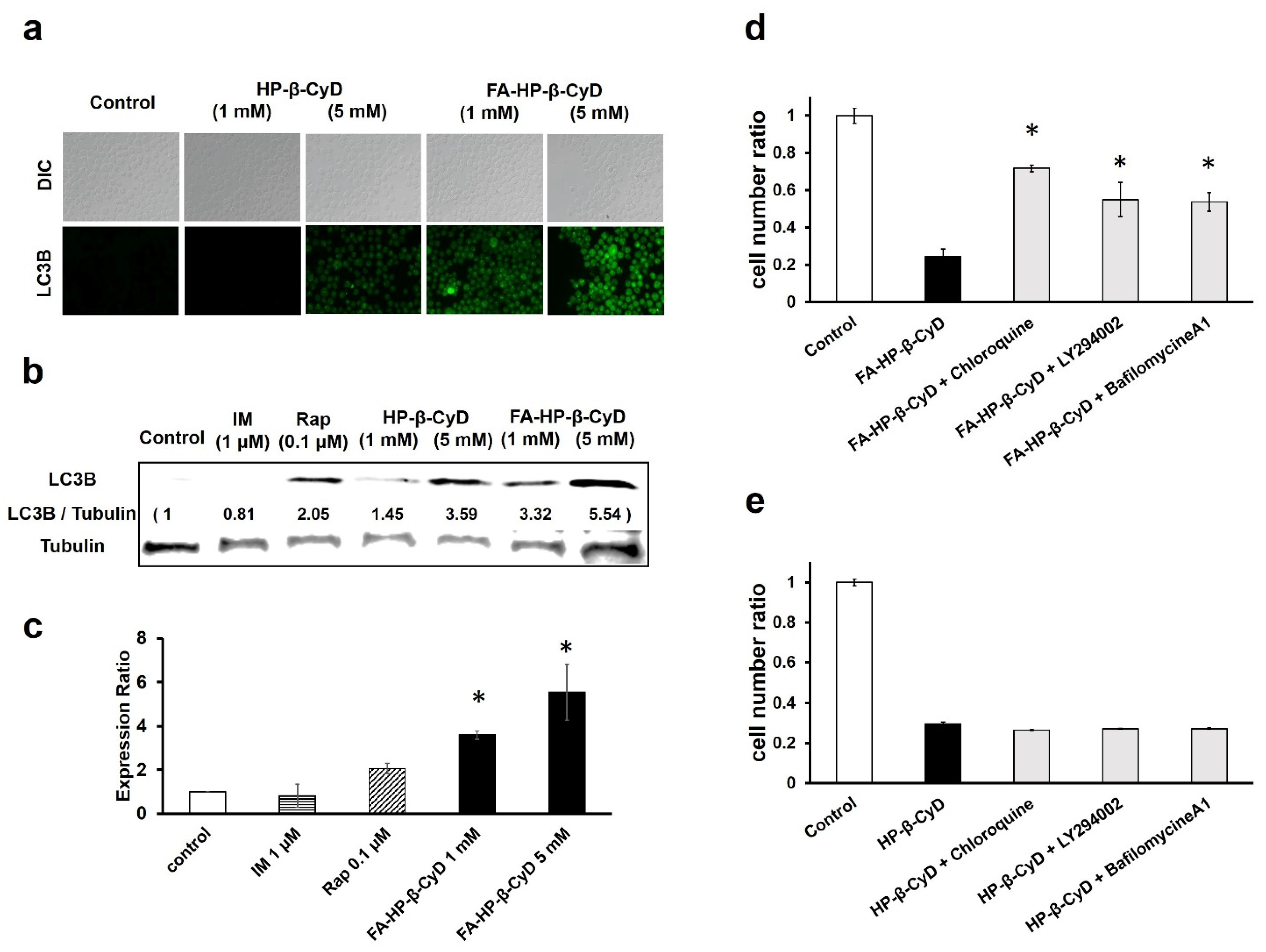
3.6. FA-HP-β-CyD Induces Autophagosome Formation
3.7. The Antitumor Activity of FA-HP-β-CyD Is Reduced by Autophagy Inhibition
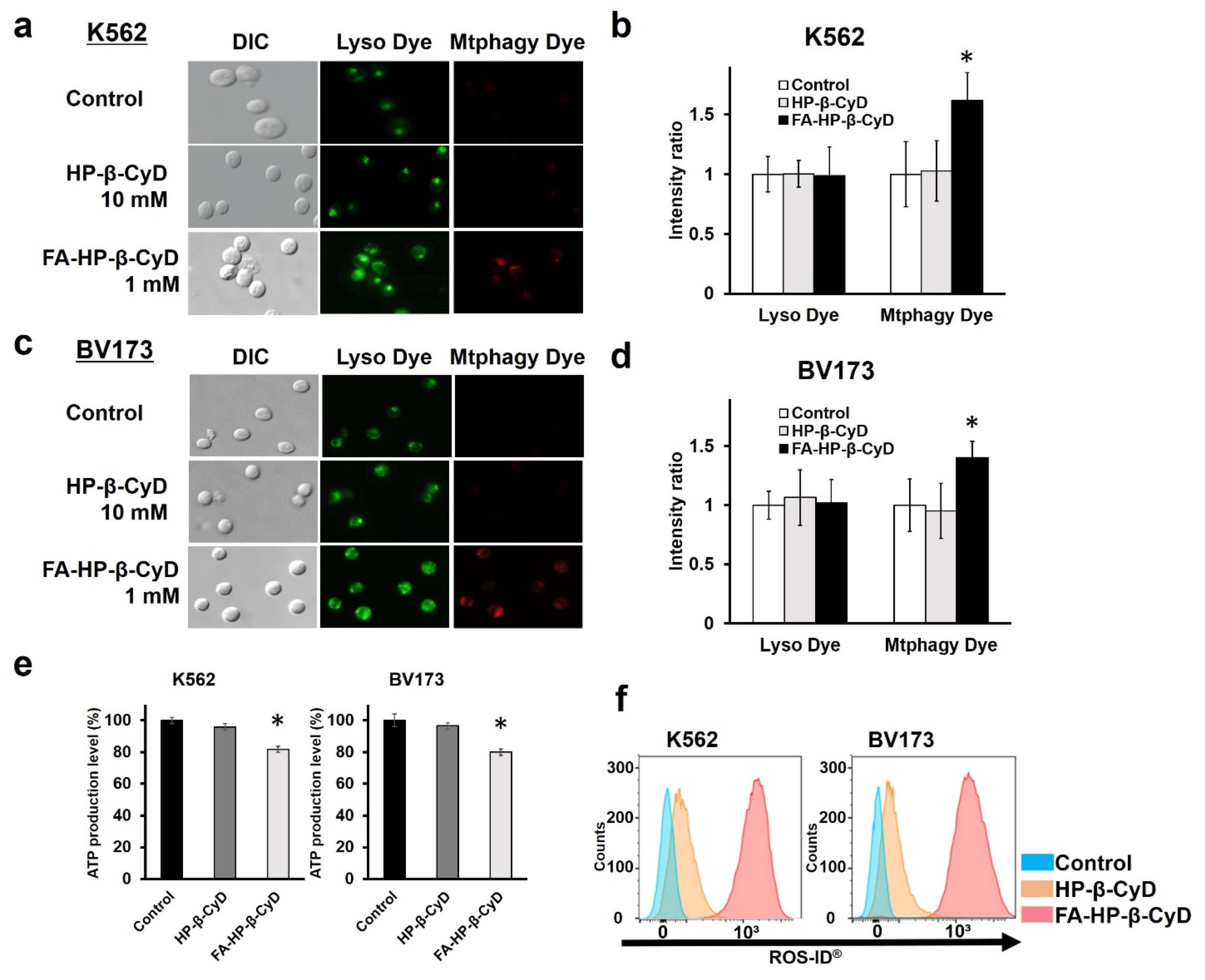
3.8. Induction of Mitophagy and Mitochondrial Dysfunction by FA-HP-β-CyD
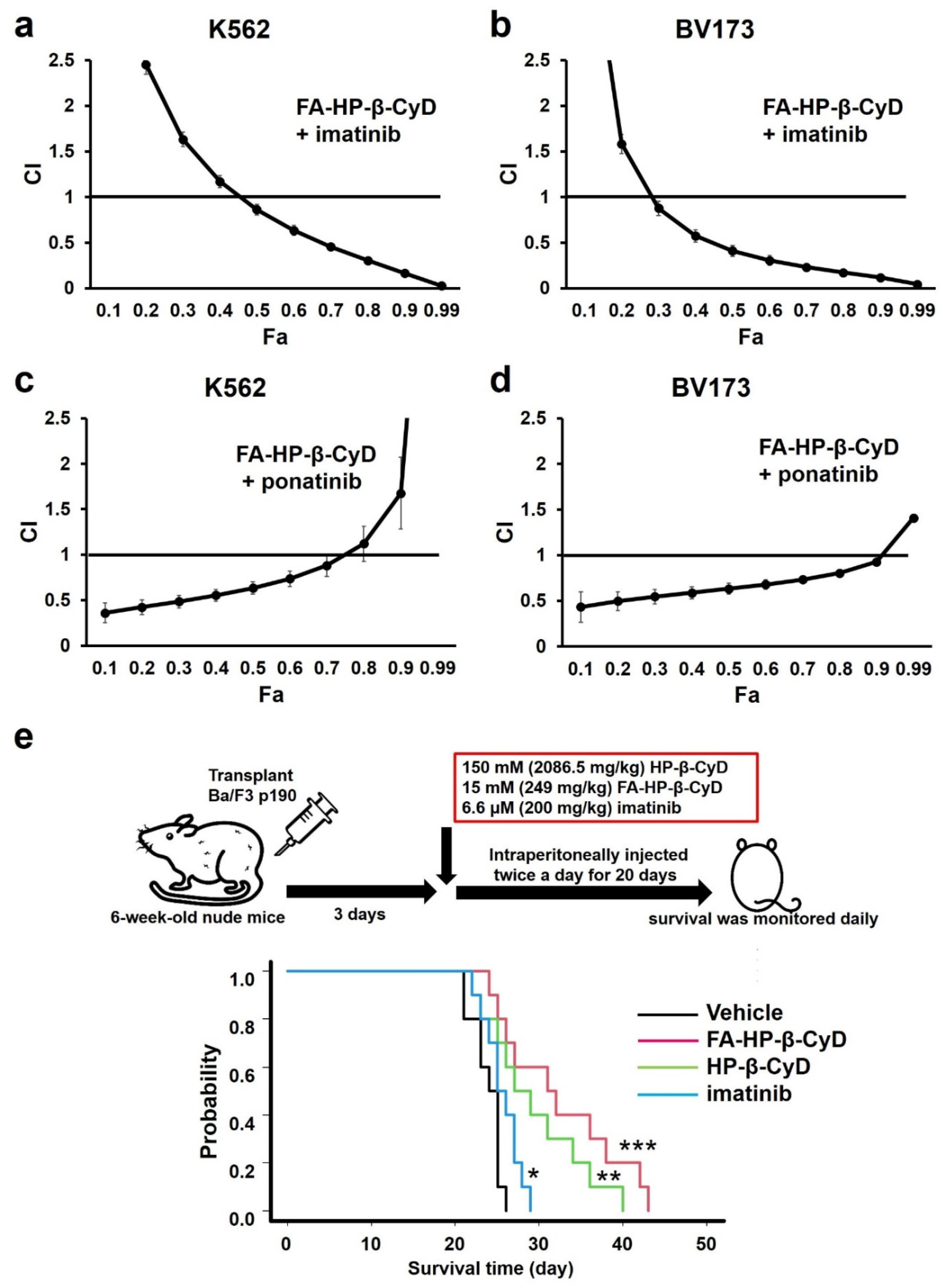
3.9. FA-HP-β-CyD Synergistically Increases the Effect of ABL Tyrosine Kinase Inhibitors
3.10. Administration of FA-HP-β-CyD Prolongs Survival in a Leukemia Mouse Model
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kimura, S.; Ando, T.; Kojima, K. Ever-advancing chronic myeloid leukemia treatment. Int. J. Clin. Oncol. 2014, 19, 3–9. [Google Scholar] [CrossRef]
- O’Brien, S.G.; Guilhot, F.; Larson, R.A.; Gathmann, I.; Baccarani, M.; Cervantes, F.; Cornelissen, J.J.; Fischer, T.; Hochhaus, A.; Hughes, T.; et al. Imatinib compared with interferon and low-dose cytarabine for newly diagnosed chronic-phase chronic myeloid leukemia. N. Engl. J. Med. 2003, 348, 994–1004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minciacchi, V.R.; Kumar, R.; Krause, D.S. Chronic Myeloid Leukemia: A Model Disease of the Past, Present and Future. Cells 2021, 10, 117. [Google Scholar] [CrossRef]
- Davis, M.E.; Brewster, M.E. Cyclodextrin-based pharmaceutics: Past, present and future. Nat. Rev. Drug Discov. 2004, 3, 1023–1035. [Google Scholar] [CrossRef]
- De Beule, K.; Van Gestel, J. Pharmacology of itraconazole. Drugs 2001, 61 (Suppl. 1), 27–37. [Google Scholar] [CrossRef]
- Matsuo, M.; Togawa, M.; Hirabaru, K.; Mochinaga, S.; Narita, A.; Adachi, M.; Egashira, M.; Irie, T.; Ohno, K. Effects of cyclodextrin in two patients with Niemann-Pick Type C disease. Mol. Genet. Metab. 2013, 108, 76–81. [Google Scholar] [CrossRef] [PubMed]
- Yamada, Y.; Ishitsuka, Y.; Kondo, Y.; Nakahara, S.; Nishiyama, A.; Takeo, T.; Nakagata, N.; Motoyama, K.; Higashi, T.; Arima, H.; et al. Differential mode of cholesterol inclusion with 2-hydroxypropyl-cyclodextrins increases safety margin in treatment of Niemann-Pick disease type C. Br. J. Pharmacol. 2021, 178, 2727–2746. [Google Scholar] [CrossRef]
- Yokoo, M.; Kubota, Y.; Motoyama, K.; Higashi, T.; Taniyoshi, M.; Tokumaru, H.; Nishiyama, R.; Tabe, Y.; Mochinaga, S.; Sato, A.; et al. 2-Hydroxypropyl-beta-Cyclodextrin Acts as a Novel Anticancer Agent. PLoS ONE 2015, 10, e0141946. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parker, N.; Turk, M.J.; Westrick, E.; Lewis, J.D.; Low, P.S.; Leamon, C.P. Folate receptor expression in carcinomas and normal tissues determined by a quantitative radioligand binding assay. Anal. Biochem. 2005, 338, 284–293. [Google Scholar] [CrossRef]
- Assaraf, Y.G.; Leamon, C.P.; Reddy, J.A. The folate receptor as a rational therapeutic target for personalized cancer treatment. Drug Resist. Updates 2014, 17, 89–95. [Google Scholar] [CrossRef]
- Onodera, R.; Motoyama, K.; Arima, H. Design and evaluation of folate-appended methyl-β-cyclodextrin as a new antitumor agent. J. Incl. Phenom. Macrocycl. Chem. 2011, 70, 321–326. [Google Scholar] [CrossRef]
- Onodera, R.; Motoyama, K.; Okamatsu, A.; Higashi, T.; Arima, H. Potential use of folate-appended methyl-beta-cyclodextrin as an anticancer agent. Sci. Rep. 2013, 3, 1104. [Google Scholar] [CrossRef] [Green Version]
- Onodera, R.; Motoyama, K.; Tanaka, N.; Ohyama, A.; Okamatsu, A.; Higashi, T.; Kariya, R.; Okada, S.; Arima, H. Involvement of autophagy in antitumor activity of folate-appended methyl-beta-cyclodextrin. Sci. Rep. 2014, 4, 4417. [Google Scholar] [CrossRef] [Green Version]
- Kameyama, K.; Motoyama, K.; Tanaka, N.; Yamashita, Y.; Higashi, T.; Arima, H. Induction of mitophagy-mediated antitumor activity with folate-appended methyl-beta-cyclodextrin. Int. J. Nanomed. 2017, 12, 3433–3446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ricci, C.; Scappini, B.; Divoky, V.; Gatto, S.; Onida, F.; Verstovsek, S.; Kantarjian, H.M.; Beran, M. Mutation in the ATP-binding pocket of the ABL kinase domain in an STI571-resistant BCR/ABL-positive cell line. Cancer Res. 2002, 62, 5995–5998. [Google Scholar]
- Ito, T.; Tanaka, H.; Kimura, A. Establishment and characterization of a novel imatinib-sensitive chronic myeloid leukemia cell line MYL, and an imatinib-resistant subline MYL-R showing overexpression of Lyn. Eur. J. Haematol. 2007, 78, 417–431. [Google Scholar] [CrossRef] [PubMed]
- Kimura, S.; Naito, H.; Segawa, H.; Kuroda, J.; Yuasa, T.; Sato, K.; Yokota, A.; Kamitsuji, Y.; Kawata, E.; Ashihara, E.; et al. NS-187, a potent and selective dual Bcr-Abl/Lyn tyrosine kinase inhibitor, is a novel agent for imatinib-resistant leukemia. Blood 2005, 106, 3948–3954. [Google Scholar] [CrossRef] [PubMed]
- Yokoo, M.; Kubota, Y.; Tabe, Y.; Kimura, S. Comparative study of the anti-leukemic effects of imatinib mesylate, Glivec tablet and its generic formulation, OHK9511. Biol. Pharm. Bull. 2015, 38, 411–416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoshiko, T.; Kubota, Y.; Akisawa, T.; Watanabe, T.; Tanigawara, K.; Yano, J.; Kimura, S. Naked antisense double-stranded DNA oligonucleotide efficiently suppresses BCR-ABL positive leukemic cells. Investig. New Drugs 2020, 38, 1012–1019. [Google Scholar] [CrossRef]
- Nishida, Y.; Maeda, A.; Kim, M.J.; Cao, L.; Kubota, Y.; Ishizawa, J.; AlRawi, A.; Kato, Y.; Iwama, A.; Fujisawa, M.; et al. The novel BMI-1 inhibitor PTC596 downregulates MCL-1 and induces p53-independent mitochondrial apoptosis in acute myeloid leukemia progenitor cells. Blood Cancer J. 2017, 7, e527. [Google Scholar] [CrossRef]
- Irie, T.; Uekama, K. Pharmaceutical applications of cyclodextrins. III. Toxicological issues and safety evaluation. J. Pharm. Sci. 1997, 86, 147–162. [Google Scholar] [CrossRef] [PubMed]
- Azad, M.B.; Chen, Y.; Gibson, S.B. Regulation of autophagy by reactive oxygen species (ROS): Implications for cancer progression and treatment. Antioxid. Redox Signal. 2009, 11, 777–790. [Google Scholar] [CrossRef] [PubMed]
- Dan Dunn, J.; Alvarez, L.A.; Zhang, X.; Soldati, T. Reactive oxygen species and mitochondria: A nexus of cellular homeostasis. Redox Biol. 2015, 6, 472–485. [Google Scholar] [CrossRef] [PubMed]
- Yun, C.W.; Lee, S.H. The Roles of Autophagy in Cancer. Int. J. Mol. Sci. 2018, 19, 3466. [Google Scholar] [CrossRef] [Green Version]
- Baquero, P.; Dawson, A.; Helgason, G.V. Autophagy and mitochondrial metabolism: Insights into their role and therapeutic potential in chronic myeloid leukaemia. FEBS J. 2019, 286, 1271–1283. [Google Scholar] [CrossRef] [PubMed]
- Baquero, P.; Dawson, A.; Mukhopadhyay, A.; Kuntz, E.M.; Mitchell, R.; Olivares, O.; Ianniciello, A.; Scott, M.T.; Dunn, K.; Nicastri, M.C.; et al. Targeting quiescent leukemic stem cells using second generation autophagy inhibitors. Leukemia 2019, 33, 981–994. [Google Scholar] [CrossRef] [Green Version]
- Altman, B.J.; Jacobs, S.R.; Mason, E.F.; Michalek, R.D.; MacIntyre, A.N.; Coloff, J.L.; Ilkayeva, O.; Jia, W.; He, Y.W.; Rathmell, J.C. Autophagy is essential to suppress cell stress and to allow BCR-Abl-mediated leukemogenesis. Oncogene 2011, 30, 1855–1867. [Google Scholar] [CrossRef] [Green Version]
- Han, W.; Sun, J.; Feng, L.; Wang, K.; Li, D.; Pan, Q.; Chen, Y.; Jin, W.; Wang, X.; Pan, H.; et al. Autophagy inhibition enhances daunorubicin-induced apoptosis in K562 cells. PLoS ONE 2011, 6, e28491. [Google Scholar] [CrossRef]
- Bosnjak, M.; Ristic, B.; Arsikin, K.; Mircic, A.; Suzin-Zivkovic, V.; Perovic, V.; Bogdanovic, A.; Paunovic, V.; Markovic, I.; Bumbasirevic, V.; et al. Inhibition of mTOR-dependent autophagy sensitizes leukemic cells to cytarabine-induced apoptotic death. PLoS ONE 2014, 9, e94374. [Google Scholar] [CrossRef]
- Carew, J.S.; Nawrocki, S.T.; Kahue, C.N.; Zhang, H.; Yang, C.; Chung, L.; Houghton, J.A.; Huang, P.; Giles, F.J.; Cleveland, J.L. Targeting autophagy augments the anticancer activity of the histone deacetylase inhibitor SAHA to overcome Bcr-Abl-mediated drug resistance. Blood 2007, 110, 313–322. [Google Scholar] [CrossRef] [Green Version]
- Isakson, P.; Bjoras, M.; Boe, S.O.; Simonsen, A. Autophagy contributes to therapy-induced degradation of the PML/RARA oncoprotein. Blood 2010, 116, 2324–2331. [Google Scholar] [CrossRef] [PubMed]
- Goussetis, D.J.; Gounaris, E.; Wu, E.J.; Vakana, E.; Sharma, B.; Bogyo, M.; Altman, J.K.; Platanias, L.C. Autophagic degradation of the BCR-ABL oncoprotein and generation of antileukemic responses by arsenic trioxide. Blood 2012, 120, 3555–3562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bellodi, C.; Lidonnici, M.R.; Hamilton, A.; Helgason, G.V.; Soliera, A.R.; Ronchetti, M.; Galavotti, S.; Young, K.W.; Selmi, T.; Yacobi, R.; et al. Targeting autophagy potentiates tyrosine kinase inhibitor-induced cell death in Philadelphia chromosome-positive cells, including primary CML stem cells. J. Clin. Investig. 2009, 119, 1109–1123. [Google Scholar] [CrossRef] [PubMed]
- Rothe, K.; Lin, H.; Lin, K.B.; Leung, A.; Wang, H.M.; Malekesmaeili, M.; Brinkman, R.R.; Forrest, D.L.; Gorski, S.M.; Jiang, X. The core autophagy protein ATG4B is a potential biomarker and therapeutic target in CML stem/progenitor cells. Blood 2014, 123, 3622–3634. [Google Scholar] [CrossRef] [PubMed]
- Karvela, M.; Baquero, P.; Kuntz, E.M.; Mukhopadhyay, A.; Mitchell, R.; Allan, E.K.; Chan, E.; Kranc, K.R.; Calabretta, B.; Salomoni, P.; et al. ATG7 regulates energy metabolism, differentiation and survival of Philadelphia-chromosome-positive cells. Autophagy 2016, 12, 936–948. [Google Scholar] [CrossRef] [Green Version]
- Vetrie, D.; Helgason, G.V.; Copland, M. The leukaemia stem cell: Similarities, differences and clinical prospects in CML and AML. Nat. Rev. Cancer 2020, 20, 158–173. [Google Scholar] [CrossRef]
- Yung, Y.; Lee, E.; Chu, H.T.; Yip, P.K.; Gill, H. Targeting Abnormal Hematopoietic Stem Cells in Chronic Myeloid Leukemia and Philadelphia Chromosome-Negative Classical Myeloproliferative Neoplasms. Int. J. Mol. Sci. 2021, 22, 659. [Google Scholar] [CrossRef]
- Yan, J.; Li, Q.F.; Wang, L.S.; Wang, H.; Xiao, F.J.; Yang, Y.F.; Wu, C.T. Methyl-beta-cyclodextrin induces programmed cell death in chronic myeloid leukemia cells and, combined with imatinib, produces a synergistic downregulation of ERK/SPK1 signaling. Anticancer Drugs 2012, 23, 22–31. [Google Scholar] [CrossRef]
- Murthy, H.; Anasetti, C.; Ayala, E. Diagnosis and Management of Leukemic and Lymphomatous Meningitis. Cancer Control 2017, 24, 33–41. [Google Scholar] [CrossRef]
- Matsuo, M.; Shraishi, K.; Wada, K.; Ishitsuka, Y.; Doi, H.; Maeda, M.; Mizoguchi, T.; Eto, J.; Mochinaga, S.; Arima, H.; et al. Effects of intracerebroventricular administration of 2-hydroxypropyl-β-cyclodextrin in a patient with Niemann-Pick Type C disease. Mol. Genet. Metab. Rep. 2014, 1, 391–400. [Google Scholar] [CrossRef] [PubMed]
- Fukaura, M.; Ishitsuka, Y.; Shirakawa, S.; Ushihama, N.; Yamada, Y.; Kondo, Y.; Takeo, T.; Nakagata, N.; Motoyama, K.; Higashi, T.; et al. Intracerebroventricular Treatment with 2-Hydroxypropyl-β-Cyclodextrin Decreased Cerebellar and Hepatic Glycoprotein Nonmetastatic Melanoma Protein B (GPNMB) Expression in Niemann-Pick Disease Type C Model Mice. Int. J. Mol. Sci. 2021, 22, 452. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| IC50 of Cell Lines | IC50 (mM) | |
|---|---|---|
| Cell Line | HP-β-CyD | FA-HP-β-CyD |
| K562 | 5.65 ± 0.96 | 0.69 ± 0.19 |
| BV173 | 3.46 ± 0.33 | 0.17 ± 0.06 |
| Ba/F3p190 BCR-ABL | 8.04 ± 0.71 | 0.91 ± 0.16 |
| Ba/F3p210 BCR-ABL | 9.21 ± 0.05 | 0.96 ± 0.02 |
| MYL-R | 8.17 ± 0.15 | 0.93 ± 0.11 |
| K562-IMR | 3.51 ± 0.12 | 0.70 ± 0.06 |
| KBM5-STI | 6.64 ± 0.18 | 0.80 ± 0.12 |
| Hepatocyte | 12.91 ± 2.73 | 20.10 ± 4.50 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hoshiko, T.; Kubota, Y.; Onodera, R.; Higashi, T.; Yokoo, M.; Motoyama, K.; Kimura, S. Folic Acid-Appended Hydroxypropyl-β-Cyclodextrin Exhibits Potent Antitumor Activity in Chronic Myeloid Leukemia Cells via Autophagic Cell Death. Cancers 2021, 13, 5413. https://doi.org/10.3390/cancers13215413
Hoshiko T, Kubota Y, Onodera R, Higashi T, Yokoo M, Motoyama K, Kimura S. Folic Acid-Appended Hydroxypropyl-β-Cyclodextrin Exhibits Potent Antitumor Activity in Chronic Myeloid Leukemia Cells via Autophagic Cell Death. Cancers. 2021; 13(21):5413. https://doi.org/10.3390/cancers13215413
Chicago/Turabian StyleHoshiko, Toshimi, Yasushi Kubota, Risako Onodera, Taishi Higashi, Masako Yokoo, Keiichi Motoyama, and Shinya Kimura. 2021. "Folic Acid-Appended Hydroxypropyl-β-Cyclodextrin Exhibits Potent Antitumor Activity in Chronic Myeloid Leukemia Cells via Autophagic Cell Death" Cancers 13, no. 21: 5413. https://doi.org/10.3390/cancers13215413