
Characterization of Genetic Heterogeneity in Recurrent Metastases of Renal Cell Carcinoma

 , , , ,
, , , , 
 , ,
, , 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Patient Cohort
2.2. Next Generation Sequencing (NGS)
2.3. Statistical Analyses
3. Results
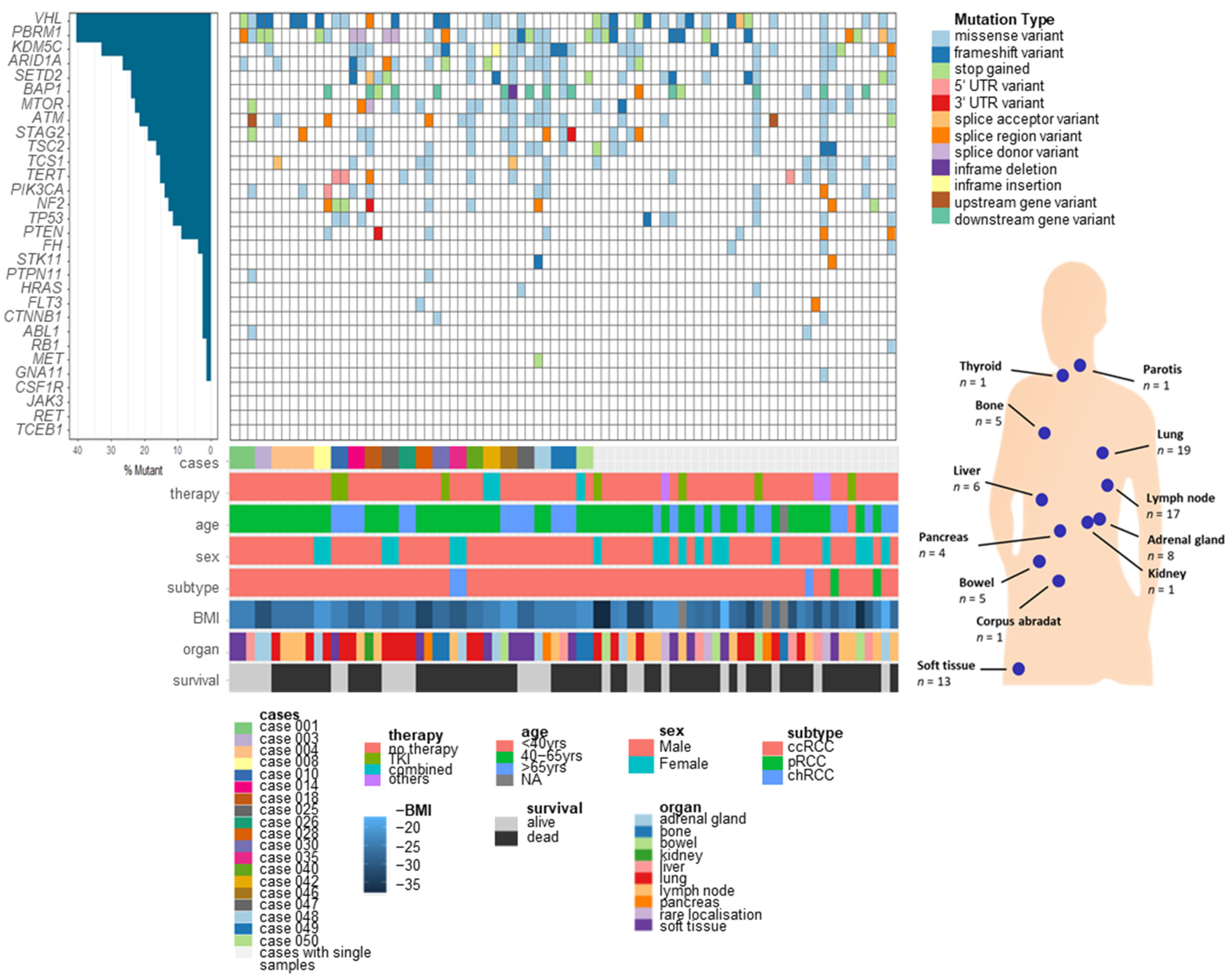
3.1. Somatic Variants of RCC Metastases in Different Organs
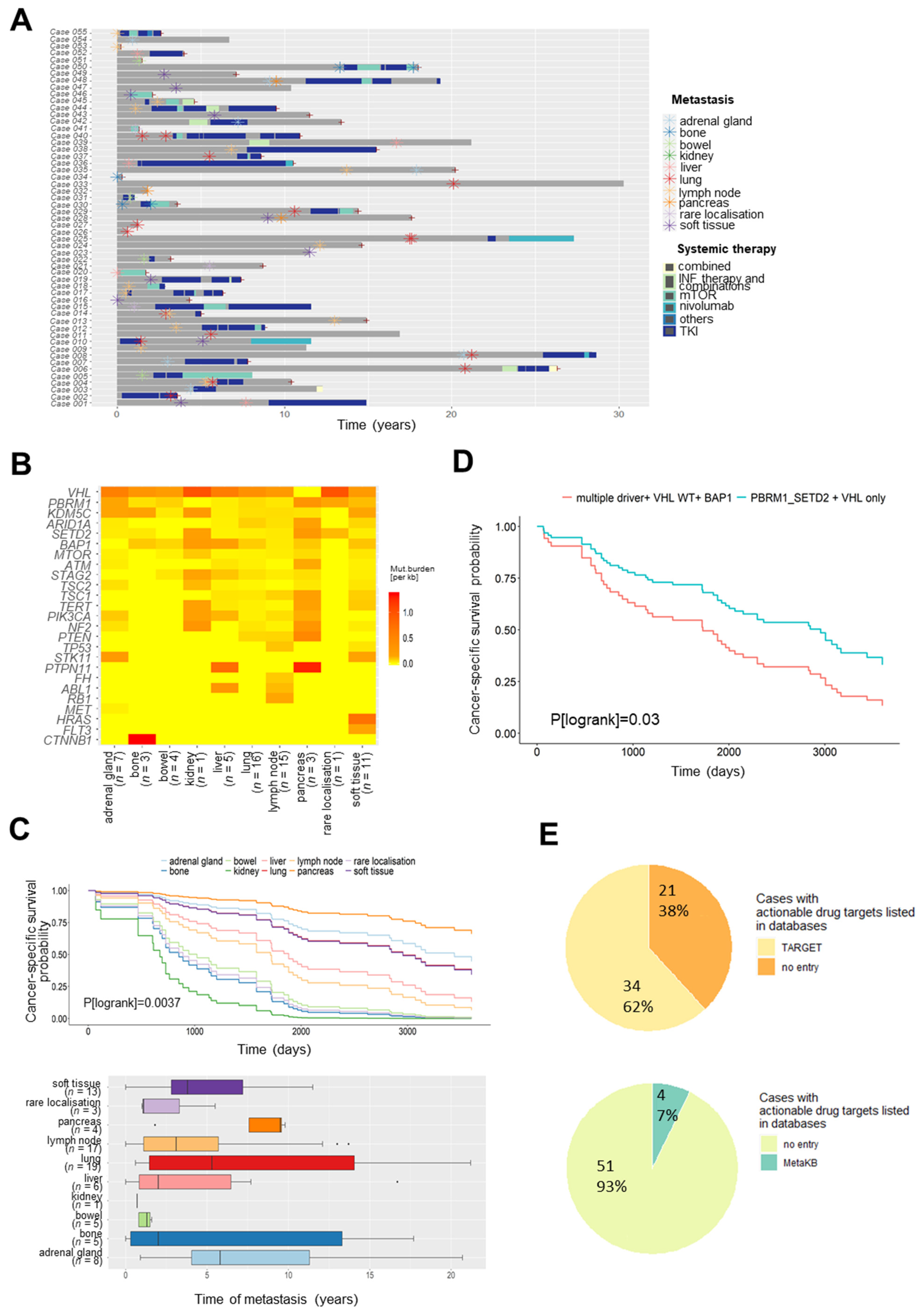
3.2. Somatic Variants in Metastases for Prediction of Survival and Personalized Therapy
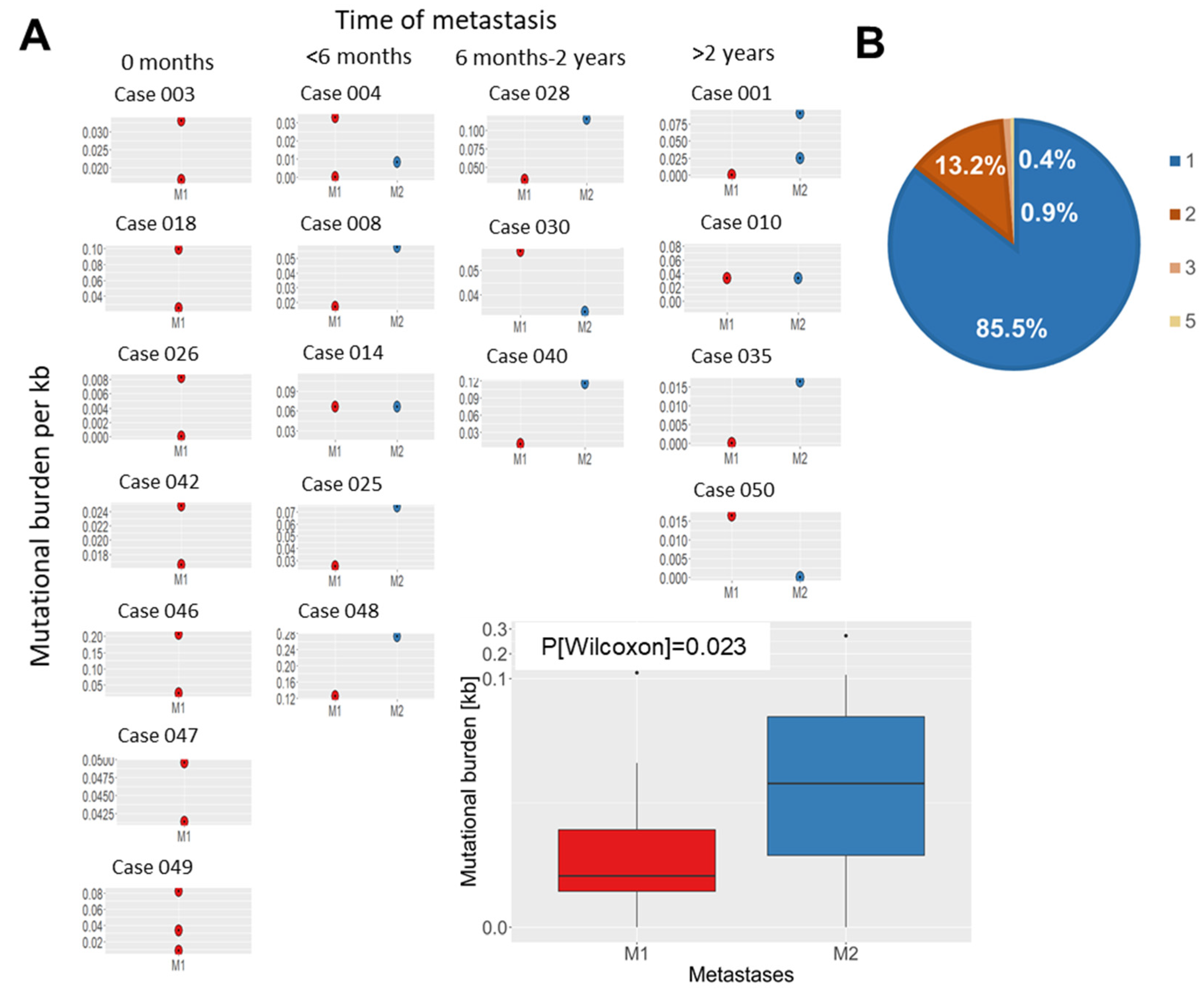
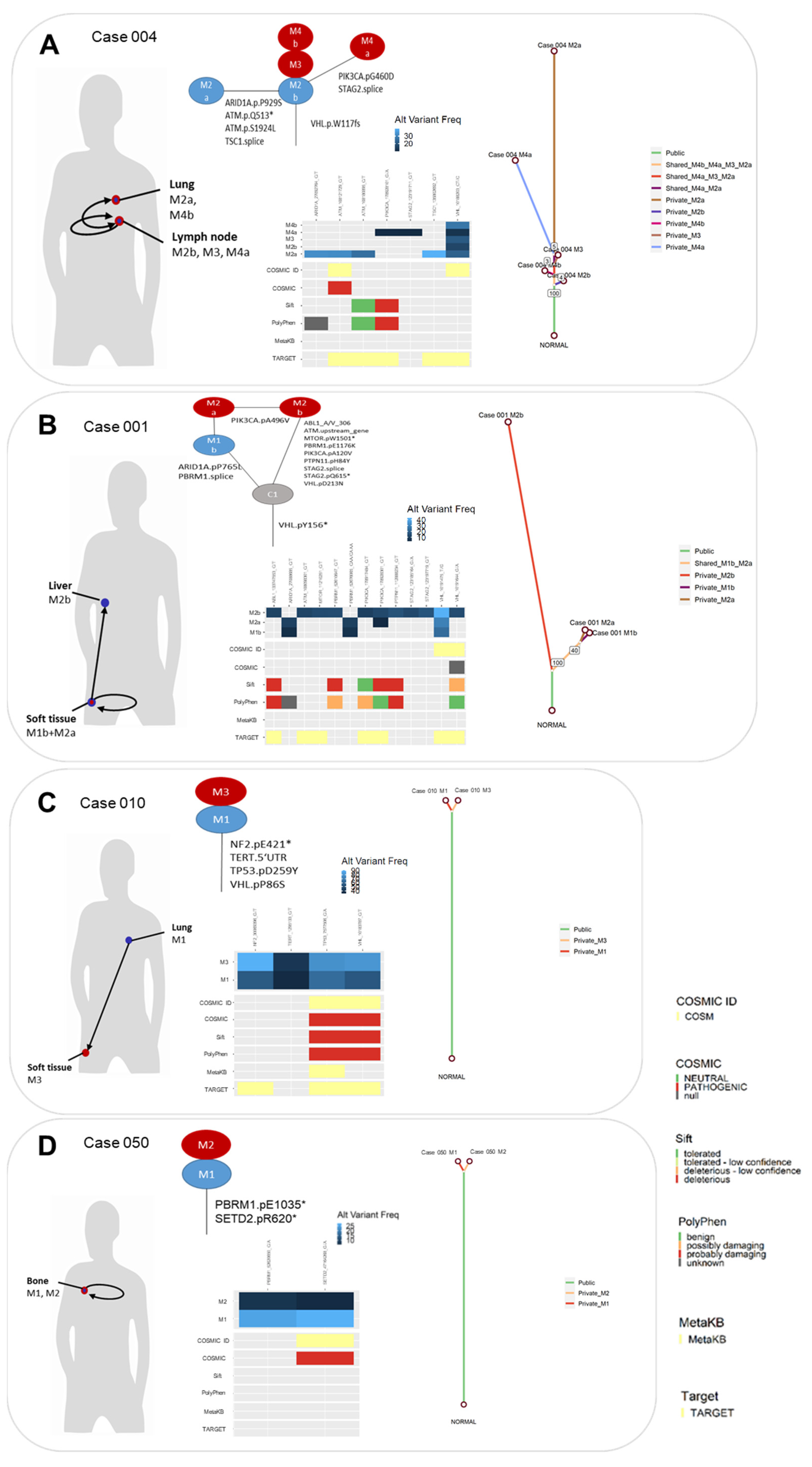
3.3. Evolution of Somatic Variants in Recurrent Metastases over Time and Therapeutic Course
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Capitanio, U.; Bensalah, K.; Bex, A.; Boorjian, S.A.; Bray, F.; Coleman, J.A.; Gore, J.L.; Sun, M.; Wood, C.G.; Russo, P. Epidemiology of Renal Cell Carcinoma. Eur. Urol. 2018, 75, 74–84. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, J.J.; Purdue, M.P.; Signoretti, S.; Swanton, C.; Albiges, L.; Schmidinger, M.; Heng, D.Y.C.; Larkin, J.M.; Ficarra, V. Renal Cell Carcinoma. Nat. Rev. Dis. Primers 2017, 3, 17009. [Google Scholar] [CrossRef] [PubMed]
- Scelo, G.; Larose, T.L. Epidemiology and Risk Factors for Kidney Cancer. J. Clin. Oncol. 2018, 36, 3574–3581. [Google Scholar] [CrossRef] [PubMed]
- McKay, R.R.; Bossé, D.; Choueiri, T.K. Evolving Systemic Treatment Landscape for Patients With Advanced Renal Cell Carcinoma. J. Clin. Oncol. 2018, 36, 3615–3623. [Google Scholar] [CrossRef]
- Bedke, J.; Albiges, L.; Capitanio, U.; Giles, R.H.; Hora, M.; Lam, T.B.; Ljungberg, B.; Marconi, L.; Klatte, T.; Volpe, A.; et al. Updated European Association of Urology Guidelines on Renal Cell Carcinoma: Nivolumab plus Cabozantinib Joins Immune Checkpoint Inhibition Combination Therapies for Treatment-naïve Metastatic Clear-Cell Renal Cell Carcinoma. Eur. Urol. 2021, 339–342. [Google Scholar] [CrossRef]
- The Cancer Genome Atlas Research Network. Comprehensive molecular characterization of clear cell renal cell carcinoma. Nature 2013, 499, 43–49. [Google Scholar] [CrossRef] [Green Version]
- Linehan, W.M.; Ricketts, C.J. The Cancer Genome Atlas of renal cell carcinoma: Findings and clinical implications. Nat. Rev. Urol. 2019, 16, 539–552. [Google Scholar] [CrossRef]
- Turajlic, S.; Xu, H.; Litchfield, K.; Rowan, A.; Horswell, S.; Chambers, T.; O’Brien, T.; Lopez, J.I.; Watkins, T.B.K.; Nicol, D.L.; et al. Deterministic Evolutionary Trajectories Influence Primary Tumor Growth: TRACERx Renal. Cell 2018, 173, 595–610. [Google Scholar] [CrossRef] [Green Version]
- Gerlinger, M.; Horswell, S.; Larkin, J.M.; Rowan, A.J.; Salm, M.P.; Varela, I.; Fisher, R.; McGranahan, N.; Matthews, N.; Santos, C.R.; et al. Genomic architecture and evolution of clear cell renal cell carcinomas defined by multiregion sequencing. Nat. Genet. 2014, 46, 225–233. [Google Scholar] [CrossRef]
- Turajlic, S.; Xu, H.; Litchfield, K.; Rowan, A.; Chambers, T.; Lopez, J.I.; Nicol, D.L.; O’Brien, T.; Larkin, J.M.; Horswell, S.; et al. Tracking Cancer Evolution Reveals Constrained Routes to Metastases: TRACERx Renal. Cell 2018, 173, 581–594.e12. [Google Scholar] [CrossRef] [Green Version]
- Ricketts, C.J.; Linehan, W.M. Multi-regional Sequencing Elucidates the Evolution of Clear Cell Renal Cell Carcinoma. Cell 2018, 173, 540–542. [Google Scholar] [CrossRef] [Green Version]
- Brooks, S.A.; Brannon, A.R.; Parker, J.S.; Fisher, J.C.; Sen, O.; Kattan, M.W.; Hakimi, A.A.; Hsieh, J.J.; Choueiri, T.K.; Tamboli, P.; et al. ClearCode34: A prognostic risk predictor for localized clear cell renal cell carcinoma. Eur. Urol. 2014, 66, 77–84. [Google Scholar] [CrossRef] [Green Version]
- Büttner, F.; Winter, S.; Rausch, S.; Hennenlotter, J.; Kruck, S.; Stenzl, A.; Scharpf, M.; Fend, F.; Agaimy, A.; Hartmann, A.; et al. Clinical utility of the S3-score for molecular prediction of outcome in non-metastatic and metastatic clear cell renal cell carcinoma. BMC Med. 2018, 16, 108. [Google Scholar] [CrossRef]
- Büttner, F.; Winter, S.; Rausch, S.; Reustle, A.; Kruck, S.; Junker, K.; Stenzl, A.; Agaimy, A.; Hartmann, A.; Bedke, J.; et al. Survival Prediction of Clear Cell Renal Cell Carcinoma Based on Gene Expression Similarity to the Proximal Tubule of the Nephron. Eur. Urol. 2015, 68, 1016–1020. [Google Scholar] [CrossRef]
- Graham, J.; Dudani, S.; Heng, D.Y.C. Prognostication in Kidney Cancer: Recent Advances and Future Directions. J. Clin. Oncol. 2018, 36, 3567–3573. [Google Scholar] [CrossRef]
- Kandabarau, S.; Leiz, J.; Krohn, K.; Winter, S.; Bedke, J.; Schwab, M.; Schaeffeler, E.; Edemir, B. Hypertonicity-Affected Genes Are Differentially Expressed in Clear Cell Renal Cell Carcinoma and Correlate with Cancer-Specific Survival. Cancers 2019, 12, 6. [Google Scholar] [CrossRef] [Green Version]
- Rini, B.; Goddard, A.; Knezevic, D.; Maddala, T.; Zhou, M.; Aydin, H.; Campbell, S.; Elson, P.; Koscielny, S.; Lopatin, M.; et al. A 16-gene assay to predict recurrence after surgery in localised renal cell carcinoma: Development and validation studies. Lancet Oncol. 2015, 16, 676–685. [Google Scholar] [CrossRef]
- Jonasch, E.; Walker, C.L.; Rathmell, W.K. Clear cell renal cell carcinoma ontogeny and mechanisms of lethality. Nat. Rev. Nephrol. 2021, 17, 245–261. [Google Scholar] [CrossRef]
- Dizman, N.; Philip, E.J.; Pal, S.K. Genomic profiling in renal cell carcinoma. Nat. Rev. Nephrol. 2020, 16, 435–451. [Google Scholar] [CrossRef]
- Lambert, A.W.; Pattabiraman, D.R.; Weinberg, R.A. Emerging Biological Principles of Metastasis. Cell 2017, 168, 670–691. [Google Scholar] [CrossRef] [Green Version]
- Yuan, S.; Norgard, R.J.; Stanger, B.Z. Cellular Plasticity in Cancer. Cancer Discov. 2019, 9, 837–851. [Google Scholar] [CrossRef] [Green Version]
- de Velasco, G.; Wankowicz, S.A.; Madison, R.; Ali, S.M.; Norton, C.K.; Duquette, A.; Ross, J.S.; Bossé, D.; Lalani, A.-K.A.; Miller, V.A.; et al. Targeted genomic landscape of metastases compared to primary tumours in clear cell metastatic renal cell carcinoma. Br. J. Cancer 2018, 118, 1238–1242. [Google Scholar] [CrossRef] [Green Version]
- Therneau, T.M.; Grambsch, P.M. Modeling Survival Data: Extending the Cox Model; Springer: New York, NY, USA, 2000. [Google Scholar]
- Kassambara, A.; Kosinski, M.; Biecek, P. Survminer: Drawing Survival Curves Using ‘ggplot2’. Available online: https://CRAN.R-project.org/package=survminer (accessed on 25 July 2020).
- Liu, M.; Chen, J.; Wang, X.; Wang, C.; Zhang, X.; Xie, Y.; Zuo, Z.; Ren, J.; Zhao, Q. MesKit: A tool kit for dissecting cancer evolution of multi-region tumor biopsies through somatic alterations. GigaScience 2021, 10, giab036. [Google Scholar] [CrossRef]
- Wagner, A.H.; Walsh, B.; Mayfield, G.; Tamborero, D.; Sonkin, D.; Krysiak, K.; Deu-Pons, J.; Duren, R.P.; Gao, J.; McMurry, J.; et al. A harmonized meta-knowledgebase of clinical interpretations of somatic genomic variants in cancer. Nat. Genet. 2020, 52, 448–457. [Google Scholar] [CrossRef] [Green Version]
- Hertz, D.L.; McLeod, H.L. Using pharmacogene polymorphism panels to detect germline pharmacodynamic markers in oncology. Clin. Cancer Res. 2014, 20, 2530–2540. [Google Scholar] [CrossRef] [Green Version]
- Neul, C.; Schaeffeler, E.; Sparreboom, A.; Laufer, S.A.; Schwab, M.; Nies, A.T. Impact of Membrane Drug Transporters on Resistance to Small-Molecule Tyrosine Kinase Inhibitors. Trends Pharmacol. Sci. 2016, 37, 904–932. [Google Scholar] [CrossRef]
- Singla, N.; Xie, Z.; Zhang, Z.; Gao, M.; Yousuf, Q.; Onabolu, O.; McKenzie, T.; Tcheuyap, V.T.; Ma, Y.; Choi, J.; et al. Pancreatic tropism of metastatic renal cell carcinoma. JCI Insight 2020, 5, e134564. [Google Scholar] [CrossRef] [Green Version]
- Tippu, Z.; Au, L.; Turajlic, S. Evolution of Renal Cell Carcinoma. Eur. Urol. Focus 2021, 7, 148–151. [Google Scholar] [CrossRef]
- Vendramin, R.; Litchfield, K.; Swanton, C. Cancer evolution: Darwin and beyond. EMBO J 2021, 40, e108389. [Google Scholar] [CrossRef]
- Nassar, A.H.; Hamieh, L.; Gray, K.P.; Thorner, A.R.; Fay, A.P.; Lasseter, K.D.; Abou Alaiwi, S.; Nuzzo, P.V.; Flippot, R.; Krajewski, K.M.; et al. Mutations and Response to Rapalogs in Patients with Metastatic Renal Cell Carcinoma. Mol. Cancer Ther. 2020, 19, 690–696. [Google Scholar] [CrossRef]
- Diekstra, M.H.M.; Swen, J.J.; Boven, E.; Castellano, D.E.; Gelderblom, H.; Mathijssen, R.H.J.; Rodriguez-Antona, C.; Garcia-Donas, J.; Rini, B.I.; Guchelaar, H.-J. CYP3A5 and ABCB1 polymorphisms as predictors for sunitinib outcome in metastatic renal cell carcinoma. Eur. Urol. 2015, 68, 621–629. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics of Patients and Primary Tumours (n = 56) | Levels/Summary Statistics | No. | % |
|---|---|---|---|
| Sex | male | 40 | 71.4 |
| female | 16 | 28.6 | |
| Age (yrs) at diagnosis of primary RCC | median (range) | 60.6 (29.2–77.5) | |
| T | 1 | 12 | 21.4 |
| 2 | 8 | 14.3 | |
| 3 | 29 | 51.8 | |
| 4 | 2 | 3.6 | |
| na | 5 | 8.9 | |
| N | 0 | 44 | 78.6 |
| 1 | 3 | 5.4 | |
| 2 | 3 | 5.4 | |
| na | 6 | 10.7 | |
| M | 0 | 39 | 69.6 |
| 1 | 12 | 21.4 | |
| na | 5 | 8.9 | |
| G | 1 | 6 | 10.7 |
| 2 | 27 | 48.2 | |
| 3 | 16 | 28.6 | |
| na | 7 | 12.5 | |
| Follow-up time (yrs) from date of diagnosis of primary RCC | median (range) | 9.15 (0.2–30.3) | |
| Cancer-related death | no | 19 | 33.9 |
| yes | 37 | 66.1 | |
| Characteristics of Metastasis/Local Recurrence Specimens (n = 81) | Levels/Summary Statistics | No. | % |
| metastatic site | Organ Group | ||
| adrenal gland | 8 | 9.9 | |
| bone | 5 | 6.2 | |
| bowel | 5 | 6.2 | |
| local recurrence (kidney) | 1 | 1.2 | |
| liver | 6 | 7.4 | |
| lung | 19 | 23.5 | |
| lymph node | 17 | 21.0 | |
| pancreas | 4 | 4.9 | |
| rare localisation | 3 | 3.7 | |
| soft tissue | 13 | 16.0 | |
| Age (yrs) at metastasis resection | median (range) | 66.9 (31.6–80.6) | |
| Follow-up time (yrs) from date of metastasis resection | median (range) | 5 (0–11.3) | |
| Systemic therapy before metastasis resection | no | 68 | 84.0 |
| yes | 13 | 16.0 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sauter-Meyerhoff, C.; Bohnert, R.; Mazzola, P.; Stühler, V.; Kandabarau, S.; Büttner, F.A.; Winter, S.; Herrmann, L.; Rausch, S.; Hennenlotter, J.; et al. Characterization of Genetic Heterogeneity in Recurrent Metastases of Renal Cell Carcinoma. Cancers 2021, 13, 6221. https://doi.org/10.3390/cancers13246221
Sauter-Meyerhoff C, Bohnert R, Mazzola P, Stühler V, Kandabarau S, Büttner FA, Winter S, Herrmann L, Rausch S, Hennenlotter J, et al. Characterization of Genetic Heterogeneity in Recurrent Metastases of Renal Cell Carcinoma. Cancers. 2021; 13(24):6221. https://doi.org/10.3390/cancers13246221
Chicago/Turabian StyleSauter-Meyerhoff, Carolin, Regina Bohnert, Pascale Mazzola, Viktoria Stühler, Siarhei Kandabarau, Florian A. Büttner, Stefan Winter, Lisa Herrmann, Steffen Rausch, Jörg Hennenlotter, and et al. 2021. "Characterization of Genetic Heterogeneity in Recurrent Metastases of Renal Cell Carcinoma" Cancers 13, no. 24: 6221. https://doi.org/10.3390/cancers13246221