
Sex Differences in Lung Cancer


1
Department of Human and Molecular Genetics, VCU Institute of Molecular Medicine, Massey Cancer Center, VCU School of Medicine, Richmond, VA 23298, USA
2
Department of Biology, Virginia State University, Petersburg, VA 23806, USA
3
Department of Internal Medicine, Division of Pulmonary Disease and Critical Care Medicine, VCU School of Medicine, Richmond, VA 23298, USA
*
Author to whom correspondence should be addressed.
Cancers 2023, 15(12), 3111; https://doi.org/10.3390/cancers15123111
Submission received: 23 May 2023
/
Revised: 2 June 2023
/
Accepted: 5 June 2023
/
Published: 8 June 2023
Abstract
:Simple Summary
Lung cancer remains the most commonly diagnosed cancer in the United States, only behind sex-specific cancers such as breast and prostate cancer. While not a sex-specific cancer, lung cancer exhibits sex-specific trends. Males are generally at a higher lifetime risk of developing lung cancer and have a higher mortality than females. There are also differences in therapeutic response between the sexes. As lung cancer is a complex disease, this difference is likely a combination of environmental factors, such as environmental exposures, diet, and smoking status, with inherent biological differences, such as the contribution of sex hormones and differences in immune responses. This narrative review focuses specifically on these biological differences and their contributions to this difference. Gaining a better understanding of the biological reasons behind this sex difference could lead to better treatment and screening decisions in the clinic that take the biological sex of the patient into consideration.
Abstract
Sex disparities in the incidence and mortality of lung cancer have been observed since cancer statistics have been recorded. Social and economic differences contribute to sex disparities in lung cancer incidence and mortality, but evidence suggests that there are also underlying biological differences that contribute to the disparity. This review summarizes biological differences which could contribute to the sex disparity. Sex hormones and other biologically active molecules, tumor cell genetic differences, and differences in the immune system and its response to lung cancer are highlighted. How some of these differences contribute to disparities in the response to therapies, including cytotoxic, targeted, and immuno-therapies, is also discussed. We end the study with a discussion of our perceived future directions to identify the key biological differences which could contribute to sex disparities in lung cancer and how these differences could be therapeutically leveraged to personalize lung cancer treatment to the individual sexes.
1. Introduction
Despite significant progress in the diagnosis and therapeutic management of cancers, significant disparities exist, with males suffering from a higher incidence and mortality compared to females worldwide [1,2]. This long-standing disparity in outcomes [3] may be driven by both social determinants and fundamental biological characteristics of the disease. Outside cancers of the reproductive system, there are several cancers including lung, melanoma, urinary tract, and glioblastoma that demonstrate great sex-specific trends in clinical presentation, progression, and mortality. These sex differences, and their implications for cancer treatment strategies, have been a focus of a European Society for Medical Oncology multi-disciplinary workshop highlighting their importance to the cancer community [4]. Lung cancer is the leading cause of cancer death [1]; thus, there is an urgent need to elucidate the underpinnings of disease initiation and progression. As such, a better understanding of the contributing clinical, biological, epigenetic, and genetic drivers of sex differences in lung cancer has the potential to inform risk stratification and screening for early disease as well as treatment strategies that take sex into consideration. In this review, we will describe potential biological bases for the observed sex differences in lung cancer and how such biology could be leveraged to tailor treatment of subjects with lung cancer.
Sex Differences in Lung Cancer—A Clinical Perspective
In the past, males were disproportionally diagnosed with lung cancer, though this gap has been closing and reversing in certain populations in recent years [5,6,7]. Later diagnoses with more advanced stages of disease are more likely to occur in males [5]. This results in a worse prognosis in males with lung cancer and higher mortality than women despite advances in therapy development, and declining mortality for all cancers (https://www.cdc.gov/cancer/, accessed on 1 May 2023). Sex differences in the survival and mortality of early stage lung cancer are reduced with low-dose computed tomography (LDCT) screening [8,9]; however, current eligibility criteria for screening do not include sex; rather, these are focused on age and smoking history [10]. Lung cancer is a complex disease, and as such, the drivers of the observed sex differences are likely to be multifactorial, including a combination of multiple environmental and biological factors. These factors can include sex-differences in behavior towards the healthcare system and environmental exposures, smoking, diet, sex hormone contributions, and differences in immune responses. Controlling for environmental contributions, age, and smoking history, men remain more susceptible to lung cancer, suggesting that underlying fundamental biological characteristics across sex may be contributing to disease [11]. Despite the confirmation of this sex difference, the biological drivers of this difference remain largely unknown.
2. Sex Hormones
Sex hormones and their signaling pathways have long been implicated in the pathogenesis of cancers [12]. Sex hormones with known roles in cancer include prolactin (PRL), luteinizing hormone (LH), follicle-stimulating hormone (FSH), gonadotropin-releasing hormone (GNRH), estrogen (E2), progesterone (P) and testosterone (T) [13]. Many of these hormones have well-documented roles in the cancer biology of reproductive organs including ovaries, endometrium, prostate, and breast cancer [14]. Several sex hormones are produced by lung cancers ectopically [15,16], which contribute to the clinical spectrum of disease (Table 1) [17]. Of these hormones, only exogenous E2, P and T have been documented to regulate lung cancer biology, and their role in the disease remains controversial in the literature.
2.1. Estrogen
Estradiol (E2) is the active form of estrogen that has been shown in vitro to induce proliferation of lung cancer cells [48,49]. ERα and ERβ are encoded by the ESR1 and ESR2 genes, respectively, and expressed as full-length proteins with several potential transcript variants [50]. ERβ is normally expressed in lung epithelium and lung fibroblasts, and is present in lung cancer cells. Conversely, ERα is typically not found, or expressed at very low levels, in lung tissues. ERα expression is known to be regulated epigenetically in lung cancer through CpG methylation [51]. One study showed significant methylation in lung cancer tissue in contrast to no methylation in normal tissue [52]. CpG methylation has also been observed in lung cancer cell lines and mouse models of lung cancer [52]. The role of ERα is somewhat controversial; despite overall lower levels of expression in some lung cancers, some studies have shown cytoplasmic ERα in non-small cell lung cancer (NSCLC), potentially due to a transcript variant [53,54].
Signaling through these receptors is initiated through E2 binding to the ER. Studies have shown that, in addition to circulating estrogen and exogenous estrogen from hormone replacement therapy (HRT), E2 can be produced by NSCLC cells via aromatase activity [18]. One study demonstrated that aromatase activity was significantly higher in adenocarcinoma, adenosquamous carcinomas, and bronchoalveolar carcinomas than in normal lung tissues [48]. An additional study demonstrated that lower levels of aromatase expression predicted a better overall survival (OS) in women over 65, as well as in younger women with no smoking history [55]. In addition to their own signaling pathways, ERs can also interact with other cell-surface receptors including epithelial growth factor receptor (EGFR), caveolin, and flotillin [18]. Interaction with these receptors can also promote cell proliferation. ERβ expression has, in general, not been associated with any specific grade of cancer, but has been associated with improved OS. In contrast, ERα expression in the cytoplasm is associated with a higher tumor grade and poorer OS [56,57].
Both ERs and aromatase have been identified as potential therapeutic targets in lung cancer. Fulvestrant, an ER competitive inhibitor, significantly reduced tumor volume of NCI-H23 NSCLC cells in a flank model of ovariectomized nude mice [57]. Erlotinib and gefitinib, both EGFR tyrosine kinase inhibitors, were incapable of controlling tumor growth; however, when combined with fulvestrant, these were superior in controlling tumor growth compared to fulvestrant alone [18]. Aromatase inhibitors such as anastrozole and exemestane alone or in combination with traditional chemotherapies also show significant tumor growth control in lung cancer [48,58].
The role of the estrogen signaling pathway in lung cancer biology is controversial, and as a result, its role in lung cancer remains unclear. Estrogens play important physiological roles in both males and females, though their overall abundance is higher in females [59]. Outside of their role in reproductive signaling, estrogens facilitate a variety of biological processes, from protein and lipid synthesis to modulating immune cell populations [60]. In the context of cancer, the involvement of estrogens is tied to estrogen receptor (ER) status. For example, breast cancers are often ER-positive, and frequently depend on signals from estrogens to grow [61,62]. Likewise, there are suggestions that lung cancers with cytoplasmic, in comparison to nuclear, localization of ER may have a worse prognosis; however, this was not observed universally in all studies [63].
It is unclear if estrogens promote or hinder lung cancer. Several studies have shown a higher risk of lung cancer in women taking HRT [19,20,21], but just as many studies show HRT or oral contraceptives having no effect on overall risk [22,64,65]. Additional studies investigating the interplay between estrogen levels and smoking show a higher risk for developing lung cancer among smokers on HRT [66,67]. Another retrospective study showed that survival was higher in women who were not on HRT at the time of diagnosis, especially in women who had a prior history of smoking [22]. Consistent with estrogens promoting lung cancer, there was a measurable increase in lung cancers in males given estrogens to treat heart disease leading to the termination of the clinical trial [68]. Estrogens can also modulate the effects of other cancer-causing factors, such as genetic mutations due to smoking [69]. The cancer-promoting effects of estrogens are further supported by common ER expression in lung cancers, the ability of estrogen to directly, or indirectly through aromatase expression, stimulate lung cancer growth cell culture studies [49]. These studies strongly suggest that estrogen promotes lung cancer growth in clinical practice and in experimental models.
In contrast, there have been several studies suggesting that there is a protective benefit against lung cancer in women who have not had an oophorectomy. A study of nearly 1000 women demonstrated that women who had a non-natural menopause (i.e., an oophorectomy for medical reasons) were at a much higher risk for developing lung cancer than those who went through a natural menopause [70]. Similarly, the same study also showed that women who entered menopause at a younger age than 45 were at a higher risk of developing lung cancer than their older counterparts [70]. Additionally, a study analyzing patients from the Surveillance, Epidemiology, and End Results (SEER) database showed that women with lung cancer who were premenopausal typically had a more advanced disease at the time of diagnosis and had a higher rate of adenocarcinoma than their postmenopausal counterparts [71]. The same study showed that premenopausal women had similar mortality rates to males in the same age range with lung cancer, but that older postmenopausal women had lower mortality rates than their age-matched males. In addition to estrogens, premenopausal ovaries are known to regulate a variety of bioactive molecules, such as progesterone, testosterone, follicular stimulating hormone (FSH), luteinizing hormone (LH), inhibin b, and anti-mullerian hormone (AMH) [72]. In addition to these, leptin, a dual hormone and cytokine, may be associated with poor outcomes in lung adenocarcinoma and squamous cell carcinoma, though more research needs to be carried out into its potential role [44]. The contrasting effects of ectopic estrogen treatments (i.e., HRT) which generally promote lung cancer, and the effects of sex hormone loss resulting from pre-menopause or ovariectomy, which increase the lung cancer risk of women, could be explained by changes in sex hormones or other bioactive molecules other than estrogen such as progesterone.
2.2. Progesterone
Progesterone, while traditionally described as a female sex hormone, is present in both males and females as an important precursor molecule for other endogenous steroids. Progesterone and its role in lung cancer has been less extensively studied compared to estrogen. As with estrogen and ERs, progesterone can be produced by NSCLC and the progesterone receptors (PRs) are expressed in NSCLC [33,73,74,75]. Analysis of PR expression in the patients’ tumor samples did not show any correlation with age, menopausal state, ER positivity, or p53 positivity; however, it was more often found in females with early stage disease and in poorly differentiated NSCLC [33]. Additionally, patients with PR-positive lung cancer had a better OS than those with PR-negative lung cancer [76]. In animal studies, progesterone was able to inhibit the growth of lung cancer cells in a flank model in nude mice, and this inhibition was associated with decreased expression of proliferation marker Ki-67, cell cycle regulators cyclin A and E, as well as an increase in cell cycle [33] (pp. 21, 27). However, studies by other groups show no correlation between PR expression and OS [33,75]. Estrogens and progesterone can also work together, as ER signaling can upregulate PR expression [77]. Despite the disagreement in the literature, studies examining the role of progesterone and PR signaling are ongoing and could provide important information and insights into the role of hormones in lung cancer.
2.3. Testosterone
In males, androgen testosterone is the primary sex hormone, and research into its role in cancer has been primarily limited to prostate cancer. Elevated testosterone has been associated with increased lung cancer risk in males in some studies [37,38]. However, other studies did not corroborate a correlation between testosterone and lung cancer risk in men [78]. One study in a mutant Kras/TP53 mouse showed that males that received E2 required a much lower dose of Adeno-Cre in order to form tumors, suggesting that androgens might potentiate the cancer-promoting effects of estrogen or progesterone [39]. In women, the ovaries and adrenal glands metabolize progesterone into testosterone, leading to the possibility that the high levels of testosterone in men, and in women on HRT, could contribute to the observed sex difference in tumor development and progression [65].
3. Genetic Factors
Key genetic differences have been identified in lung cancers from men and women. Women more frequently have lung cancers with a driver mutation such as EGFR, ALK, or KRAS [79,80,81]. Additionally, multiple studies have demonstrated differences in key DNA damage repair pathways in the lung cancers from men compared to women. For example, lung cancers in women often have mutations in p53 and female-associated polymorphisms in the cytochrome P450 gene that affect DNA damage repair efficiency [82]. Female smokers have a significantly higher level of aromatic/hydrophobic DNA adducts in non-tumor lung tissue and have higher expression of CYP1A1 in lung tissue compared to their male counterparts [83]. Higher levels of CYP1A1 in females lead to a greater metabolism of polycyclic aromatic hydrocarbons (PAHs) from cigarette smoke into carcinogenic intermediates [83]. Higher levels of polycyclic aromatic hydrocarbon DNA adducts were also found in female smokers compared to male smokers, despite there being lower levels of tobacco carcinogens found in females than males [83]. The increased sensitivity of female smokers to DNA damage could be due, in part, to estrogen signaling and its role in cell proliferation [52]. Additionally, males and females metabolize nicotine and other tobacco carcinogens differently [84]. In general, females have more efficient nicotine clearance due to greater CYP2A6 activity, which can be attributable to higher estrogen levels [85]. Additional studies have focused on the differences between males and females in the expression of the X-linked gastrin-releasing peptide receptor (GRPR), which can encourage cell proliferation. One study found that GRPR was more highly expressed in female than male non-smokers, and that in smokers, it was found at lower levels of tobacco exposure in women than in men, suggesting that female smokers’ two copies of GRPR could contribute to lung cancer susceptibility [86,87].
A detailed meta-analysis of the TCGA datasets concluded that lung adenocarcinoma (LUAD) and lung squamous cell carcinoma (LUSC) have an extensive sex disparity in gene signatures (expression, methylation status and copy number changes) [88]. Sex disparity in mutations in LUAD include a female bias for mutations in MED12, F8, DMD, FAM47A, ABCB5 and male bias for mutations in RBM10, COL21A1, ZNF521, CNTN5, SMG1 and STK11. The significance of these findings is unknown because the products of these genes have a variety of functions, including, but not limited to, transcription regulation, clotting, muscle function, membrane transport. Several significant somatic copy number amplification disparities were observed in LUSC, including 17q11.2, 4q22.1 and a deletion bias at 15q12 to females. Males have several amplifications, including 6q12, 8p11.23, 12p13.33, 17q25.1, 20q11.21, 20q11.21 and Xq28, and a deletion bias at 18q23. Gene set enrichment analysis (GSEA) in LUAD discovers gene sets annotated as androgen response, E2F targets, fatty acid metabolism, G2M checkpoint, glycolysis, Myc targets, and the unfolded protein response as being significantly enriched in females over males. LUSC observes similar enrichments in females for E2F targets, G2M checkpoint and Myc targets. The methylation status of genes in LUSC and LUAD shows a female bias and includes categories annotated as interferon alpha response, TGFβ and TNFα signaling, and apoptosis.
4. Environmental Factors and Exposures
4.1. Smoking
Cigarette smoking is one of the greatest risk factors for the development of lung cancer. In the past, males were more likely to be smokers than women, though this has decreased in recent years, which is coincident with reductions in smoking rates overall [89,90]. Within the smoking population, males are more likely to develop lung cancer than women, though some studies have suggested that the disease development is different in male smokers compared to female smokers [90,91]. Other studies have shown tobacco exposure as having no effect on risk differences between males and females, possibly due to differences in population sampling and how tobacco exposure is characterized (i.e., inhalation depth, type of cigarette, and tar content) [92]. Concurrent with smoking is secondhand smoke exposure, although there is great difficulty in measuring and characterizing secondhand smoke exposure. One meta-analysis showed that women with smoking spouses had an increased risk of developing lung cancer [93]. However, another study from the UK Million Women Study did not find a link between secondhand smoke exposure and lung cancer development [94].
4.2. Additional Environmental Factors
Asbestos, while commonly associated with mesothelioma, also contributes to lung cancer development. Occupational exposures leading to carcinogenesis are more often seen in men, but one study has suggested that non-occupational exposures can put women at a higher risk than men [95]. Many of the studies involving asbestos exposure primarily focus on males. Infections are another source of risk for the development of lung cancer. One study showed that, in a non-smoking population, women were more likely to have HPV-positive lung cancer than men, with follow-up studies showing that women with a history of HPV have increased odds for developing lung cancer as compared to men [96,97].
5. Immune Responses
The immune response to lung cancer varies between men and women. A meta-analysis of several publicly available lung cancer gene expression datasets shows that women have increased gene signatures consistent with an acute inflammatory response compared to men [98]. Additional studies of similar datasets showed elevated levels of T-cell dysfunction, inhibitory immune checkpoint molecules, and immune-suppressive cells (MDSCs, Treg) in NSCLC tumors in women compared to men [99]. Single-cell sequencing analysis has shown that tumor-associated macrophages (TAMs) have elevated immunogenicity (elevated IFNγ-producing and antigen-presenting) and are assumed to have greater antitumor activity in cells from tumors of female patients [100]. In male-derived tumor samples, TAMs had gene signatures consistent with being more immunosuppressive.
Sex differences in lung cancer, and many other cancers with a sex bias, could be in part because males and females show marked differences in their immune systems. For example, females have been shown to have immune gene set enrichments as compared to males in the context of lung cancer [101]. The underlying drivers for these differences remain largely unknown. In general, females tend to have an overall more robust innate and adaptive immune response than males [32]. Females also typically harbor higher CD4+ T cell counts and produce higher amounts of IFNγ than males in response to infections [102]. While this greater response allows for quicker clearance of infections and development of immunity, it also places females at a greater lifetime risk for the development of autoimmune diseases [103]. In addition to differences in immune responses, there are also differences in circulating immune cell populations between males and females, including greater populations of CD4+ T cells and B cells in females, and these differences can shift based on age and race [104,105]. One potential explanation for this difference is the location of multiple genes related to immune function on the X chromosome [106]. Genes that encode for the IL-2 receptor gamma subunit (IL2Rγ), Toll-like receptor (TLR)-7, TLR-8, CD40L, and the forkhead box P3 (FoxP3) are all located on the X chromosome. Because of the higher immune system activity in females, cancer cells in females are often more efficient at evading the immune system and have undergone a stringent immunoediting process to avoid detection by the immune system [107].
Sex hormones are known to have effects on immune responses and cell populations. Estrogen has been shown to have a wide range of effects, from increasing NK cell cytotoxicity in vitro but decreasing the secretion of granzyme B by NK cells in vivo [23,24], to inducing changes in T helper 1/2 (TH1/TH2) responses [25]. E2 increases the production of human peripheral blood mononuclear cell immunoglobulins via the release of IL-10 [26]. Estrogen has been shown to increase the expression of CD22, SHP-1, and Bcl-2, all of which are important for B cell survival and proliferation [27]. Estrogen levels also have direct effects on the T regulatory population size, which, in turn, can affect T cell activity and proliferation [28]. The CC chemokine receptors CCR5 and CCR1 on CD4+ T cells can be stimulated by estrogen [28]. Estrogen can also impact the differentiation and activation of dendritic cells via IFN regulatory factor (IRF)-4 production in myeloid progenitor cells [29].
Estrogen signaling is important in a variety of immune cells, with non-classical ER signaling interacting with the estrogen response elements (EREs) transcription factors NFkB, SP-1, and AP-1 [30,31]. A significant portion of activated genes in female T cells have EREs in their promoters, underlining the important role of estrogen in T cell responses [31]. PD-L1 expression can be modulated by estrogen and by several X-linked microRNAs [32]. Progesterone’s effects on the immune system are less convoluted than estrogen’s; progesterone has an overall anti-inflammatory effect, with decreases in IFNy production in NK cells and inducible nitric oxide synthase (iNOS) in macrophages [34,35,36]. Androgens, such as testosterone, also work to suppress immune cell activity and are anti-inflammatory [108,109]. Testosterone has been shown to reduce immunoglobulin production and reduces the production of IL-6, as well as generally suppressing immune responses [40,41,42]. FSH has been shown to stimulate TNF production in bone marrow granulocytes and macrophages [43]. Leptin, which is generally higher in females, can affect the survival and activation of B and T cells, and it can drive IL-2 and IFNγ production [45]. Prolactin also has effects on the immune system; it has been shown to promote CD4+ and CD8+ T cell differentiation, the release of IFNγ from NK cells, and macrophage activation via prolactin receptor activity [47]. Studies have been performed to characterize the role of the immune system in lung cancer. Lung tumors have been shown to have very low antigen presentation and low co-stimulatory molecule expression, allowing them to escape detection by the immune system [110,111].
Smoking can have significant effects on the immune system. Cigarette smoke can induce MAPK signaling, which can affect NFkB pathway activation [112]. Smoking can reduce the activity of neutrophils and the efficiency of APCs, overall T cell activity, and the amount of circulating immunoglobulins [113,114]. However, studies into sex-specific effects of smoking on the immune system have yet to be performed. These studies could provide key insights into immune cell populations that may contribute to the observed sex differences in lung cancer development and progression.
6. Differences in Response to Therapy
6.1. Immunotherapies
Immunotherapies, such as immune checkpoint blockade (ICB) therapies, are now the standard of care for a subset of subjects with lung cancer. However, the effects of sex on these therapies in lung cancer patients is an ongoing question in the cancer community. Multiple meta-analyses have been performed on clinical trial data for ICBs, which included lung cancer cohorts; however, many of these analyses fail to take into consideration study heterogeneity and different cancer types. One meta-analysis performed by Botticelli and Coll, which focused on anti-CTLA-4, anti-PD-1, and anti-PD-L1 therapies, showed no significant benefit with immunotherapies regarding OS or progression-free survival (PFS) in males vs. females, including those with lung cancer; however, this study did not account for heterogeneity between trials. This study did not demonstrate any benefits to PDL1 immunotherapy across sex in lung cancer [115]. Wu and Coll investigated the efficacy of CTLA-4 and PD-1 inhibitors vs. other therapies and demonstrated a better PFS and OS in males vs. females; however, these findings were not significant in the NSCLC cohort of the analysis [107]. A third meta-analysis focusing only on NSCLC patients with anti-PD-1 inhibitors (pembrolizumab or nivolumab) vs. chemotherapy showed a clear benefit of anti-PD1 inhibitors (pembrolizumab) in males over females [116]. In the NSCLC cohort of another meta-analysis carried out by Grassadonia et al., PFS was higher in immunotherapy-treated males than in females, and anti-CTLA-4 treatment was associated with longer OS in males [117]. Similar results were also observed Laing et al. in a more recent meta-analysis [118]. In contrast, a meta-analysis carried out by Conforti et al. showed longer OS in women than in men treated with ICBs; however, this study excluded a large number of female patients from their final analysis, making interpretation of the results difficult [119]. Multiple other meta-analyses show no benefit of ICBs as monotherapies or as combinational therapies in males vs. females with lung cancer [120,121]. The lack of agreement on the effects of immunotherapies on males vs. females suggests a need for continued research into this issue and better powered studies to elucidate potential sex differences. Gaining an understanding of which therapies might be more beneficial to a certain sex has implications for clinical practice and choosing adequate therapies for each patient.
6.2. Chemotherapies
Some studies have shown that males and females have different responses to chemotherapies, whereas other have not documented any differences [122]. In general, women have more adverse events to therapy than men [123]. In addition, there are well-documented differences in drug kinetics, clearance, and toxicity between men and women [124]. Relevant to lung cancer, platinum-based chemotherapy clinical trials in NSCLC found that women responded better to these chemotherapies than males and had a significantly better OS [125]. However, several studies did not observe these differences with platinum-based compounds [126,127]. In addition, several meta-analyses found that chemotherapy effectiveness in the adjuvant and neoadjuvant setting did not find any sex-difference in the final outcomes [128,129], nor any sex difference in concurrent platinum-based therapy compared to radiotherapy in lung cancer patients [130]. Female patients with lung carcinoma who received paclitaxel combined with carboplatin showed longer PFS than males, which could be potentially explained by a lower activity of DNA damage repair mechanisms in tumors found in females compared to those found in males [131]. Additional studies on combinational therapies with cisplatin have all shown significantly better responses in females over males with NSCLC [132,133]. As such, further investigating sex disparities in lung cancer therapeutic responses could close our knowledge gap as to why they occur and help develop sex-specific treatment strategies to mitigate the detrimental effects of these differences.
6.3. Targeted Therapies
Previously, women were shown to have better responses to EGFR inhibitors than males [134]. A recent trial showed a trend of females receiving osimertinib (a third-generation epidermal growth factor receptor tyrosine kinase inhibitor) having improved survival over males; however, this result was not statistically significant [135]. A similar result was observed in a trial studying the use of ALK inhibitors alectinib over crizotinib in patients harboring ALK-mutated NSCLC. Women trended towards improved survival over males, but the finding was not statistically significant [136]. VEGF blockade by bevacizumab showed a higher OS rate in females as compared to males with NSCLC [137]. This result underlines the need for further studies that consider sex as a variable to better understand the different responses to targeted cancer therapies between males and females.
7. Models to Study a Sex Difference in Lung Cancer
Model systems will aid in the study of mechanisms driving sex differences in cancer incidence and mortality in the clinic. Invertebrate models including D. melanogaster and C. elegans may not be useful for this purpose since, to our knowledge, no reports have been published describing a cancer sex difference in these models. In contrast, there are several vertebrate model systems with an observed sex difference in cancer incidence or mortality. In D. reno, both Myc and KRasV12 oncogenes preferentially induce liver cancer in males over females, with additional studies showing that androgens promote and estrogens inhibit cancer cell growth [138,139]. Studies in dogs and cats observed a similar sex bias to those observed in humans. Male dogs have an increase in cancers from several origins, including lymphatic, skin, genital, soft tissue, and respiratory system, whereas female dogs have an increase in mammary tumors [140,141]. In general, sprayed and neutered dogs show an increase in some non-reproductive tumors including the heart, prostate, urinary tract, lymphatic and bone cancers [142], suggesting, similarly to humans, that sex hormones influence cancer incidence rates.
A significant body of literature describes sex differences in cancer incidence in mice, including several commonly used inbred strains [143]. The spontaneous cancers in mice are predominantly lymphoma, with a lung cancer incidence at ~5% of mice necropsied at death from old age—[143]. However, in some F1 hybrid strains, it can be as high as 30% with a male bias [144]. Because of this low incidence (~5%), and long latency time (~2 years) a variety of syngeneic, chemical-induced, and genetic models have been developed to study lung cancer. In each of these induced models, a sex difference is observed that mirrors what is observed for humans.
A common syngeneic mouse lung cancer model utilized is the syngeneic Lewis lung carcinoma (LLC) model [145,146]. The use of a subcutaneous model of LLC shows no difference in growth between the sexes in some studies [147,148] or an increased growth in males over females when fed a high fat diet [149]. Ovariectomy causes subcutaneous LLC tumors to grow more quickly in female mice [150,151]. LLC orthotopic tumors grow more quickly in females than in males [152]. The urethane model is a widely used and well-characterized mouse model of carcinogen-induced lung cancer [153]. The urethane model shows a clear difference between male and female mice from several inbred backgrounds, with tumors appearing earlier in male mice than in female mice [154,155]. Ovariectomized, but not castrated, mice develop urethane-induced lung tumors more quickly than their intact counterparts, suggesting that female sex hormones suppress lung cancer growth [156].
Genetically engineered mouse models (GEMMs) have also played a significant role in lung cancer research. A sex difference has been observed in the Kras, p53 (KP model) with some studies showing gender differences in survival [157]. Ovariectomy of KP mice causes tumors to form more quickly and grow more aggressively than their intact counterparts, but administration of exogenous estrogen can also cause greater tumor formation [39]. Administration of estrogen to male KP mice resulted in higher-grade tumor formation than that observed for their vehicle counterparts, indicating an important role for estrogen in this specific model.
8. Discussion
While the literature largely agrees that there is an observed sex difference in lung cancer, the underlying mechanisms and drivers have yet to be fully elucidated. The consensus is that men have a higher tumor grade at diagnosis and an increased mortality from lung cancer compared to women. Interestingly, women tend to respond better to cytotoxic and targeted therapies, whereas men have a better response to immunotherapies. It is also clear that there are key differences in the immune systems between men and women, with women having an enhanced innate and adaptive immune response compared to men (Figure 1A). In addition to considerations of sex, racial and ethnic differences must also be taken into account. Data from both the SEER program and from the CDC clearly show higher rates of lung cancer in African Americans than in whites, and those of Hispanic ethnicity had a lower incidence of lung cancer than non-Hispanics [158]. In addition to this, African American patients have lower survival rates than white patients, and African American patients are less likely to receive surgical resection and/or timely treatment compared to white patients [46,159]. Racial and ethnic differences in smoking rates, as well as the impact of other socioeconomic factors, can also have impacts on incidence and survival [160]. In addition to socioeconomic factors, there are also genomic and epigenomic differences between racial and ethnic groups that can influence lung cancer incidence. In one example, a recent analysis of the TCGA datasets identified key gene expression, protein expression, and pathway differences which corelate with differences in LUAD survival between ethnic groups (https://doi.org/10.3390/cancers15102695 (accessed on 1 June 2023)). In other studies, patients of East Asian descent are more likely to have EGFR mutations, for example, which could affect treatment and screening decisions for this population in addition to any adjustments for sex [161]. Outside of EGFR, there are few studies that consider specific racial and ethnic differences in oncogene mutation frequencies; more work needs to be carried out in this area to identify potential differences that could influence treatment decisions. How these differences could contribute to the clinical differences in lung cancer between men and women is an area of active research.
Key areas need to be the focus of further investigations. Further work needs to be carried out to elucidate the role that estrogen and hormone replacement therapy plays in lung cancer risk in women, as well as to study the roles that progesterone and testosterone may play. The potential effects that hormone replacement therapy may have on the transgender population’s risk for lung cancer must also be considered. Transgender adults in the United States have a higher smoking rate than cisgender adults, putting them at higher risk for smoking-related diseases, and one study showed a high incidence of lung cancer in a population of transgender women after gender affirmation therapy in Amsterdam [162,163]. While research that includes and that is dedicated to the transgender population is still uncommon, more work must be carried out to include this population (Figure 1B). Future studies that consider menopausal status, not just age, and measurements of sex hormone abundance may help clarify the exact functions for menopause/sex hormones in lung cancer. Estrogen’s effects on immune cell function are an area that could be particularly important, as this could not only explain differences in cancer initiation and progression, but also immunotherapy response and potential combinations of immunotherapies with other therapies, potentially aromatase inhibitors that would prevent estrogen production. These categories also need to be considered in therapeutic clinical trials, as estrogen and/or progesterone status may affect therapeutic effectiveness in female patients; for example, sex hormones can affect PD-1 pathway activity, though the effect this may have on immunotherapy response has not been clearly defined (Figure 1B) [164]. In regard to hormone replacement therapy, studies need to be performed on both pre- and post-menopausal individuals that take HRT to assess the potential contribution that it may have to lung cancer development. The potential influences that HRT may have during treatment on both the cancer itself and immune cell behavior need to be studied to potentially develop recommendations for either continuing HRT during treatment or ceasing use. Less frequently discussed hormones such as activins, follicular-stimulating hormone (FSH), and prolactin have all been implicated in other cancer types; however, little work has been carried out as regards lung cancer (Table 1). Activin-A has been shown to be particularly important in melanoma, where it can affect metastatic potential through evasion of the immune system [165], and one study has shown that prolactin may be a possible early predictive factor in lung cancer patients with metastases [166]. It is important to also focus studies on males, not just females. As an example, the male hormone androgen has been recently identified as a key regulator of CD8 T cell exhaustion in models of bladder, melanoma, and colon cancer, but not of lung cancer [167,168]. Ultimately, it will be important to recognize that sex hormones influence a wide variety of processes in both the tumor cells and in the body. Thus, they could be playing both pro-tumor (direct effects of estrogens on lung cancer growth), and antitumor effects through an enhanced antitumor immune response (Figure 1C). Gaining a solid understanding of a mechanistic role they may play in lung cancer development and progression, immune responses to lung cancer, and therapeutic response is necessary for patient wellbeing and the development of new treatments.
While it is clear that the immune system plays an important role in lung cancer development and progression, further studies must be carried out to characterize specific differences in immune responses to lung cancer between males and females. Regarding the immune system, exploring specific pathways that could have different activity between males and females, such as apoptosis-inducing pathways, or checkpoint regulators such as PD-L1, could be a novel avenue to explore for a sex difference. As new immunotherapies continue to be developed and brought into clinical practice, knowing specific pathways that behave differently between males and females can inform not only therapy design but also novel combinations, whether with traditional chemotherapies or newer targeted therapies. Recent work using a TRAIL agonist shows promise in multiple tumor types, though the studies were only performed in female mice, and more studies would need to be performed to study their effectiveness in males [169]. Additionally, androgens have been previously shown to regulate tumor cell checkpoint expression [170,171]. Identifying specific differences in immune cell function and activity between the sexes would open up possibilities for targeted therapies that could take advantage of a pathway with greater activity towards tumor cells from one sex to improve outcomes, or that could try to activate pathways in one sex to improve the antitumor immune response. A better understanding of these differences in the immune system could, in turn, help explain the differences in the response to immunotherapies and traditional chemotherapies.
Studies on the beneficial effects of immunotherapies need to be performed with a potential sex difference in mind; study populations need to intentionally include both sexes and control for menopause in female patents—not just age—in recruitment and analysis. Several analyses have been performed to date with inconclusive and conflicting results, suggesting that not all the variables that affect immunotherapy outcomes have been defined [172]. As new immunotherapies continue to be developed, intentional work needs to be carried out prior to entering the clinic to determine any possible sex differences in response by including both sexes in any animal work and in phase 1 trials. Gaining a better understanding of what therapies are most beneficial for each sex could help guide decision making in clinical practice and ensure each patient receives the best possible treatment, whether that is an immunotherapy, a conventional chemotherapy regimen, a targeted therapy, or a combination. This is especially important as women are often more likely to experience greater toxicity from many traditional cancer treatments [124]. The development and administration of treatments must be equitable between the sexes and must therefore be based on research that considers these factors.
Studies into genetic factors have provided potential targets for therapy; however, more research is needed to continue to study the differences between males and females in DNA damage response pathways and how those can be influenced by smoking. Even though tobacco smoking rates continue to decline worldwide, newer smoking-related activities such as vaping require similar studies into their potential for causing lung cancer and whether there may be a genetic basis for any sex differences. Because vaping is such a new technology, determining its effects will take time. However, there is reason for concern; the analysis of vaping fluids shows high levels of known and probable carcinogens, and the United States may face a significant health burden as the effects of vaping fluid consumption become apparent in the future [173]. In addition to the effects of tobacco consumption and vaping, more studies into other environmental factors and the role they may play in DNA damage and epigenetic changes need to be performed.
Additionally, research needs to be carried out to characterize the contributions that the patient’s microbiome makes to lung cancer development and progression and how this could impact any observed sex differences. The microbiome has well-characterized roles in human cancer [174] and therapy effectiveness, including immunotherapies [175]. The gut microbiome regulates hormone-dependent immune cell activity, suggesting that the microbiome could be a contributing factor for sex differences in the cancer antitumor immune response [176]. In addition to the gut microbiome, lung cancer can be influenced by the lung microbiome [177]. One study has already shown that aberrations in the lung microbiome can increase the risk of malignancy, and another study has shown that treatment with antibiotics during immunotherapy can actually lower progression-free survival—though neither of these studies examined a sex difference [178,179]. The microbiome is rarely, if ever, controlled for in clinical studies and could be a significant variable in outcomes in studies investigating sex disparities. Further research in this area could contribute to a greater understanding of basic sex differences in lung cancer development and progression, as well as differences in immune responses and responses to therapies.
9. Conclusions
Lung cancer shows a clear difference in presentation and therapeutic response between the sexes, though specific biological mechanisms that would explain this phenomenon have yet to be described. This difference is likely a combination of environmental factors and innate biological factors working together in the disease process. Further work needs to be carried out to study the role of sex hormones and the role of the immune system in facilitating this difference, as well as to study differences in the therapeutic response and the potential biological reasons behind this difference. Gaining a deeper understanding of the biological reasons behind the sex difference may open the door to new therapeutic combinations or novel therapies based on differing biological mechanisms between the sexes.
Author Contributions
Conceptualization, L.M. and J.W.L.; methodology, L.M. and J.W.L.; validation, L.M. and J.W.L.; formal analysis, L.M., K.S., P.N.-S., H.L. and J.W.L.; writing—original draft preparation, L.M. and J.W.L.; writing—review and editing, L.M., K.S., P.N.-S., H.L. and J.W.L.; visualization, L.M. and J.W.L.; supervision, J.W.L.; project administration, J.W.L.; funding acquisition, P.N.-S., H.L. and J.W.L. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by internal funds from the VCU School of Medicine and an NCI Diversity Supplement to L.M., the VCU SUCEEED grant P20CA264067 and P20CA264068 to K.S., 5P20CA252717-02 funds to P.N.-S., VA Office of Research and Development and Cooperative Studies Program’s Lung Precision Oncology Program (VA ORD/CSP L0014) to H.L., and VCU School of Medicine and VCU Massey Cancer Center internal funds to J.W.L.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All data used in the paper are publicly available.
Acknowledgments
The authors would like to the members of the Shows, Nana-Sinkam, Li and Landry labs for critically reading the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Siegel, R.L.; Miller, K.D.; Fuchs, H.E.; Jemal, A. Cancer statistics. CA Cancer J. Clin. 2022, 72, 7–33. [Google Scholar] [CrossRef] [PubMed]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef]
- Ashley, D.J. Sex differences in the incidence of tumours at various sites. Br. J. Cancer. 1969, 23, 26–30. [Google Scholar] [CrossRef] [Green Version]
- Wagner, A.; Oertelt-Prigione, S.; Adjei, A.; Buclin, T.; Cristina, V.; Csajka, C.; Coukos, G.; Dafni, U.; Dotto, G.-P.; Ducreux, M.; et al. Gender medicine and oncology: Report and consensus of an ESMO workshop. Ann. Oncol. 2019, 30, 1914–1924. [Google Scholar] [CrossRef] [Green Version]
- Visbal, A.L.; Williams, B.A.; Nichols, F.C.; Marks, R.S.; Jett, J.R.; Aubry, M.-C.; Edell, E.S.; Wampfler, J.A.; Molina, J.R.; Yang, P. Gender differences in non-small-cell lung cancer survival: An analysis of 4,618 patients diagnosed between 1997 and 2002. Ann. Thorac. Surg. 2004, 78, 209–215; discussion 15. [Google Scholar] [CrossRef]
- Fu, J.B.; Kau, T.Y.; Severson, R.K.; Kalemkerian, G.P. Lung cancer in women: Analysis of the national Surveillance, Epidemiology, and End Results database. Chest 2005, 127, 768–777. [Google Scholar] [CrossRef] [Green Version]
- Kinoshita, F.L.; Ito, Y.; Morishima, T.; Miyashiro, I.; Nakayama, T. Sex differences in lung cancer survival: Long-term trends using population-based cancer registry data in Osaka, Japan. Jpn. J. Clin. Oncol. 2017, 47, 863–869. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alvarez, A.A.R.; Yuming, S.; Kothari, J.; Digumarthy, S.R.; Byrne, N.M.; Li, Y.; Christiani, D.C. Sex disparities in lung cancer survival rates based on screening status. Lung Cancer 2022, 171, 115–120. [Google Scholar] [CrossRef] [PubMed]
- Becker, N.; Motsch, E.; Trotter, A.; Heussel, C.P.; Dienemann, H.; Schnabel, P.A.; Kauczor, H.-U.; Maldonado, S.G.; Miller, A.B.; Kaaks, R.; et al. Lung cancer mortality reduction by LDCT screening-Results from the randomized German LUSI trial. Int. J. Cancer 2020, 146, 1503–1513. [Google Scholar] [CrossRef]
- Bonney, A.; Malouf, R.; Marchal, C.; Manners, D.; Fong, K.M.; Marshall, H.M.; Irving, L.B.; Manser, R. Impact of low-dose computed tomography (LDCT) screening on lung cancer-related mortality. Cochrane Database Syst. Rev. 2022, 8, CD013829. [Google Scholar] [CrossRef]
- Donington, J.S.; Colson, Y.L. Sex and gender differences in non-small cell lung cancer. Semin. Thorac. Cardiovasc. Surg. 2011, 23, 137–145. [Google Scholar] [CrossRef] [PubMed]
- Folkerd, E.J.; Dowsett, M. Influence of sex hormones on cancer progression. J. Clin. Oncol. 2010, 28, 4038–4044. [Google Scholar] [CrossRef] [PubMed]
- Capper, C.P.; Rae, J.M.; Auchus, R.J. The Metabolism, Analysis, and Targeting of Steroid Hormones in Breast and Prostate Cancer. Horm. Cancer 2016, 7, 149–164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maximov, P.Y.; Abderrahman, B.; Curpan, R.F.; Hawsawi, Y.A.; Fan, P.; Jordan, V.C. A unifying biology of sex steroid-induced apoptosis in prostate and breast cancers. Endocr. Relat. Cancer 2018, 25, R83–R113. [Google Scholar] [CrossRef] [Green Version]
- Sorenson, G.D.; Pettengill, O.S.; Brinck-Johnsen, T.; Cate, C.C.; Maurer, L.H. Hormone production by cultures of small-cell carcinoma of the lung. Cancer 1981, 47, 1289–1296. [Google Scholar] [CrossRef]
- Hattori, M.; Imura, H.; Matsukura, S.; Yoshimoto, Y.; Sekita, K.; Tomomatsu, T.; Kyogoku, M.; Kameya, T. Multiple-hormone producing lung carcinoma. Cancer 1979, 43, 2429–2437. [Google Scholar] [CrossRef]
- Mazzone, P.J.; Arroliga, A.C. Endocrine paraneoplastic syndromes in lung cancer. Curr. Opin. Pulm. Med. 2003, 9, 313–320. [Google Scholar] [CrossRef]
- Pietras, R.J.; Márquez, D.C.; Chen, H.-W.; Tsai, E.; Weinberg, O.; Fishbein, M. Estrogen and growth factor receptor interactions in human breast and non-small cell lung cancer cells. Steroids 2005, 70, 372–381. [Google Scholar] [CrossRef]
- Ettinger, B.; Friedman, G.D.; Bush, T.; Quesenberry, C.P., Jr. Reduced mortality associated with long-term postmenopausal estrogen therapy. Obstet. Gynecol. 1996, 87, 6–12. [Google Scholar] [CrossRef]
- Schabath, M.B.; Wu, X.; Vassilopoulou-Sellin, R.; Vaporciyan, A.A.; Spitz, M.R. Hormone replacement therapy and lung cancer risk: A case-control analysis. Clin. Cancer Res. 2004, 10 (1 Pt 1), 113–123. [Google Scholar] [CrossRef] [Green Version]
- Olsson, H.; Bladström, A.; Ingvar, C. Are smoking-associated cancers prevented or postponed in women using hormone replacement therapy? Obstet. Gynecol. 2003, 102, 565–570. [Google Scholar] [CrossRef] [PubMed]
- Taioli, E.; Wynder, E.L. Re: Endocrine factors and adenocarcinoma of the lung in women. JNCI J. Natl. Cancer Inst. 1994, 86, 869–870. [Google Scholar] [CrossRef] [PubMed]
- Nakaya, M.; Tachibana, H.; Yamada, K. Effect of estrogens on the interferon-gamma producing cell population of mouse splenocytes. Biosci. Biotechnol. Biochem. 2006, 70, 47–53. [Google Scholar] [CrossRef] [Green Version]
- Hao, S.; Zhao, J.; Zhou, J.; Zhao, S.; Hu, Y.; Hou, Y. Modulation of 17beta-estradiol on the number and cytotoxicity of NK cells in vivo related to MCM and activating receptors. Int. Immunopharmacol. 2007, 7, 1765–1775. [Google Scholar] [CrossRef] [PubMed]
- Straub, R.H. The complex role of estrogens in inflammation. Endocr. Rev. 2007, 28, 521–574. [Google Scholar] [CrossRef] [Green Version]
- Phiel, K.L.; Henderson, R.A.; Adelman, S.J.; Elloso, M.M. Differential estrogen receptor gene expression in human peripheral blood mononuclear cell populations. Immunol. Lett. 2005, 97, 107–113. [Google Scholar] [CrossRef]
- Dunn, S.E.; Ousman, S.S.; Sobel, R.A.; Zuniga, L.; Baranzini, S.E.; Youssef, S.; Crowell, A.; Loh, J.; Oksenberg, J.; Steinman, L. Peroxisome proliferator-activated receptor (PPAR)alpha expression in T cells mediates gender differences in development of T cell-mediated autoimmunity. J. Exp. Med. 2007, 204, 321–330. [Google Scholar] [CrossRef] [Green Version]
- Hewagama, A.; Patel, D.; Yarlagadda, S.; Strickland, F.M.; Richardson, B.C. Stronger inflammatory/cytotoxic T-cell response in women identified by microarray analysis. Genes Immun. 2009, 10, 509–516. [Google Scholar] [CrossRef] [Green Version]
- Bengtsson, A.K.; Ryan, E.J.; Giordano, D.; Magaletti, D.M.; Clark, E.A. 17beta-estradiol (E2) modulates cytokine and chemokine expression in human monocyte-derived dendritic cells. Blood 2004, 104, 1404–1410. [Google Scholar] [CrossRef]
- Hannah, M.F.; Bajic, V.B.; Klein, S.L. Sex differences in the recognition of and innate antiviral responses to Seoul virus in Norway rats. Brain Behav. Immun. 2008, 22, 503–516. [Google Scholar] [CrossRef] [Green Version]
- Klein, S.L.; Jedlicka, A.; Pekosz, A. The Xs and Y of immune responses to viral vaccines. Lancet Infect. Dis. 2010, 10, 338–349. [Google Scholar] [CrossRef] [PubMed]
- Klein, S.L.; Flanagan, K.L. Sex differences in immune responses. Nat. Rev. Immunol. 2016, 16, 626–638. [Google Scholar] [CrossRef] [PubMed]
- Ishibashi, H.; Suzuki, T.; Suzuki, S.; Niikawa, H.; Lu, L.; Miki, Y.; Moriya, T.; Hayashi, S.-I.; Handa, M.; Kondo, T.; et al. Progesterone receptor in non-small cell lung cancer—A potent prognostic factor and possible target for endocrine therapy. Cancer Res. 2005, 65, 6450–6458. [Google Scholar] [CrossRef] [Green Version]
- Butts, C.L.; Shukair, S.A.; Duncan, K.M.; Bowers, E.; Horn, J.; Belyavskaya, E.; Tonelli, L.; Sternberg, E.M. Progesterone inhibits mature rat dendritic cells in a receptor-mediated fashion. Int. Immunol. 2007, 19, 287–296. [Google Scholar] [CrossRef] [PubMed]
- Arruvito, L.; Giulianelli, S.; Flores, A.C.; Paladino, N.; Barboza, M.; Lanari, C.; Fainboim, L. NK cells expressing a progesterone receptor are susceptible to progesterone-induced apoptosis. J. Immunol. 2008, 180, 5746–5753. [Google Scholar] [CrossRef] [Green Version]
- Menzies, F.M.; Henriquez, F.L.; Alexander, J.; Roberts, C.W. Selective inhibition and augmentation of alternative macrophage activation by progesterone. Immunology 2011, 134, 281–291. [Google Scholar] [CrossRef]
- Chan, Y.X.; Alfonso, H.; Chubb, S.A.P.; Handelsman, D.J.; Fegan, P.G.; Hankey, G.J.; Golledge, J.; Flicker, L.; Yeap, B.B. Higher Dihydrotestosterone Is Associated with the Incidence of Lung Cancer in Older Men. Horm. Cancer 2017, 8, 119–126. [Google Scholar] [CrossRef] [Green Version]
- Hyde, Z.; Flicker, L.; McCaul, K.A.; Almeida, O.P.; Hankey, G.J.; Chubb, S.P.; Yeap, B.B. Associations between testosterone levels and incident prostate, lung, and colorectal cancer. A population-based study. Cancer Epidemiol. Biomark. Prev. 2012, 21, 1319–1329. [Google Scholar] [CrossRef] [Green Version]
- Hammoud, Z.; Tan, B.; Badve, S.; Bigsby, R.M. Estrogen promotes tumor progression in a genetically defined mouse model of lung adenocarcinoma. Endocr. Relat. Cancer 2008, 15, 475–483. [Google Scholar] [CrossRef]
- Roberts, C.W.; Walker, W.; Alexander, J. Sex-associated hormones and immunity to protozoan parasites. Clin. Microbiol. Rev. 2001, 14, 476–488. [Google Scholar] [CrossRef] [Green Version]
- Rettew, J.A.; Huet-Hudson, Y.M.; Marriott, I. Testosterone reduces macrophage expression in the mouse of toll-like receptor 4, a trigger for inflammation and innate immunity. Biol. Reprod. 2008, 78, 432–437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roden, A.C.; Moser, M.T.; Tri, S.D.; Mercader, M.; Kuntz, S.M.; Dong, H.; Hurwitz, A.A.; McKean, D.J.; Celis, E.; Leibovich, B.C.; et al. Augmentation of T cell levels and responses induced by androgen deprivation. J. Immunol. 2004, 173, 6098–6108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iqbal, J.; Sun, L.; Kumar, T.R.; Blair, H.C.; Zaidi, M. Follicle-stimulating hormone stimulates TNF production from immune cells to enhance osteoblast and osteoclast formation. Proc. Natl. Acad. Sci. USA 2006, 103, 14925–14930. [Google Scholar] [CrossRef] [Green Version]
- Ray, A.; Cleary, M.P. The potential role of leptin in tumor invasion and metastasis. Cytokine Growth Factor Rev. 2017, 38, 80–97. [Google Scholar] [CrossRef] [PubMed]
- Lord, G.M.; Matarese, G.; Howard, J.K.; Baker, R.J.; Bloom, S.R.; Lechler, R.I. Leptin modulates the T-cell immune response and reverses starvation-induced immunosuppression. Nature 1998, 394, 897–901. [Google Scholar] [CrossRef]
- Zullig, L.L.; Carpenter, W.R.; Provenzale, D.T.; Weinberger, M.; Reeve, B.B.; Williams, C.D.; Jackson, G.L. The association of race with timeliness of care and survival among Veterans Affairs health care system patients with late-stage non-small cell lung cancer. Cancer Manag. Res. 2013, 5, 157–163. [Google Scholar] [CrossRef] [Green Version]
- Borba, V.V.; Zandman-Goddard, G.; Shoenfeld, Y. Prolactin and Autoimmunity. Front. Immunol. 2018, 9, 73. [Google Scholar] [CrossRef]
- Weinberg, O.K.; Marquez-Garban, D.C.; Fishbein, M.C.; Goodglick, L.; Garban, H.J.; Dubinett, S.M.; Pietras, R.J. Aromatase inhibitors in human lung cancer therapy. Cancer Res. 2005, 65, 11287–11291. [Google Scholar] [CrossRef] [Green Version]
- Stabile, L.P.; Davis, A.L.; Gubish, C.T.; Hopkins, T.M.; Luketich, J.D.; Christie, N.; Finkelstein, S.; Siegfried, J.M. Human non-small cell lung tumors and cells derived from normal lung express both estrogen receptor alpha and beta and show biological responses to estrogen. Cancer Res. 2002, 62, 2141–2150. [Google Scholar]
- DeRoo, B.J.; Korach, K.S. Estrogen receptors and human disease. J. Clin. Investig. 2006, 116, 561–570. [Google Scholar] [CrossRef] [Green Version]
- Issa, J.-P.; Baylin, S.B.; Belinsky, S.A. Methylation of the estrogen receptor CpG island in lung tumors is related to the specific type of carcinogen exposure. Cancer Res. 1996, 56, 3655–3658. [Google Scholar] [PubMed]
- Chakraborty, S.; Ganti, A.K.; Marr, A.; Batra, S.K. Lung cancer in women: Role of estrogens. Expert Rev. Respir. Med. 2010, 4, 509–518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Márquez-Garbán, D.C.; Chen, H.-W.; Fishbein, M.C.; Goodglick, L.; Pietras, R.J. Estrogen receptor signaling pathways in human non-small cell lung cancer. Steroids 2007, 72, 135–143. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, A.G.; Prysak, G.M.; Murphy, V.; Lonardo, F.; Pass, H.; Schwartz, J.; Brooks, S. Nuclear estrogen receptor beta in lung cancer: Expression and survival differences by sex. Clin. Cancer Res. 2005, 11, 7280–7287. [Google Scholar] [CrossRef] [Green Version]
- Mah, V.; Seligson, D.B.; Li, A.; Márquez, D.C.; Wistuba, I.I.; Elshimali, Y.; Fishbein, M.C.; Chia, D.; Pietras, R.J.; Goodglick, L. Aromatase expression predicts survival in women with early-stage non small cell lung cancer. Cancer Res. 2007, 67, 10484–10490. [Google Scholar] [CrossRef] [Green Version]
- Kawai, H.; Ishii, A.; Washiya, K.; Konno, T.; Kon, H.; Yamaya, C.; Ono, I.; Minamiya, Y.; Ogawa, J. Estrogen receptor alpha and beta are prognostic factors in non-small cell lung cancer. Clin. Cancer Res. 2005, 11, 5084–5089. [Google Scholar] [CrossRef] [Green Version]
- Fasco, M.J.; Hurteau, G.J.; Spivack, S.D. Gender-dependent expression of alpha and beta estrogen receptors in human nontumor and tumor lung tissue. Mol. Cell Endocrinol. 2002, 188, 125–140. [Google Scholar] [CrossRef]
- Márquez-Garbán, D.C.; Chen, H.-W.; Goodglick, L.; Fishbein, M.C.; Pietras, R.J. Targeting aromatase and estrogen signaling in human non-small cell lung cancer. Ann. N. Y. Acad. Sci. 2009, 1155, 194–205. [Google Scholar] [CrossRef] [Green Version]
- Burger, H.G. Androgen production in women. Fertil. Steril. 2002, 77 (Suppl. 4), S3–S5. [Google Scholar] [CrossRef]
- Nadkarni, S.; Cooper, D.; Brancaleone, V.; Bena, S.; Perretti, M. Activation of the annexin A1 pathway underlies the protective effects exerted by estrogen in polymorphonuclear leukocytes. Arter. Thromb. Vasc. Biol. 2011, 31, 2749–2759. [Google Scholar] [CrossRef] [Green Version]
- Lin, C.-Y.; Ström, A.; Vega, V.B.; Kong, S.L.; Yeo, A.L.; Thomsen, J.S.; Chan, W.C.; Doray, B.; Bangarusamy, D.K.; Ramasamy, A.; et al. Discovery of estrogen receptor alpha target genes and response elements in breast tumor cells. Genome Biol. 2004, 5, R66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Ma, H.; Yao, J. ERalpha, A Key Target for Cancer Therapy: A Review. Onco. Targets Ther. 2020, 13, 2183–2191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsu, L.-H.; Chu, N.-M.; Kao, S.-H. Estrogen, Estrogen Receptor and Lung Cancer. Int. J. Mol. Sci. 2017, 18, 1713. [Google Scholar] [CrossRef] [PubMed]
- Adami, H.-O.; Persson, I.; Hoover, R.; Schairer, C.; Bergkvist, L. Risk of cancer in women receiving hormone replacement therapy. Int. J. Cancer 1989, 44, 833–839. [Google Scholar] [CrossRef]
- Kazmi, N.; Márquez-Garbán, D.C.; Aivazyan, L.; Hamilton, N.; Garon, E.B.; Goodglick, L.; Pietras, R.J. The role of estrogen, progesterone and aromatase in human non-small-cell lung cancer. Lung Cancer Manag. 2012, 1, 259–272. [Google Scholar] [CrossRef] [Green Version]
- Ganti, A.K.; Sahmoun, A.E.; Panwalkar, A.W.; Tendulkar, K.K.; Potti, A. Hormone replacement therapy is associated with decreased survival in women with lung cancer. J. Clin. Oncol. 2006, 24, 59–63. [Google Scholar] [CrossRef]
- Chlebowski, R.T.; Schwartz, A.G.; Wakelee, H.; Anderson, G.L.; Stefanick, M.L.; Manson, J.E.; Rodabough, R.J.; Chien, J.W.; Wactawski-Wende, J.; Gass, M.; et al. Oestrogen plus progestin and lung cancer in postmenopausal women (Women’s Health Initiative trial): A post-hoc analysis of a randomised controlled trial. Lancet 2009, 374, 1243–1251. [Google Scholar] [CrossRef] [Green Version]
- The Coronary Drug Project. Findings leading to discontinuation of the 2.5-mg day estrogen group. The coronary Drug Project Research Group. JAMA 1973, 226, 652–657. [Google Scholar] [CrossRef]
- Yager, J.D.; Davidson, N.E. Estrogen carcinogenesis in breast cancer. N. Engl. J. Med. 2006, 354, 270–282. [Google Scholar] [CrossRef] [Green Version]
- Koushik, A.; Parent, M.; Siemiatycki, J. Characteristics of menstruation and pregnancy and the risk of lung cancer in women. Int. J. Cancer. 2009, 125, 2428–2433. [Google Scholar] [CrossRef]
- Moore, K.A.; Mery, C.M.; Jaklitsch, M.T.; Estocin, A.P.; Bueno, R.; Swanson, S.J.; Sugarbaker, D.J.; Lukanich, J.M. Menopausal effects on presentation, treatment, and survival of women with non-small cell lung cancer. Ann. Thorac. Surg. 2003, 76, 1789–1795. [Google Scholar] [CrossRef] [PubMed]
- Richards, J.S.; Pangas, S.A. The ovary: Basic biology and clinical implications. J. Clin. Investig. 2010, 120, 963–972. [Google Scholar] [CrossRef] [Green Version]
- Su, J.M.; Hsu, H.K.; Chang, H.; Lin, S.L.; Chang, H.C.; Huang, M.S.; Tseng, H.H. Expression of estrogen and progesterone receptors in non-small-cell lung cancer: Immunohistochemical study. Anticancer Res. 1996, 16, 3803–3806. [Google Scholar] [PubMed]
- Kaiser, U.; Hofmann, J.; Schilli, M.; Wegmann, B.; Klotz, U.; Wedel, S.; Virmani, A.K.; Wollmer, E.; Branscheid, D.; Gazdar, A.F.; et al. Steroid-hormone receptors in cell lines and tumor biopsies of human lung cancer. Int. J. Cancer 1996, 67, 357–364. [Google Scholar] [CrossRef]
- Raso, M.G.; Behrens, C.; Herynk, M.H.; Liu, S.; Prudkin, L.; Ozburn, N.C.; Woods, D.M.; Tang, X.; Mehran, R.J.; Moran, C.; et al. Immunohistochemical expression of estrogen and progesterone receptors identifies a subset of NSCLCs and correlates with EGFR mutation. Clin. Cancer Res. 2009, 15, 5359–5368. [Google Scholar] [CrossRef] [Green Version]
- Skjefstad, K.; Richardsen, E.; Donnem, T.; Andersen, S.; Kiselev, Y.; Grindstad, T.; Hald, S.M.; Al-Shibli, K.; Bremnes, R.M.; Busund, L.-T.; et al. The prognostic role of progesterone receptor expression in non-small cell lung cancer patients: Gender-related impacts and correlation with disease-specific survival. Steroids 2015, 98, 29–36. [Google Scholar] [CrossRef]
- Marquez-Garban, D.C.; Mah, V.; Alavi, M.; Maresh, E.L.; Chen, H.-W.; Bagryanova, L.; Horvath, S.; Chia, D.; Garon, E.; Goodglick, L.; et al. Progesterone and estrogen receptor expression and activity in human non-small cell lung cancer. Steroids 2011, 76, 910–920. [Google Scholar] [CrossRef] [Green Version]
- Chang, J.; Wu, Y.; Zhou, S.; Tian, Y.; Wang, Y.; Tian, J.; Song, W.; Dong, Y.; Li, J.; Zhao, Z.; et al. Genetically predicted testosterone and cancers risk in men: A two-sample Mendelian randomization study. J. Transl. Med. 2022, 20, 573. [Google Scholar] [CrossRef]
- Yim, S.-H.; Chung, Y.-J. Molecular epidemiology of female lung cancer. Cancers 2011, 3, 1861–1876. [Google Scholar] [CrossRef] [Green Version]
- Kligerman, S.; White, C. Epidemiology of lung cancer in women: Risk factors, survival, and screening. AJR Am. J. Roentgenol. 2011, 196, 287–295. [Google Scholar] [CrossRef]
- Berardi, R.; Verdecchia, L.; Paolo, M.D.P.; Giampieri, R.; Scartozzi, M.; Pierantoni, C.; Bianconi, M.; Mazzanti, P.; Cascinu, S. Women and lung cancer: Clinical and molecular profiling as a determinate for treatment decisions: A literature review. Crit. Rev. Oncol. Hematol. 2009, 69, 223–236. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.-F.; Ronis, M.J.J.; Ingelman-Sundberg, M.; Badger, T.M. Hormonal regulation of microsomal cytochrome P4502E1 and P450 reductase in rat liver and kidney. Xenobiotica 1999, 29, 437–451. [Google Scholar] [CrossRef] [PubMed]
- Mollerup, S.; Ryberg, D.; Hewer, A.; Phillips, D.H.; Haugen, A. Sex differences in lung CYP1A1 expression and DNA adduct levels among lung cancer patients. Cancer Res. 1999, 59, 3317–3320. [Google Scholar]
- Pogun, S.; Yararbas, G. Sex differences in nicotine action. Handb. Exp. Pharmacol. 2009, 192, 261–291. [Google Scholar] [CrossRef]
- Chenoweth, M.J.; Cox, L.S.; Nollen, N.L.; Ahluwalia, J.S.; Benowitz, N.L.; Lerman, C.; Knight, J.; Tyndale, R.F. Analyses of nicotine metabolism biomarker genetics stratified by sex in African and European Americans. Sci. Rep. 2021, 11, 19572. [Google Scholar] [CrossRef]
- Uchida, K.; Kojima, A.; Morokawa, N.; Tanabe, O.; Anzai, C.; Kawakami, M.; Eto, Y.; Yoshimura, K. Expression of progastrin-releasing peptide and gastrin-releasing peptide receptor mRNA transcripts in tumor cells of patients with small cell lung cancer. J. Cancer Res. Clin. Oncol. 2002, 128, 633–640. [Google Scholar] [CrossRef]
- Shriver, S.P.; Bourdeau, H.A.; Gubish, C.T.; Tirpak, D.L.; Davis, A.L.G.; Luketich, J.D.; Siegfried, J.M. Sex-specific expression of gastrin-releasing peptide receptor: Relationship to smoking history and risk of lung cancer. J. Natl. Cancer Inst. 2000, 92, 24–33. [Google Scholar] [CrossRef] [Green Version]
- Yuan, Y.; Liu, L.; Chen, H.; Wang, Y.; Xu, Y.; Mao, H.; Li, J.; Mills, G.B.; Shu, Y.; Li, L.; et al. Comprehensive Characterization of Molecular Differences in Cancer between Male and Female Patients. Cancer Cell 2016, 29, 711–722. [Google Scholar] [CrossRef] [Green Version]
- Houston, K.A.; Henley, S.J.; Li, J.; White, M.C.; Richards, T.B. Patterns in lung cancer incidence rates and trends by histologic type in the United States, 2004–2009. Lung Cancer 2014, 86, 22–28. [Google Scholar] [CrossRef] [Green Version]
- Thun, M.J.; Hannan, L.M.; Adams-Campbell, L.L.; Boffetta, P.; Buring, J.E.; Feskanich, D.; Flanders, W.D.; Jee, S.H.; Katanoda, K.; Kolonel, L.N.; et al. Lung cancer occurrence in never-smokers: An analysis of 13 cohorts and 22 cancer registry studies. PLoS Med. 2008, 5, e185. [Google Scholar] [CrossRef]
- Marang-van de Mheen, P.J.; Smith, G.D.; Hart, C.L.; Hole, D.J. Are women more sensitive to smoking than men? Findings from the Renfrew and Paisley study. Int. J. Epidemiol. 2001, 30, 787–792. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bain, C.; Feskanich, D.; Speizer, F.E.; Thun, M.; Hertzmark, E.; Rosner, B.A.; Colditz, G.A. Lung cancer rates in men and women with comparable histories of smoking. J. Natl. Cancer Inst. 2004, 96, 826–834. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hackshaw, A.K.; Law, M.R.; Wald, N.J. The accumulated evidence on lung cancer and environmental tobacco smoke. BMJ 1997, 315, 980–988. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pirie, K.; Peto, R.; Green, J.; Reeves, G.K.; Beral, V.; Million Women Study Collaborators. Lung cancer in never smokers in the UK Million Women Study. Int. J. Cancer 2016, 139, 347–354. [Google Scholar] [CrossRef] [Green Version]
- Cheng, Y.Y.; Rath, E.M.; Linton, A.; Yuen, M.L.; Takahashi, K.; Lee, K. The Current Understanding of Asbestos-Induced Epigenetic Changes Associated With Lung Cancer. Lung Cancer 2020, 11, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Cheng, Y.W.; Chiou, H.L.; Sheu, G.T.; Hsieh, L.L.; Chen, J.T.; Chen, C.Y.; Su, J.M.; Lee, H. The association of human papillomavirus 16/18 infection with lung cancer among nonsmoking Taiwanese women. Cancer Res. 2001, 61, 2799–2803. [Google Scholar]
- Bae, J.-M.; Kim, E.H. Human papillomavirus infection and risk of lung cancer in never-smokers and women: An ‘adaptive’ meta-analysis. Epidemiol. Health 2015, 37, e2015052. [Google Scholar] [CrossRef]
- Pérez-Díez, I.; Hidalgo, M.R.; Malmierca-Merlo, P.; Andreu, Z.; Romera-Giner, S.; Farràs, R.; de la Iglesia-Vayá, M.; Provencio, M.; Romero, A.; García-García, F. Functional Signatures in Non-Small-Cell Lung Cancer: A Systematic Review and Meta-Analysis of Sex-Based Differences in Transcriptomic Studies. Cancers 2021, 13, 143. [Google Scholar] [CrossRef]
- Conforti, F.; Pala, L.; Pagan, E.; Bagnardi, V.; De Pas, T.; Queirolo, P.; Pennacchioli, E.; Catania, C.; Cocorocchio, E.; Ferrucci, P.F.; et al. Sex-Based Dimorphism of Anticancer Immune Response and Molecular Mechanisms of Immune Evasion. Clin. Cancer Res. 2021, 27, 4311–4324. [Google Scholar] [CrossRef]
- Yang, Q.; Zhang, H.; Wei, T.; Lin, A.; Sun, Y.; Luo, P.; Zhang, J. Single-Cell RNA Sequencing Reveals the Heterogeneity of Tumor-Associated Macrophage in Non-Small Cell Lung Cancer and Differences Between Sexes. Front. Immunol. 2021, 12, 756722. [Google Scholar] [CrossRef]
- Araujo, J.M.; Prado, A.; Cardenas, N.K.; Zaharia, M.; Dyer, R.; Doimi, F.; Bravo, L.; Pinillos, L.; Morante, Z.; Aguilar, A.; et al. Repeated observation of immune gene sets enrichment in women with non-small cell lung cancer. Oncotarget 2016, 7, 20282–20292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdullah, M.; Chai, P.-S.; Chong, M.-Y.; Tohit, E.R.M.; Ramasamy, R.; Pei, C.P.; Vidyadaran, S. Gender effect on in vitro lymphocyte subset levels of healthy individuals. Cell Immunol. 2012, 272, 214–219. [Google Scholar] [CrossRef] [PubMed]
- Afshan, G.; Afzal, N.; Qureshi, S. CD4+CD25(hi) regulatory T cells in healthy males and females mediate gender difference in the prevalence of autoimmune diseases. Clin. Lab. 2012, 58, 567–571. [Google Scholar] [PubMed]
- Lee, B.-W.; Yap, H.-K.; Chew, F.-T.; Quah, T.-C.; Prabhakaran, K.; Chan, G.S.; Wong, S.-C.; Seah, C.-C. Age- and sex-related changes in lymphocyte subpopulations of healthy Asian subjects: From birth to adulthood. Cytometry 1996, 26, 8–15. [Google Scholar] [CrossRef]
- Weinstein, Y.; Ran, S.; Segal, S. Sex-associated differences in the regulation of immune responses controlled by the MHC of the mouse. J. Immunol. 1984, 132, 656–661. [Google Scholar] [CrossRef]
- Libert, C.; Dejager, L.; Pinheiro, I. The X chromosome in immune functions: When a chromosome makes the difference. Nat. Rev. Immunol. 2010, 10, 594–604. [Google Scholar] [CrossRef]
- Wu, Y.; Ju, Q.; Jia, K.; Yu, J.; Shi, H.; Wu, H.; Jiang, M. Correlation between sex and efficacy of immune checkpoint inhibitors (PD-1 and CTLA-4 inhibitors). Int. J. Cancer 2018, 143, 45–51. [Google Scholar] [CrossRef] [Green Version]
- Hou, J.; Wu, F.Z. Effect of sex hormones on NK and ADCC activity of mice. Int. J. Immunopharmacol. 1988, 10, 15–22. [Google Scholar] [CrossRef]
- D’Agostino, P.; Milano, S.; Barbera, C.; DI Bella, G.; LA Rosa, M.; Ferlazzo, V.; Farruggio, R.; Miceli, D.M.; Miele, M.; Castagnetta, L.; et al. Sex hormones modulate inflammatory mediators produced by macrophages. Ann. N. Y. Acad. Sci. 1999, 876, 426–429. [Google Scholar] [CrossRef]
- Domagala-Kulawik, J.; Osinska, I.; Hoser, G. Mechanisms of immune response regulation in lung cancer. Transl. Lung Cancer Res. 2014, 3, 15–22. [Google Scholar] [PubMed]
- Domagala-Kulawik, J. The role of the immune system in non-small cell lung carcinoma and potential for therapeutic intervention. Transl. Lung Cancer Res. 2015, 4, 177–190. [Google Scholar] [PubMed]
- Zhao, J.; Harper, R.; Barchowsky, A.; Di, Y.P.P. Identification of multiple MAPK-mediated transcription factors regulated by tobacco smoke in airway epithelial cells. Am. J. Physiol. Lung Cell Mol. Physiol. 2007, 293, L480–L490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uppal, S.S.; Verma, S.; Dhot, P.S. Normal values of CD4 and CD8 lymphocyte subsets in healthy indian adults and the effects of sex, age, ethnicity, and smoking. Cytom. B Clin. Cytom. 2003, 52, 32–36. [Google Scholar] [CrossRef]
- Qiu, F.; Liang, C.-L.; Liu, H.; Zeng, Y.-Q.; Hou, S.; Huang, S.; Lai, X.; Dai, Z. Impacts of cigarette smoking on immune responsiveness: Up and down or upside down? Oncotarget 2017, 8, 268–284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, C.K.; Man, J.; Lord, S.; Cooper, W.; Links, M.; Gebski, V.; Herbst, R.S.; Gralla, R.J.; Mok, T.; Yang, J.C.-H. Clinical and Molecular Characteristics Associated with Survival Among Patients Treated With Checkpoint Inhibitors for Advanced Non-Small Cell Lung Carcinoma: A Systematic Review and Meta-analysis. JAMA Oncol. 2018, 4, 210–216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conforti, F.; Pala, L.; Pagan, E.; Corti, C.; Bagnardi, V.; Queirolo, P.; Catania, C.; De Pas, T.; Giaccone, G. Sex-based differences in response to anti-PD-1 or PD-L1 treatment in patients with non-small-cell lung cancer expressing high PD-L1 levels. A systematic review and meta-analysis of randomized clinical trials. ESMO Open 2021, 6, 100251. [Google Scholar] [CrossRef]
- Grassadonia, A.; Sperduti, I.; Vici, P.; Iezzi, L.; Brocco, D.; Gamucci, T.; Pizzuti, L.; Maugeri-Saccà, M.; Marchetti, P.; Cognetti, G.; et al. Effect of Gender on the Outcome of Patients Receiving Immune Checkpoint Inhibitors for Advanced Cancer: A Systematic Review and Meta-Analysis of Phase III Randomized Clinical Trials. J. Clin. Med. 2018, 7, 542. [Google Scholar] [CrossRef] [Green Version]
- Liang, J.; Hong, J.; Tang, X.; Qiu, X.; Zhu, K.; Zhou, L.; Guo, D. Sex difference in response to non-small cell lung cancer immunotherapy: An updated meta-analysis. Ann. Med. 2022, 54, 2606–2616. [Google Scholar] [CrossRef]
- Conforti, F.; Pala, L.; Bagnardi, V.; De Pas, T.; Martinetti, M.; Viale, G.; Gelber, R.D.; Goldhirsch, A. Cancer immunotherapy efficacy and patients’ sex: A systematic review and meta-analysis. Lancet Oncol. 2018, 19, 737–746. [Google Scholar] [CrossRef]
- Xue, C.; Zheng, S.; Dong, H.; Lu, X.; Zhang, X.; Zhang, J.; Li, J.; Cui, H. Association Between Efficacy of Immune Checkpoint Inhibitors and Sex: An Updated Meta-Analysis on 21 Trials and 12,675 Non-Small Cell Lung Cancer Patients. Front. Oncol. 2021, 11, 627016. [Google Scholar] [CrossRef]
- Madala, S.; Rasul, R.; Singla, K.; Sison, C.; Seetharamu, N.; Castellanos, M.R. Gender Differences and Their Effects on Survival Outcomes in Lung Cancer Patients Treated With PD-1/PD-L1 Checkpoint Inhibitors: A Systematic Review and Meta-Analysis. Clin. Oncol. 2022, 34, 799–809. [Google Scholar] [CrossRef]
- Schmetzer, O.; Flörcken, A. Sex differences in the drug therapy for oncologic diseases. Handb. Exp. Pharmacol. 2012, 214, 411–442. [Google Scholar] [CrossRef]
- Unger, J.M.; Vaidya, R.; Albain, K.S.; LeBlanc, M.; Minasian, L.M.; Gotay, C.C.; Henry, N.L.; Fisch, M.J.; Lee, S.M.; Blanke, C.D.; et al. Sex Differences in Risk of Severe Adverse Events in Patients Receiving Immunotherapy, Targeted Therapy, or Chemotherapy in Cancer Clinical Trials. J. Clin. Oncol. 2022, 40, 1474–1486. [Google Scholar] [CrossRef] [PubMed]
- Wagner, A.D. Sex differences in cancer chemotherapy effects, and why we need to reconsider BSA-based dosing of chemotherapy. ESMO Open 2020, 5, e000770. [Google Scholar] [CrossRef]
- Sekine, I.; Yamamoto, N.; Nishio, K.; Saijo, N. Emerging ethnic differences in lung cancer therapy. Br. J. Cancer 2008, 99, 1757–1762. [Google Scholar] [CrossRef] [Green Version]
- Rossi, A.; Di Maio, M.; Chiodini, P.; Rudd, R.M.; Okamoto, H.; Skarlos, D.V.; Früh, M.; Qian, W.; Tamura, T.; Samantas, E.; et al. Carboplatin- or cisplatin-based chemotherapy in first-line treatment of small-cell lung cancer: The COCIS meta-analysis of individual patient data. J. Clin. Oncol. 2012, 30, 1692–1698. [Google Scholar] [CrossRef] [PubMed]
- Rossi, A.; Chiodini, P.; Sun, J.-M.; O’Brien, M.E.R.; von Plessen, C.; Barata, F.; Park, K.; Popat, S.; Bergman, B.; Parente, B.; et al. Six versus fewer planned cycles of first-line platinum-based chemotherapy for non-small-cell lung cancer: A systematic review and meta-analysis of individual patient data. Lancet Oncol. 2014, 15, 1254–1262. [Google Scholar] [CrossRef]
- NSCLC Meta-analysis Collaborative Group. Preoperative chemotherapy for non-small-cell lung cancer: A systematic review and meta-analysis of individual participant data. Lancet 2014, 383, 1561–1571. [Google Scholar] [CrossRef] [Green Version]
- Burdett, S.; Pignon, J.P.; Tierney, J.; Tribodet, H.; Stewart, L.; Le Pechoux, C.; Aupérin, A.; Le Chevalier, T.; Stephens, R.J.; Arriagada, R.; et al. Adjuvant chemotherapy for resected early-stage non-small cell lung cancer. Cochrane Database Syst. Rev. 2015, 3, CD011430. [Google Scholar] [CrossRef] [Green Version]
- Sekine, I.; Sumi, M.; Ito, Y.; Tanai, C.; Nokihara, H.; Yamamoto, N.; Kunitoh, H.; Ohe, Y.; Tamura, T. Gender difference in treatment outcomes in patients with stage III non-small cell lung cancer receiving concurrent chemoradiotherapy. Jpn. J. Clin. Oncol. 2009, 39, 707–712. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, H.; Sekine, I.; Yamada, K.; Nokihara, H.; Yamamoto, N.; Kunitoh, H.; Ohe, Y.; Tamura, T. Gender differences in treatment outcomes among patients with non-small cell lung cancer given a combination of carboplatin and paclitaxel. Oncology 2008, 75, 169–174. [Google Scholar] [CrossRef] [PubMed]
- Werner-Wasik, M.; Scott, C.; Cox, J.D.; Sause, W.T.; Byhardt, R.W.; Asbell, S.; Russell, A.; Komaki, R.; Lee, J.S. Recursive partitioning analysis of 1999 Radiation Therapy Oncology Group (RTOG) patients with locally-advanced non-small-cell lung cancer (LA-NSCLC): Identification of five groups with different survival. Int. J. Radiat. Oncol. Biol. Phys. 2000, 48, 1475–1482. [Google Scholar] [CrossRef]
- O’Connell, J.P.; Kris, M.G.; Gralla, R.J.; Groshen, S.; Trust, A.; Fiore, J.J.; Kelsen, D.P.; Heelan, R.T.; Golbey, R.B. Frequency and prognostic importance of pretreatment clinical characteristics in patients with advanced non-small-cell lung cancer treated with combination chemotherapy. J. Clin. Oncol. 1986, 4, 1604–1614. [Google Scholar] [CrossRef]
- Pinto, J.A.; Vallejos, C.S.; Raez, L.E.; Mas, L.A.; Ruiz, R.; Torres-Roman, J.S.; Morante, Z.; Araujo, J.M.; Gómez, H.L.; Aguilar, A.; et al. Gender and outcomes in non-small cell lung cancer: An old prognostic variable comes back for targeted therapy and immunotherapy? ESMO Open 2018, 3, e000344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soria, J.-C.; Ohe, Y.; Vansteenkiste, J.; Reungwetwattana, T.; Chewaskulyong, B.; Lee, K.H.; Dechaphunkul, A.; Imamura, F.; Nogami, N.; Kurata, T.; et al. Osimertinib in Untreated EGFR-Mutated Advanced Non-Small-Cell Lung Cancer. N. Engl. J. Med. 2018, 378, 113–125. [Google Scholar] [CrossRef] [PubMed]
- Peters, S.; Camidge, D.R.; Shaw, A.T.; Gadgeel, S.; Ahn, J.S.; Kim, D.W.; Ou, S.H.I.; Pérol, M.; Dziadziuszko, R.; Rosell, R.; et al. Alectinib versus Crizotinib in Untreated ALK-Positive Non-Small-Cell Lung Cancer. N. Engl. J. Med. 2017, 377, 829–838. [Google Scholar] [CrossRef]
- Brahmer, J.R.; Dahlberg, S.E.; Gray, R.J.; Schiller, J.H.; Perry, M.C.; Sandler, A.; Johnson, D.H. Sex differences in outcome with bevacizumab therapy: Analysis of patients with advanced-stage non-small cell lung cancer treated with or without bevacizumab in combination with paclitaxel and carboplatin in the Eastern Cooperative Oncology Group Trial 4599. J. Thorac. Oncol. 2011, 6, 103–108. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Lu, J.-W.; Huo, X.; Li, Y.; Li, Z.; Gong, Z. Effects of sex hormones on liver tumor progression and regression in Myc/xmrk double oncogene transgenic zebrafish. Gen. Comp. Endocrinol. 2019, 277, 112–121. [Google Scholar] [CrossRef]
- Li, Y.; Li, H.; Spitsbergen, J.M.; Gong, Z. Males develop faster and more severe hepatocellular carcinoma than females in kras(V12) transgenic zebrafish. Sci. Rep. 2017, 7, 41280. [Google Scholar] [CrossRef] [Green Version]
- Pinello, K.; Pires, I.; Castro, A.F.; Carvalho, P.T.; Santos, A.; de Matos, A.; Queiroga, F.; Canadas-Sousa, A.; Dias-Pereira, P.; Catarino, J.; et al. Cross Species Analysis and Comparison of Tumors in Dogs and Cats, by Age, Sex, Topography and Main Morphologies. Data from Vet-OncoNet. Vet. Sci. 2022, 9, 167. [Google Scholar] [CrossRef]
- Merlo, D.; Rossi, L.; Pellegrino, C.; Ceppi, M.; Cardellino, U.; Capurro, C.; Ratto, A.; Sambucco, P.; Sestito, V.; Tanara, G.; et al. Cancer incidence in pet dogs: Findings of the Animal Tumor Registry of Genoa, Italy. J. Vet. Intern. Med. 2008, 22, 976–984. [Google Scholar] [CrossRef] [PubMed]
- Smith, A.N. The role of neutering in cancer development. Vet. Clin. N. Am. Small Anim. Pract. 2014, 44, 965–975. [Google Scholar] [CrossRef] [PubMed]
- Smith, G.S.; Walford, R.L.; Mickey, M.R. Lifespan and incidence of cancer and other diseases in selected long-lived inbred mice and their F 1 hybrids. J. Natl. Cancer. Inst. 1973, 50, 1195–1213. [Google Scholar] [CrossRef] [PubMed]
- Haines, D.C.; Chattopadhyay, S.; Ward, J.M. Pathology of aging B6;129 mice. Toxicol. Pathol. 2001, 29, 653–661. [Google Scholar] [CrossRef] [PubMed]
- Mayo, J.G. Biologic characterization of the subcutaneously implanted Lewis lung tumor. Cancer Chemother. Rep. 2. 1972, 3, 325–330. [Google Scholar]
- Sugiura, K.; Stock, C.C. Studies in a tumor spectrum. III. The effect of phosphoramides on the growth of a variety of mouse and rat tumors. Cancer Res. 1955, 15, 38–51. [Google Scholar]
- Zhao, X.-Z.; Liu, Y.; Zhou, L.-J.; Yang, X.-Y.; Wang, Z.-Q.; Wu, Z.-H.; Yang, X.Y. Role of estrogen in lung cancer based on the estrogen receptor-epithelial mesenchymal transduction signaling pathways. OncoTargets Ther. 2015, 8, 2849–2863. [Google Scholar] [CrossRef] [Green Version]
- da Silva, F.M.; Rosa-Caldwell, M.E.; Schrems, E.R.; Martinez, L.; Amos, M.G.; Lim, S.; Cabrera, A.R.; Brown, J.L.; Washington, T.A.; Greene, N.P. PGC-1alpha overexpression is not sufficient to mitigate cancer cachexia in either male or female mice. Appl. Physiol. Nutr. Metab. 2022, 47, 933–948. [Google Scholar] [CrossRef]
- Yakar, S.; Nunez, N.P.; Pennisi, P.; Brodt, P.; Sun, H.; Fallavollita, L.; Zhao, H.; Scavo, L.; Novosyadlyy, R.; Kurshan, N.; et al. Increased tumor growth in mice with diet-induced obesity: Impact of ovarian hormones. Endocrinology 2006, 147, 5826–5834. [Google Scholar] [CrossRef] [Green Version]
- Dou, M.; Zhu, K.; Xiufang, C.; Zhang, Y.; Chen, X.; Zhou, X.; Ding, X.; Xueliang, Z.; Gu, Z.; Guo, M.; et al. Reproductive Hormones and Their Receptors May Affect Lung Cancer. Cell Physiol. Biochem. 2017, 44, 1425–1434. [Google Scholar] [CrossRef]
- Singh, P.; Sarkar, M.; Agrawal, U.; Huhtaniemi, I.; Pal, R. The transgenic expression of the beta-subunit of human chorionic gonadotropin influences the growth of implanted tumor cells. Oncotarget 2018, 9, 34670–34680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dubois, C.; Rocks, N.; Blacher, S.; Primac, I.; Gallez, A.; García-Caballero, M.; Gérard, C.; Brouchet, L.; Noël, A.; Lenfant, F.; et al. Lymph/angiogenesis contributes to sex differences in lung cancer through oestrogen receptor alpha signalling. Endocr. Relat. Cancer 2019, 26, 201–216. [Google Scholar] [CrossRef] [PubMed]
- Gurley, K.E.; Moser, R.D.; Kemp, C.J. Induction of Lung Tumors in Mice with Urethane. Cold Spring Harb. Protoc. 2015, 2015, pdb-prot077446. [Google Scholar] [CrossRef]
- Morozkova, T.S.; Kaledin, V.I. Sex-Related Differences in the Sensitivity to Carcinogenic Effect of Urethane on the Lungs in Mice Are Reversed after Neonatal Androgenization. Bull. Exp. Biol. Med. 2015, 159, 782–785. [Google Scholar] [CrossRef]
- Stathopoulos, G.T.; Sherrill, T.P.; Cheng, D.-S.; Scoggins, R.M.; Han, W.; Polosukhin, V.V.; Connelly, L.; Yull, F.E.; Fingleton, B.; Blackwell, T.S. Epithelial NF-kappaB activation promotes urethane-induced lung carcinogenesis. Proc. Natl. Acad. Sci. USA 2007, 104, 18514–18519. [Google Scholar] [CrossRef] [Green Version]
- Liebelt, R.A.; Liebelt, A.G.; Lane, M. Hormonal Influences on Urethan Carcinogenesis in C3h/F Mice. Cancer Res. 1964, 24, 1869–1879. [Google Scholar] [PubMed]
- Zheng, S.; El-Naggar, A.K.; Kim, E.S.; Kurie, J.M.; Lozano, G. A genetic mouse model for metastatic lung cancer with gender differences in survival. Oncogene 2007, 26, 6896–6904. [Google Scholar] [CrossRef] [Green Version]
- Fairley, T.L.; Tai, E.; Townsend, J.S.; Stewart, S.L.; Steele, C.B.; Davis, S.P.; Underwood, J.M. Racial/Ethnic disparities and geographic differences in lung cancer incidence—38 States and the District of Columbia, 1998–2006. Morb. Mortal. Wkly. Rep. 2010, 59, 1434–1438. [Google Scholar]
- Farjah, F.; Wood, D.E.; Yanez, N.D.; Vaughan, T.L.; Symons, R.G.; Krishnadasan, B.; Flum, D.R. Racial disparities among patients with lung cancer who were recommended operative therapy. Arch. Surg. 2009, 144, 14–18. [Google Scholar] [CrossRef] [Green Version]
- Schabath, M.B.; Cress, W.D.; Muñoz-Antonia, T. Racial and Ethnic Differences in the Epidemiology and Genomics of Lung Cancer. Cancer Control 2016, 23, 338–346. [Google Scholar] [CrossRef] [Green Version]
- Pao, W.; Miller, V.; Zakowski, M.; Doherty, J.; Politi, K.; Sarkaria, I.; Singh, B.; Heelan, R.; Rusch, V.; Fulton, L.; et al. EGF receptor gene mutations are common in lung cancers from “never smokers” and are associated with sensitivity of tumors to gefitinib and erlotinib. Proc. Natl. Acad. Sci. USA 2004, 101, 13306–13311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Blok, C.J.; Wiepjes, C.M.; van Velzen, D.M.; Staphorsius, A.S.; Nota, N.M.; Gooren, L.J.; Kreukels, B.P.; den Heijer, M. Mortality trends over five decades in adult transgender people receiving hormone treatment: A report from the Amsterdam cohort of gender dysphoria. Lancet Diabetes Endocrinol. 2021, 9, 663–670. [Google Scholar] [CrossRef] [PubMed]
- Wheldon, C.W.; Wiseman, K.P. Tobacco Use Among Transgender and Gender Non-conforming Adults in the United States. Tob. Use Insights 2019, 12, 1179173X19849419. [Google Scholar] [CrossRef] [PubMed]
- RRubin, J.B.; Lagas, J.S.; Broestl, L.; Sponagel, J.; Rockwell, N.; Rhee, G.; Rosen, S.F.; Chen, S.; Klein, R.S.; Imoukhuede, P.; et al. Sex differences in cancer mechanisms. Biol. Sex Differ. 2020, 11, 1–29. [Google Scholar] [CrossRef] [Green Version]
- Donovan, P.; Dubey, O.A.; Kallioinen, S.; Rogers, K.W.; Muehlethaler, K.; Müller, P.; Rimoldi, D.; Constam, D.B. Paracrine Activin-A Signaling Promotes Melanoma Growth and Metastasis through Immune Evasion. J. Investig. Dermatol. 2017, 137, 2578–2587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caponnetto, S.; Iannantuono, G.M.; Barchiesi, G.; Magri, V.; Gelibter, A.; Cortesi, E. Prolactin as a Potential Early Predictive Factor in Metastatic Non-Small Cell Lung Cancer Patients Treated with Nivolumab. Oncology 2017, 93, 62–66. [Google Scholar] [CrossRef]
- Yang, C.; Jin, J.; Yang, Y.; Sun, H.; Wu, L.; Shen, M.; Hong, X.; Li, W.; Lu, L.; Cao, D.; et al. Androgen receptor-mediated CD8+ T cell stemness programs drive sex differences in antitumor immunity. Immunity 2022, 55, 1268–1283.e9. [Google Scholar] [CrossRef]
- Kwon, H.; Schafer, J.M.; Song, N.-J.; Kaneko, S.; Li, A.; Xiao, T.; Ma, A.; Allen, C.; Das, K.; Zhou, L.; et al. Androgen conspires with the CD8(+) T cell exhaustion program and contributes to sex bias in cancer. Sci. Immunol. 2022, 7, eabq2630. [Google Scholar] [CrossRef]
- Phillips, D.C.; Buchanan, F.G.; Cheng, D.; Solomon, L.R.; Xiao, Y.; Xue, J.; Tahir, S.K.; Smith, M.L.; Zhang, H.; Widomski, D.; et al. Hexavalent TRAIL Fusion Protein Eftozanermin Alfa Optimally Clusters Apoptosis-Inducing TRAIL Receptors to Induce On-Target Antitumor Activity in Solid Tumors. Cancer Res. 2021, 81, 3402–3414. [Google Scholar] [CrossRef]
- Sun, A.; Luo, Y.; Xiao, W.; Zhu, Z.; Yan, H.; Miao, C.; Zhang, W.; Bai, P.; Liu, C.; Yang, D.; et al. Androgen Receptor Transcriptionally Inhibits Programmed Death Ligand-1 Expression and Influences Immune Escape in Bladder Cancer. Lab. Investig. 2023, 103, 100148. [Google Scholar] [CrossRef]
- Jiang, G.; Shi, L.; Zheng, X.; Zhang, X.; Wu, K.; Liu, B.; Yan, P.; Liang, X.; Yu, T.; Wang, Y.; et al. Androgen receptor affects the response to immune checkpoint therapy by suppressing PD-L1 in hepatocellular carcinoma. Aging 2020, 12, 11466–11484. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Cho, H.J.; Stranger, B.E.; Huang, R.S. Sex dimorphism in response to targeted therapy and immunotherapy in non-small cell lung cancer patients: A narrative review. Transl. Lung Cancer Res. 2022, 11, 920–934. [Google Scholar] [CrossRef] [PubMed]
- Bracken-Clarke, D.; Kapoor, D.; Baird, A.M.; Buchanan, P.J.; Gately, K.; Cuffe, S.; Finn, S.P. Vaping and lung cancer—A review of current data and recommendations. Lung Cancer 2021, 153, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Sepich-Poore, G.D.; Zitvogel, L.; Straussman, R.; Hasty, J.; Wargo, J.A.; Knight, R. The microbiome and human cancer. Science 2021, 371, eabc4552. [Google Scholar] [CrossRef]
- Liu, L.; Shah, K. The Potential of the Gut Microbiome to Reshape the Cancer Therapy Paradigm: A Review. JAMA Oncol. 2022, 8, 1059–1067. [Google Scholar] [CrossRef]
- Markle, J.G.M.; Frank, D.N.; Mortin-Toth, S.; Robertson, C.E.; Feazel, L.M.; Rolle-Kampczyk, U.; von Bergen, M.; McCoy, K.D.; Macpherson, A.J.; Danska, J.S. Sex differences in the gut microbiome drive hormone-dependent regulation of autoimmunity. Science 2013, 339, 1084–1088. [Google Scholar] [CrossRef] [Green Version]
- Mao, Q.; Jiang, F.; Yin, R.; Wang, J.; Xia, W.; Dong, G.; Ma, W.; Yang, Y.; Xu, L.; Hu, J. Interplay between the lung microbiome and lung cancer. Cancer Lett. 2018, 415, 40–48. [Google Scholar] [CrossRef]
- Apopa, P.L.; Alley, L.; Penney, R.B.; Arnaoutakis, K.; Steliga, M.A.; Jeffus, S.; Bircan, E.; Gopalan, B.; Jin, J.; Patumcharoenpol, P.; et al. PARP1 Is Up-Regulated in Non-small Cell Lung Cancer Tissues in the Presence of the Cyanobacterial Toxin Microcystin. Front. Microbiol. 2018, 9, 1757. [Google Scholar] [CrossRef] [Green Version]
- Routy, B.; le Chatelier, E.; DeRosa, L.; Duong, C.P.M.; Alou, M.T.; Daillère, R.; Fluckiger, A.; Messaoudene, M.; Rauber, C.; Roberti, M.P.; et al. Gut microbiome influences efficacy of PD-1-based immunotherapy against epithelial tumors. Science 2018, 359, 91–97. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Sex differences in lung cancer growth and treatment. (A) There are distinct differences in lung cancer biology between men and women. Men have an overall increase in mortality from lung cancer and an increase in tumor grade at diagnosis compared to women. Due to increases in mutations in the EGFR pathway, women have a better response to EGFR inhibitors than men. Women also have a better response to chemotherapies compared to men, while men have a better response to immunotherapies compared to women. Some of these differences could be due to sex hormones. Estrogens can improve some aspects of the immune response to cancer; however, they can also promote tumor growth. Conversely, androgens suppress the T cell response, possibly contributing to lung cancer in men. (B) Several sex-relevant considerations must be accounted for when completing lung cancer clinical trials. These sex-relevant variables can impact treatment outcomes as described in this review. These include a sex-specific mutation status for tumors from men vs. women, the hormone status of both males and females, the inherent differences in therapy outcomes due to sex hormones, and any impacts which could result from gender affirmation therapies. (C) Estrogen is the best characterized sex hormone. It has several roles in both promoting and inhibiting lung cancer. Estrogens are a mitogen which promote (red arrow) the cell autonomous proliferation of lung cancer cells, a pro-cancer effect. Estrogens also enhance (green arrow) the cell autonomous response of cancer cells to cytotoxic chemotherapies. Lastly, there is significant literature that shows that the anti-tumor immune response is modulated by estrogen (green arrow), which could explain why women have a lower incidence and mortality for many cancers, not just lung cancer. Figure was created using Biorender.
Figure 1.
Sex differences in lung cancer growth and treatment. (A) There are distinct differences in lung cancer biology between men and women. Men have an overall increase in mortality from lung cancer and an increase in tumor grade at diagnosis compared to women. Due to increases in mutations in the EGFR pathway, women have a better response to EGFR inhibitors than men. Women also have a better response to chemotherapies compared to men, while men have a better response to immunotherapies compared to women. Some of these differences could be due to sex hormones. Estrogens can improve some aspects of the immune response to cancer; however, they can also promote tumor growth. Conversely, androgens suppress the T cell response, possibly contributing to lung cancer in men. (B) Several sex-relevant considerations must be accounted for when completing lung cancer clinical trials. These sex-relevant variables can impact treatment outcomes as described in this review. These include a sex-specific mutation status for tumors from men vs. women, the hormone status of both males and females, the inherent differences in therapy outcomes due to sex hormones, and any impacts which could result from gender affirmation therapies. (C) Estrogen is the best characterized sex hormone. It has several roles in both promoting and inhibiting lung cancer. Estrogens are a mitogen which promote (red arrow) the cell autonomous proliferation of lung cancer cells, a pro-cancer effect. Estrogens also enhance (green arrow) the cell autonomous response of cancer cells to cytotoxic chemotherapies. Lastly, there is significant literature that shows that the anti-tumor immune response is modulated by estrogen (green arrow), which could explain why women have a lower incidence and mortality for many cancers, not just lung cancer. Figure was created using Biorender.


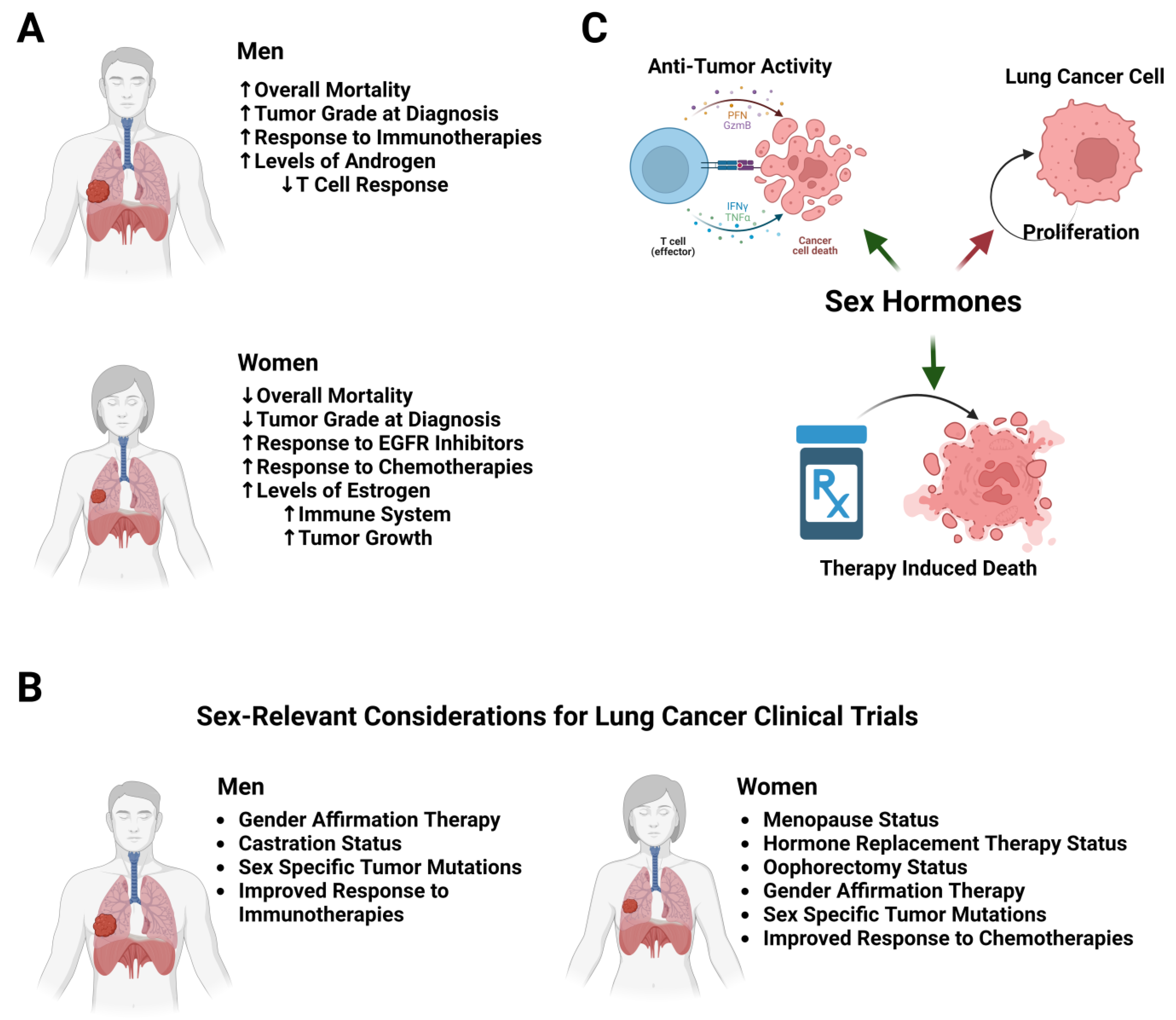{kind=link}
Table 1.
Table describing sex hormones and their effects on lung cancer growth. Sex hormones and bioactive molecules have known functions in regulating several aspects of lung cancer biology. These include the cell autonomous effects of lung cancer such as cell proliferation, cell death responses to therapy, and the ability of cancer cells to metastasize. These molecules have significant effects on the immune system, including modulating T cell activity, immunoglobulin production, and innate immune cell responses (cytokine release).
Table 1.
Table describing sex hormones and their effects on lung cancer growth. Sex hormones and bioactive molecules have known functions in regulating several aspects of lung cancer biology. These include the cell autonomous effects of lung cancer such as cell proliferation, cell death responses to therapy, and the ability of cancer cells to metastasize. These molecules have significant effects on the immune system, including modulating T cell activity, immunoglobulin production, and innate immune cell responses (cytokine release).
| Hormone | Higher in… | Role in Lung Cancer | Effects on Immune System |
|---|---|---|---|
| Estrogen | Females |
| |
| Progesterone | Females |
| |
| Testosterone | Males |
|
|
| Follicular Stimulating Hormone | Females |
|
|
| Leptin | Females |
| |
| Prolactin | Females |
|
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
May, L.; Shows, K.; Nana-Sinkam, P.; Li, H.; Landry, J.W. Sex Differences in Lung Cancer. Cancers 2023, 15, 3111. https://doi.org/10.3390/cancers15123111
AMA Style
May L, Shows K, Nana-Sinkam P, Li H, Landry JW. Sex Differences in Lung Cancer. Cancers. 2023; 15(12):3111. https://doi.org/10.3390/cancers15123111
Chicago/Turabian StyleMay, Lauren, Kathryn Shows, Patrick Nana-Sinkam, Howard Li, and Joseph W. Landry. 2023. "Sex Differences in Lung Cancer" Cancers 15, no. 12: 3111. https://doi.org/10.3390/cancers15123111
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.