
Influence of the Hypoxia-Activated Prodrug Evofosfamide (TH-302) on Glycolytic Metabolism of Canine Glioma: A Potential Improvement in Cancer Metabolism
 , , and
, , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Medical Record Review and Patient Selection
2.2. Immunohistochemical Analyses (IHC)
2.3. Cell Lines and Cultures
2.4. Protein Assay
2.5. Reagents
2.6. Cell Viability and Cytotoxicity Assay
2.7. Measurement of Cellular ATP and Lactate Levels
2.8. Experimental Animals
2.9. Murine Xenograft Models
2.10. Micro-CT Imaging
2.11. Biochemical Parameters
2.12. Metabolome Analysis
2.13. Statistical Analysis
3. Results
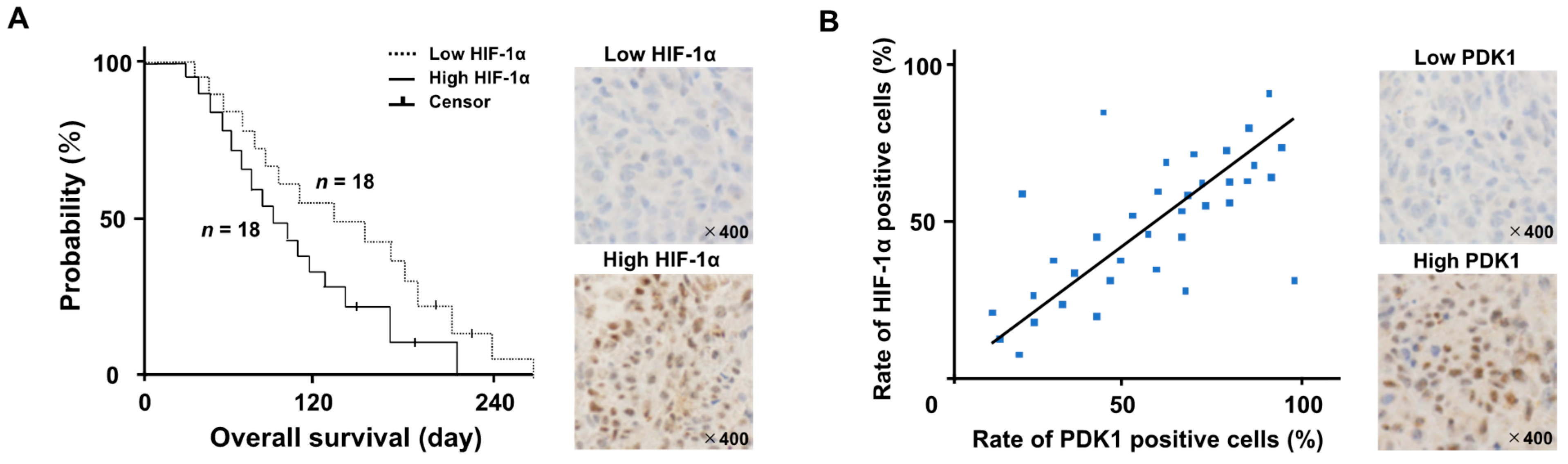
3.1. Patient Demographics and Treatment Outcomes
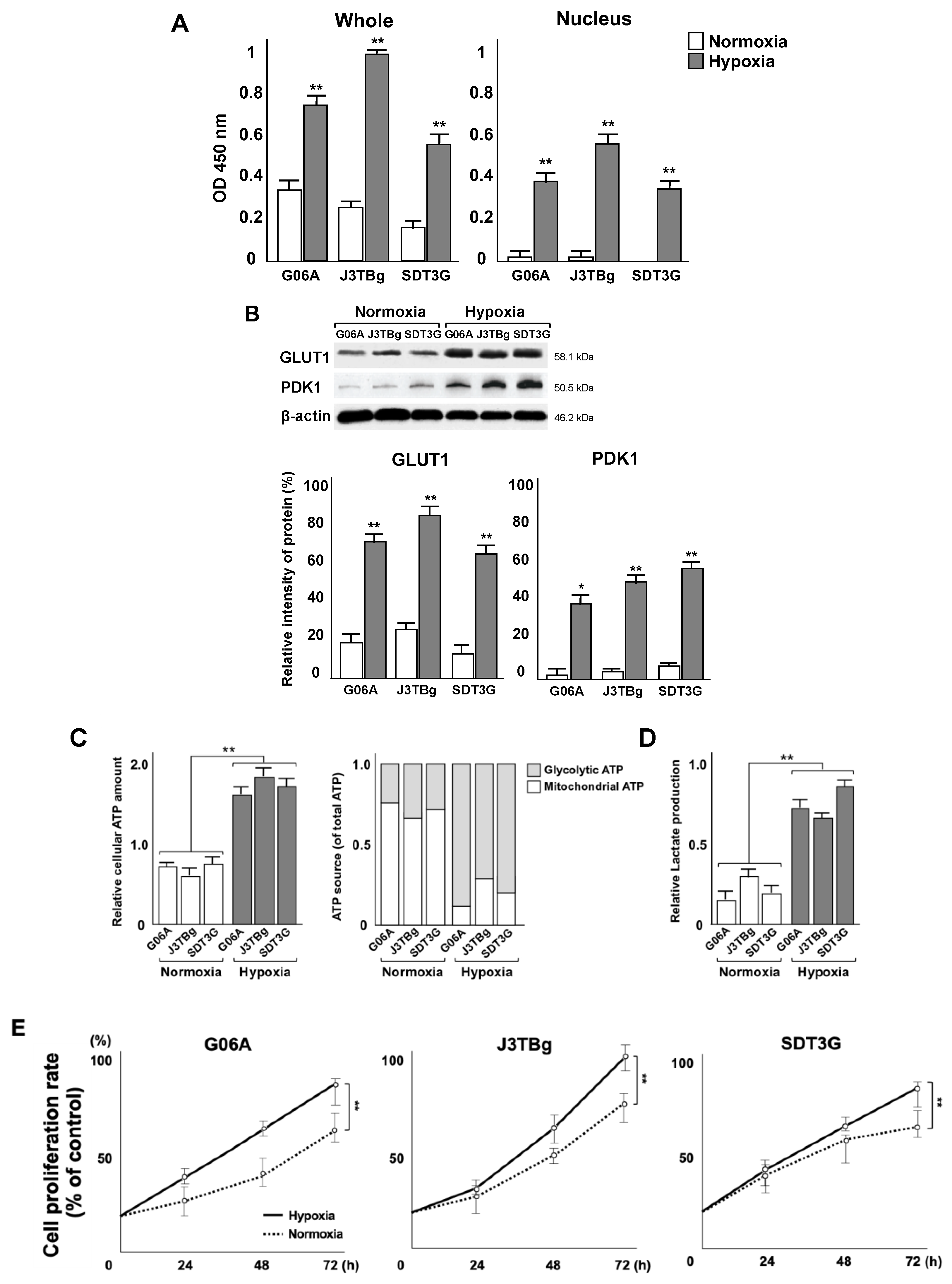
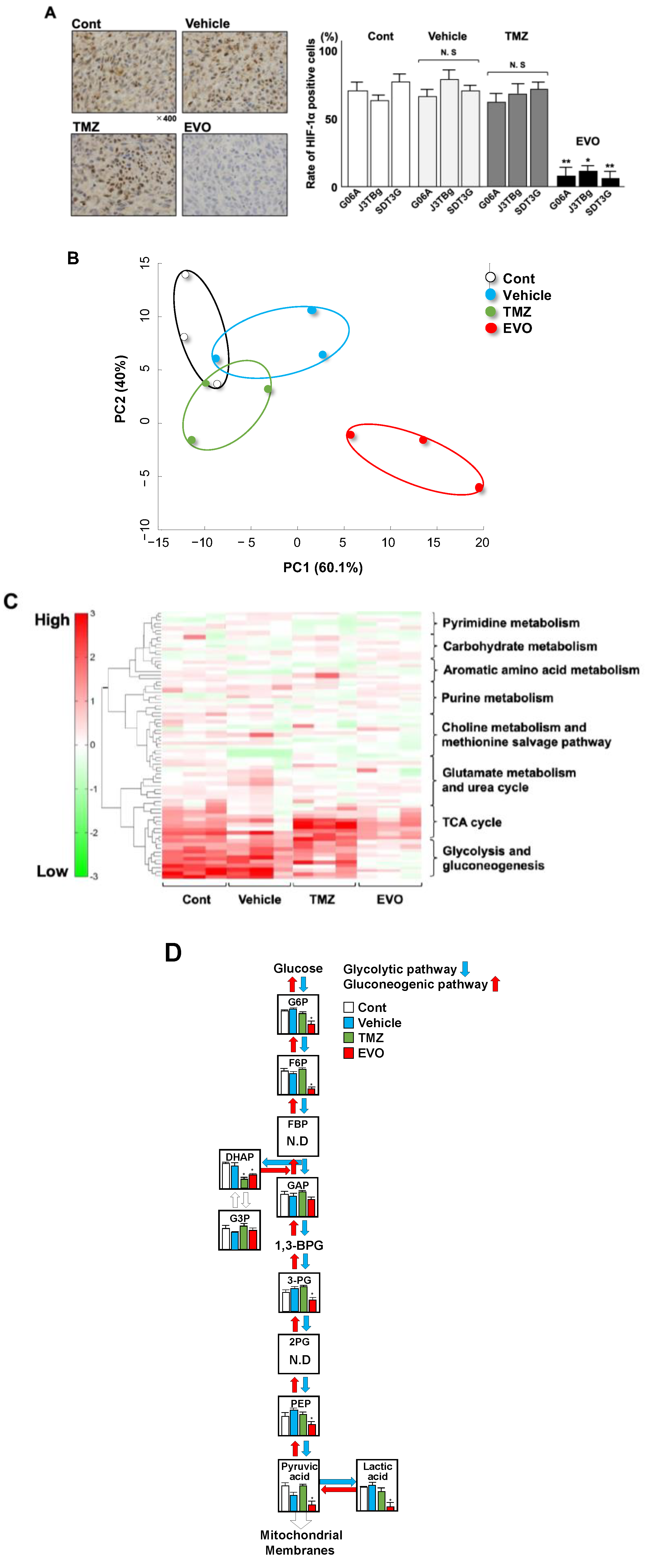
3.2. Localization of HIF-1α Protein and Metabolic Shifts
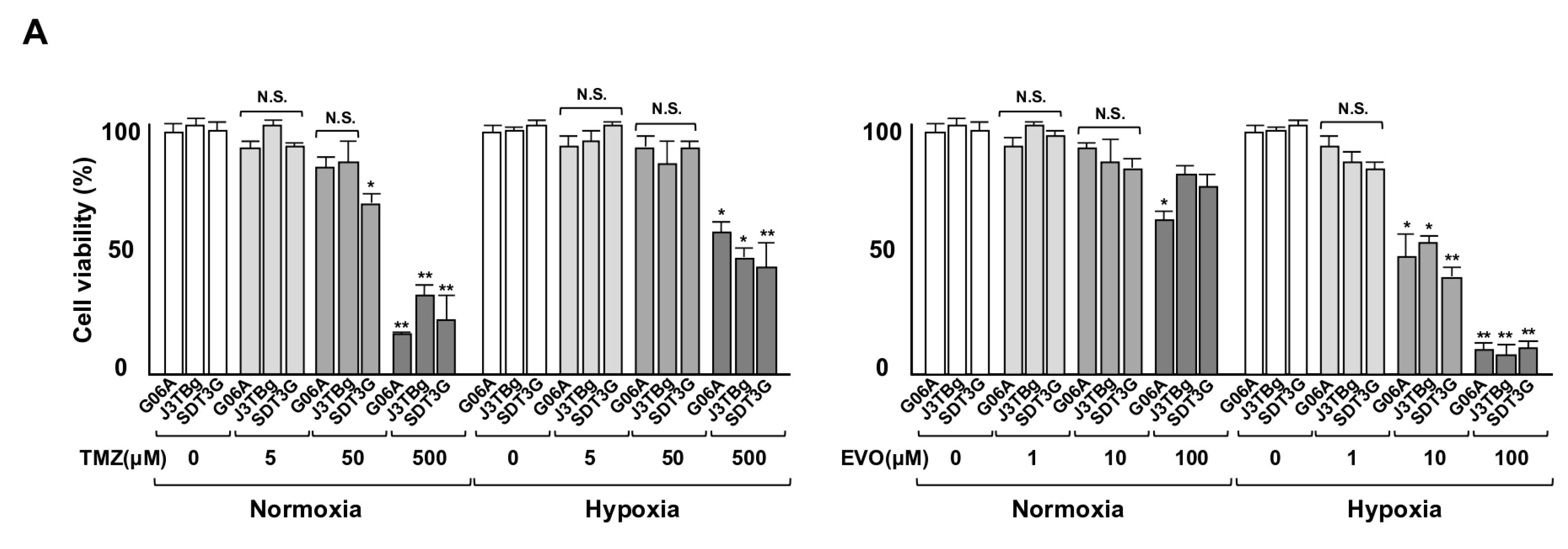
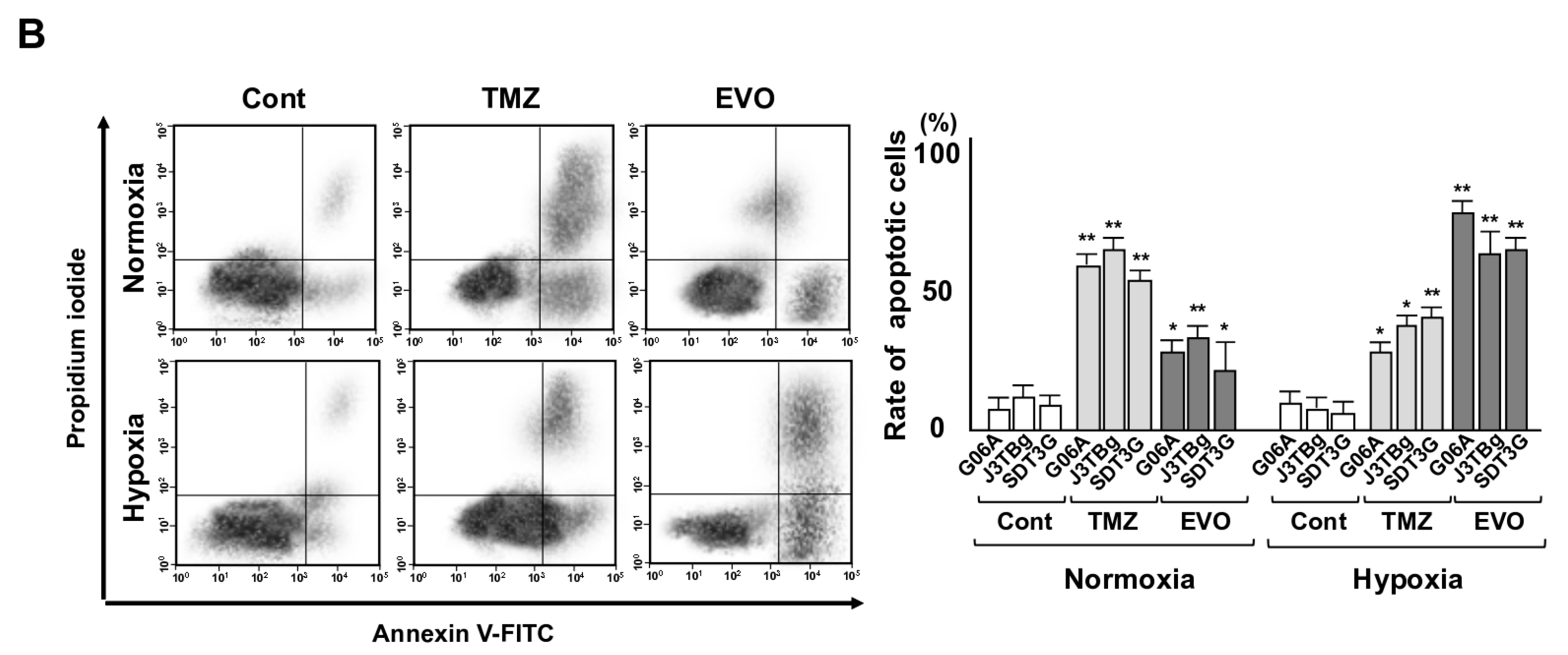
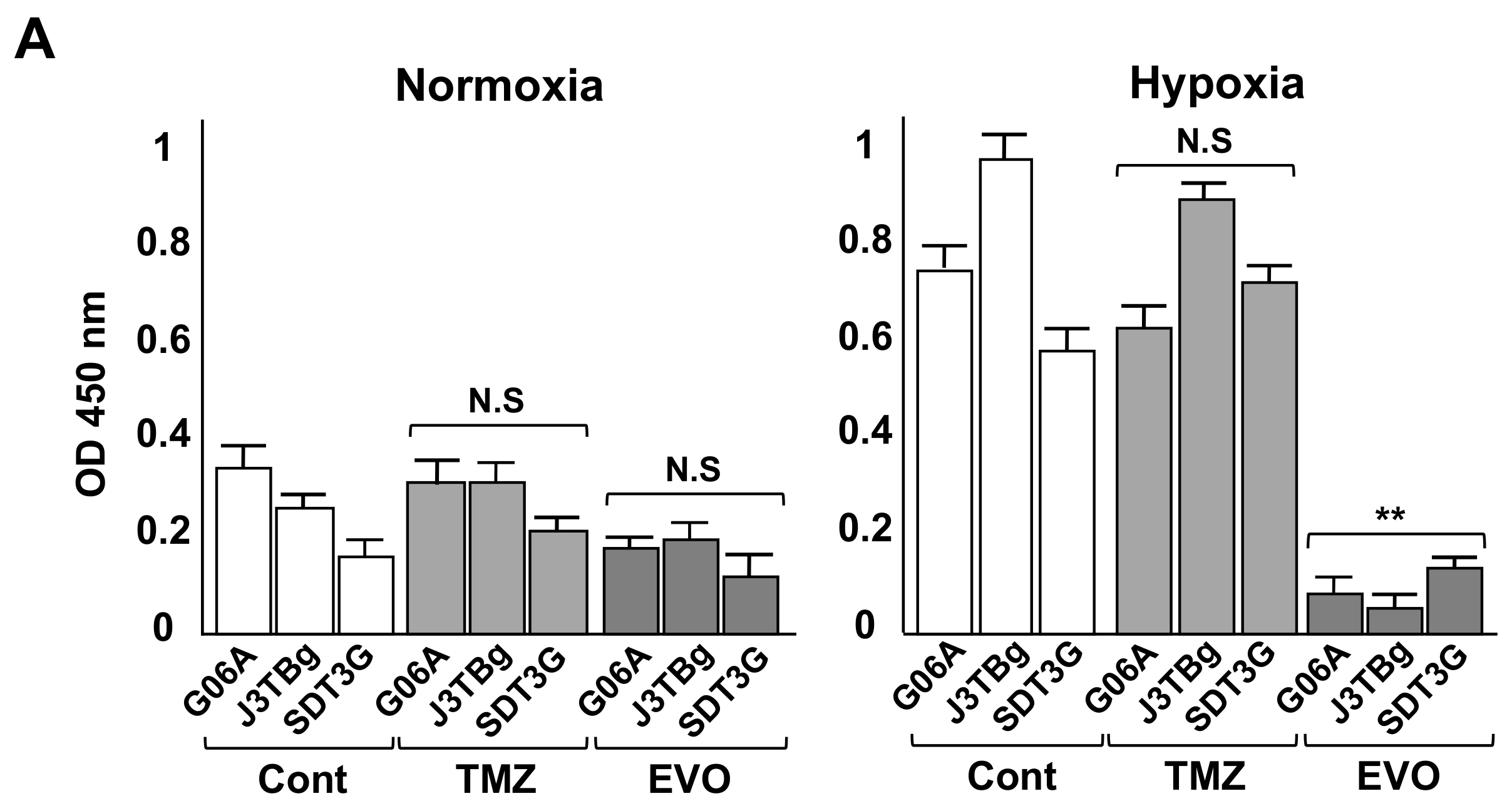
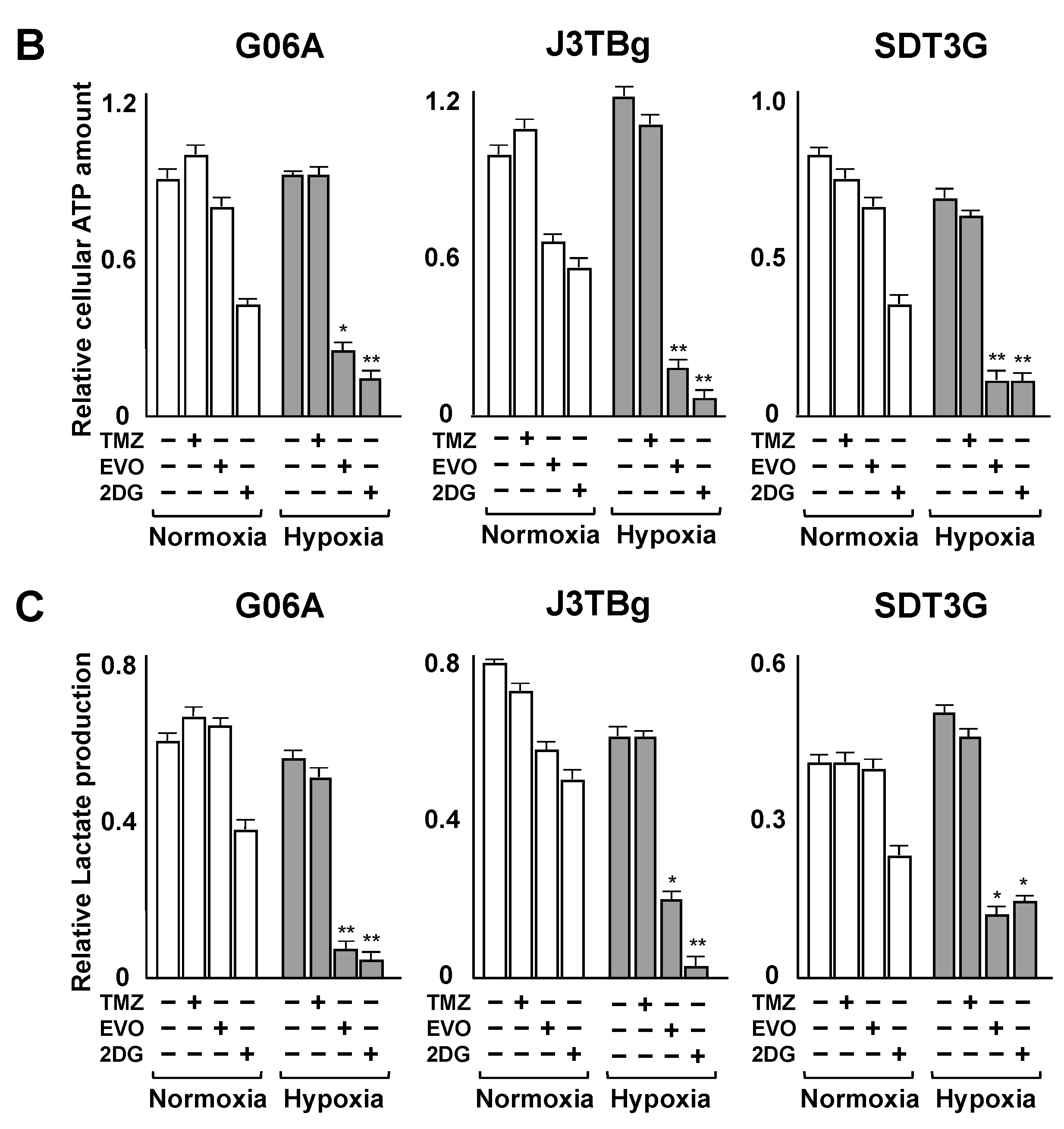
3.3. Cytotoxicity and Glycolysis Suppression
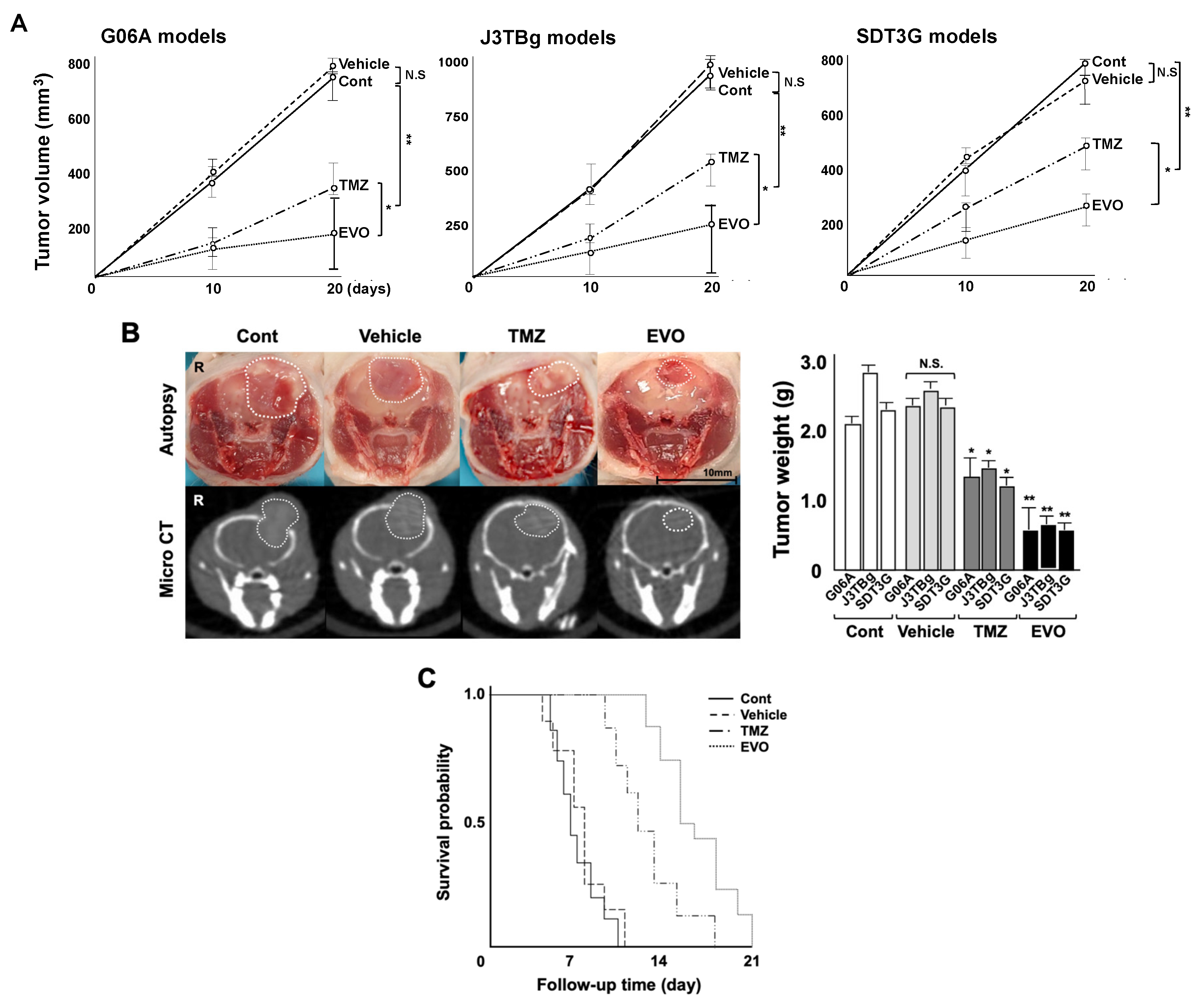
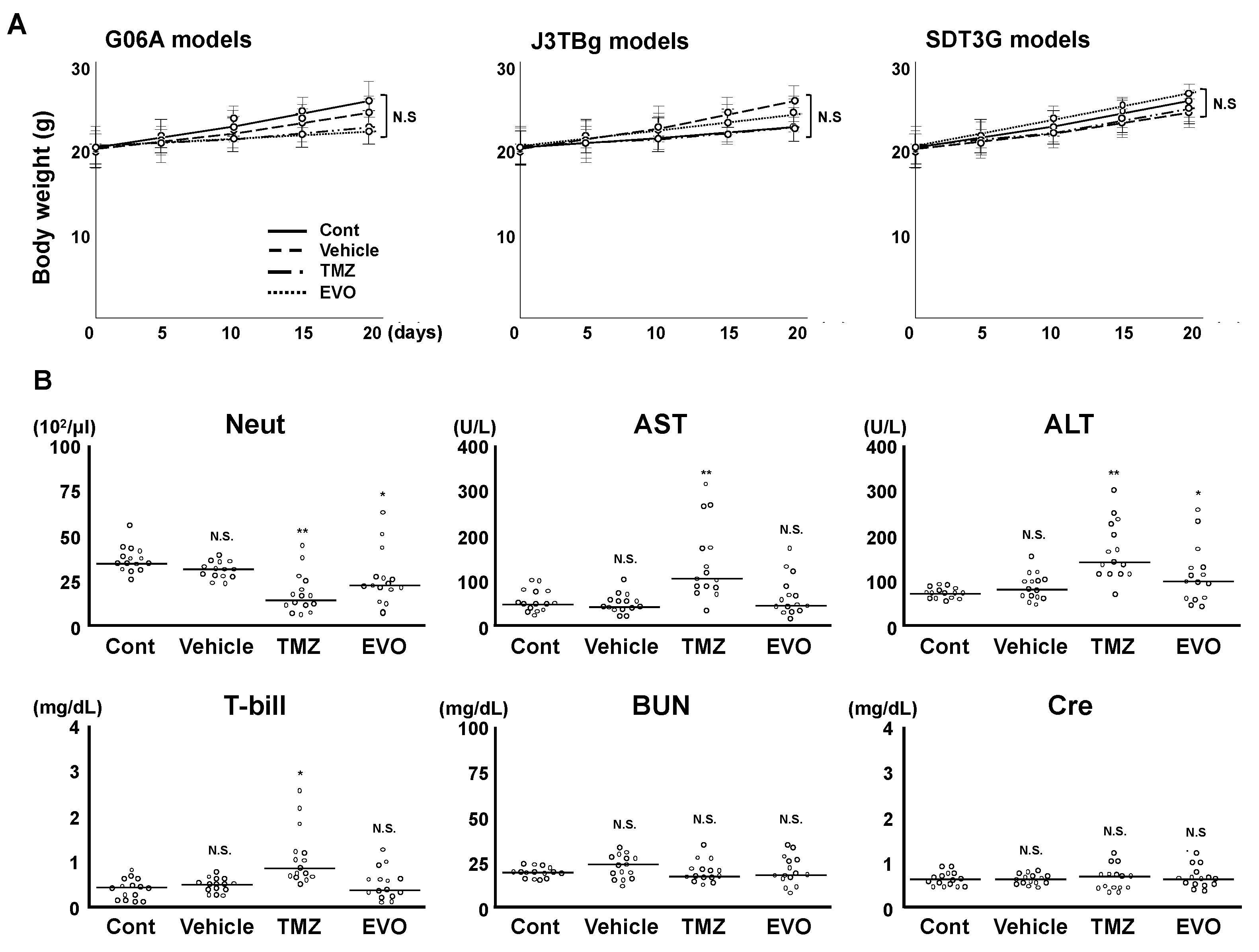
3.4. Antitumor Effects and Adverse Events
3.5. Levels of HIF-1α Protein and Influence on Tumor Metabolism
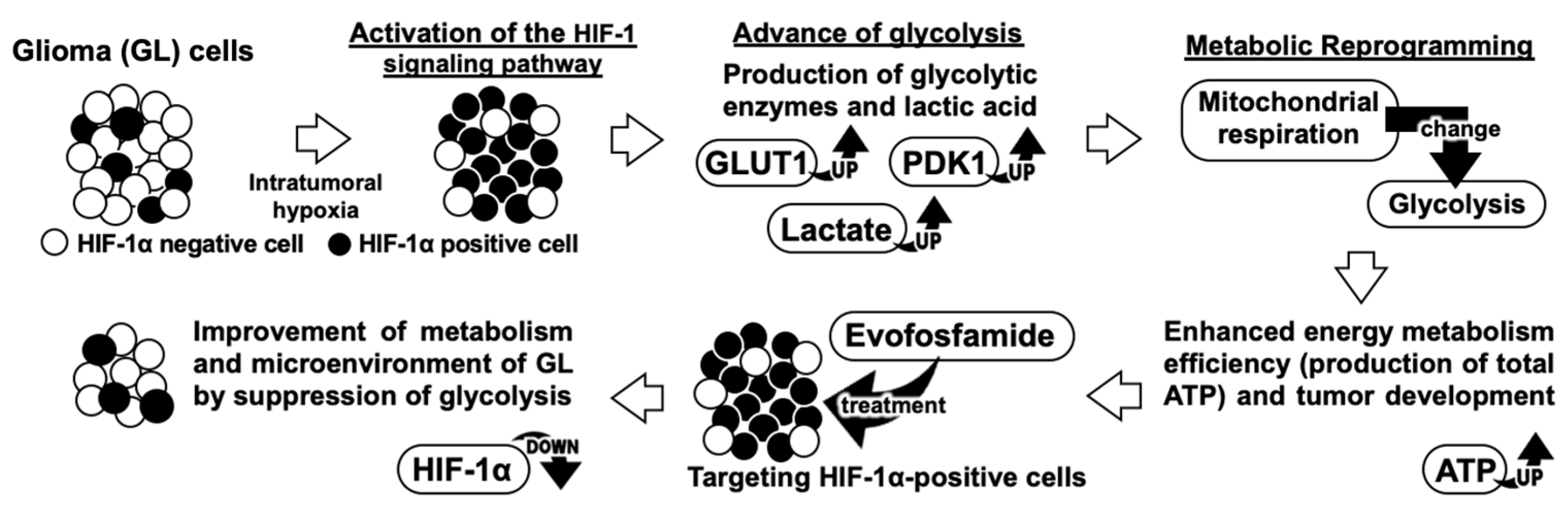
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Herranz, C.; Fernández, F.; Martín-Ibáñez, R.; Blasco, E.; Crespo, E.; De la Fuente, C.; Añor, S.; Rabanal, R.M.; Canals, J.M.; Pumarola, M. Spontaneously Arising Canine Glioma as a Potential Model for Human Glioma. J. Comp. Pathol. 2016, 154, 169–179. [Google Scholar] [CrossRef] [PubMed]
- Hubbard, M.E.; Arnold, S.; Bin Zahid, A.; McPheeters, M.; O’Sullivan, M.G.; Tabaran, A.F.; Hunt, M.A.; Pluhar, G.E. Naturally Occurring Canine Glioma as a Model for Novel Therapeutics. Cancer. Investig. 2018, 36, 415–423. [Google Scholar] [CrossRef] [PubMed]
- Ostrom, Q.T.; Cote, D.J.; Ascha, M.; Kruchko, C.; Barnholtz-Sloan, J.S. Adult Glioma Incidence and Survival by Race or Ethnicity in the United States From 2000 to 2014. JAMA Oncol. 2018, 4, 1254–1262. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Zhang, C.; Zhu, J.; Zhang, L.; Chen, H.; Qian, J.; Luo, C. Crucial Role of RLIP76 in Promoting Glycolysis and Tumorigenesis by Stabilization of HIF-1α in Glioma Cells Under Hypoxia. Mol. Neurobiol. 2022, 59, 6724–6739. [Google Scholar] [CrossRef] [PubMed]
- Lai, J.H.; Jan, H.J.; Liu, L.W.; Lee, C.C.; Wang, S.G.; Hueng, D.Y.; Cheng, Y.Y.; Lee, H.M.; Ma, H.I. Nodal regulates energy metabolism in glioma cells by inducing expression of hypoxia-inducible factor 1α. Neuro-Oncol. 2013, 15, 1330–1341. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Huang, H.; Jia, C.H.; Fan, K.; Xie, T.; Zhu, Z.Y.; Xie, M.L. Apigenin increases radiosensitivity of glioma stem cells by attenuating HIF-1α-mediated glycolysis. Med. Oncol. 2021, 38, 131. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.P.; Liu, Y.; Xiao, L.M.; Chen, L.K.; Tao, E.X.; Zeng, E.M.; Xu, C.H. Induction of cancer cell stemness in glioma through glycolysis and the long noncoding RNA HULC-activated FOXM1/AGR2/HIF-1α axis. Lab. Investig. 2022, 102, 691–701. [Google Scholar] [CrossRef] [PubMed]
- Jia, C.; Zhao, Y.; Huang, H.; Fan, K.; Xie, T.; Xie, M. Apigenin sensitizes radiotherapy of mouse subcutaneous glioma through attenuations of cell stemness and DNA damage repair by inhibiting NF-κB/HIF-1α-mediated glycolysis. J. Nutr. Biochem. 2022, 107, 109038. [Google Scholar] [CrossRef] [PubMed]
- Velpula, K.K.; Bhasin, A.; Asuthkar, S.; Tsung, A.J. Combined targeting of PDK1 and EGFR triggers regression of glioblastoma by reversing the Warburg effect. Cancer Res. 2013, 73, 7277–7289. [Google Scholar] [CrossRef]
- Shen, H.; Hau, E.; Joshi, S.; Dilda, P.J.; McDonald, K.L. Sensitization of Glioblastoma Cells to Irradiation by Modulating the Glucose Metabolism. Mol. Cancer Ther. 2015, 14, 1794–1804. [Google Scholar] [CrossRef]
- Brenner, A.; Zuniga, R.; Sun, J.D.; Floyd, J.; Hart, C.P.; Kroll, S.; Fichtel, L.; Cavazos, D.; Caflisch, L.; Gruslova, A.; et al. Hypoxia-activated evofosfamide for treatment of recurrent bevacizumab-refractory glioblastoma: A phase I surgical study. Neuro. Oncol. 2018, 20, 1231–1239. [Google Scholar] [CrossRef] [PubMed]
- Brenner, A.J.; Floyd, J.; Fichtel, L.; Michalek, J.; Kanakia, K.P.; Huang, S.; Reardon, D.; Wen, P.Y.; Lee, E.Q. Phase 2 trial of hypoxia activated evofosfamide (TH302) for treatment of recurrent bevacizumab-refractory glioblastoma. Sci. Rep. 2021, 11, 2306. [Google Scholar] [CrossRef] [PubMed]
- Yamazaki, H.; Lai, Y.C.; Tateno, M.; Setoguchi, A.; Goto-Koshino, Y.; Endo, Y.; Nakaichi, M.; Tsujimoto, H.; Miura, N. Hypoxia-activated prodrug TH-302 decreased survival rate of canine lymphoma cells under hypoxic condition. PLoS ONE 2017, 12, e01773052017. [Google Scholar] [CrossRef] [PubMed]
- Yamazaki, H.; Tanaka, T.; Nishida, H.; Hatoya, S.; Akiyoshi, H. Hypoxia-targeting therapy for intestinal T-cell lymphoma in dogs: Preclinical study using 3D in vitro models. Vet. Comp. Oncol. 2023, 21, 12–19. [Google Scholar] [CrossRef]
- Yamazaki, H.; Tanaka, T.; Nishida, H.; Hatoya, S.; Akiyoshi, H. Assessment of hypoxia-targeting therapy for gastrointestinal lymphoma in dogs: Preclinical test using murine models. Res. Vet. Sci. 2023, 154, 22–28. [Google Scholar] [CrossRef] [PubMed]
- Portwood, S.; Lal, D.; Hsu, Y.C.; Vargas, R.; Johnson, M.K.; Wetzler, M.; Hart, C.P.; Wang, E.S. Activity of the hypoxia-activated prodrug, TH-302, in preclinical human acute myeloid leukemia models. Clin. Cancer Res. 2013, 19, 6506–6519. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Tian, Y.; Zhao, Y.; Xue, C.; Zhan, J.; Liu, L.; He, X.; Zhang, L. Efficacy of the hypoxia-activated prodrug evofosfamide (TH-302) in nasopharyngeal carcinoma in vitro and in vivo. Cancer Commun. 2018, 38, 15. [Google Scholar] [CrossRef]
- Yoon, C.; Lee, H.J.; Park, D.J.; Lee, Y.J.; Tap, W.D.; Eisinger-Mathason, T.S.; Hart, C.P.; Choy, E.; Simon, M.C.; Yoon, S.S. Hypoxia-activated chemotherapeutic TH-302 enhances the effects of VEGF-A inhibition and radiation on sarcomas. Br. J. Cancer 2015, 113, 46–56. [Google Scholar] [CrossRef]
- Liu, F.; Zhou, Q.; Jiang, H.F.; Zhang, T.T.; Miao, C.; Xu, X.H.; Wu, J.X.; Yin, S.L.; Xu, S.J.; Peng, J.Y.; et al. Piperlongumine conquers temozolomide chemoradiotherapy resistance to achieve immune cure in refractory glioblastoma via boosting oxidative stress-inflamation-CD8+-T cell immunity. J. Exp. Clin. Cancer Res. 2023, 42, 118. [Google Scholar] [CrossRef]
- Gravina, G.L.; Colapietro, A.; Mancini, A.; Rossetti, A.; Martellucci, S.; Ventura, L.; Di Franco, M.; Marampon, F.; Mattei, V.; Biordi, L.A.; et al. ATX-101, a Peptide Targeting PCNA, Has Antitumor Efficacy Alone or in Combination with Radiotherapy in Murine Models of Human Glioblastoma. Cancers 2022, 14, 289. [Google Scholar] [CrossRef]
- Wei, S.; Yin, D.; Yu, S.; Lin, X.; Savani, M.R.; Du, K.; Ku, Y.; Wu, D.; Li, S.; Liu, H.; et al. Antitumor Activity of a Mitochondrial-Targeted HSP90 Inhibitor in Gliomas. Clin. Cancer Res. 2022, 28, 2180–2195. [Google Scholar] [CrossRef]
- Brauer, H.A.; Makowski, L.; Hoadley, K.A.; Casbas-Hernandez, P.; Lang, L.J.; Romàn-Pèrez, E.; D’Arcy, M.; Freemerman, A.J.; Perou, C.M.; Troester, M.A. Impact of tumor microenvironment and epithelial phenotypes on metabolism in breast cancer. Clin. Cancer Res. 2013, 19, 571–585. [Google Scholar] [CrossRef] [PubMed]
- Ikari, R.; Mukaisho, K.I.; Kageyama, S.; Nagasawa, M.; Kubota, S.; Nakayama, T.; Murakami, S.; Taniura, N.; Tanaka, H.; Kushima, R.P.; et al. Differences in the central energy metabolism of cancer cells between conventional 2D and novel 3D culture systems. Int. J. Mol. Sci. 2021, 22, 1805. [Google Scholar] [CrossRef] [PubMed]
- Rashid, M.; Zadeh, L.R.; Baradaran, B.; Molavi, O.; Ghesmati, Z.; Sabzichi, M.; Ramezani, F. Up-down regulation of HIF-1α in cancer progression. Gene 2021, 798, 145796. [Google Scholar] [CrossRef] [PubMed]
- Agnihotri, S.; Zadeh, G. Metabolic reprogramming in glioblastoma: The influence of cancer metabolism on epigenetics and unanswered questions. Neuro-Oncol. 2016, 18, 160–172. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Zhu, S.; Tong, J.; Hao, H.; Yang, J.; Liu, Z.; Wang, Y. Suppression of lactate dehydrogenase A compromises tumor progression by downregulation of the Warburg effect in glioblastoma. Neuroreport 2016, 27, 110–115. [Google Scholar] [CrossRef] [PubMed]
- Poff, A.; Koutnik, A.P.; Egan, K.M.; Sahebjam, S.; D’Agostino, D.; Kumar, N.B. Targeting the Warburg effect for cancer treatment: Ketogenic diets for management of glioma. Semin. Cancer Biol. 2019, 56, 135–148. [Google Scholar] [CrossRef] [PubMed]
- Benito, J.; Ramirez, M.S.; Millward, N.Z.; Velez, J.; Harutyunyan, K.G.; Lu, H.; Shi, Y.X.; Matre, P.; Jacamo, R.; Ma, H.; et al. Hypoxia-Activated Prodrug TH-302 Targets Hypoxic Bone Marrow Niches in Preclinical Leukemia Models. Clin. Cancer Res. 2016, 22, 1687–1698. [Google Scholar] [CrossRef]
- Marcucci, F.; Rumio, C. Glycolysis-induced drug resistance in tumors—A response to danger signals? Neoplasia 2021, 23, 234–245. [Google Scholar] [CrossRef]
- Zhang, J.; Chen, G.; Gao, Y.; Liang, H. HOTAIR/miR-125 axis-mediated Hexokinase 2 expression promotes chemoresistance in human glioblastoma. J. Cell. Mol. Med. 2020, 24, 5707–5717. [Google Scholar] [CrossRef]
- Branco, M.; Linhares, P.; Carvalho, B.; Santos, P.; Costa, B.M.; Vaz, R. Serum lactate levels are associated with glioma malignancy grade. Clin. Neurol. Neurosurg. 2019, 186, 105546. [Google Scholar] [CrossRef] [PubMed]
- Sitter, B.; Forsmark, A.; Solheim, O. Elevated Serum Lactate in Glioma Patients: Associated Factors. Front Oncol. 2022, 12, 831079. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Cao, P. Clinical and prognostic significance of HIF-1α in glioma patients: A meta-analysis. Int. J. Clin. Exp. Med. 2015, 8, 22073–22083. [Google Scholar] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | High HIF-1α (n = 18) | Low HIF-1α (n = 18) | p Value | |
|---|---|---|---|---|
| Age (year): median (range) | 10.6 (4.3–14.5) | 11.4 (6.4–16.5) | 0.774 | |
| Weight (kg): median (range) | 9.8 (2.4–42.3) | 8.3 (3.1–38.6) | 0.531 | |
| Sex (n) | Male | 10 (56%) | 11 (61%) | 0.469 |
| Female | 8 (44%) | 7 (39%) | 0.482 | |
| Tumor volume (cm3): median (range) | 1.56 (0.89–3.74) | 1.48 (0.82–3.86) | 0.085 | |
| Tumor type | Oligodendroglioma (14) | Oligodendroglioma (15) | 0.362 | |
| Astrocytoma (1) | Astrocytoma (1) | |||
| Undefined (3) | Undefined (2) | |||
| Tumor location | Hemispheric (15) | Hemispheric (14) | 0.545 | |
| Diencephalon (2) | Diencephalon (3) | |||
| Infratentorial (1) | Infratentorial (1) | |||
| Parietal (6) | Parietal (5) | 0.249 | ||
| Temporal (5) | Temporal (7) | |||
| Front-olfactory (7) | Front-olfactory (6) | |||
| Neurological sign (n) | 12 (67%) | 7 (39%) | 0.078 | |
| Group | OS; Median Days (Range) | Hazard Ratio | 95%CI | p-Value |
|---|---|---|---|---|
| Group 1 (Control) | 7.3 (6–10) | 1 | - | - |
| Group 2 (Vehicle) | 8.1 (5–11) | 1.12 | 0.82–1.36 | 0.802 |
| Group 3 (TMZ) | 11.8 (10–18) | 0.78 | 0.60–0.96 | 0.013 |
| Group 4 (EVO) | 16.0 (13–21) | 0.64 | 0.58–0.85 | 0.007 |
| Group 3 (TMZ) | - | 1 | - | - |
| Group 4 (EVO) | - | 0.82 | 0.73–0.92 | 0.024 |
| Pathway Cluster | Compound Name | Comparative Analysis (Ratio) | ||
|---|---|---|---|---|
| Vehicle * vs. Control | TMZ vs. Control | EVO vs. Control | ||
| Glycolysis and gluconeogenesis | Glucose 6-phosphate (G6P) | 0.81 | 0.45 | 8 × 10−3 * |
| Fructose 6-phosphate (F6P) | 0.71 | 0.78 | 3 × 10−3 * | |
| Dihydroxyacetone phosphate (DHAP) | 0.84 | 2 × 10−2 * | 3 × 10−2 * | |
| Glyceraldehyde-3-phosphate dehydrogenase (G3P) | 0.65 | 0.56 | 0.12 | |
| Glyceraldehyde-3-phosphate (GAP) | 0.34 | 0.67 | 0.21 | |
| 3-Phosphoglyceric acid (3-PG) | 0.54 | 0.48 | 4 × 10−3 * | |
| Phosphoenolpyruvic acid (PEP) | 0.38 | 0.21 | 6 × 10−3 * | |
| Pyruvic acid | 0.45 | 0.08 | 2 × 10−3 * | |
| Lactic acid | 0.39 | 0.55 | 3 × 10−3 * | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yamazaki, H.; Onoyama, S.; Gotani, S.; Deguchi, T.; Tamura, M.; Ohta, H.; Iwano, H.; Nishida, H.; Dickinson, P.J.; Akiyoshi, H. Influence of the Hypoxia-Activated Prodrug Evofosfamide (TH-302) on Glycolytic Metabolism of Canine Glioma: A Potential Improvement in Cancer Metabolism. Cancers 2023, 15, 5537. https://doi.org/10.3390/cancers15235537
Yamazaki H, Onoyama S, Gotani S, Deguchi T, Tamura M, Ohta H, Iwano H, Nishida H, Dickinson PJ, Akiyoshi H. Influence of the Hypoxia-Activated Prodrug Evofosfamide (TH-302) on Glycolytic Metabolism of Canine Glioma: A Potential Improvement in Cancer Metabolism. Cancers. 2023; 15(23):5537. https://doi.org/10.3390/cancers15235537
Chicago/Turabian StyleYamazaki, Hiroki, Seio Onoyama, Shunichi Gotani, Tatsuya Deguchi, Masahiro Tamura, Hiroshi Ohta, Hidetomo Iwano, Hidetaka Nishida, Peter J. Dickinson, and Hideo Akiyoshi. 2023. "Influence of the Hypoxia-Activated Prodrug Evofosfamide (TH-302) on Glycolytic Metabolism of Canine Glioma: A Potential Improvement in Cancer Metabolism" Cancers 15, no. 23: 5537. https://doi.org/10.3390/cancers15235537