
Prenatal Hyperhomocysteinemia Induces Glial Activation and Alters Neuroinflammatory Marker Expression in Infant Rat Hippocampus

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Animals
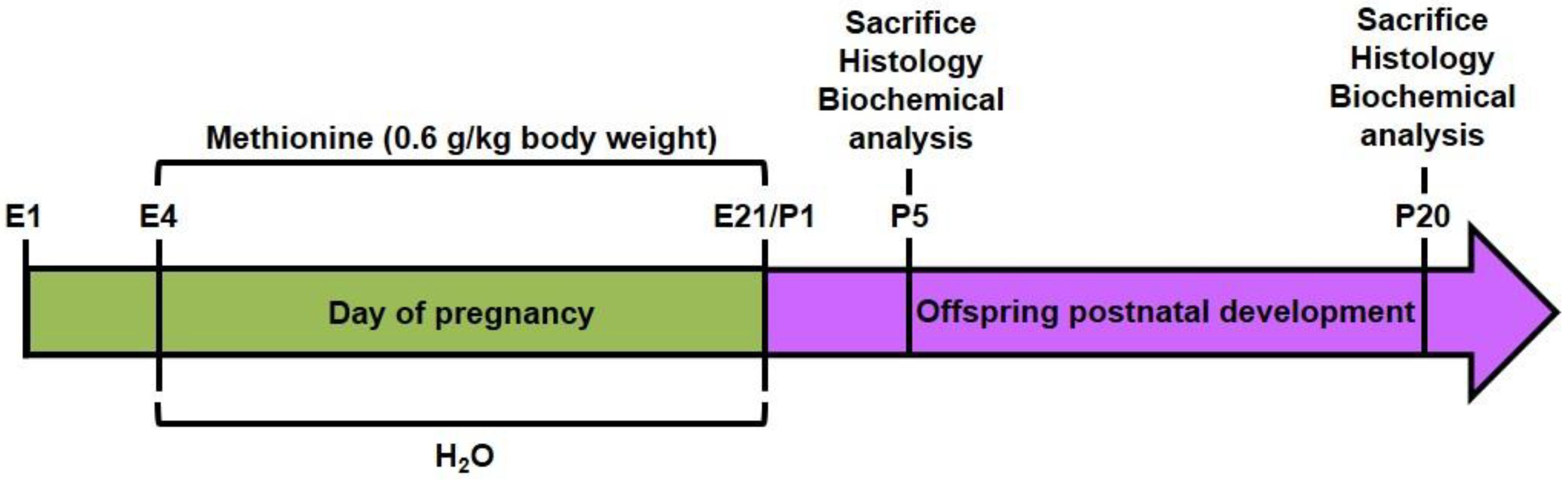
2.2. Chronic Methionine Treatment
2.3. Brain Tissue Preparation for Microscopy
2.4. Light Microscopy
2.5. Immunochemistry
2.6. Electron Microscopy
2.7. Cytokine Assay
2.8. Western Blot Analysis
2.9. Caspase-3 Activity
2.10. Acetylcholinesterase Activity
2.11. Statistical Analysis
3. Results
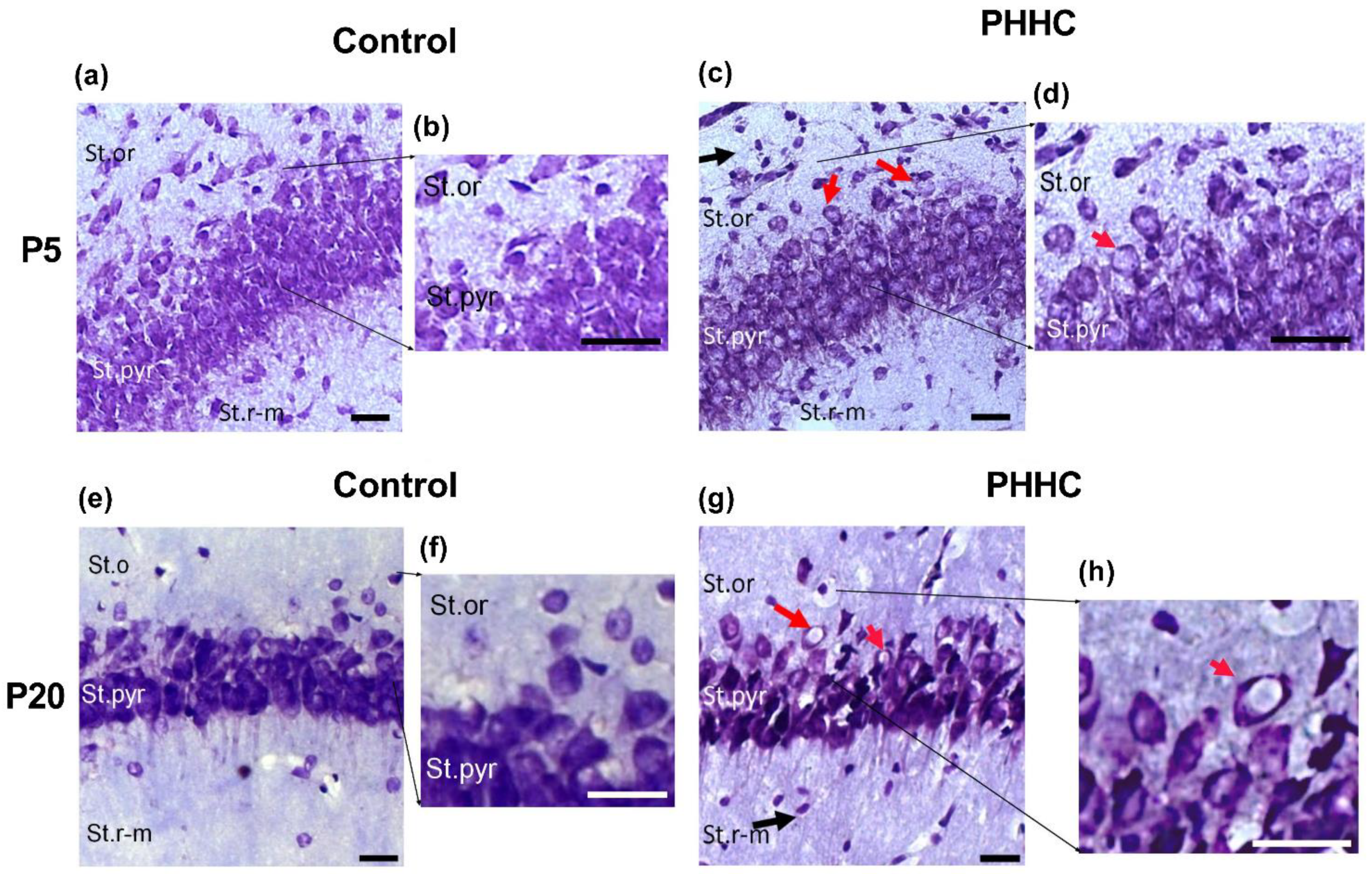
3.1. Light Microscopy
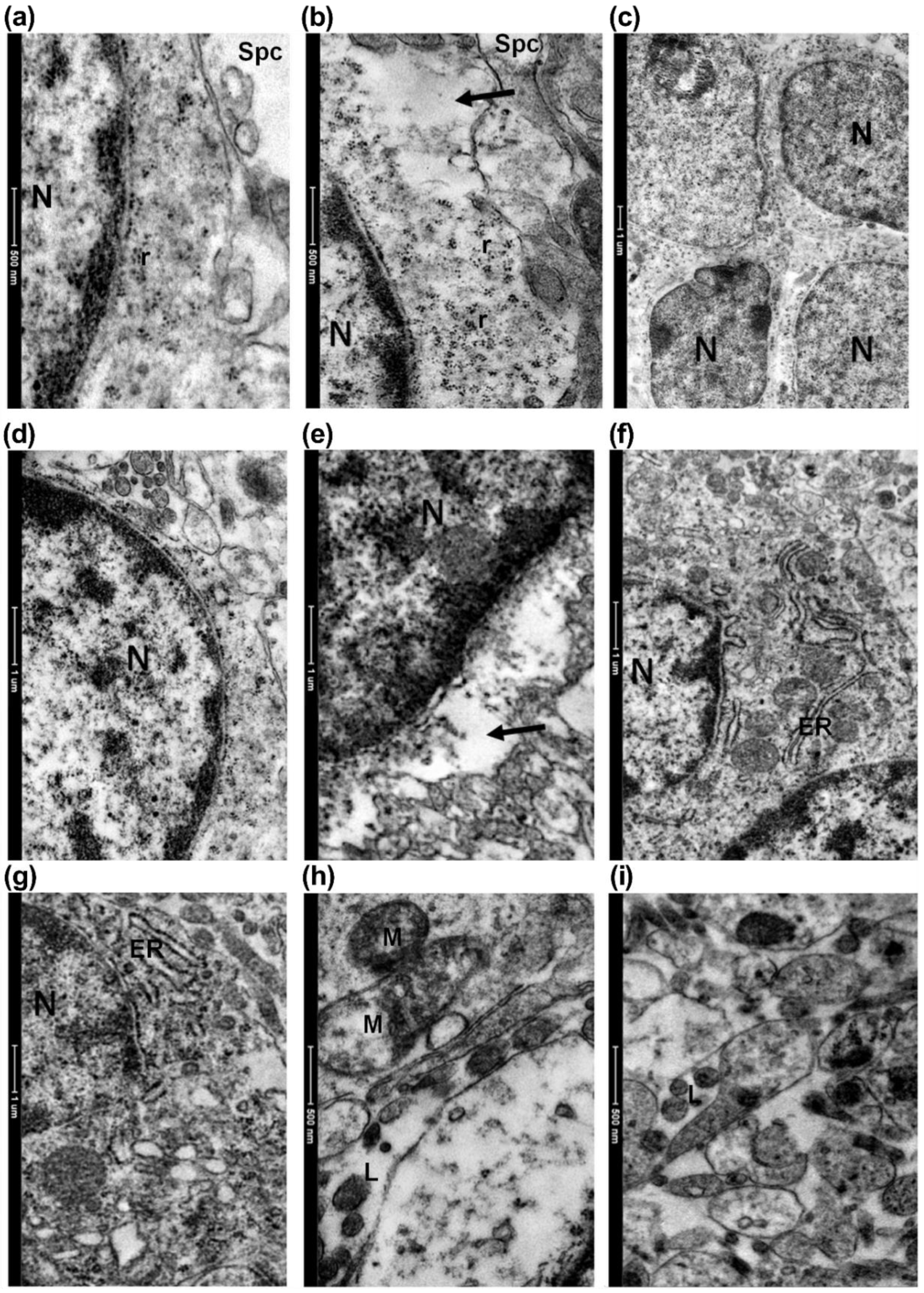
3.2. Electron Microscopy
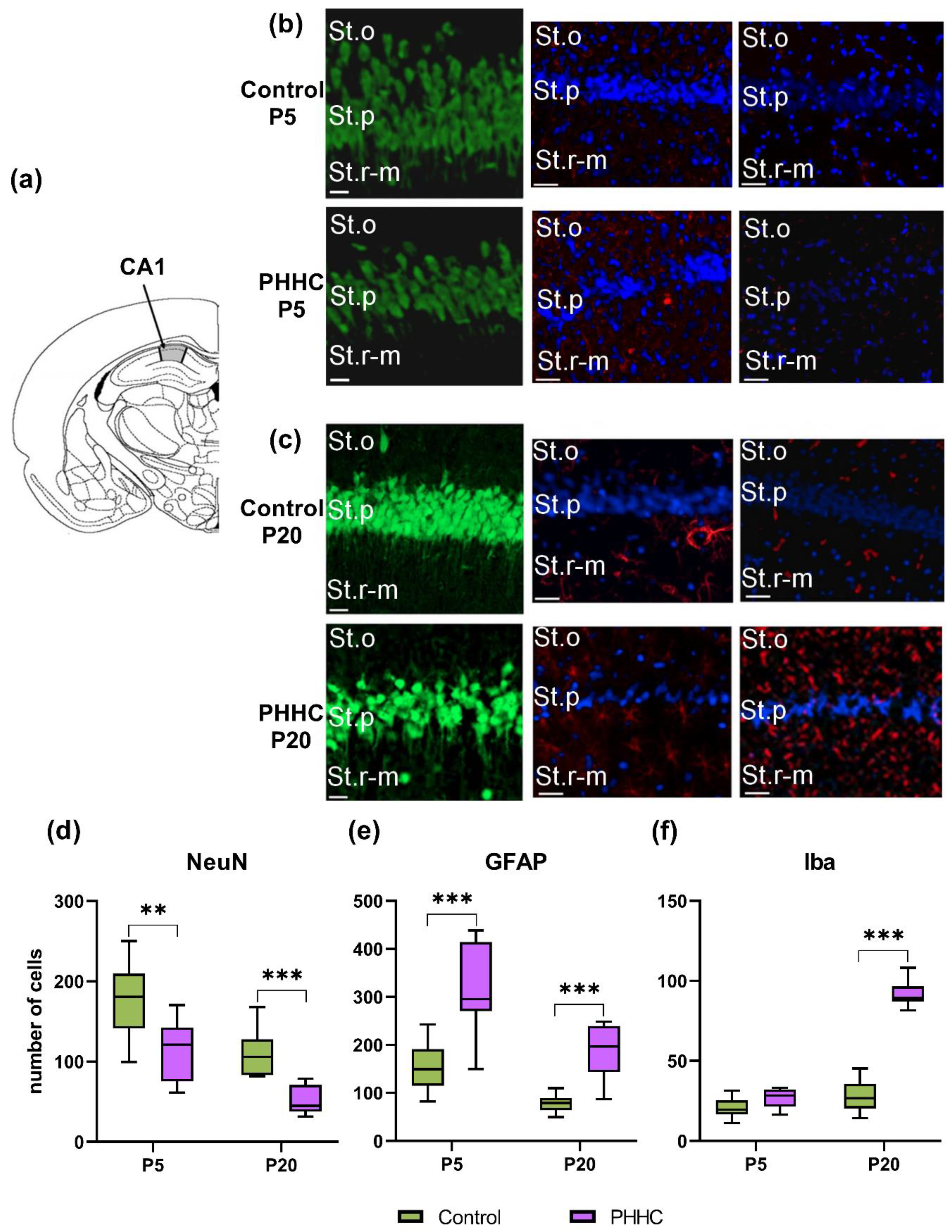
3.3. PHHC-Induced Neuronal Cell Loss and Microglia Activation
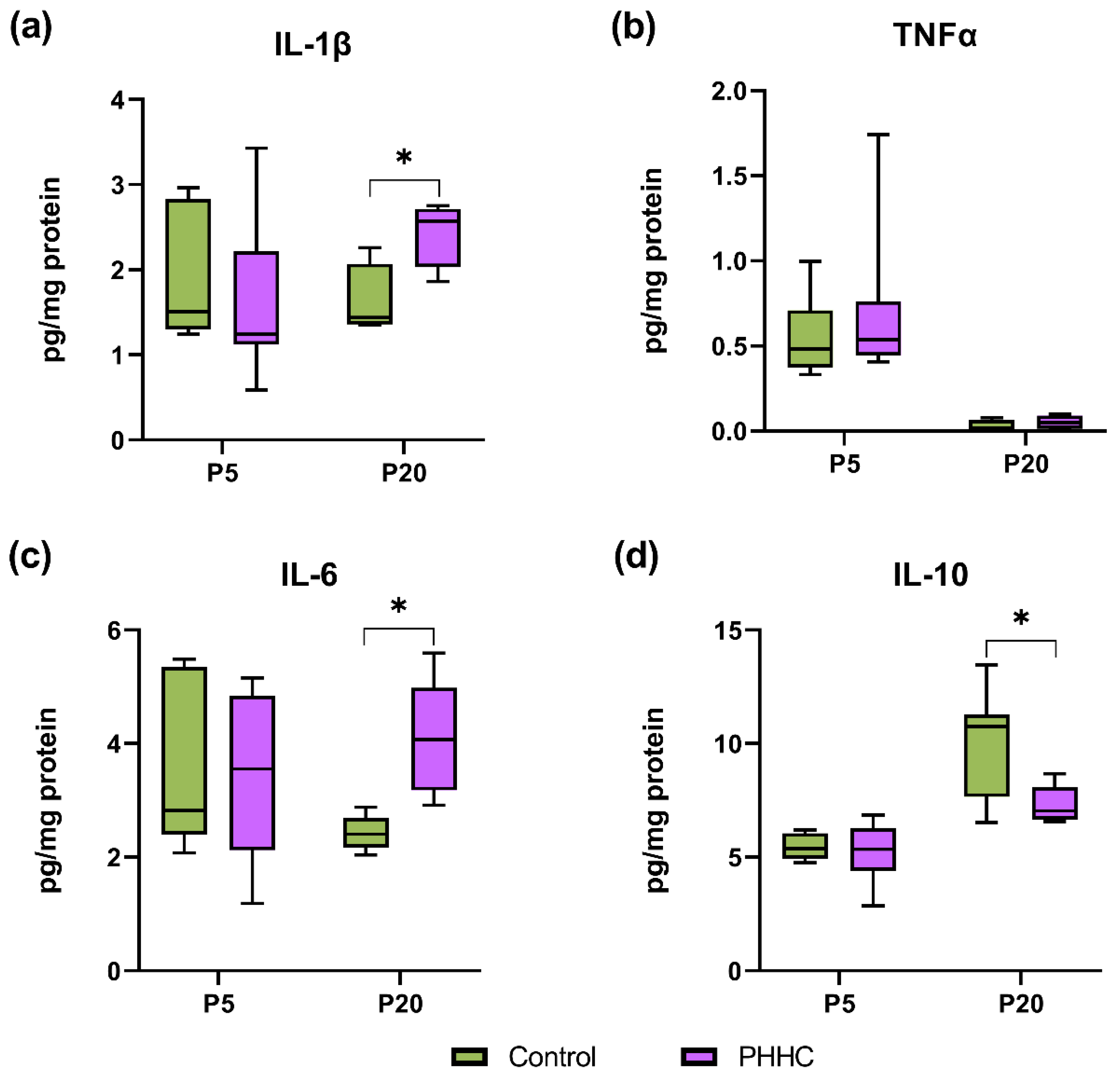
3.4. Pro- and Anti-Inflammatory Cytokine Content
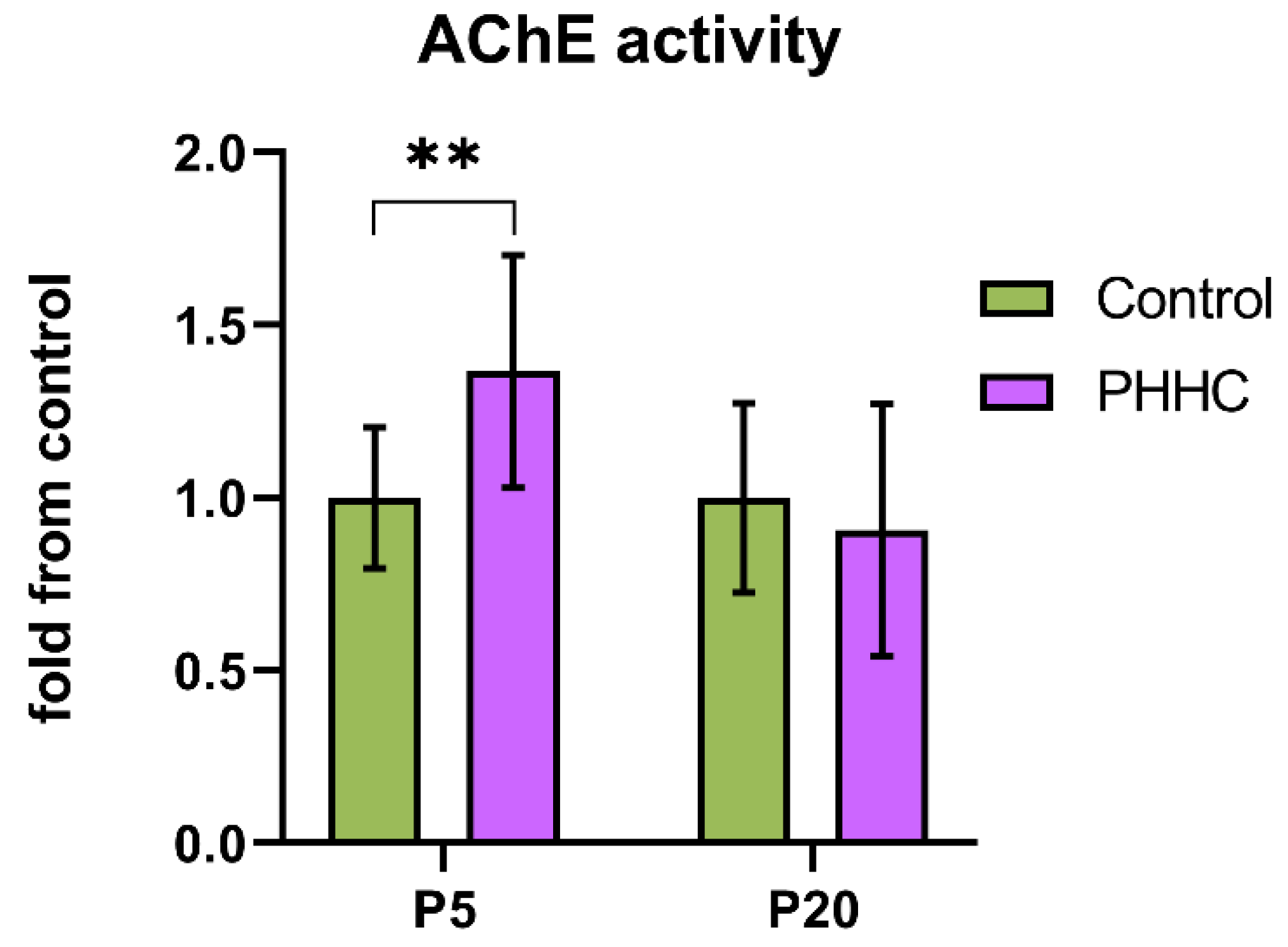
3.5. Acetylcholinesterase Activity
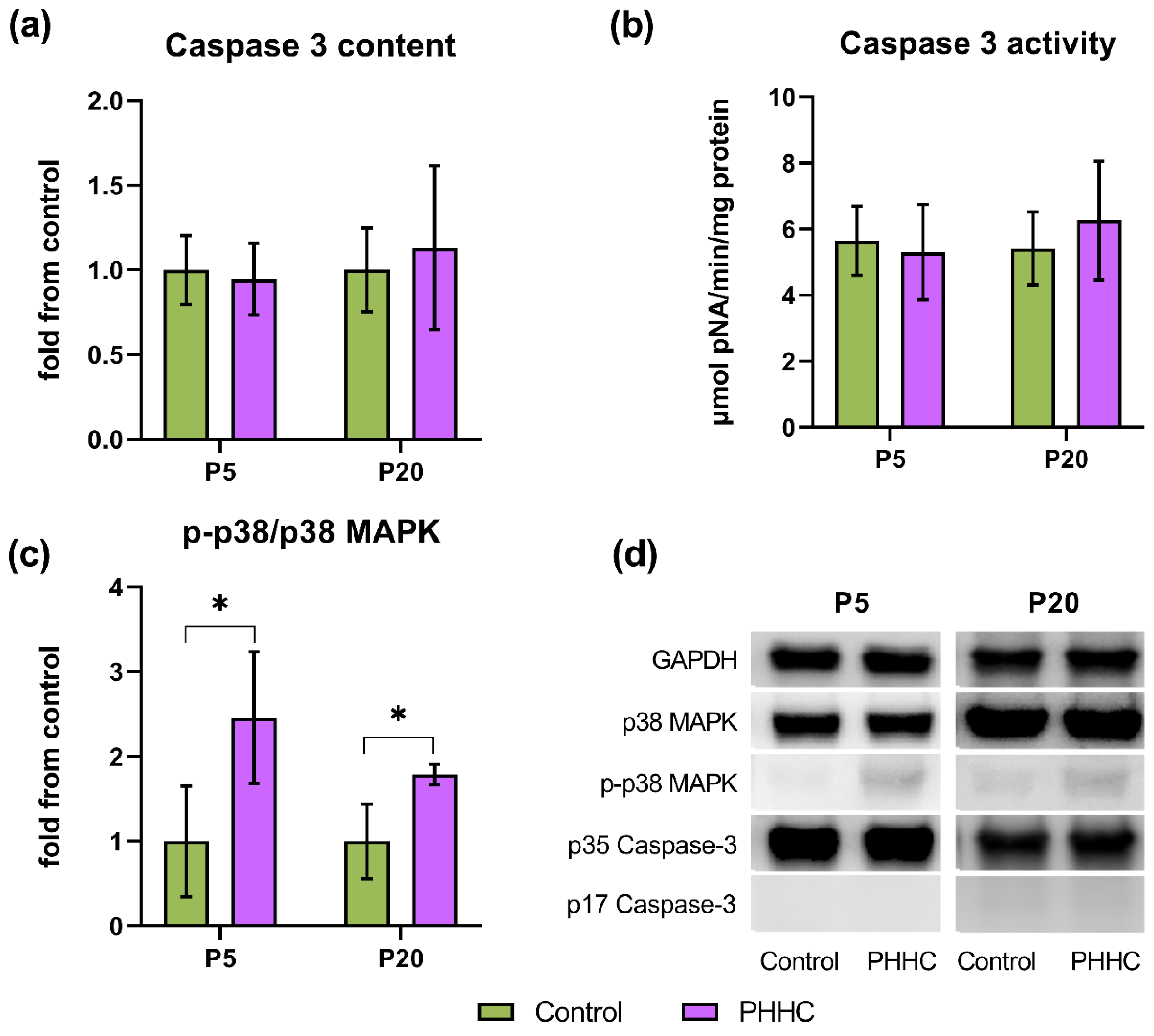
3.6. Caspase-3 Content and Activity
3.7. p38 MAPK Content
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AChE | acetylcholinesterase |
| EPR | endoplasmatic reticulum |
| GAPDH | glyceraldehyde 3-phosphate dehydrogenase |
| GFAP | glial fibrillary acidic protein |
| Hcy | homocysteine |
| HHC | hyperhomocysteinemia |
| Iba1 | ionized calcium-binding adapter molecule |
| IL-1β | interleukin-1β |
| IL-6 | interleukin-6 |
| IL-10 | interleukin-10 |
| MAPK | mitogen-activated protein kinase |
| Mthfr | methylenetetrahydrofolate reductase |
| NeuN | neuronal nuclear marker protein |
| PHHC | prenatal hyperhomocysteinemia |
| SAH | S-adenosyl homocysteine |
| SDS | sodium dodecyl sulfate |
| STAT3 | signal transducer and activator of transcription 3 |
| TNFα | tumor necrosis factor alpha |
References
- Arutjunyan, A.; Kozina, L.; Stvolinskiy, S.; Bulygina, Y.; Mashkina, A.; Khavinson, V. Pinealon protects the rat offspring from prenatal hyperhomocysteinemia. Int. J. Clin. Exp. Med. 2012, 5, 179–185. [Google Scholar]
- Baydas, G.; Koz, S.T.; Tuzcu, M.; Nedzvetsky, V.S. Melatonin prevents gestational hyperhomocysteinemia-associated alterations in neurobehavioral developments in rats. J. Pineal Res. 2008, 44, 181–188. [Google Scholar] [CrossRef]
- Blaise, S.A.; Nedelec, E.; Schroeder, H.; Alberto, J.M.; Bossenmeyer-Pourie, C.; Gueant, J.L.; Daval, J.L. Gestational vita-min B deficiency leads to homocysteine-associated brain apoptosis and alters neurobehavioral development in rats. Am. J. Pathol. 2007, 170, 667–679. [Google Scholar] [CrossRef] [Green Version]
- Jadavji, N.; Deng, L.; Malysheva, O.; Caudill, M.; Rozen, R. MTHFR deficiency or reduced intake of folate or choline in pregnant mice results in impaired short-term memory and increased apoptosis in the hippocampus of wild-type offspring. Neuroscience 2015, 300, 1–9. [Google Scholar] [CrossRef]
- Yakovleva, O.V.; Ziganshina, A.R.; Dmitrieva, S.A.; Arslanova, A.N.; Yakovlev, A.V.; Minibayeva, F.V.; Khaertdinov, N.N.; Ziyatdinova, G.K.; Giniatullin, R.A.; Sitdikova, G.F. Hydrogen sulfide ameliorates developmental impairments of rat offspring with prenatal hyperhomocysteinemia. Oxidative Med. Cell. Longev. 2018, 2018, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Schweinberger, B.M.; Rodrigues, A.F.; dos Santos, T.M.; Rohden, F.; Barbosa, S.; Soster, P.R.D.L.; Partata, W.A.; Faccioni-Heuser, M.C.; Wyse, A.T.S. Methionine administration in pregnant rats causes memory deficit in the offspring and alters ultrastructure in brain tissue. Neurotox. Res. 2017, 33, 239–246. [Google Scholar] [CrossRef] [PubMed]
- Shcherbitskaya, A.D.; Milyutina, Y.P.; Zaloznyaya, I.V.; Arutjunyan, A.V.; Nalivaeva, N.N.; Zhuravin, I.A. The effects of prenatal hyperhomocysteinemia on the formation of memory and the contents of biogenic amines in the rat hippocampus. Neurochem. J. 2017, 11, 296–301. [Google Scholar] [CrossRef]
- Ars, C.L.; Nijs, I.M.; Marroun, H.E.; Muetzel, R.; Schmidt, M.; Graaff, J.S.-D.; Van Der Lugt, A.; Jaddoe, V.W.; Hofman, A.; Steegers, E.A.; et al. Prenatal folate, homocysteine and vitamin B12 levels and child brain volumes, cognitive development and psychological functioning: The Generation R Study. Br. J. Nutr. 2016, 122, S1–S9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pizzolo, F.; Blom, H.J.; Choi, S.W.; Girelli, D.; Guarini, P.; Martinelli, N.; Stanzial, A.M.; Corrocher, R.; Olivieri, O.; Friso, S. Folic acid effects on s-adenosylmethionine, s-adenosylhomocysteine, and DNA methylation in patients with intermediate hyperhomocysteinemia. J. Am. Coll. Nutr. 2011, 30, 11–18. [Google Scholar] [CrossRef] [PubMed]
- Pickell, L.; Brown, K.; Li, D.; Wang, X.-L.; Deng, L.; Wu, Q.; Selhub, J.; Luo, L.; Jerome-Majewska, L.; Rozen, R. High intake of folic acid disrupts embryonic development in mice. Birth Defects Res. Part A Clin. Mol. Teratol. 2010, 91, 8–19. [Google Scholar] [CrossRef]
- Valera-Gran, D.; Navarrete-Muñoz, E.M.; Garcia de la Hera, M.; Fernández-Somoano, A.; Tardón, A.; Ibarluzea, J.; Ball-uerka, N.; Murcia, M.; González-Safont, L.; Romaguera, D.; et al. Effect of maternal high dosages of folic acid sup-plements on neurocognitive development in children at 4–5 years of age: The prospective birth cohort Infancia y Medio Ambi-ente (INMA) study. Am. J. Clin. Nutr. 2017, 106, 878–887. [Google Scholar]
- Cornet, D.; Clement, A.; Clement, P.; Menezo, Y. High doses of folic acid induce a pseudo-methylenetetrahydrofolate syndrome. SAGE Open Med. Case Rep. 2019, 7, 2050313x1985043. [Google Scholar] [CrossRef]
- Navarrete-Muñoz, E.M.; Valera-Gran, D.; Garcia-de-la-Hera, M.; Gonzalez-Palacios, S.; Riaño, I.; Murcia, M.; Lertxundi, A.; Guxens, M.; Tardón, A.; Amiano, P.; et al. High doses of folic acid in the periconceptional pe-riod and risk of low weight for gestational age at birth in a population based cohort study. Eur. J. Nutr. 2017, 58, 241–251. [Google Scholar] [CrossRef]
- Molnár, Z.; Clowry, G. Cerebral cortical development in rodents and primates. Prog. Brain Res. 2012, 195, 45–70. [Google Scholar]
- Zhan, Y.; Paolicelli, R.C.; Sforazzini, F.; Weinhard, L.; Bolasco, G.; Pagani, F.; Vyssotski, A.; Bifone, A.; Gozzi, A.; Ragozzino, D.A.; et al. Deficient neuron-microglia signaling results in impaired functional brain connectivity and social behavior. Nat. Neurosci. 2014, 17, 400–406. [Google Scholar] [CrossRef]
- Rice, D.; Barone, S. Critical periods of vulnerability for the developing nervous system: Evidence from humans and ani-mal models. Environ. Health Perspect. 2000, 108, 511–533. [Google Scholar]
- Arutjunyan, A.V.; Milyutina, Y.P.; Shcherbitskaia, A.; Kerkeshko, G.O.; Zalozniaia, I.V.; Mikhel, A.V. Neurotrophins of the fetal brain and placenta in prenatal hyperhomocysteinemia. Biochemistry 2020, 85, 213–223. [Google Scholar] [CrossRef] [PubMed]
- Shcherbitskaia, A.D.; Vasilev, D.S.; Milyutina, Y.P.; Tumanova, N.L.; Zalozniaia, I.V.; Kerkeshko, G.O.; Arutjunyan, A.V. Maternal hyperhomocysteinemia induces neuroinflammation and neuronal death in the rat offspring cortex. Neurotox. Res. 2020, 38, 408–420. [Google Scholar] [CrossRef] [PubMed]
- Paxinos, G.; Watson, C.R.; Emson, P.C. AChE-stained horizontal sections of the rat brain in stereotaxic coordinates. J. Neurosci. Methods 1980, 3, 129–149. [Google Scholar] [CrossRef]
- Vasilev, D.; Tumanova, N.L.; Kim, K.K.; Lavrentyeva, V.V.; Lukomskaya, N.Y.; Zhuravin, I.A.; Magazanik, L.G.; Zaitsev, A.V. Transient morphological alterations in the hippocampus after pentylenetetrazole-induced seizures in rats. Neurochem. Res. 2018, 43, 1671–1682. [Google Scholar] [CrossRef] [PubMed]
- Bass, J.J.; Wilkinson, D.J.; Rankin, D.; Phillips, B.E.; Szewczyk, N.J.; Smith, K.; Atherton, P.J. An overview of technical considerations for Western blotting applications to physiological research. Scand. J. Med. Sci. Sports 2017, 27, 4–25. [Google Scholar] [CrossRef]
- Ellman, G.L.; Courtney, K.D.; Andres, V.; Featherstone, R.M. A new and rapid colorimetric determination of acetylcho-linesterase activity. Biochem. Pharmacol. 1961, 7, 88–95. [Google Scholar] [CrossRef]
- Nalivaeva, N.N.; Makova, N.Z.; Kochkina, E.G.; John, D.; Arutyunov, V.A.; Kozina, L.S.; Arutjunyan, A.V.; Zhuravin, I.A. Effects of geroprotective peptides on the activity of cholinesterases and formation of the soluble form of the amyloid precur-sor protein in human neuroblastoma SH-SY5Y cells. Neurochem. J. 2011, 5, 176–182. [Google Scholar] [CrossRef]
- Craft, J.M.; Watterson, D.M.; Van Eldik, L.J. Neuroinflammation: A potential therapeutic target. Expert Opin. Ther. Targets 2005, 9, 887–900. [Google Scholar] [CrossRef] [PubMed]
- Moore, A.H.; O’Banion, M. Neuroinflammation and anti-inflammatory therapy for Alzheimer’s disease. Adv. Drug Deliv. Rev. 2002, 54, 1627–1656. [Google Scholar] [CrossRef]
- Walter, L.; Neumann, H. Role of microglia in neuronal degeneration and regeneration. Semin. Immunopathol. 2009, 31, 513–525. [Google Scholar] [CrossRef]
- Wojtera, M.; Sikorska, B.; Sobow, T.; Liberski, P.P. Microglial cells in neurodegenerative disorders. Folia Neuropathol. 2005, 43, 311–321. [Google Scholar]
- Soliman, M.L.; Combs, C.K.; Rosenberger, T.A. Modulation of Inflammatory Cytokines and Mitogen-activated Protein Kinases by Acetate in Primary Astrocytes. J. Neuroimmune Pharmacol. 2013, 8, 287–300. [Google Scholar] [CrossRef] [Green Version]
- Longoni, A.; Bellaver, B.; Bobermin, L.D.; Santos, C.L.; Nonose, Y.; Kolling, J.; De Assis, A.M.; Quincozes-Santos, A.; Wyse, A.T.S. Homocysteine induces glial reactivity in adult rat astrocyte cultures. Mol. Neurobiol. 2017, 55, 1966–1976. [Google Scholar] [CrossRef]
- Da Cunha, A.A.; Ferreira, A.G.K.; Wyse, A.T.S. Increased inflammatory markers in brain and blood of rats subjected to acute homocysteine administration. Metab. Brain Dis. 2010, 25, 199–206. [Google Scholar] [CrossRef]
- Schweinberger, B.M.; Rodrigues, A.F.; Turcatel, E.; Pierozan, P.; Pettenuzzo, L.F.; Grings, M.; Scaini, G.; Parisi, M.M.; Leipnitz, G.; Streck, E.L.; et al. Maternal hypermethioninemia affects neurons number, neurotro-phins levels, energy metabolism, and Na(+),K(+)-ATPase expression/content in brain of rat offspring. Mol. Neurobiol. 2018, 55, 980–988. [Google Scholar] [CrossRef]
- Ma, X.C.; Gottschall, P.E.; Chen, L.T.; Wiranowska, M.; Phelps, C.P. Role and mechanisms of interleukin-1 in the modulation of Neurotoxicity. Neuroimmunomodulation 2002, 10, 199–207. [Google Scholar] [CrossRef]
- Qiu, Z.; Sweeney, D.D.; Netzeband, J.G.; Gruol, D.L. Chronic Interleukin-6 alters NMDA receptor-mediated membrane responses and enhances neurotoxicity in developing CNS Neurons. J. Neurosci. 1998, 18, 10445–10456. [Google Scholar] [CrossRef]
- Makhro, A.V.; Mashkina, A.P.; Solenaya, O.A.; Trunova, O.A.; Kozina, L.S.; Arutyunian, A.V.; Bulygina, E.R. Prenatal hyperhomocysteinemia as a model of oxidative stress of the brain. Bull. Exp. Biol. Med. 2008, 146, 33–35. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, L.; Li, X.; Geng, J. Preliminary analysis of immunoregulatory mechanism of hyperhomocyste-inemiainduced brain injury in Wistar-Kyoto rats. Exp. Ther. Med. 2021, 21, 483. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, L.; Zhou, X.; Geng, J.; Li, X. The immunomodulatory mechanism of brain injury induced by hyperho-mocysteinemia in spontaneously hypertensive rats. J. Cell. Biochem. 2019, 120, 9421–9429. [Google Scholar] [CrossRef]
- dos Santos, T.M.; Júnior, O.V.R.; Alves, V.S.; Coutinho-Silva, R.; Savio, L.E.B.; Wyse, A.T. Hyperhomocysteinemia alters cytokine gene expression, cytochrome c oxidase activity and oxidative stress in striatum and cerebellum of rodents. Life Sci. 2021, 277, 119386. [Google Scholar] [CrossRef]
- Nkpaa, K.W.; Owoeye, O.; Amadi, B.A.; Adedara, I.A.; Abolaji, A.O.; Wegwu, M.O.; Farombi, E.O. Ethanol exacerbates manganeseinduced oxidative/nitrosative stress, proinflammatory cytokines, nuclear factor-κB activation, and apoptosis induction in rat cerebellar cortex. J. Biochem. Mol. Toxicol. 2020, 35, e22681. [Google Scholar] [PubMed]
- Rahim, N.S.; Lim, S.M.; Mani, V.; Hazalin, N.A.M.N.; Majeed, A.B.A.; Ramasamy, K. Virgin coconut oil-induced neuroprotection in lipopolysaccharide-challenged rats is mediated, in part, through cholinergic, anti-oxidative and anti-inflammatory pathways. J. Diet. Suppl. 2020, 14, 1–27. [Google Scholar] [CrossRef]
- Scherer, E.B.S.; Loureiro, S.O.; Vuaden, F.C.; Da Cunha, A.A.; Schmitz, F.; Kolling, J.; Savio, L.E.B.; Bogo, M.R.; Bonan, C.D.; Netto, C.A.; et al. Mild hyperhomocysteinemia increases brain acetylcholinesterase and proinflammatory cytokine levels in different tissues. Mol. Neurobiol. 2014, 50, 589–596. [Google Scholar] [CrossRef] [PubMed]
- Moreira, D.D.S.; Figueiró, P.W.; Siebert, C.; Prezzi, C.A.; Rohden, F.; Guma, F.C.R.; Manfredini, V.; Wyse, A.T.S. Chronic mild hyperhomocysteinemia alters inflammatory and oxidative/nitrative status and causes protein/dna damage, as well as ultrastructural changes in cerebral cortex: Is acetylsalicylic acid neuroprotective? Neurotox. Res. 2018, 33, 580–592. [Google Scholar] [CrossRef]
- Borovikova, L.V.; Ivanova, S.; Zhang, M.; Yang, H.; Botchkina, G.I.; Watkins, L.R.; Wang, H.; Abumrad, N.; Eaton, J.W.; Tracey, K.J. Vagus nerve stimulation attenuates the systemic inflammatory response to endotoxin. Nat. Cell Biol. 2000, 405, 458–462. [Google Scholar] [CrossRef]
- Wang, J.; Zhang, H.Y.; Tang, X.C. Huperzine a improves chronic inflammation and cognitive decline in rats with cerebral hypoperfusion. J. Neurosci. Res. 2009, 88, 807–815. [Google Scholar] [CrossRef]
- Wessler, I.; Reinheimer, T.; Klapproth, H.; Schneider, F.-J.; Racké, K.; Hammer, R. Mammalian glial cells in culture synthesize acetylcholine. Naunyn-Schmiedeberg’s Arch. Pharmacology 1997, 356, 694–697. [Google Scholar]
- Shytle, R.D.; Mori, T.; Townsend, K.P.; Vendrame, M.; Sun, N.; Zeng, J.; Ehrhart, J.; Silver, A.A.; Sanberg, P.R.; Tan, J. Cholinergic modulation of microglial activation by α7 nicotinic receptors. J. Neurochem. 2004, 89, 337–343. [Google Scholar] [CrossRef]
- Jha, S.K.; Jha, N.K.; Kar, R.; Ambasta, R.K.; Kumar, P. p38 MAPK and PI3K/AKT Signalling Cascades in Parkinson’s Disease. Int. J. Mol. Cell. Med. 2015, 4, 67–86. [Google Scholar]
- Lee, J.K.; Kim, N.J. Recent advances in the inhibition of p38 MAPK as a potential strategy for the treatment of Alzheimer’s disease. Molecules 2017, 22, 1287. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.; Dong, Z.; Cheng, M.; Zhao, Y.; Wang, M.; Sai, N.; Wang, X.; Liu, H.; Huang, G.; Zhang, X. Homocysteine exaggerates microglia activation and neuroinflammation through microglia localized STAT3 overactivation following ischemic stroke. J. Neuroinflamm. 2017, 14, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gee, M.S.; Kim, S.-W.; Kim, N.; Lee, S.J.; Oh, M.S.; Jin, H.K.; Bae, J.-S.; Inn, K.-S.; Kim, N.-J.; Kil Lee, J. A novel and selective p38 mitogen-activated protein kinase inhibitor attenuates LPS-induced neuroinflammation in BV2 microglia and a mouse model. Neurochem. Res. 2018, 43, 2362–2371. [Google Scholar] [CrossRef]
- Millot, P.; San, C.; Bennana, E.; Porte, B.; Vignal, N.; Hugon, J.; Paquet, C.; Hosten, B.; Mouton-Liger, F. STAT3 inhibition protects against neuroinflammation and BACE1 upregulation induced by systemic inflammation. Immunol. Lett. 2020, 228, 129–134. [Google Scholar] [CrossRef]
- Lang, R.; Patel, D.; Morris, J.J.; Rutschman, R.L.; Murray, P.J. Shaping gene expression in activated and resting primary macrophages by IL-10. J. Immunol. 2002, 169, 2253–2263. [Google Scholar] [CrossRef] [Green Version]
- Deng, Y.; Lu, J.; Sivakumar, V.; Ling, E.A.; Kaur, C. Amoeboid microglia in the periventricular white matter induce oli-godendrocyte damage through expression of proinflammatory cytokines via MAP kinase signaling pathway in hypoxic neonatal rats. Brain Pathol. 2008, 18, 387–400. [Google Scholar] [CrossRef]
- Shi, S.; Cui, Q.; Xu, J.; Tang, Z.; Shi, B.; Liu, Z. Baicalin suppresses bilirubin-induced apoptosis and inflammation by regulating p38 mitogen-activated protein kinases (MAPK) signaling in neonatal neurons. Med Sci. Monit. 2020, 26, 26. [Google Scholar] [CrossRef]
- Irving, E.A.; Bamford, M. Role of mitogen- and stress-activated kinases in ischemic injury. Br. J. Pharmacol. 2002, 22, 631–647. [Google Scholar] [CrossRef] [Green Version]
- Hommes, D.W.; Peppelenbosch, M.; Van Deventer, S.J.H. Mitogen activated protein (MAP) kinase signal transduction pathways and novel anti-inflammatory targets. Gut 2003, 52, 144–151. [Google Scholar] [CrossRef] [Green Version]
- Zhou, L.; Wang, D.; Qiu, X.; Zhang, W.; Gong, Z.; Wang, Y.; Xu, X. DHZCP Modulates microglial M1/M2 polarization via the p38 and TLR4/NF-κB signaling pathways in LPS-stimulated microglial cells. Front. Pharmacol. 2020, 11, 1126. [Google Scholar] [CrossRef] [PubMed]
- Bu, X.; Huang, P.; Qi, Z.; Zhang, N.; Han, S.; Fang, L.; Li, J. Cell type-specific activation of p38 MAPK in the brain regions of hypoxic preconditioned mice. Neurochem. Int. 2007, 51, 459–466. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.; Ge, M.; Li, C.; Yu, L.; Gu, Y.; Hu, Y.; Cao, Z. Effects of p38 MAPK signaling pathway on cognitive function and recovery of neuronal function after hypoxic-ischemic brain injury in newborn rats. J. Clin. Neurosci. 2020, 78, 365–370. [Google Scholar] [CrossRef] [PubMed]
- van der Bruggen, T.; Nijenhuis, S.; van Raaij, E.; Verhoef, J.; van Asbeck, B.S. Lipopolysaccharide-induced tumor necrosis factor alpha production by human monocytes involves the raf-1/MEK1-MEK2/ERK1-ERK2 pathway. Infect. Immun. 1999, 67, 3824–3829. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bachstetter, A.D.; Van Eldik, L.J. The p38 MAP kinase family as regulators of proinflammatory cytokine production in degenerative diseases of the CNS. Aging Dis. 2010, 1, 199–211. [Google Scholar]
- Duan, W.; Ladenheim, B.; Cutler, R.G.; Kruman, I.I.; Cadet, J.L.; Mattson, M.P. Dietary folate deficiency and elevated ho-mocysteine levels endanger dopaminergic neurons in models of Parkinson’s disease. J. Neurochem. 2002, 80, 101–110. [Google Scholar] [CrossRef]
- Maler, J.; Seifert, W.; Hüther, G.; Wiltfang, J.; Rüther, E.; Kornhuber, J.; Bleich, S. Homocysteine induces cell death of rat astrocytes in vitro. Neurosci. Lett. 2003, 347, 85–88. [Google Scholar] [CrossRef]
- Suhara, T.; Fukuo, K.; Yasuda, O.; Tsubakimoto, M.; Takemura, Y.; Kawamoto, H.; Yokoi, T.; Mogi, M.; Kaimoto, T.; Ogihara, T. Homocysteine enhances endothelial apoptosis via upregulation of fas-mediated pathways. Hypertension 2004, 43, 1208–1213. [Google Scholar] [CrossRef]
- Koz, S.T.; Gouwy, N.T.; Demir, N.; Nedzvetsky, V.S.; Etem, E.; Baydas, G. Effects of maternal hyperhomocysteinemia induced by methionine intake on oxidative stress and apoptosis in pup rat brain. Int. J. Dev. Neurosci. 2010, 28, 325–329. [Google Scholar] [CrossRef] [PubMed]
- Endo, N.; Nishiyama, K.; Okabe, M.; Matsumoto, M.; Kanouchi, H.; Oka, T. Vitamin B6 suppresses apoptosis of NM-1 bovine endothelial cells induced by homocysteine and copper. Biochim. Biophys. Acta (BBA) Gen. Subj. 2007, 1770, 571–577. [Google Scholar] [CrossRef] [PubMed]
- Hirashima, Y.; Seshimo, S.; Fujiki, Y.; Okabe, M.; Nishiyama, K.; Matsumoto, M.; Kanouchi, H.; Oka, T. Homocysteine and copper induce cellular apoptosis via caspase activation and nuclear translocation of apoptosis-inducing factor in neuronal cell line SH-SY5Y. Neurosci. Res. 2010, 67, 300–306. [Google Scholar] [CrossRef] [PubMed]
- Gitto, E.; Pellegrino, S.; Gitto, P.; Barberi, I.; Reiter, R.J. Oxidative stress of the newborn in the pre- and postnatal period and the clinical utility of melatonin. J. Pineal Res. 2009, 46, 128–139. [Google Scholar] [CrossRef] [PubMed]
- Jakubowski, H. Molecular basis of homocysteine toxicity in humans. Cell. Mol. Life Sci. 2004, 61, 470–487. [Google Scholar] [CrossRef] [PubMed]
- Boldyrev, A.A. Molecular mechanisms of homocysteine toxicity. Biochemistry 2009, 74, 589–598. [Google Scholar] [CrossRef] [PubMed]
- Poddar, R.; Paul, S. Novel crosstalk between ERK MAPK and p38 MAPK leads to homocysteine-NMDA receptor-mediated neuronal cell death. J. Neurochem. 2013, 124, 558–570. [Google Scholar] [CrossRef] [Green Version]
- McGee, M.; Bainbridge, S.; Fontaine-Bisson, B. A crucial role for maternal dietary methyl donor intake in epigenetic pro-gramming and fetal growth outcomes. Nutr. Rev. 2018, 76, 469–478. [Google Scholar] [CrossRef]
- James, S.J.; Melnyk, S.; Pogribna, M.; Pogribny, I.P.; Caudill, M.A. Elevation in S-adenosylhomocysteine and DNA hypo-methylation: Potential epigenetic mechanism for homocysteine-related pathology. J. Nutr. 2002, 132, 2361S–2366S. [Google Scholar] [CrossRef]
- Heil, S.G.; Herzog, E.M.; Griffioen, P.H.; Van Zelst, B.; Willemsen, S.P.; De Rijke, Y.B.; Steegers-Theunissen, R.P.M.; Steegers, E.A.P. Lower S-adenosylmethionine levels and DNA hypomethylation of placental growth factor (PlGF) in placental tissue of early-onset preeclampsia-complicated pregnancies. PLoS ONE 2019, 14, e0226969. [Google Scholar] [CrossRef] [Green Version]
- Lin, N.; Qin, S.; Luo, S.; Cui, S.; Huang, G.; Zhang, X. Homocysteine induces cytotoxicity and proliferation inhibition in neural stem cells via DNA methylation in vitro. FEBS J. 2014, 281, 2088–2096. [Google Scholar] [CrossRef]
- Perla-Kajan, J.; Jakubowski, H. Dysregulation of epigenetic mechanisms of gene expression in the pathologies of hyper-homocysteinemia. Int. J. Mol. Sci. 2019, 20, 3140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scott, J.M.; Molloy, A.; Kennedy, D.G.; Kennedy, S.; Weir, D.G. Effects of the disruption of transmethylation in the central nervous system: An animal model. Acta Neurol. Scand. 1994, 89, 27–31. [Google Scholar] [CrossRef] [PubMed]
- Schatz, R.A.; Wilens, T.E.; Sellinger, O.Z. Decreased transmethylation of biogenic amines after in vivo elevation of brain s-adenosyl-l-homocysteine. J. Neurochem. 1981, 36, 1739–1748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dragovich, T.; Rudin, C.M.; Thompson, C.B. Signal transduction pathways that regulate cell survival and cell death. Oncogene 1998, 17, 3207–3213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Figueiró, P.W.; Moreira, D.D.S.; dos Santos, T.M.; Prezzi, C.A.; Rohden, F.; Faccioni-Heuser, M.C.; Manfredini, V.; Netto, C.A.; Wyse, A.T. The neuroprotective role of melatonin in a gestational hypermethioninemia model. Int. J. Dev. Neurosci. 2019, 78, 198–209. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Li, W.; Li, S.; Yan, J.; Wilson, J.X.; Huang, G. Maternal folic acid supplementation during pregnancy improves neurobehavioral development in rat offspring. Mol. Neurobiol. 2017, 55, 2676–2684. [Google Scholar] [CrossRef]
- Chen, S.; Dong, Z.; Zhao, Y.; Sai, N.; Wang, X.; Liu, H.; Huang, G.; Zhang, X. Homocysteine induces mitochondrial dysfunction involving the crosstalk between oxidative stress and mitochondrial pSTAT3 in rat ischemic brain. Sci. Rep. 2017, 7, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Salissou, M.T.M.; Mahaman, Y.A.R.; Zhu, F.; Huang, F.; Wang, Y.; Xu, Z.; Ke, D.; Wang, Q.; Liu, R.; Wang, J.Z.; et al. Methanolic extract of Tamarix Gallica attenuates hyperhomocysteinemia induced AD-like pathology and cognitive im-pairments in rats. Aging 2018, 10, 3229–3248. [Google Scholar] [CrossRef] [PubMed]
- Kovalska, M.; Hnilicova, P.; Kalenska, D.; Tothova, B.; Adamkov, M.; Lehotsky, J. Effect of methionine diet on metabolic and histopathological changes of rat hippocampus. Int. J. Mol. Sci. 2019, 20, 6234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riancho, J.; Soto, M.R.; Villagrã¡, N.T.; Berciano, J.; Berciano, M.T.; Lafarga, M. Compensatory motor neuron response to chromatolysis in the murine hsod1g93a model of amyotrophic lateral sclerosis. Front. Cell. Neurosci. 2014, 8, 346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palanca, A.; Casafont, I.; Berciano, M.T.; Lafarga, M. Proteasome inhibition induces DNA damage and reorganizes nuclear architecture and protein synthesis machinery in sensory ganglion neurons. Cell. Mol. Life Sci. 2014, 71, 1961–1975. [Google Scholar] [CrossRef] [PubMed]
- Castejón, O.J. Electron Microscopy Study of Nerve Cell Death Types in Some Central Nervous System Diseases. A Review. Am. J. Biomed. Sci. Res. 2019, 3, 73–83. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shcherbitskaia, A.D.; Vasilev, D.S.; Milyutina, Y.P.; Tumanova, N.L.; Mikhel, A.V.; Zalozniaia, I.V.; Arutjunyan, A.V. Prenatal Hyperhomocysteinemia Induces Glial Activation and Alters Neuroinflammatory Marker Expression in Infant Rat Hippocampus. Cells 2021, 10, 1536. https://doi.org/10.3390/cells10061536
Shcherbitskaia AD, Vasilev DS, Milyutina YP, Tumanova NL, Mikhel AV, Zalozniaia IV, Arutjunyan AV. Prenatal Hyperhomocysteinemia Induces Glial Activation and Alters Neuroinflammatory Marker Expression in Infant Rat Hippocampus. Cells. 2021; 10(6):1536. https://doi.org/10.3390/cells10061536
Chicago/Turabian StyleShcherbitskaia, Anastasiia D., Dmitrii S. Vasilev, Yulia P. Milyutina, Natalia L. Tumanova, Anastasiia V. Mikhel, Irina V. Zalozniaia, and Alexander V. Arutjunyan. 2021. "Prenatal Hyperhomocysteinemia Induces Glial Activation and Alters Neuroinflammatory Marker Expression in Infant Rat Hippocampus" Cells 10, no. 6: 1536. https://doi.org/10.3390/cells10061536