
The Glycoprotein M6a Is Associated with Invasiveness and Radioresistance of Glioblastoma Stem Cells

 ,
,  ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. GB Patient-Derived Cells
2.2. Three-Dimensional Invasion Assays
2.3. Western Blot Analysis
2.4. Immunofluorescence Staining and Microscopy
2.5. Transfection, RNA Extraction, Reverse Transcription and Real-Time PCR
2.6. Genes Correlations
2.7. 3D Spheroid Formation
2.8. 3D Survival Assay under Radiation
3. Results
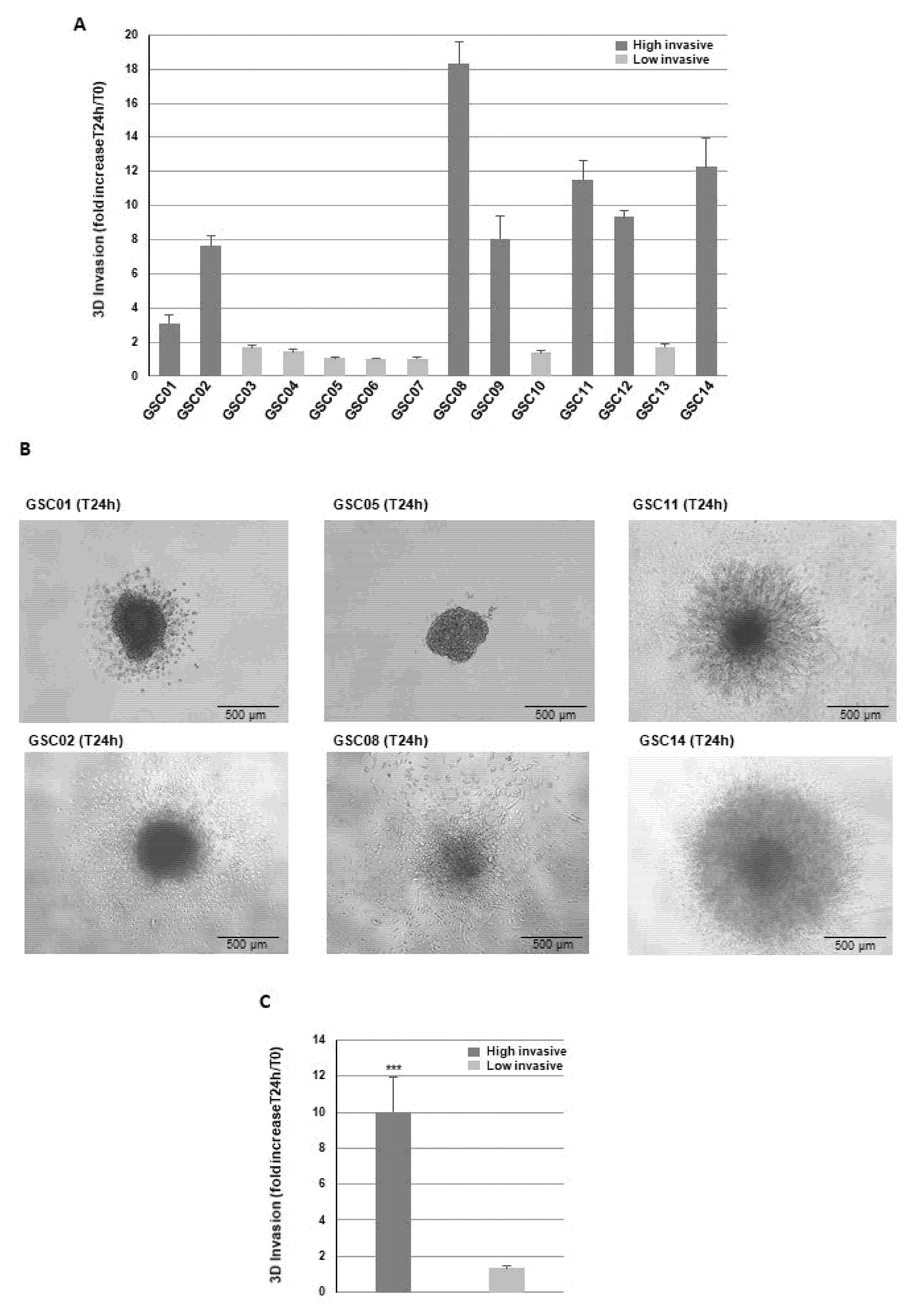
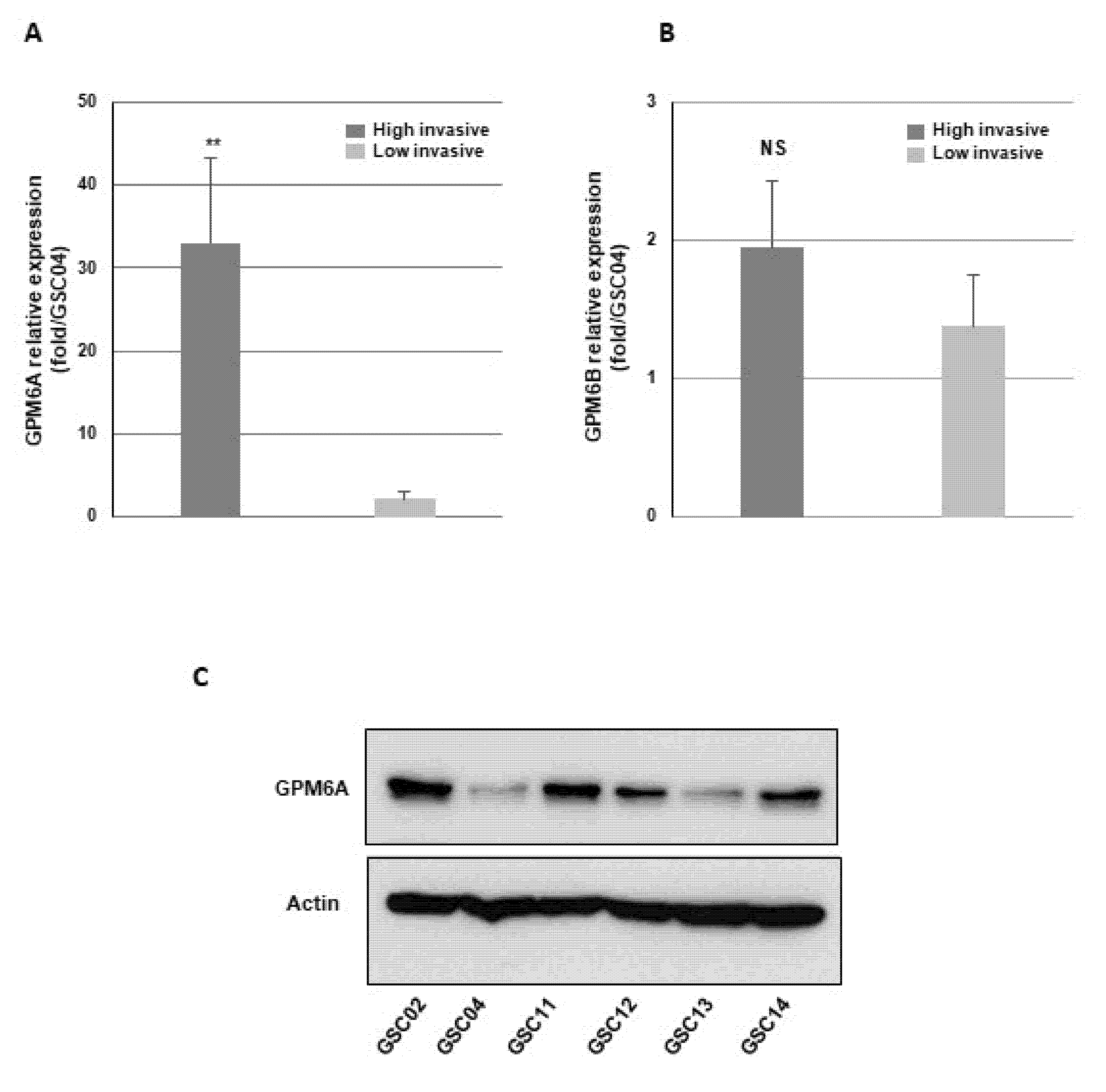
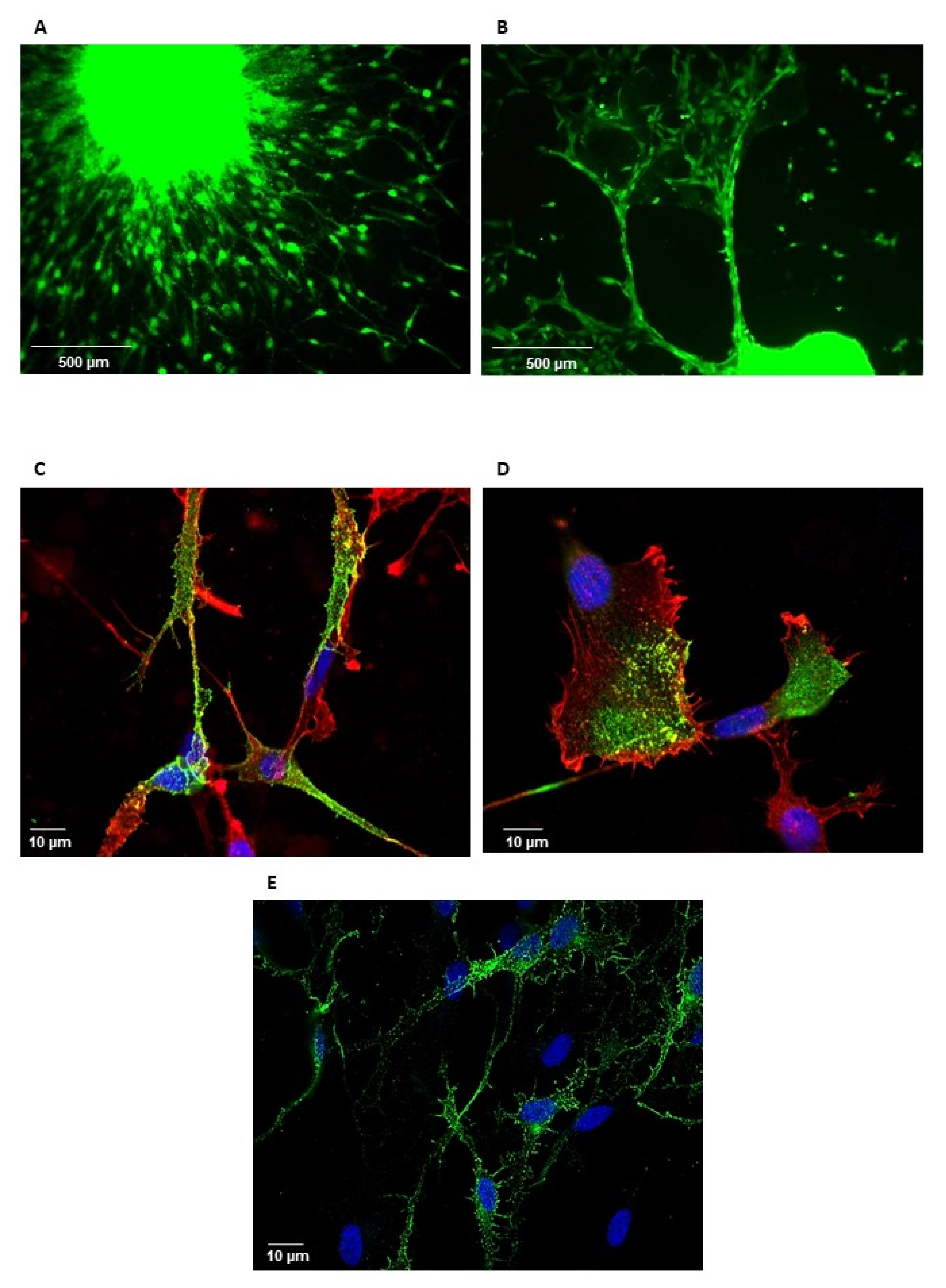
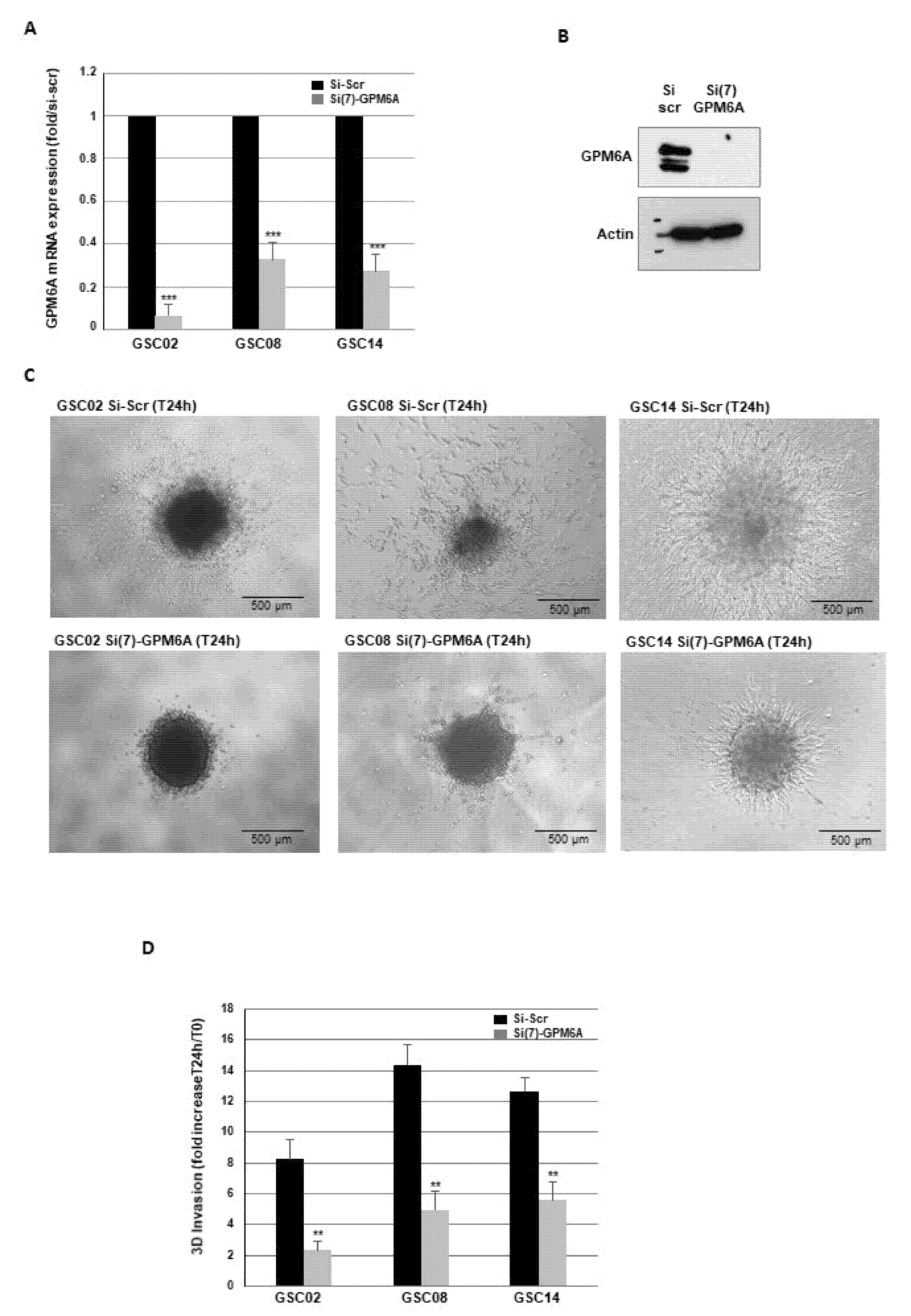
3.1. Blocking GPM6A Expression Represses Invasion of GBSC
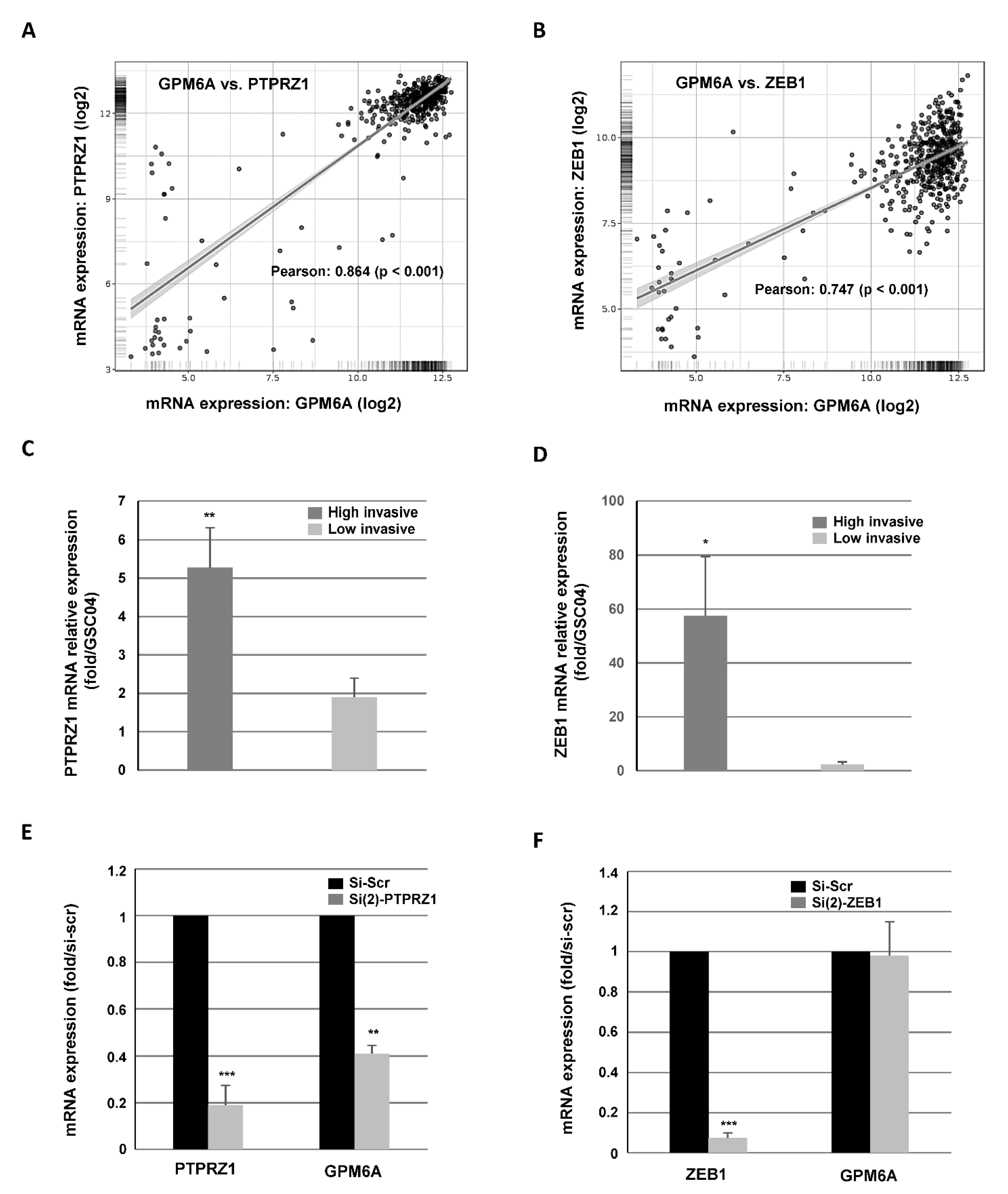
3.2. Targeting PTPRZ1 Inhibits GPM6A Expression and GBSC Invasion
3.3. Down-Regulation of GPM6A or PTPRZ1 Gene Expression Decreases Sphere-Forming Ability of GBSC
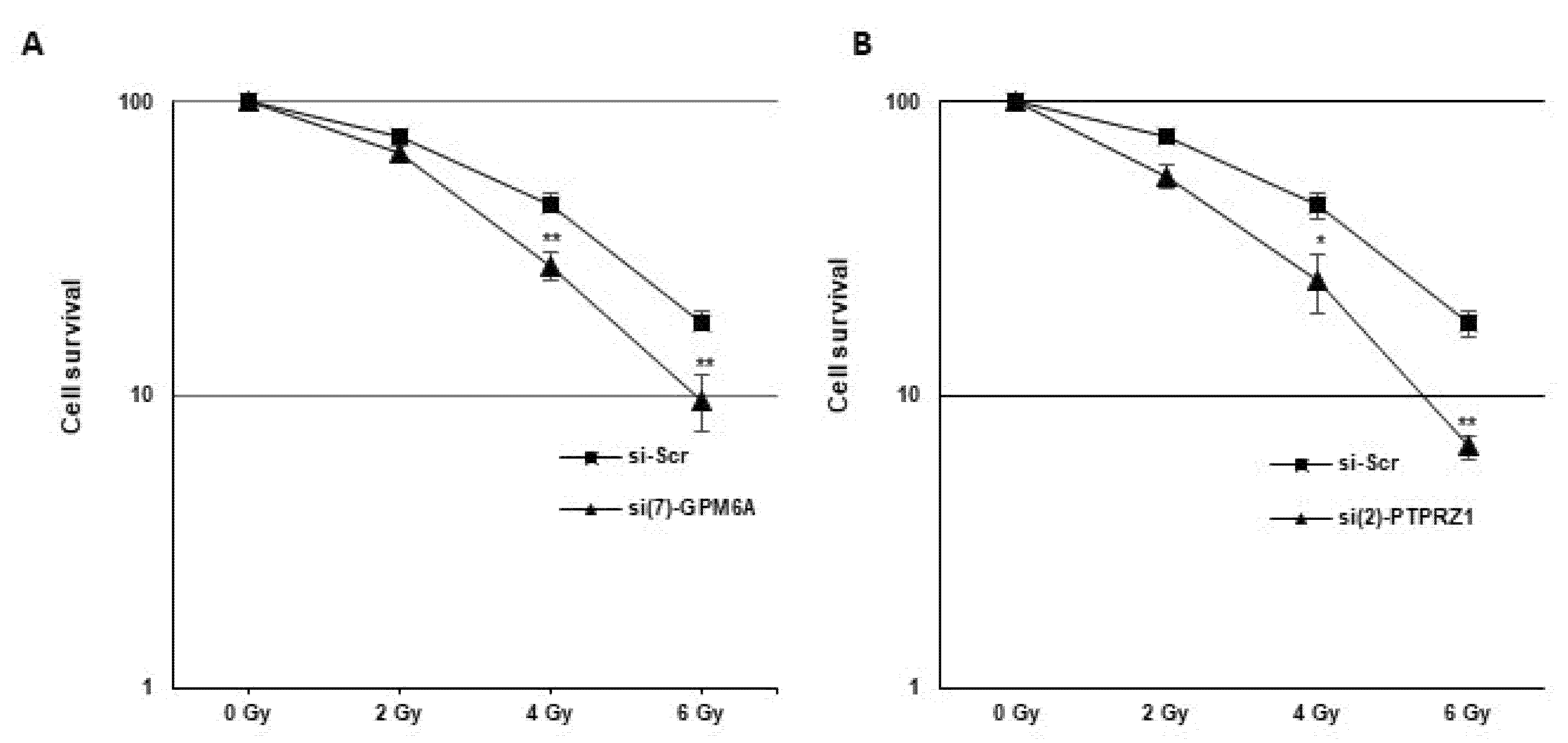
3.4. Blocking GPM6A or PTPRZ1 Radiosensitizes GBSC
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Weller, M.; Butowski, N.; Tran, D.D.; Recht, L.D.; Lim, M.; Hirte, H.; Ashby, L.; Mechtler, L.; Goldlust, S.A.; Iwamoto, F.; et al. Rindopepimut with temozolomide for patients with newly diagnosed, EGFRvIII-expressing glioblastoma (ACT IV): A randomised, double-blind, international phase 3 trial. Lancet Oncol. 2017, 18, 1373–1385. [Google Scholar] [CrossRef] [Green Version]
- Khalifa, J.; Tensaouti, F.; Lusque, A.; Plas, B.; Lotterie, J.A.; Benouaich-Amiel, A.; Uro-Coste, E.; Lubrano, V.; Cohen-Jonathan Moyal, E. Subventricular zones: New key targets for glioblastoma treatment. Radiat. Oncol. 2017, 12, 67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Armocida, D.; Pesce, A.; Palmieri, M.; D’Andrea, G.; Salvati, M.; Santoro, A.; Frati, A. Periventricular zone involvement as a predictor of survival in glioblastoma patients: A single centre cohort-comparison investigation concerning a distinct clinical entity. Interdiscip. Neurosurg. 2021, 25, 101185. [Google Scholar] [CrossRef]
- Armocida, D.; Pesce, A.; Di Giammarco, F.; Frati, A.; Salvati, M.; Santoro, A. Histological, molecular, clinical and outcomes characteristics of multiple lesion glioblastoma. A retrospective monocentric study and review of literature. Neurocirugia 2021, 32, 114–123. [Google Scholar] [CrossRef] [PubMed]
- Bao, S.; Wu, Q.; McLendon, R.E.; Hao, Y.; Shi, Q.; Hjelmeland, A.B.; Dewhirst, M.W.; Bigner, D.D.; Rich, J.N. Glioma stem cells promote radioresistance by preferential activation of the DNA damage response. Nature 2006, 444, 756–760. [Google Scholar] [CrossRef]
- Ortensi, B.; Setti, M.; Osti, D.; Pelicci, G. Cancer stem cell contribution to glioblastoma invasiveness. Stem. Cell Res. Ther. 2013, 4, 18. [Google Scholar] [CrossRef] [Green Version]
- Ito, Y.; Honda, A.; Igarashi, M. Glycoprotein M6a as a signaling transducer in neuronal lipid rafts. Neurosci. Res. 2018, 128, 19–24. [Google Scholar] [CrossRef]
- Alvarez Julia, A.; Frasch, A.C.; Fuchsova, B. Neuronal filopodium formation induced by the membrane glycoprotein M6a (Gpm6a) is facilitated by coronin-1a, Rac1, and p21-activated kinase 1 (Pak1). J. Neurochem. 2016, 137, 46–61. [Google Scholar] [CrossRef] [Green Version]
- Scorticati, C.; Formoso, K.; Frasch, A.C. Neuronal glycoprotein M6a induces filopodia formation via association with cholesterol-rich lipid rafts. J. Neurochem. 2011, 119, 521–531. [Google Scholar] [CrossRef]
- Alfonso, J.; Fernandez, M.E.; Cooper, B.; Flugge, G.; Frasch, A.C. The stress-regulated protein M6a is a key modulator for neurite outgrowth and filopodium/spine formation. Proc. Natl. Acad. Sci. USA 2005, 102, 17196–17201. [Google Scholar] [CrossRef] [Green Version]
- Michibata, H.; Okuno, T.; Konishi, N.; Wakimoto, K.; Kyono, K.; Aoki, K.; Kondo, Y.; Takata, K.; Kitamura, Y.; Taniguchi, T. Inhibition of mouse GPM6A expression leads to decreased differentiation of neurons derived from mouse embryonic stem cells. Stem Cells Dev. 2008, 17, 641–651. [Google Scholar] [CrossRef] [PubMed]
- Charfi, C.; Edouard, E.; Rassart, E. Identification of GPM6A and GPM6B as potential new human lymphoid leukemia-associated oncogenes. Cell Oncol. 2014, 37, 179–191. [Google Scholar] [CrossRef] [PubMed]
- Falch, C.M.; Sundaram, A.Y.M.; Oystese, K.A.; Normann, K.R.; Lekva, T.; Silamikelis, I.; Eieland, A.K.; Andersen, M.; Bollerslev, J.; Olarescu, N.C. Gene expression profiling of fast- and slow-growing non-functioning gonadotroph pituitary adenomas. Eur. J. Endocrinol. 2018, 178, 295–307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lv, J.; Wang, J.; Shang, X.; Liu, F.; Guo, S. Survival prediction in patients with colon adenocarcinoma via multi-omics data integration using a deep learning algorithm. Biosci. Rep. 2020, 40, BSR20201482. [Google Scholar] [CrossRef]
- Ye, Z.; Li, Y.; Xie, J.; Feng, Z.; Yang, X.; Wu, Y.; Pu, Y.; Gao, J.; Xu, X.; Zhu, Z.; et al. Integrated bioinformatics identifies the dysregulation induced by aberrant gene methylation in colorectal carcinoma. Genes Dis. 2021, 8, 521–530. [Google Scholar] [CrossRef]
- Castells, X.; Acebes, J.J.; Boluda, S.; Moreno-Torres, A.; Pujol, J.; Julia-Sape, M.; Candiota, A.P.; Arino, J.; Barcelo, A.; Arus, C. Development of a predictor for human brain tumors based on gene expression values obtained from two types of microarray technologies. OMICS 2010, 14, 157–164. [Google Scholar] [CrossRef] [Green Version]
- Avril, T.; Vauleon, E.; Hamlat, A.; Saikali, S.; Etcheverry, A.; Delmas, C.; Diabira, S.; Mosser, J.; Quillien, V. Human glioblastoma stem-like cells are more sensitive to allogeneic NK and T cell-mediated killing compared with serum-cultured glioblastoma cells. Brain Pathol. 2012, 22, 159–174. [Google Scholar] [CrossRef]
- Vinci, M.; Box, C.; Eccles, S.A. Three-dimensional (3D) tumor spheroid invasion assay. J. Vis. Exp. 2015, 99, e52686. [Google Scholar] [CrossRef] [Green Version]
- Kowalski-Chauvel, A.; Gouaze-Andersson, V.; Baricault, L.; Martin, E.; Delmas, C.; Toulas, C.; Cohen-Jonathan-Moyal, E.; Seva, C. Alpha6-integrin regulates FGFR1 expression through the ZEB1/YAP1 transcription complex in glioblastoma stem cells resulting in enhanced proliferation and stemness. Cancers 2019, 11, 406. [Google Scholar] [CrossRef] [Green Version]
- Data Visualization Tools for Brain Tumor Datasets. Available online: http://gliovis.bioinfo.cnio.es/ (accessed on 25 May 2022).
- Nagai, K.; Fujii, M.; Kitazume, S. Protein tyrosine phosphatase receptor type z in central nervous system disease. Int. J. Mol. Sci. 2022, 23, 4414. [Google Scholar] [CrossRef]
- Majc, B.; Sever, T.; Zaric, M.; Breznik, B.; Turk, B.; Lah, T.T. Epithelial-to-mesenchymal transition as the driver of changing carcinoma and glioblastoma microenvironment. Biochim. Biophys. Acta Mol. Cell Res. 2020, 1867, 118782. [Google Scholar] [CrossRef] [PubMed]
- Kowalski-Chauvel, A.; Lacore, M.G.; Arnauduc, F.; Delmas, C.; Toulas, C.; Cohen-Jonathan-Moyal, E.; Seva, C. The m6A RNA demethylase ALKBH5 promotes radioresistance and invasion capability of glioma stem cells. Cancers 2020, 13, 40. [Google Scholar] [CrossRef] [PubMed]
- Fujikawa, A.; Sugawara, H.; Tanaka, T.; Matsumoto, M.; Kuboyama, K.; Suzuki, R.; Tanga, N.; Ogata, A.; Masumura, M.; Noda, M. Targeting PTPRZ inhibits stem cell-like properties and tumorigenicity in glioblastoma cells. Sci. Rep. 2017, 7, 5609. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujikawa, A.; Nagahira, A.; Sugawara, H.; Ishii, K.; Imajo, S.; Matsumoto, M.; Kuboyama, K.; Suzuki, R.; Tanga, N.; Noda, M.; et al. Small-molecule inhibition of PTPRZ reduces tumor growth in a rat model of glioblastoma. Sci. Rep. 2016, 6, 20473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Honda, A.; Ito, Y.; Takahashi-Niki, K.; Matsushita, N.; Nozumi, M.; Tabata, H.; Takeuchi, K.; Igarashi, M. Extracellular signals induce glycoprotein M6a clustering of lipid rafts and associated signaling molecules. J. Neurosci. 2017, 37, 4046–4064. [Google Scholar] [CrossRef]
- Lathia, J.D.; Li, M.; Hall, P.E.; Gallagher, J.; Hale, J.S.; Wu, Q.; Venere, M.; Levy, E.; Rani, M.R.; Huang, P.; et al. Laminin alpha 2 enables glioblastoma stem cell growth. Ann. Neurol. 2012, 72, 766–778. [Google Scholar] [CrossRef]
- Liu, J.; Liu, D.; Yang, Z.; Yang, Z. High LAMC1 expression in glioma is associated with poor prognosis. Onco Targets Ther. 2019, 12, 4253–4260. [Google Scholar] [CrossRef] [Green Version]
- Sun, T.; Patil, R.; Galstyan, A.; Klymyshyn, D.; Ding, H.; Chesnokova, A.; Cavenee, W.K.; Furnari, F.B.; Ljubimov, V.A.; Shatalova, E.S.; et al. Blockade of a laminin-411-notch axis with CRISPR/Cas9 or a nanobioconjugate inhibits glioblastoma growth through tumor-microenvironment cross-talk. Cancer Res. 2019, 79, 1239–1251. [Google Scholar] [CrossRef] [Green Version]
- Yu, Q.; Xiao, W.; Sun, S.; Sohrabi, A.; Liang, J.; Seidlits, S.K. Extracellular matrix proteins confer cell adhesion-mediated drug resistance through integrin alpha v in glioblastoma cells. Front. Cell Dev. Biol. 2021, 9, 616580. [Google Scholar] [CrossRef]
- Maeda, N.; Hamanaka, H.; Shintani, T.; Nishiwaki, T.; Noda, M. Multiple receptor-like protein tyrosine phosphatases in the form of chondroitin sulfate proteoglycan. FEBS Lett. 1994, 354, 67–70. [Google Scholar] [CrossRef] [Green Version]
- Muller, S.; Kunkel, P.; Lamszus, K.; Ulbricht, U.; Lorente, G.A.; Nelson, A.M.; von Schack, D.; Chin, D.J.; Lohr, S.C.; Westphal, M.; et al. A role for receptor tyrosine phosphatase zeta in glioma cell migration. Oncogene 2003, 22, 6661–6668. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ulbricht, U.; Brockmann, M.A.; Aigner, A.; Eckerich, C.; Muller, S.; Fillbrandt, R.; Westphal, M.; Lamszus, K. Expression and function of the receptor protein tyrosine phosphatase zeta and its ligand pleiotrophin in human astrocytomas. J. Neuropathol. Exp. Neurol. 2003, 62, 1265–1275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foehr, E.D.; Lorente, G.; Kuo, J.; Ram, R.; Nikolich, K.; Urfer, R. Targeting of the receptor protein tyrosine phosphatase beta with a monoclonal antibody delays tumor growth in a glioblastoma model. Cancer Res. 2006, 66, 2271–2278. [Google Scholar] [CrossRef] [PubMed] [Green Version]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lacore, M.G.; Delmas, C.; Nicaise, Y.; Kowalski-Chauvel, A.; Cohen-Jonathan-Moyal, E.; Seva, C. The Glycoprotein M6a Is Associated with Invasiveness and Radioresistance of Glioblastoma Stem Cells. Cells 2022, 11, 2128. https://doi.org/10.3390/cells11142128
Lacore MG, Delmas C, Nicaise Y, Kowalski-Chauvel A, Cohen-Jonathan-Moyal E, Seva C. The Glycoprotein M6a Is Associated with Invasiveness and Radioresistance of Glioblastoma Stem Cells. Cells. 2022; 11(14):2128. https://doi.org/10.3390/cells11142128
Chicago/Turabian StyleLacore, Marie Geraldine, Caroline Delmas, Yvan Nicaise, Aline Kowalski-Chauvel, Elizabeth Cohen-Jonathan-Moyal, and Catherine Seva. 2022. "The Glycoprotein M6a Is Associated with Invasiveness and Radioresistance of Glioblastoma Stem Cells" Cells 11, no. 14: 2128. https://doi.org/10.3390/cells11142128