
Differential Susceptibility of Retinal Neurons to the Loss of Mitochondrial Biogenesis Factor Nrf1


1
Ruiz Department of Ophthalmology and Visual Science, McGovern Medical School at The University of Texas Health Science Center at Houston (UTHealth), 6431 Fannin St., MSB 7.024, Houston, TX 77030, USA
2
Department of Ophthalmology, Baylor College of Medicine, One Baylor Plaza, Houston, TX 77030, USA
3
The MD Anderson Cancer Center/UTHealth Graduate School of Biomedical Sciences, Houston, TX 77030, USA
*
Author to whom correspondence should be addressed.
Cells 2022, 11(14), 2203; https://doi.org/10.3390/cells11142203
Submission received: 27 April 2022
/
Revised: 10 July 2022
/
Accepted: 13 July 2022
/
Published: 14 July 2022
(This article belongs to the Collection Determinants of Neuronal Susceptibility to Mitochondrial Disease)
Abstract
:The retina, the accessible part of the central nervous system, has served as a model system to study the relationship between energy utilization and metabolite supply. When the metabolite supply cannot match the energy demand, retinal neurons are at risk of death. As the powerhouse of eukaryotic cells, mitochondria play a pivotal role in generating ATP, produce precursors for macromolecules, maintain the redox homeostasis, and function as waste management centers for various types of metabolic intermediates. Mitochondrial dysfunction has been implicated in the pathologies of a number of degenerative retinal diseases. It is well known that photoreceptors are particularly vulnerable to mutations affecting mitochondrial function due to their high energy demand and susceptibility to oxidative stress. However, it is unclear how defective mitochondria affect other retinal neurons. Nuclear respiratory factor 1 (Nrf1) is the major transcriptional regulator of mitochondrial biogenesis, and loss of Nrf1 leads to defective mitochondria biogenesis and eventually cell death. Here, we investigated how different retinal neurons respond to the loss of Nrf1. We provide in vivo evidence that the disruption of Nrf1-mediated mitochondrial biogenesis results in a slow, progressive degeneration of all retinal cell types examined, although they present different sensitivity to the deletion of Nrf1, which implicates differential energy demand and utilization, as well as tolerance to mitochondria defects in different neuronal cells. Furthermore, transcriptome analysis on rod-specific Nrf1 deletion uncovered a previously unknown role of Nrf1 in maintaining genome stability.
1. Introduction
Mitochondria biogenesis is a subcellular process through which mitochondria are replenished and remodeled through continuously importing and incorporating new proteins and lipids, replicating mitochondrial DNA, transcribing mitochondrion-encoded genes, and fusing and dividing in response to cellular demands and bioenergetics loads. This process is essential for maintaining a healthy and functional mitochondria network for energy metabolism, calcium homeostasis, biosynthesis of amino acids, cholesterol, and phospholipids, eliminating excessive reactive oxygen species, and apoptosis [1,2,3,4,5,6].
A number of transcriptional regulators, including peroxisome proliferative activated receptor gamma coactivator 1 (PGC-1) family members, nuclear respiratory factor 1 (Nrf1), and nuclear respiratory factor 2 (Nrf2/GABP), have been identified as key regulators for mitochondria biogenesis in different contexts [7,8,9,10]. Among them, Nrf1 is identified as an evolutionarily conserved transcription activator that binds to GC-rich DNA elements in promoters of a host of nuclear genes encoding proteins involved in mitochondrial structure and functions [11,12,13,14,15]. Multiple studies using ChIP-seq analysis have identified distinct sets of Nrf1’s target genes in different cell types, suggesting that Nrf1 acts in a context-dependent manner in regulating cell growth, differentiation, and mitochondrial biogenesis [16,17,18,19]. In addition, Nrf1 interacts with other proteins involved in different cellular functions. For example, it interacts directly with Auts2 to confer transcriptional activity for non-canonical polycomb repressive complex ncPRC1.3 [17]. It also interacts with poly(ADP-ribose) polymerase 1 (PARP-1) and PARP-1 PARylates DNA-binding domain of Nrf1 for transcriptional regulation [20]. Post-translational modifications on various Nrf1 domains may in addition regulate transcriptional activity in different cellular contexts [21].
Previously, we have demonstrated that defective Nrf1-mediated mitochondrial biogenesis significantly affects cell proliferation, migration, and metabolic pathways in retinal progenitor cells (RPCs) during retinal development and uncovered essential roles of Nrf1 in the survival of newly specified retinal ganglion cells (RGCs) and neurite outgrowth in retinal explants. In rod photoreceptor cells (PRs), Nrf1 deletion causes defective mitochondrial morphology, position, and function, leading to a slow, progressive degeneration of all rod PRs after 4–5 months of age. In a separate study, we further found that when Nrf1 is deleted in a subset of Tbr1-expressing OFF-RGCs [22], these RGCs also displayed a similar pattern of slow degeneration [17]. In the same study, we have also observed the reduction of Tbr1-expressing hippocampal neurons.
In the current study, we expand previous studies by covering additional types of retinal neurons and find in them differential sensitivity to Nrf1 deletion. Our findings provide in vivo evidence that Nrf1-mediated pathways have cell-type-specific functions in different retinal neural types. Through transcriptomic analyses, we further discovered a small subset of early responsive genes in Nrf1 deleted rod PRs and uncovered a novel role of Nrf1 in maintaining genome stability.
2. Materials and Methods
2.1. Animal Breeding
Nrf1flox, RhoiCre, Opn4cre, HRGP-Cre, and Pcp2-Cre mice were described previously [23,24,25,26,27,28]. We have confirmed that the expression of Cre recombinase in these lines do not contribute to the cell death phenotype described in this study. All animal procedures followed the US Public Health Service Policy on Humane Care and Use of Laboratory Animals and were approved by the Animal Welfare Committee at The University of Texas Health Science Center at Houston and the Animal Welfare Committee at the Baylor College of Medicine.
2.2. Immunohistochemistry
Flat-mounted retinas were fixed with 4% paraformaldehyde (PFA; Electron Microscopy Sciences). Retinas were then embedded in 4% low melting agarose and sectioned into 100 μm thickness with Leica VT1000S vibratome (Leica Biosystems, Deer Park, IL, USA). Retinal sections or flat-mounted retinas were incubated with the primary antibodies for 3 days at 4 °C. The primary antibodies used were rabbit anti-R/G opsin (1:1000, catalog #AB5405, Sigma), rabbit anti-B opsin (1:1000, catalog #AB5407, Sigma, St. Louis, MO, USA), rabbit anti-melanopsin (1:1000, catalog #AB-N38, Advanced Targeting Systems, Carlsbad, CA, USA), chicken anti-GFP (1:1000, catalog #A10262, Thermo Fisher Scientific, Waltham, MA, USA), rabbit anti-PKCα (1:600, catalog #P4334, Sigma), mouse anti-Syt2 (1:1000, catalog #Znp-1, DSHB, The University of Iowa, Iowa City, IA, USA), and mouse anti-Nrf1 (1:300, catalog #PCRP-NFR1-3D4; DSHB). Secondary antibodies conjugated with Alexa-488 and -555 (Thermo Fisher Scientific) were used at 1:800 dilution. DAPI (2.5 μg/mL, catalog #D1306; Thermo Fisher Scientific) was used to stain nuclei. Images were captured using a Zeiss LSM 780 confocal microscope (Carl Zeiss, White Plains, NY, USA) and exported as TIFF files into Adobe Photoshop (Adobe Systems, San Jose, CA, USA). Cell counting was conducted using the cell counter plugin in the NIH ImageJ (NIH, Bethesda, MD, USA).
2.3. Photopic Electroretinography
Photopic ERG recordings were performed on mice dark-adapted for two hours. Briefly, mice were anesthetized with ketamine/xylazine (150/10 mg/kg; ip), and the pupils were dilated for 10 min with sequential topical eye drops of 1% tropicamide and 2.5% phenylephrine (Bausch & Lomb, Tampa, FL, USA). A total of 5 μL of filtered 1X PBS was added to the surface of the eyes to prevent corneal clouding and cataract formation. Body temperature was monitored by a rectal probe (Braintree Scientific, Braintree, MA, USA) and maintained at 35 °C to 37 °C using a plastic heating coil with 43 °C circulating water. ERG from both eyes was recorded simultaneously (UTAS BigShotTM system; LKC Technologies, Gaithersburg, MD, USA) after exposure to a background illumination of 30 cd m−2 white light for 10 min. Typically, the difference in the photopic b-wave amplitudes between the two eyes were <10%. Averaged responses to 90 flashes of the intensity of 25 cd s m−2 delivered at 1 Hz were recorded. A typical recording session lasted about 20 min for each animal.
2.4. RNA-Sequencing Analysis
Four retinas isolated from male or female wild-type (WT) and Nrf1f/f;RhoiCre at 4 weeks old were pooled in the RNA later solution (Qiagen, Germantown, MD, USA). Three sets of pooled retinas were independently used for RNA-seq. Male and female retinas were sequenced separately to determine whether gender is a determining biological variable. Total RNA was extracted using an RNeasy mini kit (Qiagen, Germantown, MD, USA), and RNA sequencing was performed by Novogene (Sacramento, CA, USA). RNA libraries were prepared using standard Illumina protocols. Paired-end reads (150 nt) were obtained using an Illumina Novaseq 6000 Next Generation Sequencing instrument. Sequence reads were trimmed free of adaptor sequences and masked for low-complexity or low-quality sequences, then mapped to the mouse mm10 whole genome using the STAR (ver2.5) software (https://github.com/alexdobin/STAR). HITSeq v0.6.1 was used to count the read numbers mapped to each gene. FPKM of each gene was calculated based on the length of the gene and the read count mapped to this gene. Differential expression analyses of two groups were performed using the DESeq2 R package (2_1.6.3) (https://bioconductor.org/packages/release/bioc/html/DESeq2.html). The resulting p-values were adjusted using Benjamini and Hochberg’s approach for controlling the false discovery rate. Nrf1-dependent genes (fold change > 1.6667; adjusted p-value < 0.05) were analyzed by using the PANTHER Overrepresentation Test (Released 20220202). The raw datasets and normalized count data for each sample have been deposited in NCBI (Geo dataset: GSE150258).
2.5. Quantitative Reverse Transcriptase PCR
Two retinas from WT or Nrf1f/f;RhoiCre of multiple littermates at 4 weeks old were extracted using TRI reagent (MilliporeSigma, Burlington, MA, USA). First-strand cDNA was synthesized using iScript Reverse Transcription Super Mix for RT-qPCR (Bio-Rad, Hercules, CA, USA). Real-time PCR was performed using the CFX Connect Real-Time System (Bio-Rad, Hercules, CA, USA) with iTaq Universal SYBR Green Supermix (Bio-Rad, Hercules, CA, USA). Relative RNA levels were normalized to that of β-actin.
2.6. Terminal Deoxynucleotidyl Transferase dUTP Nick-End Labeling (TUNEL) Assay
Eye balls were extracted and fixed with 4% PFA for 1.5 hrs, and washed with PBS and cryo-sectioned into 30 μm thickness. Sections were washed with PBS then proceeded to TUNEL assay using the “In Situ Cell Death Detection Kit” (Roche Molecular Systems, Pleasanton, CA, USA). DAPI was used to stain nuclei.
2.7. Statistical Analysis
All data are presented as mean ± SD for each genotype. For all comparisons between genotypes, a two-tailed, two-sample Student’s t-test was used for all measurements and conducted in Microsoft Excel (Redmond, WA, USA). Results were considered significant when p < 0.05.
3. Results
To determine how different retinal neurons respond to the loss of Nrf1, we generated several cell type-specific Nrf1 conditional knockouts by breeding Nrf1flox allele [29] to HRGP-Cre, Opn4Cre, or Pcp2-Cre to delete Nrf1 in cone photoreceptors, intrinsically photosensitive retinal ganglion cells (ipRGCs), and a subset of cone and rod bipolar cells that express Pcp2, respectively.
3.1. Progressive Degeneration in Nrf1-Deficient Cone Photoreceptors
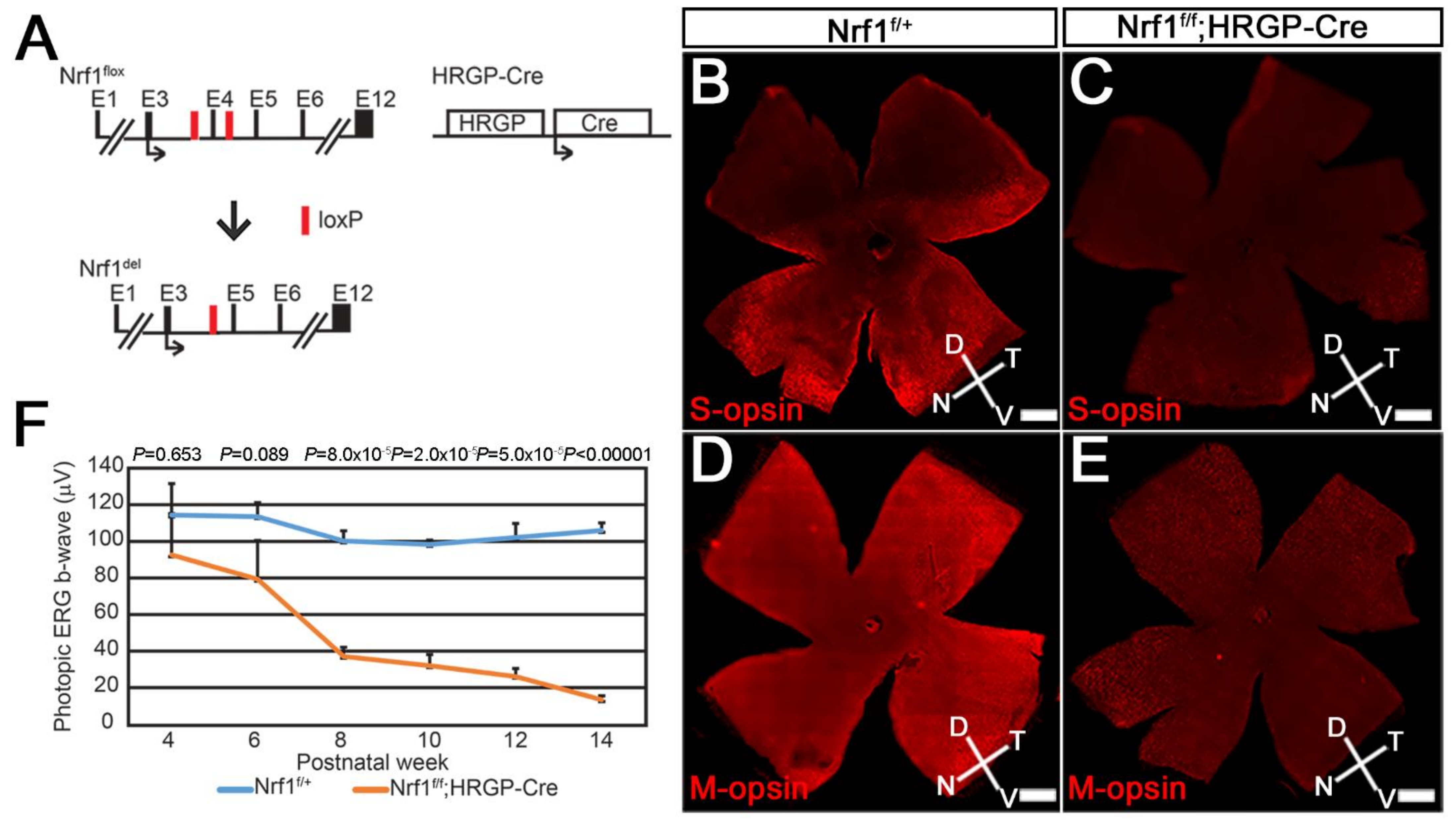
First, we examined the Nrf1 deletion mediated by HRGP-Cre (Nrf1f/f;HRGP-Cre) (Figure 1A), a transgenic mouse line in which the Cre recombinase gene is driven by human red/green pigment (HRGP) promoter specifically in M-opsin-expressing cone photoreceptors [26]. The majority of cone photoreceptors co-express middle/long wavelength-sensitive opsin (M-opsin) and short wavelength-sensitive opsin (S-opsin) in mouse retina [30]; thus, Nrf1 should be deleted in most of the cone photoreceptors in Nrf1f/f;HRGP-Cre retinas. We first examined the distribution of S-opsin+ or M-opsin+ cone photoreceptors by immuno-fluorescent staining using anti-blue opsin or anti-red/green opsin antibodies, respectively. In 3-month-old Nrf1f/+ control retinas, S-opsin+ cone cells (S-cones) were distributed in a ventral-high, dorsal-low manner (Figure 1B), consistent with previous reports [30,31,32,33]. In contrast, M-opsin+ cone cells (M-cones) were distributed in a slightly dorsal-high and ventral-low manner in the control retina (Figure 1D), although this gradient is less apparent [30]. In Nrf1f/f;HRGP-Cre retinas, the expression of S-opsin and M-opsin were significantly reduced compared to Nrf1f/+ retinas (Figure 1C,E). The reduction of S- and M-cones appears to be occurring uniformly in the entire retina without a noticeable sign of regional differences. Because of the severe reduction of S- and M-cones in 3-month-old Nrf1f/f;HRGP-Cre retinas, we next examined the functions of cone photoreceptors using photopic electroretinography (ERG). Photopic ERG b-wave amplitudes from light-adapted 4-week-old control and Nrf1-deficient mice were comparable (Figure 1F). At 6 weeks old, the photopic EGR b-wave of Nrf1-deficient mice started to decline gradually, and eventually reduced to ~10% that of the control mice at 14 weeks old (Figure 1F). ERG data indicate that deleting Nrf1 in cone photoreceptors leads to the gradual loss of cone cells and cone-mediated function.
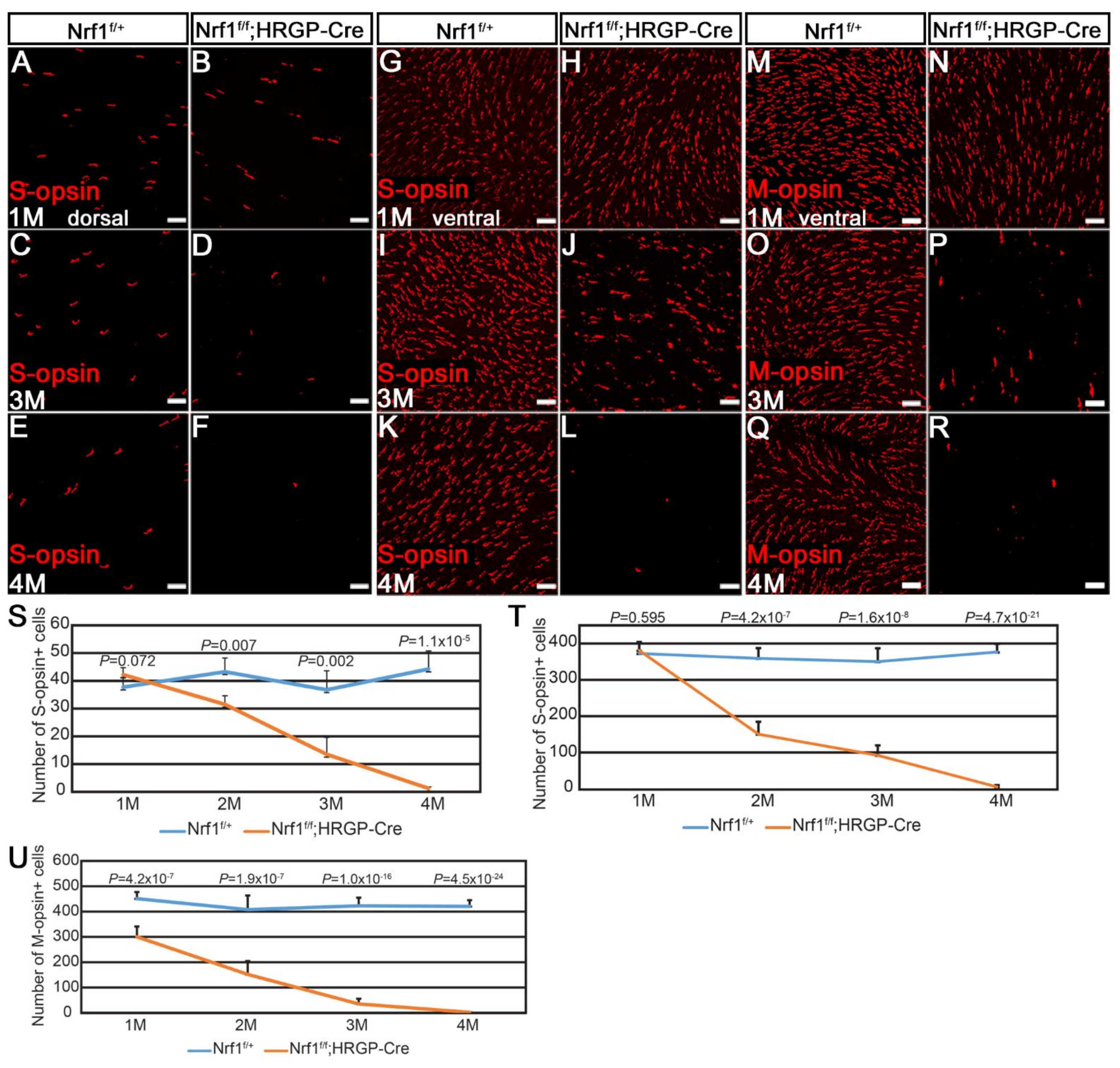
Next, we asked whether S- and M-cones respond to Nrf1 deletion differently by examining them in 1- to 4-month-old Nrf1f/+ and Nrf1f/f;HRGP-Cre retinas. Immunostaining images from different areas (dorsal and ventral for S-opsin; dorsal, ventral, nasal, and temporal for M-opsin) were counted. At 1 month old, there was no noticeable difference of S-cones between Nrf1f/+ and Nrf1f/f;HRGP-Cre retinas, both in the dorsal and ventral areas (compare Figure 2A with Figure 2B, Figure 2G with Figure 2H, and Figure 2S with Figure 2T). At 2 months old, S-cones in the ventral retinas of Nrf1-deficient mice declined to ~40% that of the Nrf1f/+ retina (Figure 2T), while in dorsal retinas, a slight, but less significant reduction of S-cones was observed (Figure 2S). At 3 months old, the number of S-cones in both dorsal and ventral retinas decreased to ~30–35% that of the Nrf1f/+ retina (compare Figure 2C with Figure 2D, Figure 2I with Figure 2J, and Figure 2S with Figure 2T). At 4 months old, hardly any S-cones can be detected in both the dorsal and ventral areas of Nrf1-deficient retinas (compare Figure 2E with Figure 2F, Figure 2K with Figure 2L, and Figure 2S with Figure 2T).
M-cones in Nrf1f/f;HRGP-Cre at 1 month old were slightly but statistically significantly less than those in the Nrf1f/+ retina (compare Figure 2M with Figure 2N, and Figure 2U). The number of M-cones decreased to ~10% that of the Nrf1f/+ retinas at 3 months old (compare Figure 2O with Figure 2P) and was almost undetectable at 4 months old (compare Figure 2Q with Figure 2R). No region-specific difference in the number and pattern of the reduction of M-cones was observed (data not shown). Together, these data indicate that the loss of Nrf1 affects both S- and M-cones in the entire retina and the loss of S-cones in the ventral retinas is slightly faster than that in the dorsal retinas.
3.2. Slow and Progressive Reduction of Nrf1-Depleted ipRGC
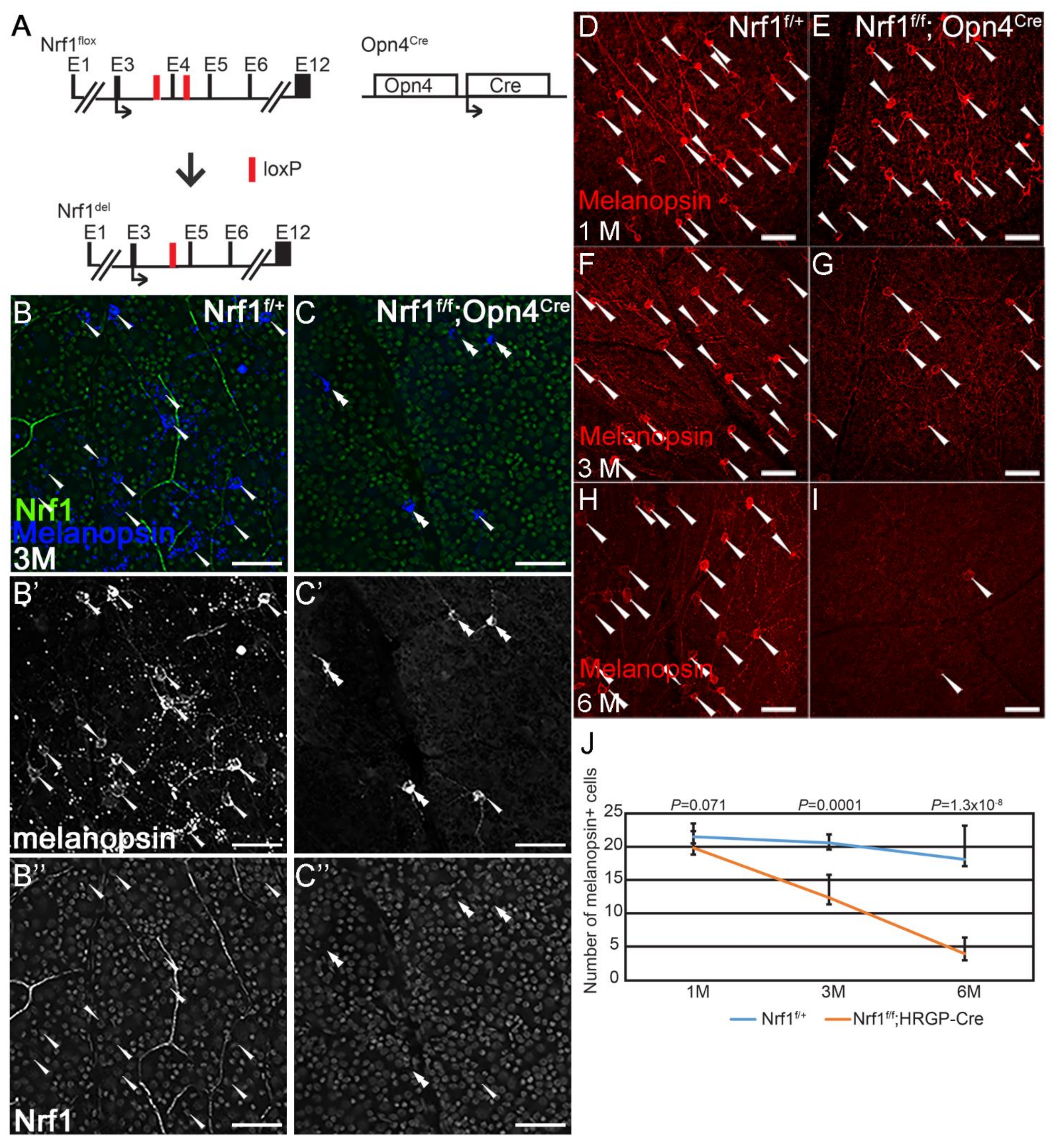
Next, we examined the effect of Nrf1-deleted ipRGCs using Opn4Cre (Figure 3A). Opn4Cre is a Cre knock-in line in which the expression of Cre is under the control of the intrinsic Opn4 promoter and regulatory regions. Opn4Cre in conjunction with Z/EG reporter has been used to detect all six types of intrinsically photosensitive retinal ganglion cells (ipRGCs) [25,34]. We first confirmed that Nrf1 is deleted in Opn4-expressing cells by conducting co-immunostaining on 3-month-old Nrf1f/+ and Nrf1f/f;Opn4Cre retinas with anti-melanopsin and anti-Nrf1 antibodies (Figure 3B,C). In Nrf1f/+ retina, all melanopsin+ ipRGCs express Nrf1 (Figure 3B–B”, n = 38). In Nrf1f/f;Opn4Cre retina, the number of melanopsin+ cells are much less than that of Nrf1f/+ (compared Figure 3B′,C′). Among the 16 melanopsin+ ipRGCs examined in Nrf1f/f;Opn4Cre retina, 15 lacked Nrf1 staining (Figure 3C′,C”), suggesting that Nrf1 has been efficiently deleted in Nrf1f/f;Opn4Cre retina.
Next, we conducted a longitudinal study to examine how Nrf1 deletion affects the survival of ipRGCs over time. We immuno-labeled 1-month, 3-month, and 6-month-old Nrf1f/+ and Nrf1f/f;Opn4Cre retinas with an anti-melanopsin antibody, then counted melanopsin+ ipRGCs. In 1-month-old retinas, the number of melanopsin+ cells in Nrf1f/+ and Nrf1f/f;Opn4Cre retinas are comparable (Figure 3D,E,J). The number of melanopsin+ cells in Nrf1f/f;Opn4Cre decreased to ~50% at 3 months old (Figure 3F,G,J), and further dropped to ~20% at 6 months old compared to Nrf1f/+ (Figure 3H–J). These results indicate that the depletion of Nrf1 caused a slower but progressive ipRGC degeneration, and unlike cones and rods, ipRGCs are more resistant to defective mitochondria biogenesis.
3.3. Highly Sensitive Type 2 and 6 Cone Bipolar Cells to Nrf1 Deletion
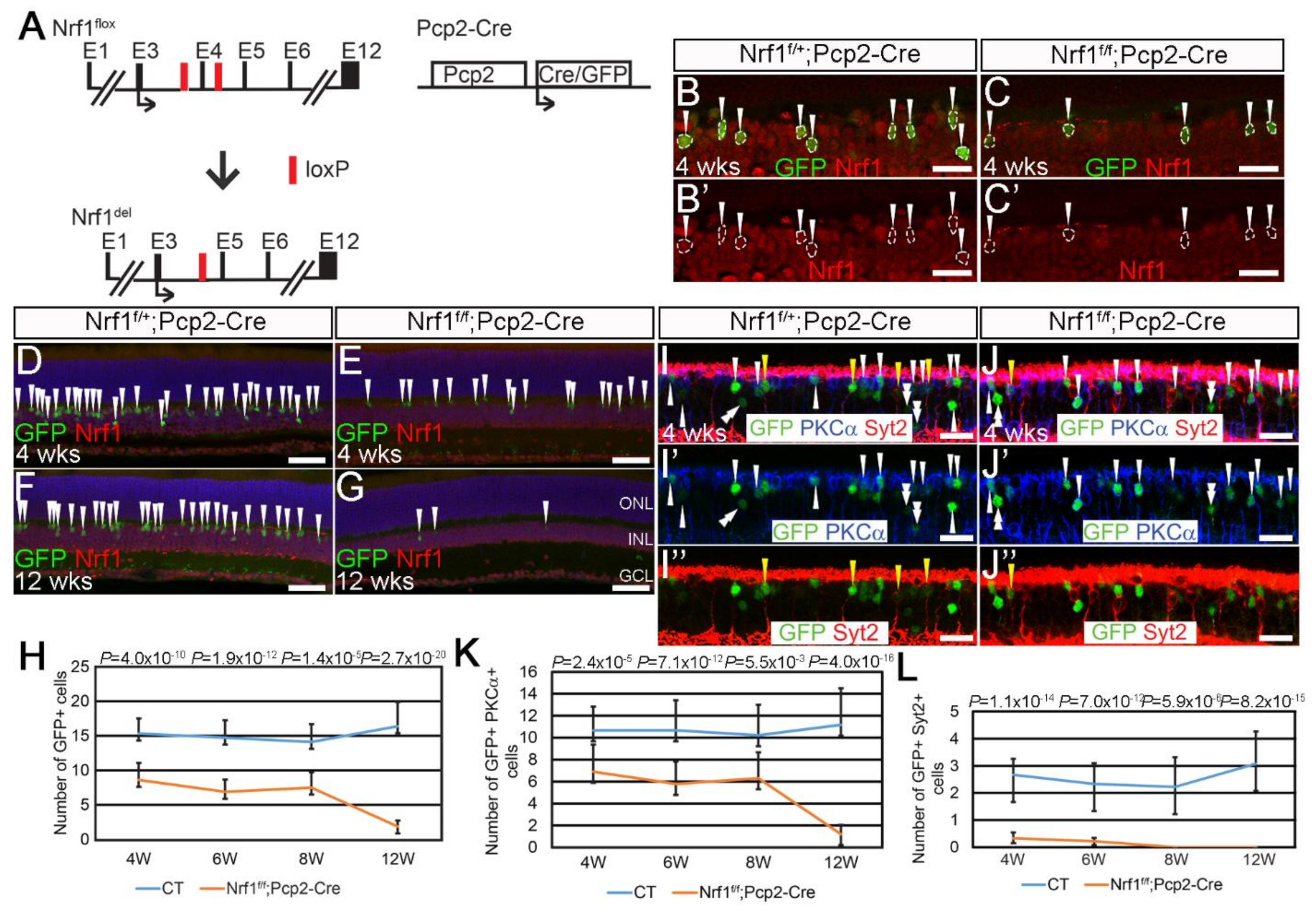
Next, we examined the Nrf1-deleted retinal bipolar cells (BCs), which are a less energy demanding cell type in retinas [35]. We used a Pcp2-Cre line to delete Nrf1 (Nrf1f/f;Pcp2-Cre) (Figure 4A) in retinal bipolar cells. The Pcp2-Cre line, when bred with the Ai9 reporter, has been shown to activate tdTomato mainly in rod BPs as well as type 2 and 6 cone BPs [36]. The Pcp2-Cre mouse line also carries a GFP reporter gene whose expression in BCs matches the tdTomato expression in Pcp2-Cre;Ai9. To confirm Nrf1 deletion in Pcp2-Cre cells, we first conducted immunostaining for GFP and Nrf1 expression on Nrf1f/+;Pcp2-Cre (control) and Nrf1f/f;Pcp2-Cre (mutant) retinas at 4 weeks old (Figure 4B,C). In control retinas, all GFP+ cells co-express Nrf1 (Figure 4B′), whereas all GFP+ cells in mutant retinas lacked Nrf1 (Figure 4C′), indicating that Pcp2-Cre is effective in deleting Nrf1 in bipolar cells.
Next, we compared the number of GFP+ BP cells between controls and mutants. At 4 weeks old, the number of GFP+ cells in mutant retinal sections was ~50% that of the controls (Figure 4D,E,H). This ratio did not change through 4- to 8-week-old retinas (Figure 4H). However, at 12 weeks old, the number of GFP+ cells in mutants dropped to ~10% that of the controls (Figure 4F–H). To distinguish whether rod or cone BPs have different sensitivity to Nrf1 deletion, we checked GFP co-expression patterns with PKCα, a rod BC marker, and Syt2, which labels type 2 and 6 cone BCs. In 4-week-old Pcp2-Cre retinas, we found that GFP+PKCα+ rod BCs and GFP+Syt2+ cone BCs account for approximately 58.8% and 23.6%, respectively, of all GFP+ cells in the inner nuclear layer (INL). The other 17.6% of PKCα−Syt2−GFP+ cells in INL are likely type 5 and 7 BCs (data not shown). On this basis, we compared GFP co-expression with PKCα and Syt2 at various ages to examine how Nrf1 deletion affects the survival of different BC subtypes over time. We conducted immunostaining on 4- to 12-week-old control and mutant retinas for rod BPs and types 2 and 6 cone BCs. At 4 weeks old, GFP+PKCα+ rod BP cells in mutants were ~60% that of the controls (Figure 4I,J,I′,J′,K). The ratio of GFP+PKCα+ cells between the controls and mutants did not change through 4 to 8 weeks. Eventually, at 12 weeks of age, GFP+PKCα+ rod BPs drop to ~10% in mutants compared to that of the controls (Figure 4K). In sharp contrast, at 4 weeks, the number of GFP+Syt2+ cone BPs in mutants dropped to ~20% compared to that of the controls (Figure 4I,J,I”,J”,L). In 8- and 12-week-old mutant retinas, we cannot detect any GFP+Syt2+ cells. These results revealed a previously unknown cellular property of Syt2+ type 2 and 6 cone BCs, i.e., that they are extremely sensitive to defective mitochondria biogenesis compared to rods, cones, RGCs, and rod BCs, suggesting that they may have higher demand for mitochondria-mediated biological functions, or that Nrf1 has a different function in them to support their functions and survival.
3.4. The Most Sensitive Genes in Nrf1-Deficient Rod Photoreceptors
Because Nrf1 deletion in all of the retinal neurons tested led to a similar pattern of progressive or rapid cell death, to understand how neurons respond to Nrf1 deletion at the transcriptional level, we used rod photoreceptors (PRs) as a model to examine how Nrf1 deletions affect gene expression. Previously, we found that in rod PR-specific Nrf1 deletion mice (Nrf1f/f;RhoiCre), the number, morphology, and location of mitochondria in rod PRs are abnormal compared to that of the controls; consequently, mitochondria-mediated energy production is impaired and severe rod degeneration takes place, followed by cone degeneration [29].
We selected 4 weeks postnatal age as the experimental time point because the onset of PR degeneration in Nrf1f/f;RhoiCre retinas has not started, while the Nrf1 has been effectively removed by RhoiCre (Figure 5A) [29]. We reasoned that those genes most sensitive to the loss of Nrf1 would display greater changes in expression levels. Six 4-week-old Nrf1f/+ (CTL) and Nrf1f/f;Rho-iCre (rod-Nrf1 deletion) retinas were collected to conduct total RNA-seq.
We first filtered the gene list by adjusting the p-value to < 0.05 and identified 1505 differentially expressed genes (DEGs), of which 695 are upregulated and 810 are downregulated in rod PR-specific Nrf1 mutants (Figure 5B). Next, we selected fold-change ≥ 2 (|(log2FC)| ≥ 1) as the cut-off to reveal Nrf1-dependent genes from these 1505 DEGs. By these stringent criteria, we only detected 25 genes whose expression between control and mutant groups was changed (Table 1). To facilitate a meaningful gene ontology analysis, we reduced the cut-off of the fold-change to ≥1.6667 (|(log2FC)| ≥ 0.7369) to define the most sensitive, early responsive Nrf1-dependent genes in rod-Nrf1 deletion. By such a criterion, we identified 58 downregulated and 28 upregulated genes in Nrf1-deficient rod PRs (Figure 5C) (Table 1). A heatmap was plotted to show the hierarchical clustering of these 86 DEGs (Figure 5D). We then compared these 86 genes with Nrf1-bound target genes identified in the P0 mouse brain (GSE161808) and found that 57 out of 86 genes are direct targets of Nrf1 (Figure 5C), placing these 57 genes as the early responsive target genes of Nrf1-deleted rod PRs (Table 1; gene names in bold). Next, because of the abnormal, mispositioned mitochondria and defective energy production observed in Nrf1-deficient PRs, we anticipated that genes involved in various mitochondrial functions would also likely be sensitive to rod-Nrf1 deletion. We compared the 86 most sensitive genes with gene lists in MitoCarta 2.0 [37,38] and revealed that only 7 genes were involved in mitochondrial functions (Figure 5C) (Table 1; genes marked with asterisk). However, when we used a more lenient criterion by setting the fold-change to ≥1.25 (|(log2FC)| ≥ 0.3219), we recovered 52 mitochondrial genes whose expression was altered in Nrf1-deficient rod PRs, and among them, 34 are direct targets of Nrf1 (Table 2; gene names in bold). We color-coded and mapped them in the functional subdomain of the mitochondria (Figure 5E). To quantitatively evaluate RNA-seq data relevant to these genes, we performed qRT-PCR analyses on a subset of these mitochondrial genes and found that these qRT-PCR analyses results were consistent with the RNA-seq data (Figure 5F).
Next, we analyzed the 86 most sensitive genes from the more stringent criteria using gene ontology biological process analysis (GO-BP). The top four categories revealed in GO terms are genes involved in the regulation of double-strand break repair and DNA recombination (Table 3). Interestingly, the genes involved in these processes, including Pot1b, Rad51ap1, Fignl1, Rbbp8, Zcwpw1, Prkdc, Cdc7, Slx4, Rad51ap1, Rbbp8, and Ino80c, are direct targets of Nrf1, suggesting that Nrf1 deletion in rod PRs led to the misregulation of its direct target genes involved in DNA repair and recombination, conceivably leading to DNA damages in rod photoreceptors. To test whether DNA damage in Nrf1-deficient rod PRs precedes the onset of other phenotypes we revealed previously through histology, electron microscopy imaging of mitochondria, and scotopic ERG [29], we conducted TUNEL assay on P21 control and rod-Nrf1 deletion retinas. Consistent with the transcriptomic data, we detected many TUNEL+ rod PRs in P21 rod-Nrf1 deletion retinas but not in the control retinas (Figure 5G,H), suggesting DNA breakage is the earliest detectable phenotype found in Nrf1 deletion rod PRs. In 7-week-old rod-Nrf1 deletion retinas, significantly more TUNEL+ cells were detected in rod PRS of rod-Nrf1 deletion retinas compared to that of control retinas (Figure 5I,J).
4. Discussion
In this report, we tested the susceptibility of various retinal neuronal subtypes to defective Nrf1-mediated biological processes including mitochondria biogenesis. We conducted a longitudinal study and tested how quickly retinal neurons degenerate when Nrf1 is genetically deleted in a cell type-specific manner. It is conceivable that all retinal neurons eventually die when Nrf1 is removed. However, our data revealed differential susceptibility of different types of retinal neurons to Nrf1 deletion.
Photoreceptors are known to be an extremely energy-demanding cell type to support several principal energy-consuming functions, such as phototransduction, outer segment disk regeneration, and active ion transport, to repolarize the plasma membrane and re-establish the transmembrane ionic gradient for re-activation [39,40,41]. In Nrf1-deleted cone photoreceptors (cone PRs), we found that their tolerance for Nrf1 deletion is similar to that of rod photoreceptors (rod PRs) [29]. In both cases, PRs degenerate within 4 to 5 months of ages following Nrf1 deletion, even though cones consume more energy than rods in bright light. In rod-Nrf1-mutants, cone PRs also degenerate, owing to the lack of survival factors from RPs [42]. In contrast, in cone-Nrf1-mutants, rod PRs remain histologically intact and functional by ERG analysis (data not shown), despite the fact that cone PRs mediate electrical coupling through a rod-cone-rod pathway [43]. It is conceivable that the major cause of PR degeneration in Nrf1-mutants is cell-autonomous and due to the defective mitochondrial biogenesis that fails to provide sufficient energy. Furthermore, the defective mitochondrial biogenesis in Nrf1-mutant PRs may lead to a defective mitochondrial network, resulting in a barrier for incoming light. A recent study found that mitochondria in cone PRs of ground squirrels may serve as microscopic lenses to help focus light on the photoreceptor pigments. In rod-Nrf1-mutants, we have observed misshaped and mispositioned mitochondria in the inner segments of rods. Whether these anomalies in mitochondrial structure and position in Nrf1-mutants directly disturb the focus of light and hence cause the decline of ERG activity remains unknown.
Among all GFP+ bipolar cells found in the Pcp2-GFP-Cre line (58% PKCα+ rod bipolar cells, 23% Syt2+ type 2 OFF- and type 6 ON-BCs, and 17% PKCa−Syt2− BCs that may be type 5 or 7) [36], we found that Syt2+ cone BCs are much more sensitive to Nrf1 deletion than rod BCs and the other subset of cone BCs. At 4 weeks of age, the number of Syt2+ cone BCs in Nrf1f/f;Pcp2-Cre retinas has significantly decreased to ~20% compared to that of the Pcp2-Cre control, while the number of the other cone BCs and rod BCs only reduced to ~60% at this stage. This observation is intriguing because even in the rod- or cone-specific Nrf1 mutant retinas, the number and their functionalities characterized by ERG remain similar to that of control littermates at this stage. Most strikingly, in 2-month-old Nrf1f/f;Pcp2-Cre retinas, Syt2+ BCs completely disappeared, while other BC types only partially degenerated. This observation reveals a previously unknown character of Syt2+ cone BCs: they are extremely sensitive to Nrf1 deletion and/or defective mitochondrial biogenesis, implicating a high demand for energy and/or mitochondrial biogenesis in this population. In a recent study using the complex-1 component Ndufs4 knockout mouse model [44], death of rod BCs was found taking place at P20, preceding the inflammatory wave that first leads to the death of amacrine cells and then RGCs at P42, suggesting that rod BCs are indeed a sensitive cell type to mitochondrial defect. Our present study supports this notion and further suggests that Syt2+ cone BCs are much more sensitive to defective mitochondrial biogenesis than rod BCs and other retinal neuronal types. Among the two Syt2+ cone BC subtypes found in Pcp2-Cre retinas, it is worth noting that the type-2 OFF BCs possess an interesting cellular property through which they import transcription factor Otx2 from photoreceptors and incorporate it into mitochondria to protect themselves from glutamate excitotoxicity and to support mitochondrial ATP synthesis [45,46]. Whether Nrf1-mediated mitochondrial biogenesis is involved in such a unique intercellular transporting process remains to be studied.
In ganglion cell populations, we previously used a sparse labeling strategy and reported that ~50% of Nrf1-deleted Tbr1-expressing OFF RGCs survive for at least 3 months following Nrf1 deletion [17]. In the present study, we measure the number of melanopsin-expressing ipRGCs and found similar results. Although these two experiments were conducted in different RGC subpopulations using different experimental settings, the similar outcome supports a notion that RGCs are relatively more resistant to defective mitochondrial biogenesis than other retinal neurons. This is rather surprising given that RGCs are most vulnerable to mitochondrial damages because of their unique cellular architecture, including the elaborate dendritic arbors in the inner plexiform layer and the long axons that extend into the brain and connect with other neurons. At the active sites of both regions, the energy demand for producing neurotransmitters, organizing synaptic vesicles, restoring ion gradients, and buffering calcium is extensive. In several human diseases caused by mutations in genes essential for mitochondrial function, including approximately 90% of Leber’s hereditary optic neuropathy cases with point mutations in mitochondria DNA [47] and 75% of autosomal dominant optic atrophy patients with mutations in OPA1 [48,49], RGCs are the most sensitive neurons and die as diseases progress. In the N-methyl-D-aspartic acid-induced excitotoxicity model in retinas, RGCs are also the most sensitive neurons due to the glutamate-evoked rise of calcium mediated by NMDA-type glutamate receptors in RGCs [50,51,52]. A simple explanation is that RGCs may have a backup mechanism temporarily compensating for the loss of Nrf1-mediated mitochondrial biogenesis pathways to some extent but not forever. Alternatively, those patients with OPA1 or mitochondrial mutations may also have unrecognized, defective neurons in other parts of the CNS that do not present notable neurological symptoms and pathology [53].
The GO-BP analysis of the 86 most sensitive genes found in Nrf1-deficient PRs reveals that the top affected biological processes in gene ontology terms encompass double-strand break repair and DNA recombination. This early response at the transcription level implies that Nrf1 plays a pivotal role in maintaining genome integrity in the central nervous system. Consistent with this idea, Nrf1 has been shown to be physically associated with Prkdc, Xrcc5/Ku70, and Xrcc6 (Ku80) [20], further strengthening Nrf1’s role in maintaining genome integrity at transcriptional and post-transcriptional levels. Additionally, genes with neuroprotective function, such as Mlf1 and Bcl2l12, were upregulated in the Nrf1-mutant, likely due to DNA damage and other cellular stresses prior to the onset of degeneration.
When we used a more relaxed criterion in differential gene expression analysis, we found more mitochondrial genes whose expression is modestly affected in Nrf1-deleted rod PRs. These genes are different from those found in Nrf1-deficient retinal progenitor cells (RPCs) in embryonic retina [29]. For example, expression of several ribosomal or translation-related genes that may be involved in the mitochondrial translation system, including a subset of Mrpl genes and Rpl10a, Rpusd3, Sars2, Mtg2, and Vars2, were altered in Nrf1-deficient rod PRs but not in Nrf1-deficient RPCs. 8-Oxoguanine (8-oxoG) is one of the major base lesions in oxidative stressed DNA [54]. 8-oxoG can pair with adenine and cytosine, leading to base-substitution mutagenesis [55]. Mutyh, an adenine DNA glycosylase, removes misincorporated adenine in template DNA that pairs with 8-oxoG in the opposite strand [56]. The upregulation of Mutyh in rod-Nrf1-deleted retinas supports the notion that the damaged PRs activate DNA repair machinery. Hexokinase 1 (HK1), which functions at the first step of the glycolysis pathway, was upregulated in Nrf1-deficient rods but downregulated in Nrf1-deficient RPCs, suggesting that Nrf1 deletion affects metabolic pathways differently in neural development and degeneration states. Additionally, photoreceptors are known to be glycolytic [57,58,59]. We also found that the expression of 6 genes involved in glycolysis pathway were modestly affected in rod-Nrf1-deleted retinas (Table 4), suggesting that rod-specific Nrf1 deletion moderately affects glycolysis and mitochondrial energy production pathways. In addition, RNA-seq revealed a subset of misregulated genes involved in neurodegeneration in CNS. For example, Bardet-Biedl syndrome 2 protein homolog Bbs2 is downregulated by ~0.5 fold in rod-Nrf1-deleted retinas. It has been shown that Bbs2 mutation causes photoreceptor degeneration with mislocalized rhodopsin [60]. Ska3, an important kinetochore component involved in proper chromosome segregation during mitosis [61], is upregulated in the Nrf1-mutant to ~1.8 fold. It has been shown that Ska3 upregulation activates PI3K/Akt signaling pathway, which promotes cell proliferation in cervical cancer [62]. Whether the upregulation of Ska3 in Nrf1-deleted photoreceptor leads to aberrant cell cycle re-entry and cell death remains unclear [63].
Overall, our findings offer a deeper understanding of how defective Nrf1-mediated mitochondrial biogenesis might contribute to the pathology and progression of neurodegeneration in the retina, and suggest that a fine-tuned Nrf1 expression level is essential for safeguarding genetic fidelity indispensable for the long life of mature neurons.
Author Contributions
Conceptualization, primary experimentation design and data analysis, and writing, all authors; Transcriptome data generation and analysis, T.K. and C.-A.M. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by grants from the National Institutes of Health-National Eye Institute to C.A.M. (EY024376) and C.K.C. (EY026930), and in part by National Eye Institute Vision Core Grants P30EY028102 (UTHealth) and P30EY002520 (BCM). The Ophthalmology Department of Baylor College Medicine received an unrestricted grant from Research to Prevent Blindness, Inc. C.K.C. is the Alice McPherson Retinal Research Foundation Chair at BCM.
Institutional Review Board Statement
The animal study protocols were approved by the Animal Welfare Committee at The University of Texas Health Science Center at Houston (AWC-21–102; Date of approval: 8 October 2021) and the Animal Welfare Committee at the Baylor College of Medicine (AN-6373, date of approval: 15 August 2019).
Informed Consent Statement
Not applicable.
Data Availability Statement
RNA-seq data described in this paper have been deposited in the GEO database (GSE150258).
Acknowledgments
The authors acknowledge Tiffany Wang for proofreading and editing the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Chan, D.C. Mitochondrial fusion and fission in mammals. Annu. Rev. Cell Dev. Biol. 2006, 22, 79–99. [Google Scholar] [CrossRef] [Green Version]
- Miranda, M.; Bonekamp, N.A.; Kühl, I. Starting the engine of the powerhouse: Mitochondrial transcription and beyond. Biol. Chem. 2022, 403, 779–805. [Google Scholar] [CrossRef] [PubMed]
- Bomba-Warczak, E.; Savas, J.N. Long-lived mitochondrial proteins and why they exist. Trends Cell Biol. 2022. [Google Scholar] [CrossRef] [PubMed]
- Vafai, S.B.; Mootha, V.K. Mitochondrial disorders as windows into an ancient organelle. Nature 2012, 491, 374–383. [Google Scholar] [CrossRef] [PubMed]
- Giarmarco, M.M.; Brock, D.C.; Robbings, B.M.; Cleghorn, W.M.; Tsantilas, K.A.; Kuch, K.C.; Brockerhoff, S.E. Daily mitochondrial dynamics in cone photoreceptors. Proc. Natl. Acad. Sci. USA 2020, 117, 28816–28827. [Google Scholar] [CrossRef]
- Chang, J.Y.; Shi, L.; Ko, M.L.; Ko, G.Y. Circadian Regulation of Mitochondrial Dynamics in Retinal Photoreceptors. J. Biol. Rhythm. 2018, 33, 151–165. [Google Scholar] [CrossRef]
- Wu, Z.; Puigserver, P.; Andersson, U.; Zhang, C.; Adelmant, G.; Mootha, V.; Troy, A.; Cinti, S.; Lowell, B.; Scarpulla, R.C.; et al. Mechanisms controlling mitochondrial biogenesis and respiration through the thermogenic coactivator PGC-1. Cell 1999, 98, 115–124. [Google Scholar] [CrossRef] [Green Version]
- Virbasius, C.A.; Virbasius, J.V.; Scarpulla, R.C. NRF-1, an activator involved in nuclear-mitochondrial interactions, utilizes a new DNA-binding domain conserved in a family of developmental regulators. Genes Dev. 1993, 7, 2431–2445. [Google Scholar] [CrossRef]
- Hock, M.B.; Kralli, A. Transcriptional control of mitochondrial biogenesis and function. Annu. Rev. Physiol. 2009, 71, 177–203. [Google Scholar] [CrossRef] [Green Version]
- Spiegelman, B.M. Transcriptional control of mitochondrial energy metabolism through the PGC1 coactivators. Novartis Found. Symp. 2007, 287, 60–63. [Google Scholar]
- Calzone, F.J.; Hoog, C.; Teplow, D.B.; Cutting, A.E.; Zeller, R.W.; Britten, R.J.; Davidson, E.H. Gene regulatory factors of the sea urchin embryo. I. Purification by affinity chromatography and cloning of P3A2, a novel DNA-binding protein. Development 1991, 112, 335–350. [Google Scholar] [CrossRef] [PubMed]
- DeSimone, S.M.; White, K. The Drosophila erect wing gene, which is important for both neuronal and muscle development, encodes a protein which is similar to the sea urchin P3A2 DNA binding protein. Mol. Cell Biol. 1993, 13, 3641–3649. [Google Scholar]
- Becker, T.S.; Burgess, S.M.; Amsterdam, A.H.; Allende, M.L.; Hopkins, N. not really finished is crucial for development of the zebrafish outer retina and encodes a transcription factor highly homologous to human Nuclear Respiratory Factor-1 and avian Initiation Binding Repressor. Development 1998, 125, 4369–4378. [Google Scholar] [CrossRef] [PubMed]
- Schaefer, L.; Engman, H.; Miller, J.B. Coding sequence, chromosomal localization, and expression pattern of Nrf1: The mouse homolog of Drosophila erect wing. Mamm. Genome 2000, 11, 104–110. [Google Scholar] [CrossRef] [PubMed]
- Evans, M.J.; Scarpulla, R.C. NRF-1: A trans-activator of nuclear-encoded respiratory genes in animal cells. Genes Dev. 1990, 4, 1023–1034. [Google Scholar] [CrossRef] [Green Version]
- Satoh, J.; Kawana, N.; Yamamoto, Y. Pathway Analysis of ChIP-Seq-Based NRF1 Target Genes Suggests a Logical Hypothesis of their Involvement in the Pathogenesis of Neurodegenerative Diseases. Gene Regul. Syst. Biol. 2013, 7, 139–152. [Google Scholar] [CrossRef] [Green Version]
- Liu, S.; Aldinger, K.A.; Cheng, C.V.; Kiyama, T.; Dave, M.; McNamara, H.K.; Zhao, W.; Stafford, J.M.; Descostes, N.; Lee, P.; et al. NRF1 association with AUTS2-Polycomb mediates specific gene activation in the brain. Mol. Cell 2021, 81, 4663–4676.e8. [Google Scholar] [CrossRef]
- Takayama, K.I.; Kosaka, T.; Suzuki, T.; Hongo, H.; Oya, M.; Fujimura, T.; Suzuki, Y.; Inoue, S. Subtype-specific collaborative transcription factor networks are promoted by OCT4 in the progression of prostate cancer. Nat. Commun. 2021, 12, 3766. [Google Scholar] [CrossRef]
- Zhang, J.; Wang, C.; Chen, X.; Takada, M.; Fan, C.; Zheng, X.; Wen, H.; Liu, Y.; Pestell, R.G.; Aird, K.M.; et al. EglN2 associates with the NRF1-PGC1α complex and controls mitochondrial function in breast cancer. EMBO J. 2015, 34, 2953–2970. [Google Scholar] [CrossRef]
- Hossain, M.B.; Ji, P.; Anish, R.; Jacobson, R.H.; Takada, S. Poly(ADP-ribose) Polymerase 1 Interacts with Nuclear Respiratory Factor 1 (NRF-1) and Plays a Role in NRF-1 Transcriptional Regulation. J. Biol. Chem. 2009, 284, 8621–8632. [Google Scholar] [CrossRef] [Green Version]
- Herzig, R.P.; Andersson, U.; Scarpulla, R.C. Dynein light chain interacts with NRF-1 and EWG, structurally and functionally related transcription factors from humans and drosophila. J. Cell Sci. 2000, 113, 4263–4273. [Google Scholar] [CrossRef] [PubMed]
- Kiyama, T.; Long, Y.; Chen, C.K.; Whitaker, C.M.; Shay, A.; Wu, H.; Badea, T.C.; Mohsenin, A.; Parker-Thornburg, J.; Klein, W.H.; et al. Essential Roles of Tbr1 in the Formation and Maintenance of the Orientation-Selective J-RGCs and a Group of OFF-Sustained RGCs in Mouse. Cell Rep. 2019, 27, 900–915.e5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scarpulla, R.C.; Vega, R.B.; Kelly, D.P. Transcriptional integration of mitochondrial biogenesis. Trends Endocrinol. Metab. 2012, 23, 459–466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, S.; Chen, D.; Sauvé, Y.; McCandless, J.; Chen, Y.J.; Chen, C.K. Rhodopsin-iCre transgenic mouse line for Cre-mediated rod-specific gene targeting. Genesis 2005, 41, 73–80. [Google Scholar] [CrossRef]
- Ecker, J.L.; Dumitrescu, O.N.; Wong, K.Y.; Alam, N.M.; Chen, S.K.; LeGates, T.; Renna, J.M.; Prusky, G.T.; Berson, D.M.; Hattar, S. Melanopsin-expressing retinal ganglion-cell photoreceptors: Cellular diversity and role in pattern vision. Neuron 2010, 67, 49–60. [Google Scholar] [CrossRef] [Green Version]
- Le, Y.Z.; Ash, J.D.; Al-Ubaidi, M.R.; Chen, Y.; Ma, J.X.; Anderson, R.E. Targeted expression of Cre recombinase to cone photoreceptors in transgenic mice. Mol. Vis. 2004, 10, 1011–1018. [Google Scholar]
- Barski, J.J.; Dethleffsen, K.; Meyer, M. Cre recombinase expression in cerebellar Purkinje cells. Genesis 2000, 28, 93–98. [Google Scholar] [CrossRef]
- Lewis, P.M.; Gritli-Linde, A.; Smeyne, R.; Kottmann, A.; McMahon, A.P. Sonic hedgehog signaling is required for expansion of granule neuron precursors and patterning of the mouse cerebellum. Dev. Biol. 2004, 270, 393–410. [Google Scholar] [CrossRef] [Green Version]
- Kiyama, T.; Chen, C.K.; Wang, S.W.; Pan, P.; Ju, Z.; Wang, J.; Takada, S.; Klein, W.H.; Mao, C.A. Essential roles of mitochondrial biogenesis regulator Nrf1 in retinal development and homeostasis. Mol. Neurodegener. 2018, 13, 56. [Google Scholar] [CrossRef]
- Applebury, M.L.; Antoch, M.P.; Baxter, L.C.; Chun, L.L.; Falk, J.D.; Farhangfar, F.; Kage, K.; Krzystolik, M.G.; Lyass, L.A.; Robbins, J.T. The murine cone photoreceptor: A single cone type expresses both S and M opsins with retinal spatial patterning. Neuron 2000, 27, 513–523. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.V.; Weick, M.; Demb, J.B. Spectral and temporal sensitivity of cone-mediated responses in mouse retinal ganglion cells. J. Neurosci. 2011, 31, 7670–7681. [Google Scholar] [CrossRef] [PubMed]
- Chang, L.; Breuninger, T.; Euler, T. Chromatic coding from cone-type unselective circuits in the mouse retina. Neuron 2013, 77, 559–571. [Google Scholar] [CrossRef] [Green Version]
- Nadal-Nicolas, F.M.; Kunze, V.P.; Ball, J.M.; Peng, B.T.; Krishnan, A.; Zhou, G.; Dong, L.; Li, W. True S-cones are concentrated in the ventral mouse retina and wired for color detection in the upper visual field. eLife 2020, 9, e56840. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, T.M.; Chen, S.K.; Hattar, S. Intrinsically photosensitive retinal ganglion cells: Many subtypes, diverse functions. Trends Neurosci. 2011, 34, 572–580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wong-Riley, M.T. Energy metabolism of the visual system. Eye Brain 2010, 2, 99–116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, Q.; Ivanova, E.; Ganjawala, T.H.; Pan, Z.H. Cre-mediated recombination efficiency and transgene expression patterns of three retinal bipolar cell-expressing Cre transgenic mouse lines. Mol. Vis. 2013, 19, 1310–1320. [Google Scholar]
- Pagliarini, D.J.; Calvo, S.E.; Chang, B.; Sheth, S.A.; Vafai, S.B.; Ong, S.E.; Walford, G.A.; Sugiana, C.; Boneh, A.; Chen, W.K.; et al. A mitochondrial protein compendium elucidates complex I disease biology. Cell 2008, 134, 112–123. [Google Scholar] [CrossRef] [Green Version]
- Calvo, S.E.; Clauser, K.R.; Mootha, V.K. MitoCarta2.0: An updated inventory of mammalian mitochondrial proteins. Nucleic Acids Res. 2016, 44, D1251–D1257. [Google Scholar] [CrossRef] [Green Version]
- LaVail, M.M. Rod outer segment disk shedding in rat retina: Relationship to cyclic lighting. Science 1976, 194, 1071–1074. [Google Scholar] [CrossRef]
- Okawa, H.; Sampath, A.P.; Laughlin, S.B.; Fain, G.L. ATP consumption by mammalian rod photoreceptors in darkness and in light. Curr. Biol. 2008, 18, 1917–1921. [Google Scholar] [CrossRef] [Green Version]
- Niven, J.E.; Laughlin, S.B. Energy limitation as a selective pressure on the evolution of sensory systems. J. Exp. Biol. 2008, 211, 1792–1804. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ait-Ali, N.; Fridlich, R.; Millet-Puel, G.; Clerin, E.; Delalande, F.; Jaillard, C.; Blond, F.; Perrocheau, L.; Reichman, S.; Byrne, L.C.; et al. Rod-derived cone viability factor promotes cone survival by stimulating aerobic glycolysis. Cell 2015, 161, 817–832. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, N.; Zhang, Z.; Keung, J.; Youn, S.B.; Ishibashi, M.; Tian, L.M.; Marshak, D.W.; Solessio, E.; Umino, Y.; Fahrenfort, I.; et al. Molecular and functional architecture of the mouse photoreceptor network. Sci. Adv. 2020, 6, eaba7232. [Google Scholar] [CrossRef] [PubMed]
- Song, L.; Yu, A.; Murray, K.; Cortopassi, G. Bipolar cell reduction precedes retinal ganglion neuron loss in a complex 1 knockout mouse model. Brain Res. 2017, 1657, 232–244. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.T.; Kim, S.J.; Sohn, Y.I.; Paik, S.S.; Caplette, R.; Simonutti, M.; Moon, K.H.; Lee, E.J.; Min, K.W.; Kim, M.J.; et al. Mitochondrial Protection by Exogenous Otx2 in Mouse Retinal Neurons. Cell Rep. 2015, 13, 990–1002. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.T.; Prochiantz, A.; Kim, J.W. Donating Otx2 to support neighboring neuron survival. BMB Rep. 2016, 49, 69–70. [Google Scholar] [CrossRef] [Green Version]
- Brown, M.D.; Starikovskaya, E.; Derbeneva, O.; Hosseini, S.; Allen, J.C.; Mikhailovskaya, I.E.; Sukernik, R.I.; Wallace, D.C. The role of mtDNA background in disease expression: A new primary LHON mutation associated with Western Eurasian haplogroup J. Hum. Genet. 2002, 110, 130–138. [Google Scholar] [CrossRef]
- Delettre, C.; Lenaers, G.; Griffoin, J.M.; Gigarel, N.; Lorenzo, C.; Belenguer, P.; Pelloquin, L.; Grosgeorge, J.; Turc-Carel, C.; Perret, E.; et al. Nuclear gene OPA1, encoding a mitochondrial dynamin-related protein, is mutated in dominant optic atrophy. Nat. Genet. 2000, 26, 207–210. [Google Scholar] [CrossRef]
- Alexander, C.; Votruba, M.; Pesch, U.E.; Thiselton, D.L.; Mayer, S.; Moore, A.; Rodriguez, M.; Kellner, U.; Leo-Kottler, B.; Auburger, G.; et al. OPA1, encoding a dynamin-related GTPase, is mutated in autosomal dominant optic atrophy linked to chromosome 3q28. Nat. Genet. 2000, 26, 211–215. [Google Scholar] [CrossRef]
- Siliprandi, R.; Canella, R.; Carmignoto, G.; Schiavo, N.; Zanellato, A.; Zanoni, R.; Vantini, G. N-methyl-D-aspartate-induced neurotoxicity in the adult rat retina. Vis. Neurosci. 1992, 8, 567–573. [Google Scholar] [CrossRef]
- Lipton, S.A.; Rosenberg, P.A. Excitatory amino acids as a final common pathway for neurologic disorders. N. Engl. J. Med. 1994, 330, 613–622. [Google Scholar] [CrossRef] [PubMed]
- Shaw, P.J. Excitatory amino acid receptors, excitotoxicity, and the human nervous system. Curr. Opin. Neurol. Neurosurg. 1993, 6, 414–422. [Google Scholar] [PubMed]
- Yu-Wai-Man, P.; Griffiths, P.G.; Gorman, G.S.; Lourenco, C.M.; Wright, A.F.; Auer-Grumbach, M.; Toscano, A.; Musumeci, O.; Valentino, M.L.; Caporali, L.; et al. Multi-system neurological disease is common in patients with OPA1 mutations. Brain 2010, 133, 771–786. [Google Scholar] [CrossRef] [PubMed]
- Kasai, H.; Hayami, H.; Yamaizumi, Z.; Saitô, H.; Nishimura, S. Detection and identification of mutagens and carcinogens as their adducts with guanosine derivatives. Nucleic Acids Res. 1984, 12, 2127–2136. [Google Scholar] [CrossRef] [Green Version]
- Maki, H. Origins of spontaneous mutations: Specificity and directionality of base-substitution, frameshift, and sequence-substitution mutageneses. Annu. Rev. Genet. 2002, 36, 279–303. [Google Scholar] [CrossRef]
- Nakabeppu, Y.; Kajitani, K.; Sakamoto, K.; Yamaguchi, H.; Tsuchimoto, D. MTH1, an oxidized purine nucleoside triphosphatase, prevents the cytotoxicity and neurotoxicity of oxidized purine nucleotides. DNA Repair 2006, 5, 761–772. [Google Scholar] [CrossRef]
- Chinchore, Y.; Begaj, T.; Wu, D.; Drokhlyansky, E.; Cepko, C.L. Glycolytic reliance promotes anabolism in photoreceptors. eLife 2017, 6, e25946. [Google Scholar] [CrossRef]
- Winkler, B.S. Glycolytic and oxidative metabolism in relation to retinal function. J. Gen. Physiol. 1981, 77, 667–692. [Google Scholar] [CrossRef] [Green Version]
- Bisbach, C.M.; Hass, D.T.; Robbings, B.M.; Rountree, A.M.; Sadilek, M.; Sweet, I.R.; Hurley, J.B. Succinate Can Shuttle Reducing Power from the Hypoxic Retina to the O2-Rich Pigment Epithelium. Cell Rep. 2020, 31, 107606. [Google Scholar] [CrossRef]
- Nishimura, D.Y.; Fath, M.; Mullins, R.F.; Searby, C.; Andrews, M.; Davis, R.; Andorf, J.L.; Mykytyn, K.; Swiderski, R.E.; Yang, B.; et al. Bbs2-null mice have neurosensory deficits, a defect in social dominance, and retinopathy associated with mislocalization of rhodopsin. Proc. Natl. Acad. Sci. USA 2004, 101, 16588–16593. [Google Scholar] [CrossRef] [Green Version]
- Abad, M.A.; Zou, J.; Medina-Pritchard, B.; Nigg, E.A.; Rappsilber, J.; Santamaria, A.; Jeyaprakash, A.A. Ska3 Ensures Timely Mitotic Progression by Interacting Directly with Microtubules and Ska1 Microtubule Binding Domain. Sci. Rep. 2016, 6, 34042. [Google Scholar] [CrossRef] [PubMed]
- Hu, R.; Wang, M.Q.; Niu, W.B.; Wang, Y.J.; Liu, Y.Y.; Liu, L.Y.; Wang, M.; Zhong, J.; You, H.Y.; Wu, X.H.; et al. SKA3 promotes cell proliferation and migration in cervical cancer by activating the PI3K/Akt signaling pathway. Cancer Cell Int. 2018, 18, 183. [Google Scholar] [CrossRef] [PubMed]
- Kruman, I.I.; Wersto, R.P.; Cardozo-Pelaez, F.; Smilenov, L.; Chan, S.L.; Chrest, F.J.; Emokpae, R., Jr.; Gorospe, M.; Mattson, M.P. Cell cycle activation linked to neuronal cell death initiated by DNA damage. Neuron 2004, 41, 549–561. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Nrf1 conditional knockout by HRGP-Cre causes impaired cone photoreceptor function. (A) Schematic diagram showing Nrf1flox allele, Nrf1-deleted allele and HRGP-Cre allele. Exons are indicated as E1–E12. Black arrows indicate the translational start sites for Nrf1 and Cre protein. Red boxes indicate loxP sites. (B–E) Immunostaining of 3-month-old Nrf1f/+ (B,D) and Nrf1f/f;HRGP-Cre (C,E) flatmount retinas showing the expression of S-opsin (B,C) and M-opsin (D,E). (F) ERG of Nrf1f/+ and Nrf1f/f;HRGP-Cre mice under light-adapted conditions. Scale bars: 500 μm.
Figure 1.
Nrf1 conditional knockout by HRGP-Cre causes impaired cone photoreceptor function. (A) Schematic diagram showing Nrf1flox allele, Nrf1-deleted allele and HRGP-Cre allele. Exons are indicated as E1–E12. Black arrows indicate the translational start sites for Nrf1 and Cre protein. Red boxes indicate loxP sites. (B–E) Immunostaining of 3-month-old Nrf1f/+ (B,D) and Nrf1f/f;HRGP-Cre (C,E) flatmount retinas showing the expression of S-opsin (B,C) and M-opsin (D,E). (F) ERG of Nrf1f/+ and Nrf1f/f;HRGP-Cre mice under light-adapted conditions. Scale bars: 500 μm.

Figure 2.
Nrf1 conditional knockout by HRGP-Cre causes degeneration of S- and M-opsin. (A–R) Immunostaining of Nrf1f/+ (A,C,E,G,I,K,M,O,Q) and Nrf1f/f;HRGP-Cre (B,D,F,H,J,L,N,P,R) retinal flat-mounts with anti-B opsin (A–L) and anti-RG opsin (M–R) at 1 month, 3 months, and 4 months old. (S,T) Number of S-opsin+ cells at dorsal area (S) and ventral area (T) of Nrf1f/+ and Nrf1f/f;HRGP-Cre retinal flatmount. (U) Number of M-opsin+ cells of Nrf1f/+ and Nrf1f/f;HRGP-Cre retinal flatmount. Scale bars: 20 μm.
Figure 2.
Nrf1 conditional knockout by HRGP-Cre causes degeneration of S- and M-opsin. (A–R) Immunostaining of Nrf1f/+ (A,C,E,G,I,K,M,O,Q) and Nrf1f/f;HRGP-Cre (B,D,F,H,J,L,N,P,R) retinal flat-mounts with anti-B opsin (A–L) and anti-RG opsin (M–R) at 1 month, 3 months, and 4 months old. (S,T) Number of S-opsin+ cells at dorsal area (S) and ventral area (T) of Nrf1f/+ and Nrf1f/f;HRGP-Cre retinal flatmount. (U) Number of M-opsin+ cells of Nrf1f/+ and Nrf1f/f;HRGP-Cre retinal flatmount. Scale bars: 20 μm.

Figure 3.
Nrf1 conditional deletion by Opn4cre causes slow degeneration of ipRGCs. (A) Schematic representation of Nrf1flox allele, Nrf1-deleted allele, and Opn4Cre allele. Exons are indicated as E1–E12. Black arrows indicate the translational start sites for Nrf1 and Cre protein. Red boxes indicate loxP sites. (B,C) Immunostaining of 3-month-old Nrf1f/+ (B) and Nrf1f/f;Opn4cre (C) retinal flat-mounts with anti-melanopsin (B′,C′) and anti-Nrf1 (B″,C″) antibodies. (B,C) merged images. Melanopsin+ and Nrf1+ cells are indicated with arrowheads. Melanopsin+ Nrf1− cells are indicated with double arrowheads. (D–I) Immunostaining of Nrf1f/+ (D,F,H) and Nrf1f/f;Opn4cre (E,G,I) retinal flat-mounts with anti-melanopsin antibody. Melanopsin+ cells are indicated with arrowhead. (J) Number of melanopsin+ cells of Nrf1f/+ and Nrf1f/f;Opn4cre retinal flat-mounts at 1 month, 3 months, and 6 months old. Scale bars: 50 μm.
Figure 3.
Nrf1 conditional deletion by Opn4cre causes slow degeneration of ipRGCs. (A) Schematic representation of Nrf1flox allele, Nrf1-deleted allele, and Opn4Cre allele. Exons are indicated as E1–E12. Black arrows indicate the translational start sites for Nrf1 and Cre protein. Red boxes indicate loxP sites. (B,C) Immunostaining of 3-month-old Nrf1f/+ (B) and Nrf1f/f;Opn4cre (C) retinal flat-mounts with anti-melanopsin (B′,C′) and anti-Nrf1 (B″,C″) antibodies. (B,C) merged images. Melanopsin+ and Nrf1+ cells are indicated with arrowheads. Melanopsin+ Nrf1− cells are indicated with double arrowheads. (D–I) Immunostaining of Nrf1f/+ (D,F,H) and Nrf1f/f;Opn4cre (E,G,I) retinal flat-mounts with anti-melanopsin antibody. Melanopsin+ cells are indicated with arrowhead. (J) Number of melanopsin+ cells of Nrf1f/+ and Nrf1f/f;Opn4cre retinal flat-mounts at 1 month, 3 months, and 6 months old. Scale bars: 50 μm.

Figure 4.
Nrf1 conditional deletion by Pcp2-Cre causes rapid degeneration of type 2 and 6 cone bipolar cells. (A) Schematic illustration of Nrf1flox, Nrf1del, and Pcp2-Cre alleles. Exons are indicated as E1–E12. Black arrows indicate the translational start sites for Nrf1 and Cre protein. Red boxes indicate loxP sites. (B,C) Immunostaining of 4-week-old Nrf1f/+;Pcp2-Cre (B,B′) and Nrf1f/f;Pcp2-Cre (C,C′) retinas with anti-GFP (B,C) and anti-Nrf1 (B,B′,C,C′) antibodies. GFP+ cells are indicated with arrowhead. (D–G) Immunostaining of 4- and 12-week-old Nrf1f/+;Pcp2-Cre (D,F) and Nrf1f/f;Pcp2-Cre (E,G) retinas with anti-GFP and anti-Nrf1 antibodies. GFP+ cells are indicated with arrowheads. (H) Number of GFP+ cells of Nrf1f/+;Pcp2-Cre and Nrf1f/f;Pcp2-Cre retinas at 4, 6, 8, and 12 weeks old. (I,J) Immunostaining of Nrf1f/+;Pcp2-Cre (I,I′,I″) and Nrf1f/f;Pcp2-Cre (J,J′,J″) retinas at 4 weeks old with anti-GFP (I–I″,J–J″), anti-PKCα (I,I′,J,J′), and anti-Syt2 (I,I″,J,J″) antibodies. GFP+PKCα+ cells, GFP+Syt2+ cells, and GFP+PKCα−Syt2− cells are indicated with white arrowheads, yellow arrowheads, and white double-arrowheads, respectively. (K) Number of GFP+PKCα+ cells of Nrf1f/+;Pcp2-Cre and Nrf1f/f;Pcp2-Cre retinas at 4, 6, 8, and 12 weeks old. (L) Number of GFP+Syt2+ cells of Nrf1f/+;Pcp2-Cre and Nrf1f/f;Pcp2-Cre retinas at 4, 6, 8, and 12 weeks old. Scale bars: 20 μm in (B,B′,C,C′,I,I′,I″,J,J′,J″) and 50 μm in (D–G).
Figure 4.
Nrf1 conditional deletion by Pcp2-Cre causes rapid degeneration of type 2 and 6 cone bipolar cells. (A) Schematic illustration of Nrf1flox, Nrf1del, and Pcp2-Cre alleles. Exons are indicated as E1–E12. Black arrows indicate the translational start sites for Nrf1 and Cre protein. Red boxes indicate loxP sites. (B,C) Immunostaining of 4-week-old Nrf1f/+;Pcp2-Cre (B,B′) and Nrf1f/f;Pcp2-Cre (C,C′) retinas with anti-GFP (B,C) and anti-Nrf1 (B,B′,C,C′) antibodies. GFP+ cells are indicated with arrowhead. (D–G) Immunostaining of 4- and 12-week-old Nrf1f/+;Pcp2-Cre (D,F) and Nrf1f/f;Pcp2-Cre (E,G) retinas with anti-GFP and anti-Nrf1 antibodies. GFP+ cells are indicated with arrowheads. (H) Number of GFP+ cells of Nrf1f/+;Pcp2-Cre and Nrf1f/f;Pcp2-Cre retinas at 4, 6, 8, and 12 weeks old. (I,J) Immunostaining of Nrf1f/+;Pcp2-Cre (I,I′,I″) and Nrf1f/f;Pcp2-Cre (J,J′,J″) retinas at 4 weeks old with anti-GFP (I–I″,J–J″), anti-PKCα (I,I′,J,J′), and anti-Syt2 (I,I″,J,J″) antibodies. GFP+PKCα+ cells, GFP+Syt2+ cells, and GFP+PKCα−Syt2− cells are indicated with white arrowheads, yellow arrowheads, and white double-arrowheads, respectively. (K) Number of GFP+PKCα+ cells of Nrf1f/+;Pcp2-Cre and Nrf1f/f;Pcp2-Cre retinas at 4, 6, 8, and 12 weeks old. (L) Number of GFP+Syt2+ cells of Nrf1f/+;Pcp2-Cre and Nrf1f/f;Pcp2-Cre retinas at 4, 6, 8, and 12 weeks old. Scale bars: 20 μm in (B,B′,C,C′,I,I′,I″,J,J′,J″) and 50 μm in (D–G).

Figure 5.
Nrf1 conditional deletion by RhoiCre causes dynamic gene expression alteration before the onset of photoreceptor degeneration. (A) Schematic representation of Nrf1flox allele, Nrf1 deleted allele, and RhoiCre allele. Exons are indicated as E1–E12. Black arrows indicate the translational start sites for Nrf1 and iCre protein. Red boxes indicate loxP sites. (B) Volcano plot showing the differentially expressed genes. The x-axis indicates the log2 fold change of genes between mutant and control. The y-axis indicates the −log10(adjusted p-value). Each dot represents a gene. Blue dots indicate no significant difference; red dots indicate upregulated DEGs; green dots indicate downregulated DEGs. (C) Venn diagram showing coexpressed genes among the three gene lists described in the text. (D) Hierarchical clustering heatmap showing the 86 most sensitive, early responsive DEGs in rod-specific Nrf1-deleted retinas. (E) Schematic mapping of mitochondrion and functional annotation of upregulated (red) and downregulated (green) genes in Nrf1f/f;RhoiCre retinas detected by RNA-seq. (F) qRT-PCR analysis of wild-type and Nrf1f/f;RhoiCre retinas, confirming the changes of mRNA levels detected by RNA-seq. (G–J) TUNEL assay of 3- and 7-week-old Nrf1f/+ (G,I) and Nrf1f/f;RhoiCre (H,J) retinas. ONL: outer nuclear layer. INL: inner nuclear layer. Scale bars: 50 μm.
Figure 5.
Nrf1 conditional deletion by RhoiCre causes dynamic gene expression alteration before the onset of photoreceptor degeneration. (A) Schematic representation of Nrf1flox allele, Nrf1 deleted allele, and RhoiCre allele. Exons are indicated as E1–E12. Black arrows indicate the translational start sites for Nrf1 and iCre protein. Red boxes indicate loxP sites. (B) Volcano plot showing the differentially expressed genes. The x-axis indicates the log2 fold change of genes between mutant and control. The y-axis indicates the −log10(adjusted p-value). Each dot represents a gene. Blue dots indicate no significant difference; red dots indicate upregulated DEGs; green dots indicate downregulated DEGs. (C) Venn diagram showing coexpressed genes among the three gene lists described in the text. (D) Hierarchical clustering heatmap showing the 86 most sensitive, early responsive DEGs in rod-specific Nrf1-deleted retinas. (E) Schematic mapping of mitochondrion and functional annotation of upregulated (red) and downregulated (green) genes in Nrf1f/f;RhoiCre retinas detected by RNA-seq. (F) qRT-PCR analysis of wild-type and Nrf1f/f;RhoiCre retinas, confirming the changes of mRNA levels detected by RNA-seq. (G–J) TUNEL assay of 3- and 7-week-old Nrf1f/+ (G,I) and Nrf1f/f;RhoiCre (H,J) retinas. ONL: outer nuclear layer. INL: inner nuclear layer. Scale bars: 50 μm.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
The most sensitive genes in Nrf1f/f;RhoiCre retina. Gene names in bold font are direct targets of Nrf1 found in P0 brain (GEO dataset: GSE161808). Genes marked with an asterisk encode proteins found in mitochondria.
Table 1.
The most sensitive genes in Nrf1f/f;RhoiCre retina. Gene names in bold font are direct targets of Nrf1 found in P0 brain (GEO dataset: GSE161808). Genes marked with an asterisk encode proteins found in mitochondria.
| Gene Name | Description | Log2 Fold Change (Nrf1f/f;RhoiCre/Nrf1f/+) | Adjusted p Value |
|---|---|---|---|
| Ccdc87 | coiled-coil domain containing 87 | −1.6234 | 5.89 × 10−36 |
| Rnf207 | ring finger protein 207 | −1.3834 | 1.10 × 10−44 |
| AI429214 | expressed sequence AI429214 | −1.3369 | 6.84 × 10−24 |
| C030017K20Rik | RIKEN cDNA C030017K20 gene | −1.2807 | 1.19 × 10−31 |
| Zfp182 | zinc finger protein 182 | −1.2239 | 4.54 × 10−40 |
| Tbc1d24 | TBC1 domain family member 24 | −1.2234 | 9.22 × 10−120 |
| Cdc7 | cell division cycle 7 (S. cerevisiae) | −1.2132 | 4.45 × 10−43 |
| Samhd1 | SAM domain and HD domain 1 | −1.139 | 2.41 × 10−82 |
| Zc2hc1c | zinc finger C2HC-type containing 1C | −1.1341 | 4.26 × 10−23 |
| Wdr5b | WD repeat domain 5B | −1.1292 | 8.21 × 10−26 |
| Sprtn | SprT-like N-terminal domain | −1.1122 | 6.12 × 10−39 |
| H2-K2 | histocompatibility 2 K region locus 2 | −1.0906 | 1.58 × 10−22 |
| Shisa2 | shisa family member 2 | −1.0698 | 2.44 × 10−25 |
| Mlh3 | mutL homolog 3 (E. coli) | −1.0473 | 1.15 × 10−44 |
| Ankrd26 * | ankyrin repeat domain 26 | −1.0158 | 1.68 × 10−55 |
| Cog6 | component of oligomeric golgi complex 6 | −1.0071 | 4.78 × 10−49 |
| Zfp287 | zinc finger protein 287 | −0.99342 | 7.33 × 10−27 |
| Mtg2 * | mitochondrial ribosome associated GTPase 2 | −0.97742 | 5.34 × 10−33 |
| Katnal2 | katanin p60 subunit A-like 2 | −0.97416 | 3.88 × 10−34 |
| Mthfsd | methenyltetrahydrofolate synthetase domain containing | −0.97072 | 8.13 × 10−31 |
| Zfp791 | zinc finger protein 791 | −0.96542 | 3.25 × 10−19 |
| Zfp329 | zinc finger protein 329 | −0.9606 | 4.01 × 10−54 |
| Pot1b | protection of telomeres 1B | −0.95894 | 9.62 × 10−21 |
| Bbs2 | Bardet-Biedl syndrome 2 (human) | −0.95414 | 1.22 × 10−63 |
| Dynlrb2 | dynein light chain roadblock-type 2 | −0.94971 | 6.89 × 10−14 |
| Abhd5 | abhydrolase domain containing 5 | −0.9241 | 1.01 × 10−26 |
| Marveld3 | MARVEL (membrane-associating) domain containing 3 | −0.91459 | 6.08 × 10−15 |
| Ccdc166 | coiled-coil domain containing 166 | −0.90586 | 1.81 × 10−31 |
| Exoc8 | exocyst complex component 8 | −0.90494 | 6.33 × 10−36 |
| Stk38 | serine/threonine kinase 38 | −0.89765 | 2.81 × 10−60 |
| Tctex1d2 | Tctex1 domain containing 2 | −0.89357 | 9.03 × 10−14 |
| Gm26572 | predicted gene 26572 | −0.88514 | 1.06 × 10−16 |
| Rabif | RAB interacting factor | −0.87374 | 6.53 × 10−43 |
| Dynlt1c | dynein light chain Tctex-type 1C | −0.87284 | 3.64 × 10−11 |
| Spint2 | serine protease inhibitor Kunitz type 2 | −0.87063 | 2.51 × 10−43 |
| Fam98a | family with sequence similarity 98 member A | −0.86343 | 2.20 × 10−35 |
| Slx4 | SLX4 structure-specific endonuclease subunit homolog (S. cerevisiae) | −0.85413 | 9.39 × 10−34 |
| Mtrr | 5-methyltetrahydrofolate-homocysteine methyltransferase reductase | −0.84843 | 1.96 × 10−18 |
| Setd4 | SET domain containing 4 | −0.83398 | 1.40 × 10−17 |
| Gcfc2 | GC-rich sequence DNA binding factor 2 | −0.83387 | 2.49 × 10−15 |
| Sox12 | SRY (sex determining region Y)-box 12 | −0.82933 | 7.00 × 10−21 |
| Sbf2 | SET binding factor 2 | −0.82821 | 2.80 × 10−50 |
| Nubp1 | nucleotide binding protein 1 | −0.82325 | 3.47 × 10−19 |
| Angptl6 | angiopoietin-like 6 | −0.82273 | 9.67 × 10−13 |
| Mrs2 * | MRS2 magnesium homeostasis factor homolog (S. cerevisiae) | −0.81398 | 6.57 × 10−35 |
| Pdrg1 | p53 and DNA damage regulated 1 | −0.79343 | 1.45 × 10−27 |
| Zfyve16 | zinc finger FYVE domain containing 16 | −0.78442 | 1.04 × 10−24 |
| Ints7 | integrator complex subunit 7 | −0.783 | 5.35 × 10−33 |
| Shmt2 * | serine hydroxymethyltransferase 2 (mitochondrial) | −0.78075 | 2.29 × 10−29 |
| Mettl22 | methyltransferase like 22 | −0.77958 | 2.18 × 10−16 |
| Ccdc93 | coiled-coil domain containing 93 | −0.77053 | 2.11 × 10−27 |
| Smg9 | smg-9 homolog nonsense mediated mRNA decay factor (C. elegans) | −0.76729 | 3.83 × 10−24 |
| Tmem143 * | transmembrane protein 143 | −0.75985 | 4.64 × 10−20 |
| Prkdc | protein kinase DNA activated catalytic polypeptide | −0.75349 | 1.45 × 10−18 |
| 2310061I04Rik * | RIKEN cDNA 2310061I04 gene | −0.75092 | 6.93 × 10−17 |
| Pls1 | plastin 1 (I-isoform) | −0.74708 | 2.22 × 10−10 |
| Nrtn | neurturin | −0.74655 | 2.40 × 10−12 |
| Nr2c1 | nuclear receptor subfamily 2 group C member 1 | −0.74308 | 2.67 × 10−19 |
| Hist1h1a | histone cluster 1 H1a | 0.74952 | 7.45 × 10−8 |
| Gm13421 | predicted gene 13421 | 0.77879 | 2.71 × 10−14 |
| Hist1h4c | histone cluster 1 H4c | 0.78665 | 1.79 × 10−8 |
| Bpifb6 | BPI fold containing family B member 6 | 0.79491 | 2.04 × 10−8 |
| Hist1h2be | histone cluster 1 H2be | 0.7962 | 2.31 × 10−14 |
| Nhej1 | nonhomologous end-joining factor 1 | 0.80335 | 1.01 × 10−8 |
| BC051142 | cDNA sequence BC051142 | 0.81214 | 1.83 ×10−10 |
| Hist1h2bj | histone cluster 1 H2bj | 0.83377 | 3.03 ×10−9 |
| Ska3 | spindle and kinetochore associated complex subunit 3 | 0.84721 | 1.54 ×10−9 |
| Gm11961 | predicted gene 11961 | 0.87529 | 2.79 × 10−15 |
| Hist1h1e | histone cluster 1 H1e | 0.93176 | 2.49 × 10−15 |
| Gm9959 | predicted gene 9959 | 0.93908 | 1.03 × 10−11 |
| Rbbp8 | retinoblastoma binding protein 8 | 0.97505 | 3.74 × 10−21 |
| Mlf1 | myeloid leukemia factor 1 | 0.98101 | 3.09 × 10−15 |
| Bcl2l12 | BCL2-like 12 (proline rich) | 0.98105 | 9.18 × 10−13 |
| Gm17101 | predicted gene 17101 | 0.98366 | 7.78 × 10−13 |
| Gm12480 | predicted gene 12480 | 0.98839 | 5.94 × 10−13 |
| Ino80c | INO80 complex subunit C | 0.99506 | 8.83 × 10−55 |
| Gm8251 | predicted gene 8251 | 1.1522 | 1.03 × 10−18 |
| Dsn1 | DSN1 MIND kinetochore complex component homolog (S. cerevisiae) | 1.1532 | 1.85 × 10−18 |
| Loxl4 | lysyl oxidase-like 4 | 1.2333 | 3.80 × 10−21 |
| Mutyh * | mutY homolog (E. coli) | 1.2731 | 3.28 × 10−38 |
| Zcwpw1 | zinc finger CW type with PWWP domain 1 | 1.4136 | 1.66 × 10−67 |
| Rad51ap1 | RAD51 associated protein 1 | 1.5557 | 1.07 × 10−41 |
| Cenpi | centromere protein I | 1.8589 | 4.28 × 10−52 |
| Fignl1 | fidgetin-like 1 | 2.0243 | 2.37 × 10−73 |
| Casc5 | cancer susceptibility candidate 5 | 2.7592 | 6.88 × 10−109 |
Table 2.
Mitochondrial genes with modest changed expression levels in Nrf1f/f;RhoiCre retina. Gene names in bold font are direct targets of Nrf1 found in P0 brain (GEO dataset: GSE161808).
Table 2.
Mitochondrial genes with modest changed expression levels in Nrf1f/f;RhoiCre retina. Gene names in bold font are direct targets of Nrf1 found in P0 brain (GEO dataset: GSE161808).
| Gene Name | Description | Log2 Fold change (Nrf1f/f;RhoiCre/Nrf1f/+) | Adjusted p Value |
|---|---|---|---|
| Ankrd26 | ankyrin repeat domain-containing protein 26 | −1.0158 | 1.68 × 10−55 |
| Mtg2 | mitochondrial ribosome associated GTPase 2 | −0.97742 | 5.34 × 10−33 |
| Mrs2 | MRS2 magnesium homeostasis factor homolog (S. cerevisiae) | −0.81398 | 6.57 × 10−35 |
| Shmt2 | serine hydroxymethyltransferase 2 (mitochondrial) | −0.78075 | 2.29 × 10−29 |
| Tmem143 | transmembrane protein 143 | −0.75985 | 4.64 × 10−20 |
| 2310061I04Rik | RIKEN cDNA 2310061I04 gene | −0.75092 | 4.12 × 10−19 |
| Fastkd3 | FAST kinase domains 3 | −0.73257 | 6.15 × 10−12 |
| Pstk | phosphoseryl tRNA kinase | −0.73031 | 1.28 × 10−16 |
| Alas1 | aminolevulinic acid synthase 1 | −0.70115 | 9.42 × 10−25 |
| Hsdl2 | hydroxysteroid dehydrogenase like 2 | −0.59599 | 1.75 × 10−20 |
| D2hgdh | D2 hydroxyglutarate dehydrogenase | −0.58842 | 1.56 × 10−11 |
| Sdr39u1 | short chain dehydrogenase/reductase family 39U member 1 | −0.55177 | 1.20 × 10−14 |
| Sdha | succinate dehydrogenase complex subunit A flavoprotein (Fp) | −0.54497 | 5.27 × 10−32 |
| Cecr5 | cat eye syndrome chromosome region candidate 5 | −0.53991 | 1.65 × 10−11 |
| Slc25a13 | solute carrier family 25 (mitochondrial carrier adenine nucleotide translocator) member 13 | −0.53462 | 4.91 × 10−4 |
| Acad10 | acyl Coenzyme A dehydrogenase family member 10 | −0.53414 | 2.61 × 10−6 |
| Pccb | propionyl Coenzyme A carboxylase beta polypeptide | −0.52005 | 2.09 ×10−15 |
| Exog | endo/exonuclease (5′-3′) endonuclease G-like | −0.51228 | 1.76 × 10−9 |
| Ide | insulin degrading enzyme | −0.50673 | 1.97 × 10−22 |
| Mcat | malonyl CoA:ACP acyltransferase (mitochondrial) | −0.48803 | 1.60 × 10−7 |
| Afg3l1 | AFG3-like AAA ATPase 1 | −0.48699 | 6.23 × 10−16 |
| 2810006K23Rik | RIKEN cDNA 2810006K23 gene | −0.47985 | 2.65 × 10−7 |
| Vars2 | valyl-tRNA synthetase 2 mitochondrial (putative) | −0.45605 | 9.55 × 10−7 |
| Mtpap | mitochondrial poly(A)_polymerase | −0.45337 | 1.37 × 10−5 |
| Dna2 | DNA replication helicase 2 homolog (yeast) | −0.44603 | 0.003128 |
| Mrpl49 | mitochondrial ribosomal protein L49 | −0.43346 | 1.64 × 10−8 |
| Nfu1 | NFU1 iron sulfur cluster scaffold homolog (S. cerevisiae) | −0.42766 | 7.19 × 10−9 |
| Rpusd3 | RNA pseudouridylate synthase domain containing 3 | −0.41583 | 1.73 × 10−4 |
| Abcb10 | ATP-binding cassette subfamily B (MDR/TAP) member 10 | −0.41548 | 3.37 × 10−12 |
| Ppa2 | pyrophosphatase (inorganic) 2 | −0.40886 | 9.23 × 10−6 |
| Bphl | biphenyl hydrolase like (serine hydrolase breast epithelial mucin associated antigen) | −0.39912 | 9.78 × 10−4 |
| Dhrs7b | dehydrogenase/reductase (SDR family) member 7B | −0.39831 | 3.88 × 10−6 |
| Cisd1 | CDGSH iron sulfur domain 1 | −0.3853 | 5.47 × 10−6 |
| Stx17 | syntaxin 17 | −0.38146 | 4.69 × 10−7 |
| Gpi1 | glucose phosphate isomerase 1 | −0.35742 | 2.27 × 10−6 |
| Ndufa8 | NADH dehydrogenase (ubiquinone) 1 alpha subcomplex 8 | −0.34492 | 2.18 × 10−4 |
| Acaa1a | acetyl-Coenzyme A acyltransferase 1A | −0.33944 | 2.45 × 10−9 |
| Idh3a | isocitrate dehydrogenase 3 (NAD+) alpha | −0.32204 | 5.08 × 10−9 |
| Iba57 | IBA57 iron sulfur cluster assembly homolog (S. cerevisiae) | 0.33473 | 2.31 × 10−4 |
| Tha1 | threonine aldolase 1 | 0.34796 | 4.24 × 10−3 |
| Amacr | alpha methylacyl CoA racemase | 0.36987 | 1.82 × 10−3 |
| Aldh1l2 | aldehyde dehydrogenase 1 family member L2 | 0.37356 | 3.79 × 10−3 |
| Tfb1m | transcription factor B1 mitochondrial | 0.37658 | 4.31 × 10−3 |
| Mrpl54 | mitochondrial ribosomal protein L54 | 0.38391 | 2.29 × 10−4 |
| Mrpl36 | mitochondrial ribosomal protein L36 | 0.41616 | 9.30 × 10−5 |
| Nucb2 | nucleobindin 2 | 0.48404 | 4.16 × 10−16 |
| Sars2 | seryl-aminoacyl-tRNA synthetase 2 | 0.54314 | 2.87 × 10−10 |
| Ephx2 | epoxide hydrolase 2 cytoplasmic | 0.56004 | 1.71 × 10−14 |
| Ccdc51 | coiled-coil domain containing 51 | 0.57846 | 1.49 × 10−7 |
| Ccdc127 | coiled-coil domain containing 127 | 0.58535 | 1.02 × 10−22 |
| Rpl10a | ribosomal protein L10A | 0.64403 | 1.49 × 10−42 |
| Mutyh | mutY homolog (E. coli) | 1.2731 | 3.28 × 10−38 |
Table 3.
Top four GO terms from analysis of the 86 most sensitive genes in Nrf1f/f;RhoiCre retina.
| GO Biological Process ID | Number of Focused Genes | Fold Enrichment | Raw p-Value | FDR | Gene Name |
|---|---|---|---|---|---|
| regulation of double-strand break repair (GO:2000779) | 6 | 18.07 | 1.35 × 10−6 | 3.05 × 10−3 | Pot1b, Rad51ap1, Fignl1, Rbbp8, Zcwpw1, Prkdc |
| double-strand break repair via homologous recombination (GO:0000724) | 5 | 16.42 | 1.69 × 10−5 | 1.91 × 10−2 | Cdc7, Slx4, Samhd1, Rad51ap1, Rbbp8 |
| regulation of DNA recombination (GO:0000018) | 6 | 15.04 | 3.75 × 10−6 | 6.57 × 10−3 | Pot1b, Rad51ap1, Fignl1, Rbbp8, Zcwpw1, Hist1h1a |
| positive regulation of DNA repair (GO:0045739) | 5 | 14.79 | 2.74 × 10−5 | 2.70 × 10−2 | Rad51ap1, Rbbp8, Zcwpw1, Ino80c, Prkdc |
Table 4.
Genes involved in glycolysis pathway with altered expression in Nrf1f/f;RhoiCre retina.
| Gene Name | Description | Fold Change (Nrf1f/f;RhoiCre/Nrf1f/+) | Adjusted p Value |
|---|---|---|---|
| Hk1 | Hexokinase 1 | 1.24 | 5.35 × 10−10 |
| Gpi1 | Glucose-6-phsphate isomerase 1 | 0.78 | 2.27 × 10−6 |
| Pgam1 | Phosphoglycerate mutase 1 | 0.81 | 1.57 × 10−4 |
| Pgm2 | Phosphoglucomutase-2 | 0.80 | 1.34 × 10−3 |
| Eno1 | Enolase 1, alpha non-neuron | 0.82 | 5.76 × 10−3 |
| Pkm | Pyruvate kinase, muscle | 0.80 | 5.76 × 10−6 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Kiyama, T.; Chen, C.-K.; Zhang, A.; Mao, C.-A. Differential Susceptibility of Retinal Neurons to the Loss of Mitochondrial Biogenesis Factor Nrf1. Cells 2022, 11, 2203. https://doi.org/10.3390/cells11142203
AMA Style
Kiyama T, Chen C-K, Zhang A, Mao C-A. Differential Susceptibility of Retinal Neurons to the Loss of Mitochondrial Biogenesis Factor Nrf1. Cells. 2022; 11(14):2203. https://doi.org/10.3390/cells11142203
Chicago/Turabian StyleKiyama, Takae, Ching-Kang Chen, Annie Zhang, and Chai-An Mao. 2022. "Differential Susceptibility of Retinal Neurons to the Loss of Mitochondrial Biogenesis Factor Nrf1" Cells 11, no. 14: 2203. https://doi.org/10.3390/cells11142203
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.