
Deletion of Cd44 Inhibits Metastasis Formation of Liver Cancer in Nf2-Mutant Mice

 , , , , , , and
, , , , , , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experiments Involving Animals
2.2. Detection of α-Fetoprotein (AFP), Alanine Aminotransferase (ALT), and Bilirubin in Mouse Serum
2.3. Histochemistry and Immunohistochemistry
2.4. TUNEL Assay
2.5. Isolation of Primary Liver Cells
2.6. Sources of Other Cells and Cell Culture Conditions
2.7. Clonogenic Assay under Cell Attachment Conditions
2.8. Colony Growth under Nonadherent Conditions/Anoikis Assay
2.9. Scratch Migration Assay
2.10. Tumor Cell Transendothelial Migration Assay
2.11. Western Blot
2.12. Immunocytochemistry
2.13. Antibodies
2.14. RNA Sequencing and Gene Expression Analysis
2.15. Real-Time Quantitative Reverse Transcription PCR (Real-Time qRT-PCR)
2.16. Small Interfering RNA (siRNA) Transfections
2.17. Statistical Analysis
3. Results
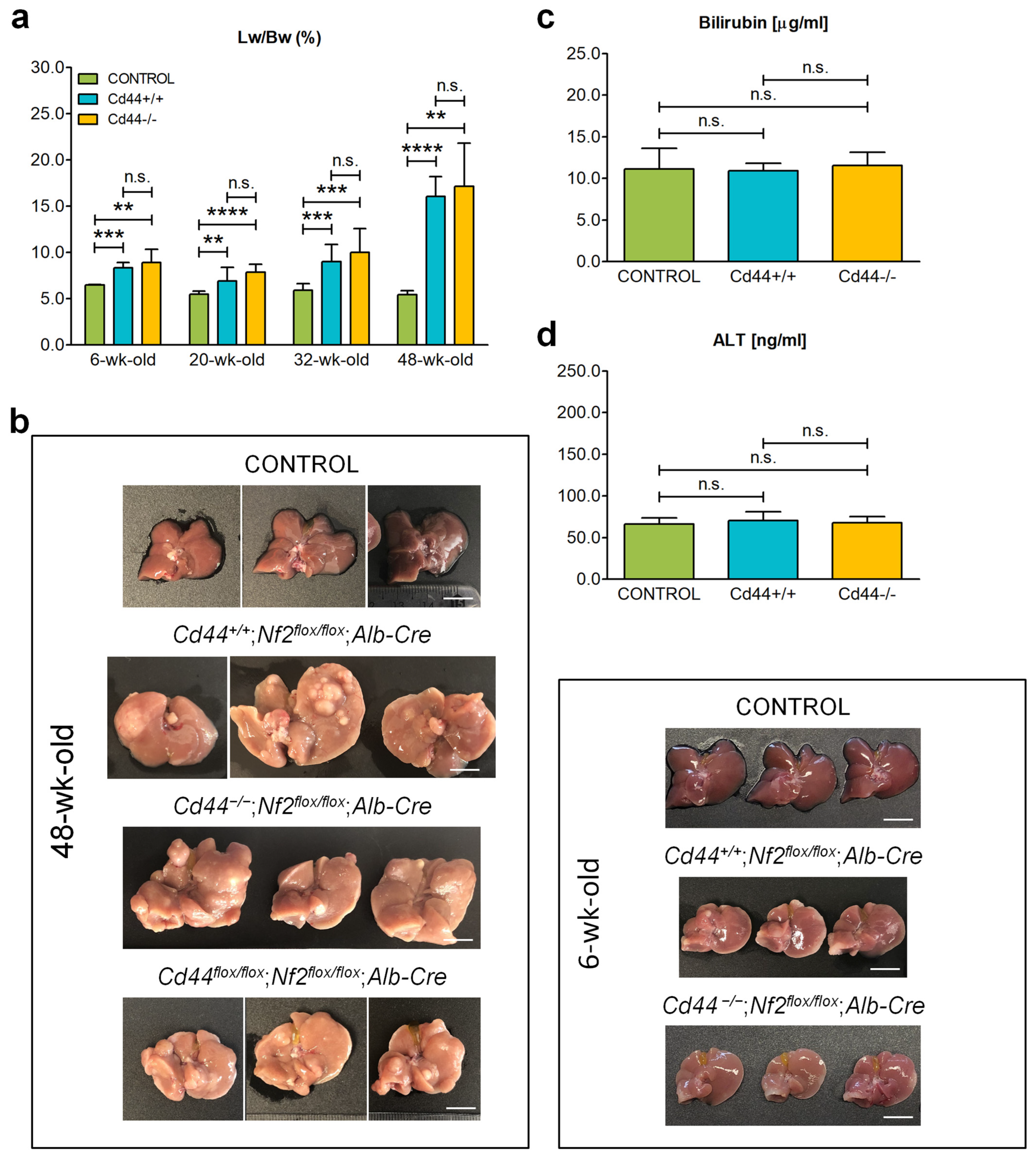
3.1. Deletion of Cd44 Does Not Influence Progressive Liver Enlargement in Nf2flox/flox;Alb-Cre Mice
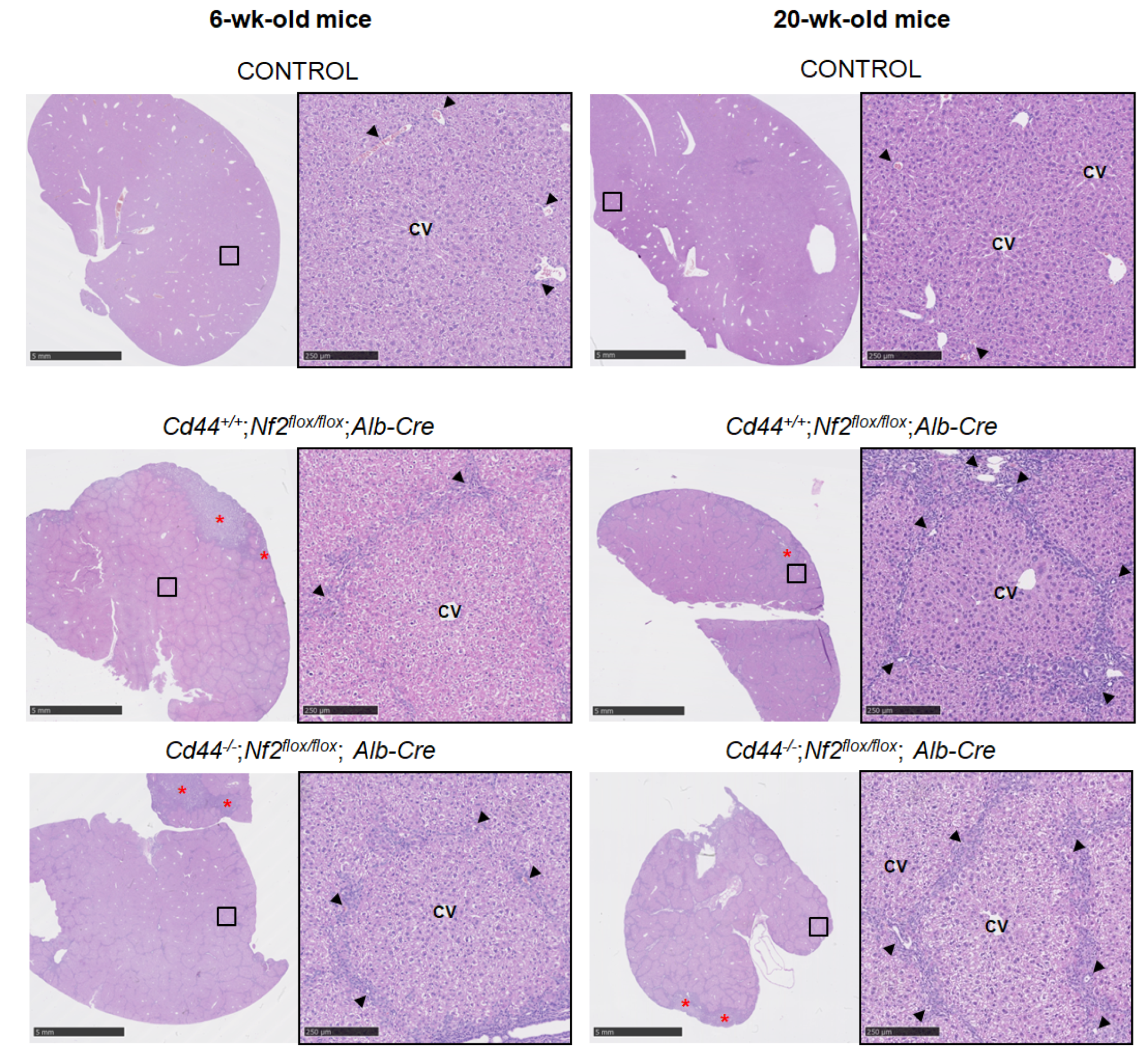
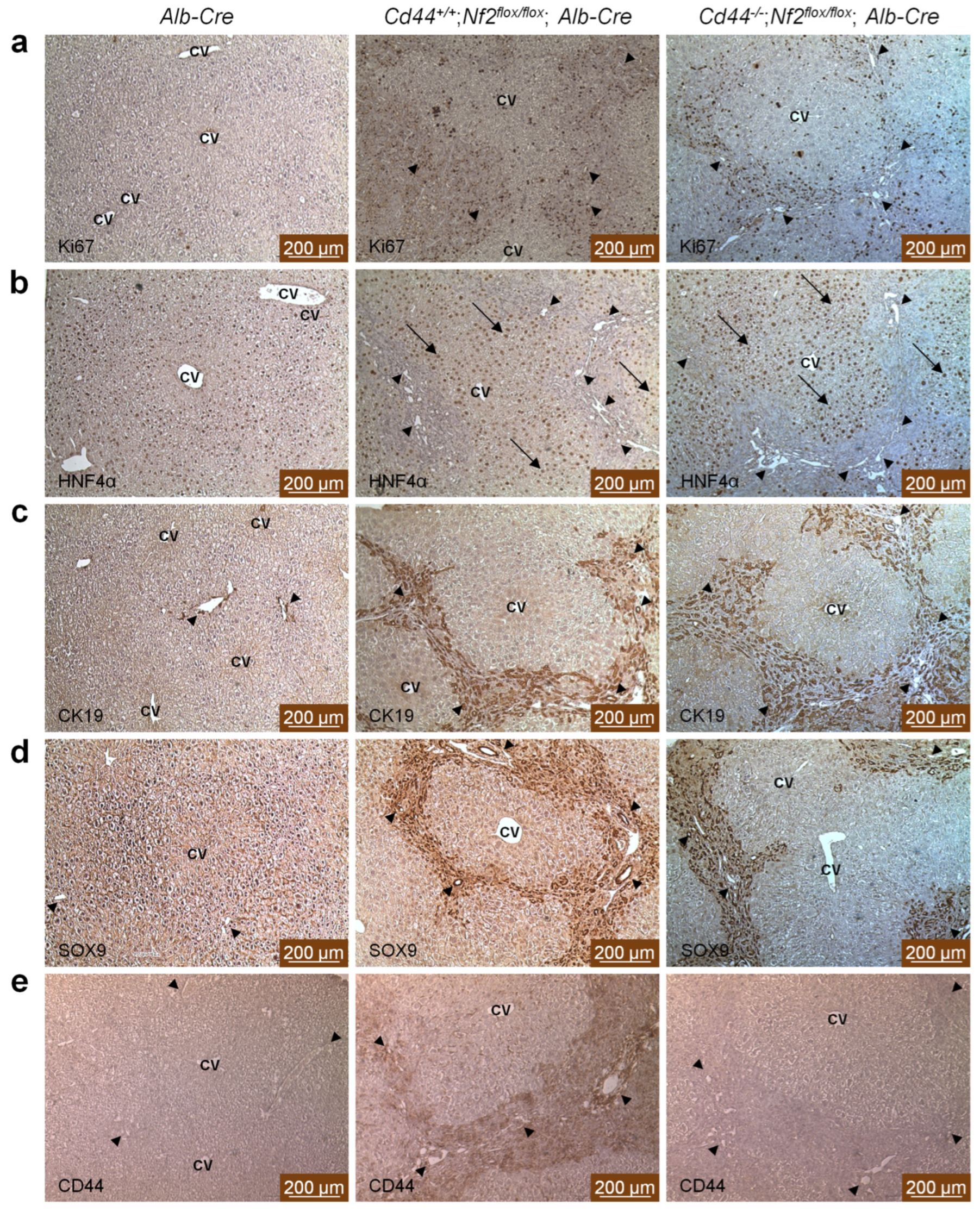
3.2. Ductular Proliferation Observed in Nf2flox/flox;Alb-Cre Mice Is Not Influenced by Loss of Cd44
3.3. CD44 Exhibits Similar Expression Pattern to Ductular/Liver Progenitor Cell Markers, but Is Not Required for Ductular Process in Nf2flox/flox;Alb-Cre Mice
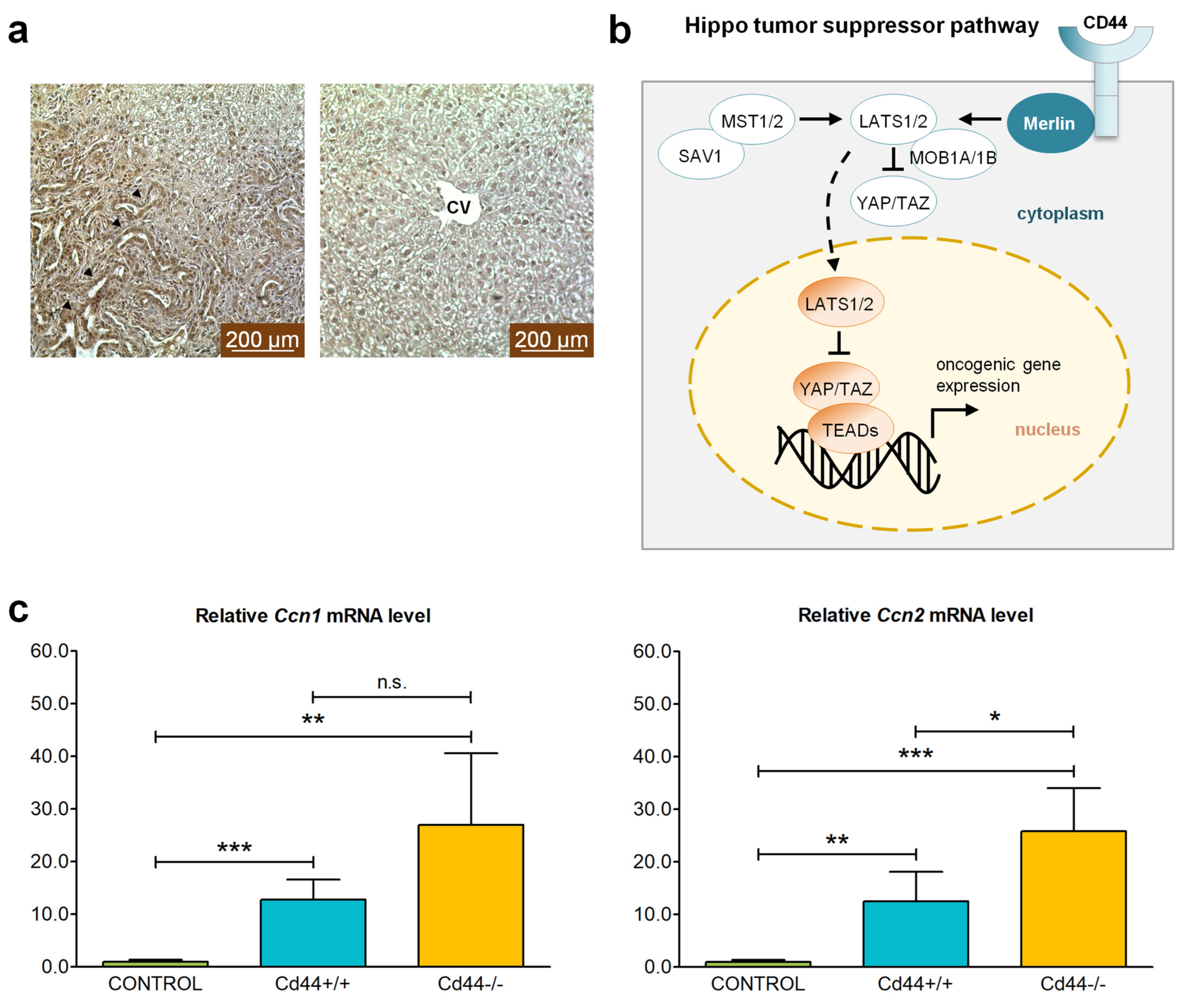
3.4. YAP Signaling Is Elevated in Nf2-Deficient Livers
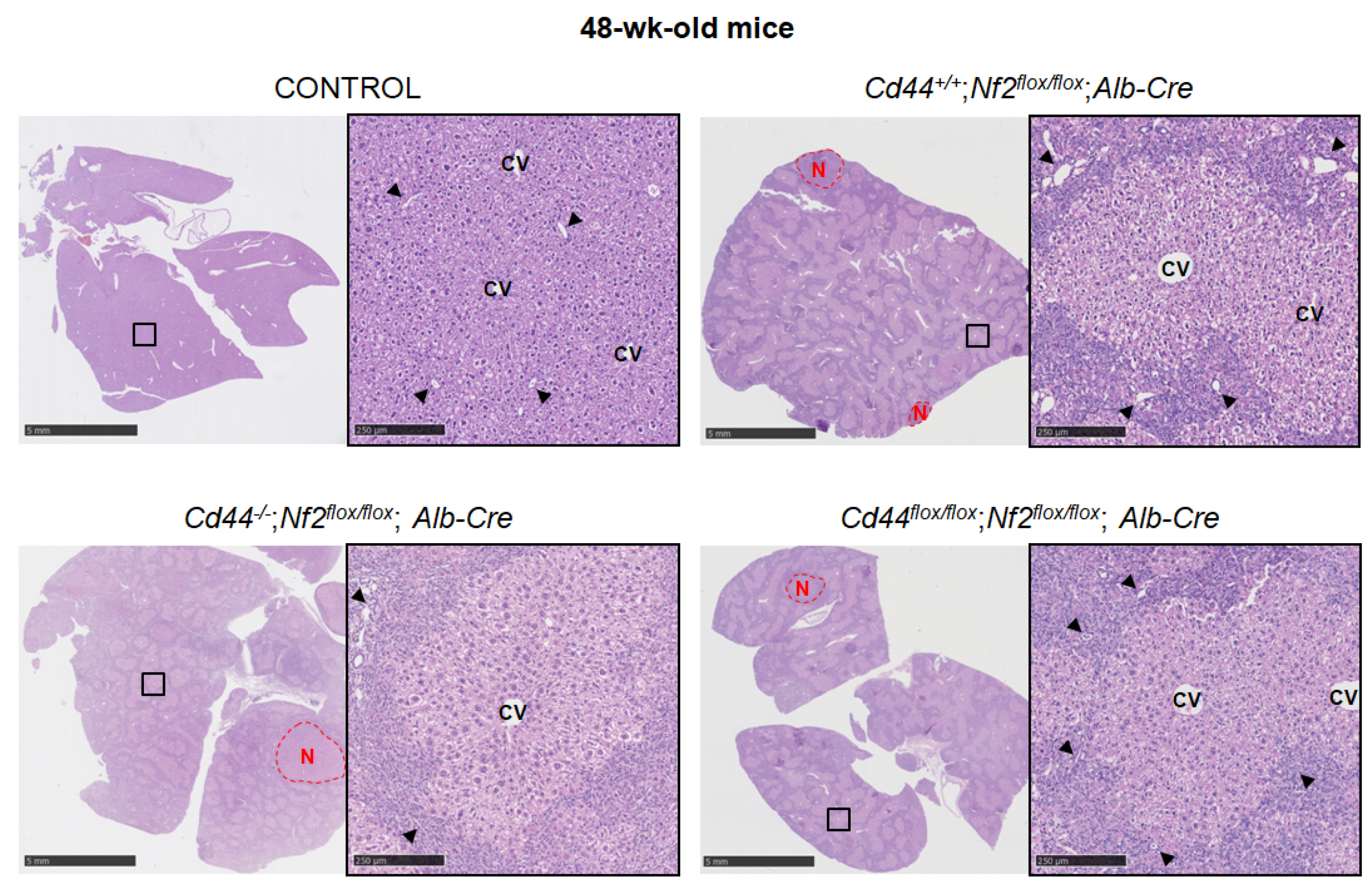
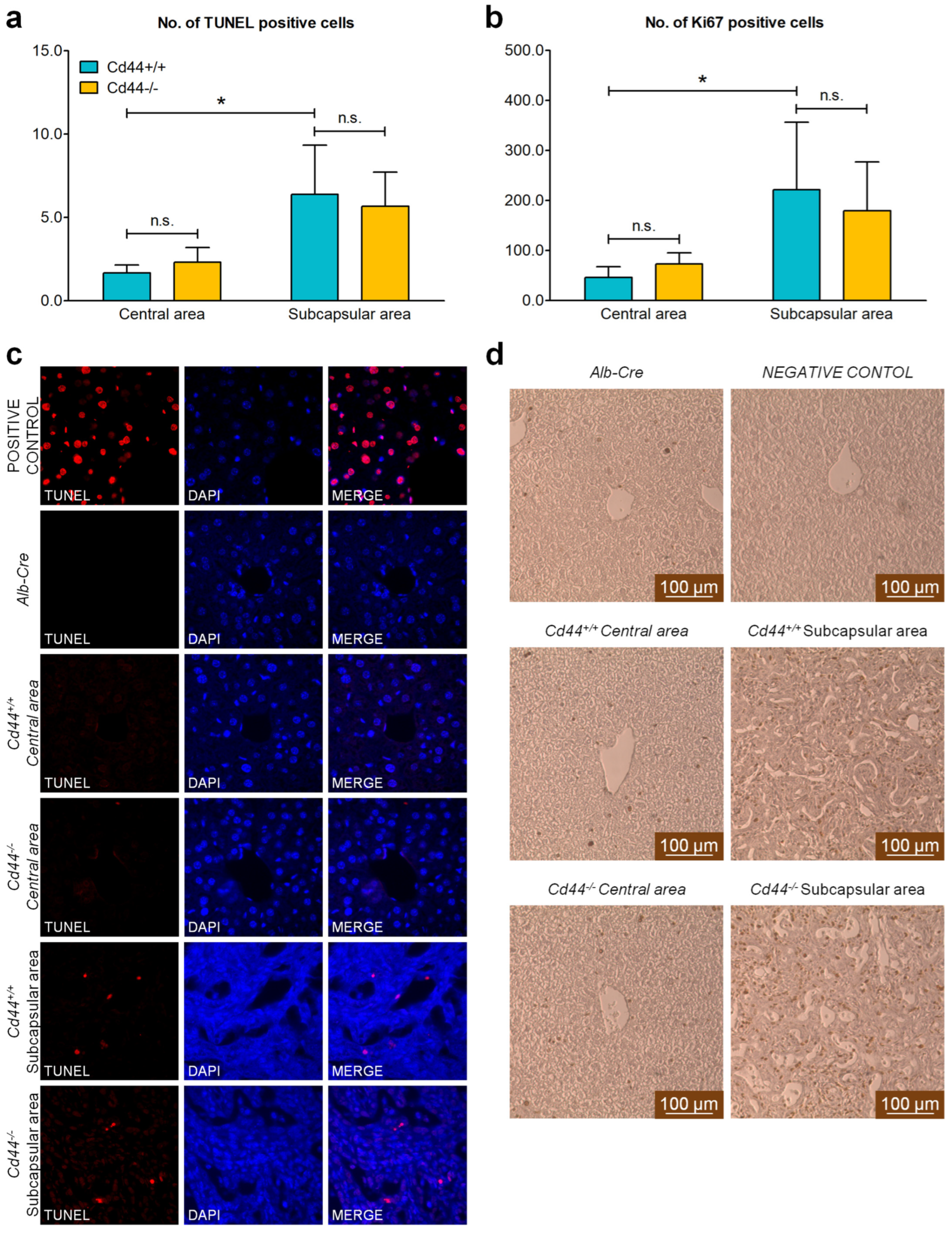
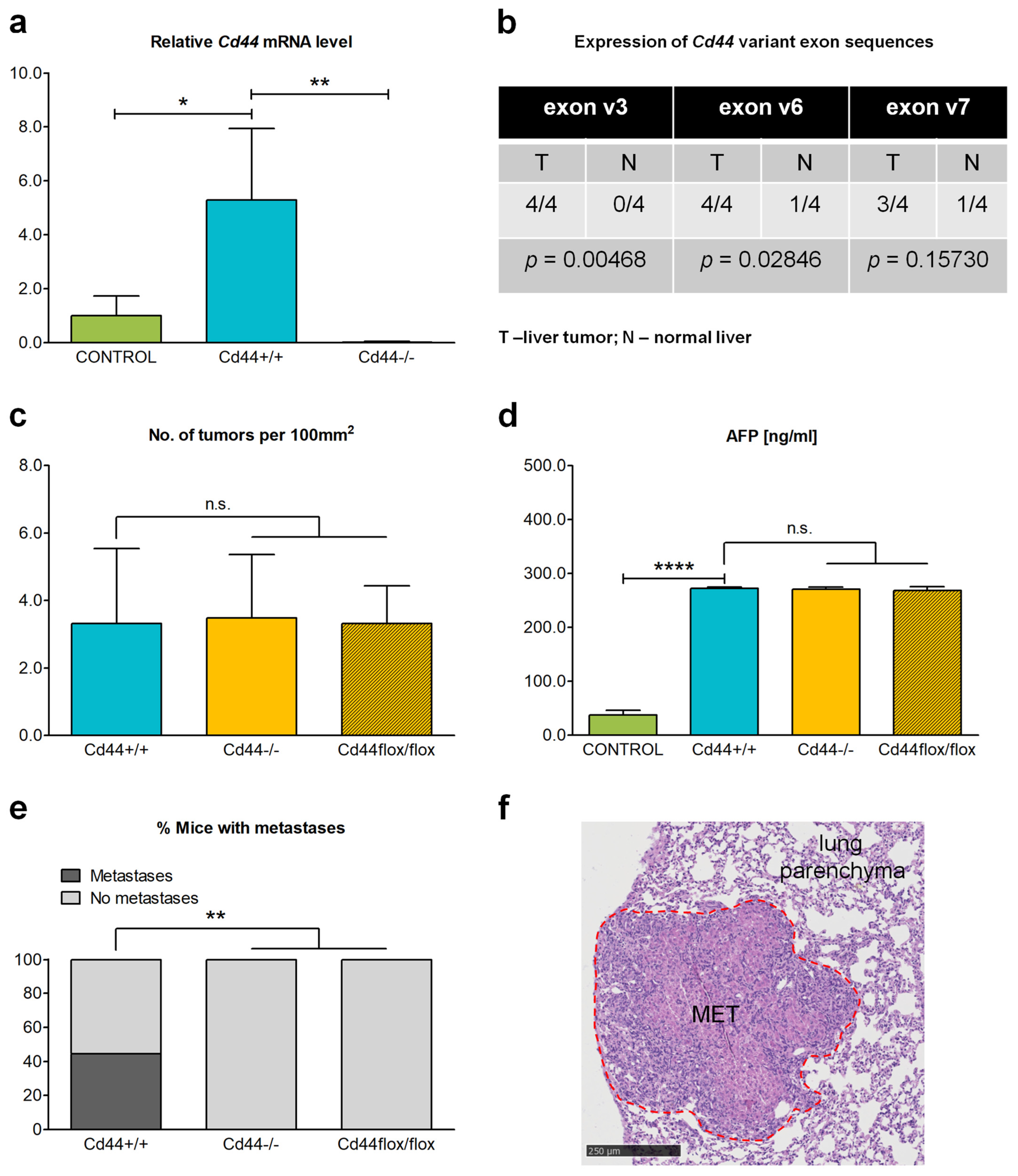
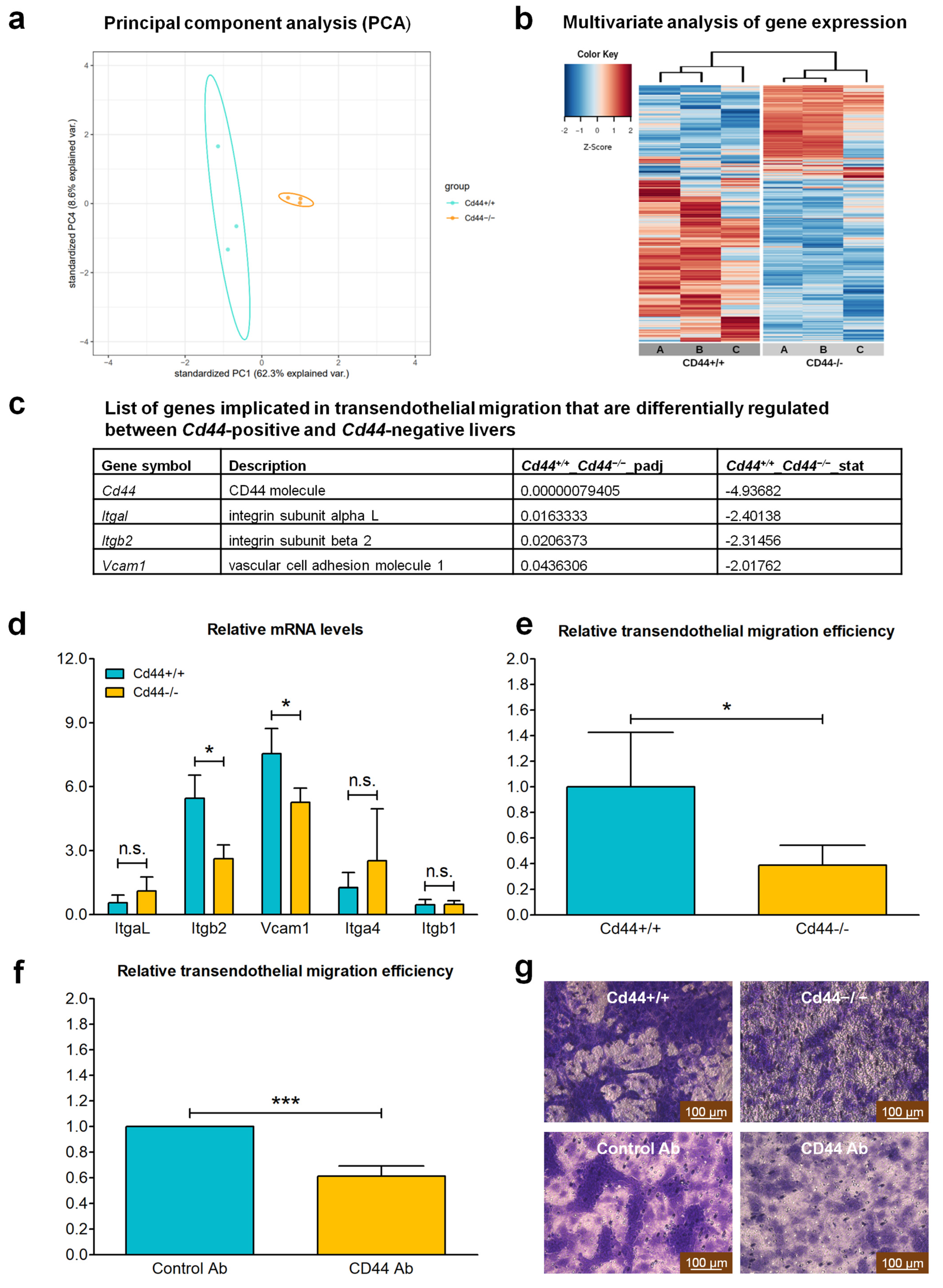
3.5. Deletion of Cd44 Does Not Influence Primary Liver Tumor Generation but Virtually Aborts Metastasis Formation in Nf2flox/flox;Alb-Cre Mice
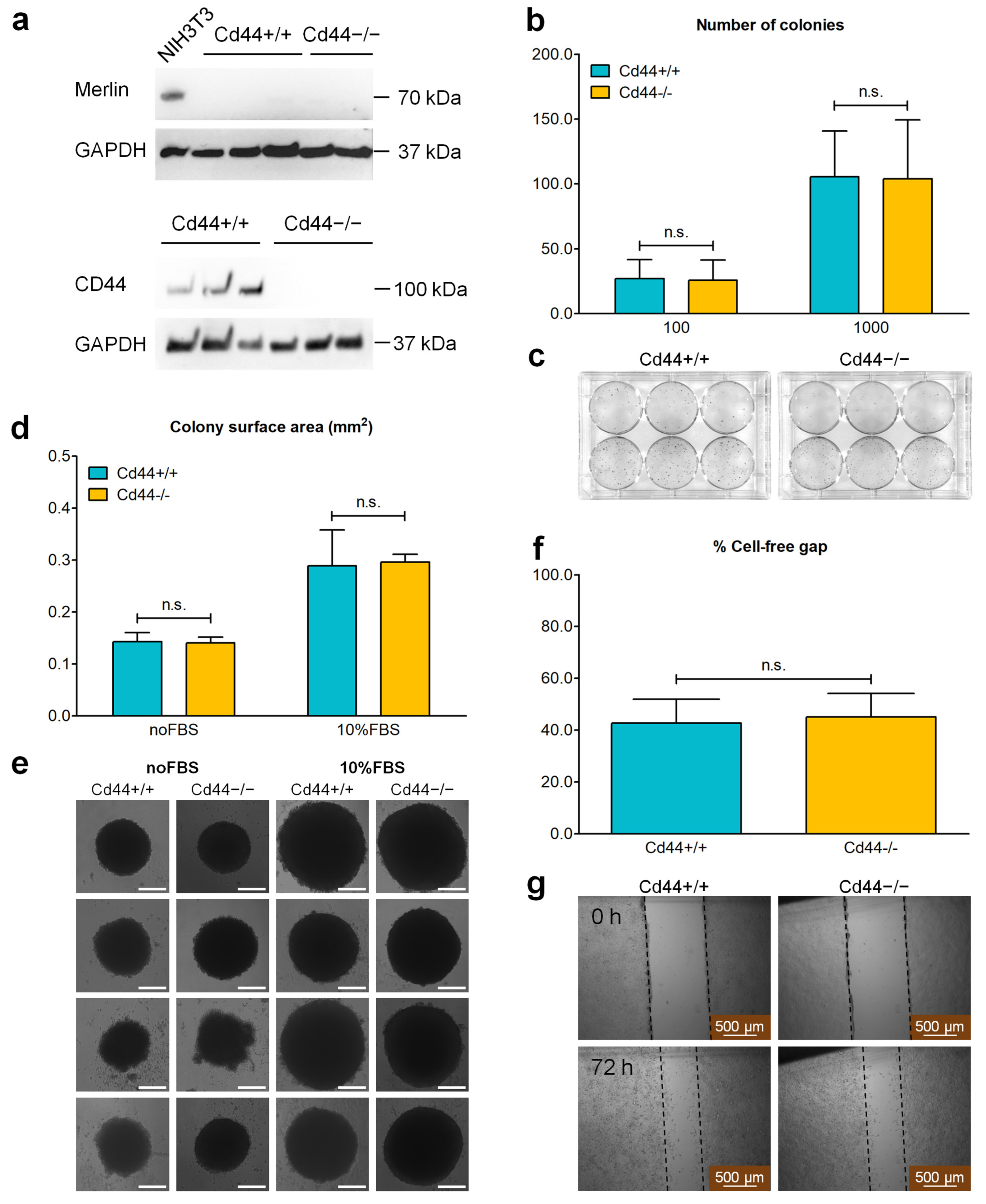
3.6. Deletion of the Cd44 Gene Does Not Influence Colony-Forming Ability or Migration into Cell-Free Gap of Nf2-Negative Liver Cells
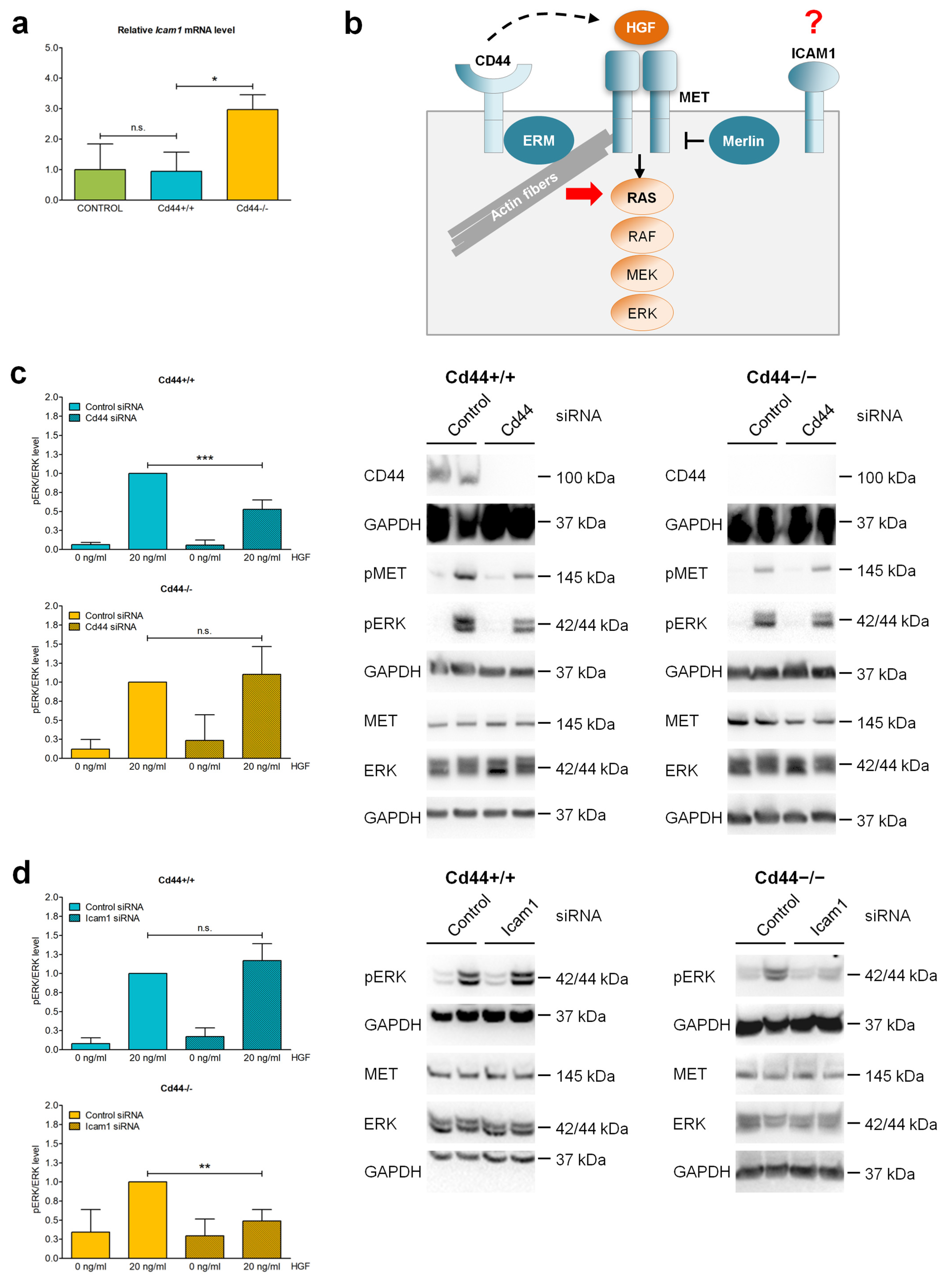
3.7. CD44 Contributes to Transendothelial Migration of Nf2-Negative Liver Cells by Upregulating Levels of Integrin Subunit Beta 2
3.8. ICAM-1 May Overtake CD44 Function as a Coreceptor of Receptor Tyrosine Kinase MET in Cd44-Negative Liver Cells
4. Discussion
4.1. Deletion of Nf2 in the Liver Leads to CD44-Independent Liver Enlargement
4.2. Origin of Active Cells during Ductular Reaction Observed in Nf2flox/flox;Alb-Cre Mice
4.3. CD44 Does Not Influence Primary Liver Tumor Development in Nf2flox/flox;Alb-Cre Mice
4.4. Compensatory Molecules May Substitute for CD44 Function during Primary Tumor Development in Cd44-Knockout Nf2flox/flox;Alb-Cre Mice
4.5. CD44 Facilitates Metastatic Dissemination of Nf2-Deficient Liver Tumors
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AFP | α-fetoprotein |
| AKT | AK mouse plus transforming or thymoma |
| Alb | albumin |
| Amot | angiomotin |
| AraC | cytosine b-D-arabinofuranoside |
| ARID1A | AT-rich interactive domain-containing protein 1A |
| BCA | bicinchoninic acid |
| BIRC5 | baculoviral IAP repeat containing 5 |
| BMI1 | B lymphoma Mo-MLV insertion region 1 homolog |
| BSA | bovine serum albumin |
| CD44 | cluster of differentiation 44 |
| CK19 | cytokeratin 19 |
| CPI-17 | C-kinase potentiated protein phosphatase-1 Inhibitor |
| Cre | causes recombination |
| CRL4 | Cullin-RING ubiquitin ligase 4 |
| CSC | cancer stem cells |
| CTGF | connective tissue growth factor |
| CTLA-4 | cytotoxic T-lymphocyte-associated protein 4 |
| CYR61 | cysteine-rich angiogenic inducer 61 |
| DAPI | 4’,6-diamidino-2-phenylindole |
| DCAF1 | DDB1- and Cul4-associated factor 1 |
| DMEM | Dulbecco’s Modified Eagle’s Medium |
| EGFR | epidermal growth factor receptor |
| ELISA | enzyme-linked immunosorbent assay |
| EMT | epithelial-mesenchymal transition |
| ERK | extracellular signal-regulated kinase |
| ERM | Ezrin-Radixin-Moesin |
| FAK | focal adhesion kinase |
| FAT1-4 | FAT atypical cadherin 1-4 |
| FBS | fetal bovine serum |
| FERM | 4.1 protein, ezrin, radixin, moesin |
| FGFR | fibroblast growth factor receptor |
| H&E | hematoxylin and eosin |
| HA | hyaluronic acid |
| HBV | hepatitis B virus |
| HCC | hepatocellular carcinoma |
| HCV | hepatitis C virus |
| HEPES | 2-(4-(2-hydroxyethyl)piperazin-1-yl)ethanesulfonic acid |
| HGF | hepatocyte growth factor |
| HNF4α | hepatocyte nuclear factor 4 alpha |
| HPRT1 | hypoxanthine phosphoribosyltransferase 1 |
| HRP | horseradish peroxidase |
| ICAM-1 | intercellular adhesion molecule 1 |
| iCCA | intrahepatic cholangiocarcinoma |
| IDH | isocitrate dehydrogenase |
| IL-6 | interleukin-6 |
| JAG1 | Jagged 1 |
| JAK | janus kinase |
| JNK | c-JUN N-terminal kinase |
| KIBRA | kidney and brain expressed protein |
| KRAS | Kirsten rat sarcoma virus |
| LATS1/2 LFA-1 | large tumor suppressor kinase 1 and 2 lymphocyte function-associated antigen 1 |
| MAPK | mitogen-activated protein kinase |
| MEK | mitogen-activated protein kinase kinase |
| Merlin | Moesin-Ezrin-Radixin-like protein |
| MET | mesenchymal epithelial transition factor receptor |
| MOB1A/1B | Mps one binder homolog A and B |
| MST1/2 | mammalian STE20-like protein kinase 1 and 2 |
| mTOR | mammalian target of rapamycin |
| MYPT1 | myosin phosphatase targeting subunit 1 |
| NF2 | neurofibromatosis type 2 |
| NIH | National Institutes of Health |
| PAK | p21-activated kinase |
| PBS | phosphate-buffered saline |
| PCR | polymerase chain reaction |
| PD-1 | programmed cell death protein 1 |
| PD-L1 | programmed death ligand 1 |
| PFA | paraformaldehyde |
| PI3K | phosphoinositide 3-kinase |
| PKA | protein kinase A |
| PP1δ | Protein Phosphatase 1 δ |
| PTPN3 | tyrosine-protein phosphatase non-receptor type 3 |
| RAC | Ras-related C3 botulinum toxin substrate |
| RAF | rapidly accelerated fibrosarcoma |
| RAS | rat sarcoma |
| RASSF | RAS association domain family |
| qRT-PCR | quantitative reverse transcription PCR |
| RT | room temperature |
| RTK | receptor tyrosine kinase |
| RUNX | Runt-related transcription factor |
| SAV1 | Salvador homolog 1 |
| SMAD | mothers against ecapentaplegic homolog 2 |
| SOX9 | SRY-box transcription factor 9 |
| STAT | signal transducer and activator of transcription |
| TACE | transarterial chemoembolization |
| TAZ | transcriptional co-activator with PDZ-binding motif |
| TBP | TATA box binding protein |
| TEAD | TEA domain |
| TERT | telomerase reverse transcriptase |
| TGFβ | transforming growth factor β |
| TNFα | tumor necrosis factor-α |
| TNM | tumor, node, metastasis |
| TP53 | tumor protein 53 |
| TUNEL | terminal deoxynucleotidyl transferase (TdT) dUTP nick-end labeling |
| VCAM–1 | vascular cell adhesion molecule–1 |
| VEGF | vascular endothelial growth factor |
| WNT | wingless-related integration site |
| YAP | Yes-associated protein |
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA A Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef]
- Wang, G.; Wang, Q.; Liang, N.; Xue, H.; Yang, T.; Chen, X.; Qiu, Z.; Zeng, C.; Sun, T.; Yuan, W.; et al. Oncogenic driver genes and tumor microenvironment determine the type of liver cancer. Cell Death Dis. 2020, 11, 313. [Google Scholar] [CrossRef]
- Sidali, S.; Trepo, E.; Sutter, O.; Nault, J.C. New concepts in the treatment of hepatocellular carcinoma. United Eur. Gastroenterol. J. 2022, 10, 765–774. [Google Scholar] [CrossRef]
- Gelband, H.; Chen, C.J.; Chen, W.; Franceschi, S.; Hall, S.A.; London, W.T.; McGlynn, K.A.; Wild, C.P. Liver Cancer. In Cancer: Disease Control Priorities, 3rd ed.; Gelband, H., Jha, P., Sankaranarayanan, R., Horton, S., Eds.; The International Bank for Reconstruction and Development/The World Bank: Washington, DC, USA, 2015; Volume 3. [Google Scholar]
- Zhang, N.; Bai, H.; David, K.K.; Dong, J.; Zheng, Y.; Cai, J.; Giovannini, M.; Liu, P.; Anders, R.A.; Pan, D. The Merlin/NF2 tumor suppressor functions through the YAP oncoprotein to regulate tissue homeostasis in mammals. Dev. Cell 2010, 19, 27–38. [Google Scholar] [CrossRef] [PubMed]
- Benhamouche, S.; Curto, M.; Saotome, I.; Gladden, A.B.; Liu, C.H.; Giovannini, M.; McClatchey, A.I. Nf2/Merlin controls progenitor homeostasis and tumorigenesis in the liver. Genes Dev. 2010, 24, 1718–1730. [Google Scholar] [CrossRef] [PubMed]
- Zhou, D.; Zhang, Y.; Wu, H.; Barry, E.; Yin, Y.; Lawrence, E.; Dawson, D.; Willis, J.E.; Markowitz, S.D.; Camargo, F.D.; et al. Mst1 and Mst2 protein kinases restrain intestinal stem cell proliferation and colonic tumorigenesis by inhibition of Yes-associated protein (Yap) overabundance. Proc. Natl. Acad. Sci. USA 2011, 108, E1312–E1320. [Google Scholar] [CrossRef] [PubMed]
- Song, H.; Mak, K.K.; Topol, L.; Yun, K.; Hu, J.; Garrett, L.; Chen, Y.; Park, O.; Chang, J.; Simpson, R.M.; et al. Mammalian Mst1 and Mst2 kinases play essential roles in organ size control and tumor suppression. Proc. Natl. Acad. Sci. USA 2010, 107, 1431–1436. [Google Scholar] [CrossRef] [PubMed]
- Lu, L.; Li, Y.; Kim, S.M.; Bossuyt, W.; Liu, P.; Qiu, Q.; Wang, Y.; Halder, G.; Finegold, M.J.; Lee, J.S.; et al. Hippo signaling is a potent in vivo growth and tumor suppressor pathway in the mammalian liver. Proc. Natl. Acad. Sci. USA 2010, 107, 1437–1442. [Google Scholar] [CrossRef]
- Chen, Q.; Zhang, N.; Xie, R.; Wang, W.; Cai, J.; Choi, K.S.; David, K.K.; Huang, B.; Yabuta, N.; Nojima, H.; et al. Homeostatic control of Hippo signaling activity revealed by an endogenous activating mutation in YAP. Genes Dev. 2015, 29, 1285–1297. [Google Scholar] [CrossRef]
- Yi, J.; Lu, L.; Yanger, K.; Wang, W.; Sohn, B.H.; Stanger, B.Z.; Zhang, M.; Martin, J.F.; Ajani, J.A.; Chen, J.; et al. Large tumor suppressor homologs 1 and 2 regulate mouse liver progenitor cell proliferation and maturation through antagonism of the coactivators YAP and TAZ. Hepatology 2016, 64, 1757–1772. [Google Scholar] [CrossRef]
- Nishio, M.; Sugimachi, K.; Goto, H.; Wang, J.; Morikawa, T.; Miyachi, Y.; Takano, Y.; Hikasa, H.; Itoh, T.; Suzuki, S.O.; et al. Dysregulated YAP1/TAZ and TGF-β signaling mediate hepatocarcinogenesis in Mob1a/1b-deficient mice. Proc. Natl. Acad. Sci. USA 2016, 113, E71–E80. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.P.; Lee, J.H.; Kim, T.S.; Kim, T.H.; Park, H.D.; Byun, J.S.; Kim, M.C.; Jeong, W.I.; Calvisi, D.F.; Kim, J.M.; et al. The Hippo-Salvador pathway restrains hepatic oval cell proliferation, liver size, and liver tumorigenesis. Proc. Natl. Acad. Sci. USA 2010, 107, 8248–8253. [Google Scholar] [CrossRef] [PubMed]
- Zender, L.; Spector, M.S.; Xue, W.; Flemming, P.; Cordon-Cardo, C.; Silke, J.; Fan, S.T.; Luk, J.M.; Wigler, M.; Hannon, G.J.; et al. Identification and validation of oncogenes in liver cancer using an integrative oncogenomic approach. Cell 2006, 125, 1253–1267. [Google Scholar] [CrossRef]
- Dong, J.; Feldmann, G.; Huang, J.; Wu, S.; Zhang, N.; Comerford, S.A.; Gayyed, M.F.; Anders, R.A.; Maitra, A.; Pan, D. Elucidation of a universal size-control mechanism in Drosophila and mammals. Cell 2007, 130, 1120–1133. [Google Scholar] [CrossRef] [PubMed]
- Camargo, F.D.; Gokhale, S.; Johnnidis, J.B.; Fu, D.; Bell, G.W.; Jaenisch, R.; Brummelkamp, T.R. YAP1 increases organ size and expands undifferentiated progenitor cells. Curr. Biol. CB 2007, 17, 2054–2060. [Google Scholar] [CrossRef] [PubMed]
- Mranda, G.M.; Xiang, Z.P.; Liu, J.J.; Wei, T.; Ding, Y. Advances in prognostic and therapeutic targets for hepatocellular carcinoma and intrahepatic cholangiocarcinoma: The hippo signaling pathway. Front. Oncol. 2022, 12, 937957. [Google Scholar] [CrossRef]
- Zhao, B.; Wei, X.; Li, W.; Udan, R.S.; Yang, Q.; Kim, J.; Xie, J.; Ikenoue, T.; Yu, J.; Li, L.; et al. Inactivation of YAP oncoprotein by the Hippo pathway is involved in cell contact inhibition and tissue growth control. Genes Dev. 2007, 21, 2747–2761. [Google Scholar] [CrossRef]
- Xu, M.Z.; Yao, T.J.; Lee, N.P.; Ng, I.O.; Chan, Y.T.; Zender, L.; Lowe, S.W.; Poon, R.T.; Luk, J.M. Yes-associated protein is an independent prognostic marker in hepatocellular carcinoma. Cancer 2009, 115, 4576–4585. [Google Scholar] [CrossRef] [PubMed]
- Petrilli, A.M.; Fernandez-Valle, C. Role of Merlin/NF2 inactivation in tumor biology. Oncogene 2016, 35, 537–548. [Google Scholar] [CrossRef]
- You, H.; Ding, W.; Dang, H.; Jiang, Y.; Rountree, C.B. c-Met represents a potential therapeutic target for personalized treatment in hepatocellular carcinoma. Hepatology 2011, 54, 879–889. [Google Scholar] [CrossRef] [PubMed]
- Luo, Z.L.; Cheng, S.Q.; Shi, J.; Zhang, H.L.; Zhang, C.Z.; Chen, H.Y.; Qiu, B.J.; Tang, L.; Hu, C.L.; Wang, H.Y.; et al. A splicing variant of Merlin promotes metastasis in hepatocellular carcinoma. Nat. Commun. 2015, 6, 8457. [Google Scholar] [CrossRef]
- Curto, M.; McClatchey, A.I. Nf2/Merlin: A coordinator of receptor signalling and intercellular contact. Br. J. Cancer 2008, 98, 256–262. [Google Scholar] [CrossRef]
- Sato, T.; Sekido, Y. NF2/Merlin Inactivation and Potential Therapeutic Targets in Mesothelioma. Int. J. Mol. Sci. 2018, 19, 988. [Google Scholar] [CrossRef]
- Cooper, J.; Giancotti, F.G. Molecular insights into NF2/Merlin tumor suppressor function. FEBS Lett. 2014, 588, 2743–2752. [Google Scholar] [CrossRef] [PubMed]
- Ahronowitz, I.; Xin, W.; Kiely, R.; Sims, K.; MacCollin, M.; Nunes, F.P. Mutational spectrum of the NF2 gene: A meta-analysis of 12 years of research and diagnostic laboratory findings. Hum. Mutat. 2007, 28, 1–12. [Google Scholar] [CrossRef]
- Morrison, H.; Sherman, L.S.; Legg, J.; Banine, F.; Isacke, C.; Haipek, C.A.; Gutmann, D.H.; Ponta, H.; Herrlich, P. The NF2 tumor suppressor gene product, merlin, mediates contact inhibition of growth through interactions with CD44. Genes Dev. 2001, 15, 968–980. [Google Scholar] [CrossRef]
- Bai, Y.; Liu, Y.J.; Wang, H.; Xu, Y.; Stamenkovic, I.; Yu, Q. Inhibition of the hyaluronan-CD44 interaction by merlin contributes to the tumor-suppressor activity of merlin. Oncogene 2007, 26, 836–850. [Google Scholar] [CrossRef] [PubMed]
- Hartmann, M.; Parra, L.M.; Ruschel, A.; Bohme, S.; Li, Y.; Morrison, H.; Herrlich, A.; Herrlich, P. Tumor Suppressor NF2 Blocks Cellular Migration by Inhibiting Ectodomain Cleavage of CD44. Mol. Cancer Res. MCR 2015, 13, 879–890. [Google Scholar] [CrossRef] [PubMed]
- Orian-Rousseau, V. CD44 Acts as a Signaling Platform Controlling Tumor Progression and Metastasis. Front. Immunol. 2015, 6, 154. [Google Scholar] [CrossRef]
- Senbanjo, L.T.; Chellaiah, M.A. CD44: A Multifunctional Cell Surface Adhesion Receptor Is a Regulator of Progression and Metastasis of Cancer Cells. Front. Cell Dev. Biol. 2017, 5, 18. [Google Scholar] [CrossRef]
- Luo, Y.; Tan, Y. Prognostic value of CD44 expression in patients with hepatocellular carcinoma: Meta-analysis. Cancer Cell Int. 2016, 16, 47. [Google Scholar] [CrossRef]
- Morine, Y.; Imura, S.; Ikemoto, T.; Iwahashi, S.; Saito, Y.U.; Shimada, M. CD44 Expression Is a Prognostic Factor in Patients with Intrahepatic Cholangiocarcinoma After Surgical Resection. Anticancer Res. 2017, 37, 5701–5705. [Google Scholar] [CrossRef]
- Kunlabut, K.; Vaeteewoottacharn, K.; Wongkham, C.; Khuntikeo, N.; Waraasawapati, S.; Pairojkul, C.; Wongkham, S. Aberrant expression of CD44 in bile duct cancer correlates with poor prognosis. Asian Pac. J. Cancer Prev. 2012, 13, 95–99. [Google Scholar]
- Kim, R.; Kim, S.B.; Cho, E.H.; Park, S.H.; Park, S.B.; Hong, S.K.; Chae, G. CD44 expression in patients with combined hepatocellular cholangiocarcinoma. Ann. Surg. Treat. Res. 2015, 89, 9–16. [Google Scholar] [CrossRef] [PubMed]
- Gu, M.J.; Jang, B.I. Clinicopathologic significance of Sox2, CD44 and CD44v6 expression in intrahepatic cholangiocarcinoma. Pathol. Oncol. Res. 2014, 20, 655–660. [Google Scholar] [CrossRef]
- Ponta, H.; Sherman, L.; Herrlich, P.A. CD44: From adhesion molecules to signalling regulators. Nat. Rev. Mol. Cell Biol. 2003, 4, 33–45. [Google Scholar] [CrossRef]
- Orian-Rousseau, V.; Chen, L.; Sleeman, J.P.; Herrlich, P.; Ponta, H. CD44 is required for two consecutive steps in HGF/c-Met signaling. Genes Dev. 2002, 16, 3074–3086. [Google Scholar] [CrossRef]
- Matzke, A.; Herrlich, P.; Ponta, H.; Orian-Rousseau, V. A five-amino-acid peptide blocks Met- and Ron-dependent cell migration. Cancer Res. 2005, 65, 6105–6110. [Google Scholar] [CrossRef]
- Lammich, S.; Okochi, M.; Takeda, M.; Kaether, C.; Capell, A.; Zimmer, A.K.; Edbauer, D.; Walter, J.; Steiner, H.; Haass, C. Presenilin-dependent intramembrane proteolysis of CD44 leads to the liberation of its intracellular domain and the secretion of an Aβ-like peptide. J. Biol. Chem. 2002, 277, 44754–44759. [Google Scholar] [CrossRef] [PubMed]
- Giovannini, M.; Robanus-Maandag, E.; van der Valk, M.; Niwa-Kawakita, M.; Abramowski, V.; Goutebroze, L.; Woodruff, J.M.; Berns, A.; Thomas, G. Conditional biallelic Nf2 mutation in the mouse promotes manifestations of human neurofibromatosis type 2. Genes Dev. 2000, 14, 1617–1630. [Google Scholar] [CrossRef] [PubMed]
- Dhar, D.; Antonucci, L.; Nakagawa, H.; Kim, J.Y.; Glitzner, E.; Caruso, S.; Shalapour, S.; Yang, L.; Valasek, M.A.; Lee, S.; et al. Liver Cancer Initiation Requires p53 Inhibition by CD44-Enhanced Growth Factor Signaling. Cancer Cell 2018, 33, 1061–1077.e6. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Klemm, J.; Gerardo-Ramirez, M.; Frappart, L.; Castven, D.; Becker, D.; Zoch, A.; Parent, R.; Bartosch, B.; Minnich, K.; et al. Cluster of differentiation 44 promotes osteosarcoma progression in mice lacking the tumor suppressor Merlin. Int. J. Cancer 2020, 147, 2564–2577. [Google Scholar] [CrossRef] [PubMed]
- Bentley, D.R.; Balasubramanian, S.; Swerdlow, H.P.; Smith, G.P.; Milton, J.; Brown, C.G.; Hall, K.P.; Evers, D.J.; Barnes, C.L.; Bignell, H.R.; et al. Accurate whole human genome sequencing using reversible terminator chemistry. Nature 2008, 456, 53–59. [Google Scholar] [CrossRef] [PubMed]
- Pfaffl, M.W.; Horgan, G.W.; Dempfle, L. Relative expression software tool (REST) for group-wise comparison and statistical analysis of relative expression results in real-time PCR. Nucleic Acids Res. 2002, 30, e36. [Google Scholar] [CrossRef]
- Kellendonk, C.; Opherk, C.; Anlag, K.; Schutz, G.; Tronche, F. Hepatocyte-specific expression of Cre recombinase. Genesis 2000, 26, 151–153. [Google Scholar] [CrossRef]
- Postic, C.; Magnuson, M.A. DNA excision in liver by an albumin-Cre transgene occurs progressively with age. Genesis 2000, 26, 149–150. [Google Scholar] [CrossRef]
- Schmits, R.; Filmus, J.; Gerwin, N.; Senaldi, G.; Kiefer, F.; Kundig, T.; Wakeham, A.; Shahinian, A.; Catzavelos, C.; Rak, J.; et al. CD44 regulates hematopoietic progenitor distribution, granuloma formation, and tumorigenicity. Blood 1997, 90, 2217–2233. [Google Scholar] [CrossRef]
- Protin, U.; Schweighoffer, T.; Jochum, W.; Hilberg, F. CD44-deficient mice develop normally with changes in subpopulations and recirculation of lymphocyte subsets. J. Immunol. 1999, 163, 4917–4923. [Google Scholar] [CrossRef]
- Nguyen, Q.; Anders, R.A.; Alpini, G.; Bai, H. Yes-associated protein in the liver: Regulation of hepatic development, repair, cell fate determination and tumorigenesis. Dig Liver Dis. 2015, 47, 826–835. [Google Scholar] [CrossRef]
- Franken, N.A. Clonogenic assay of cells in vitro. Nat. Protoc. 2006, 1, 2315–2319. [Google Scholar] [CrossRef]
- Kim, Y.N.; Koo, K.H.; Sung, J.Y.; Yun, U.J.; Kim, H. Anoikis resistance: An essential prerequisite for tumor metastasis. Int. J. Cell Biol. 2012, 2012, 306879. [Google Scholar] [CrossRef] [PubMed]
- Siegelman, M.H.; Stanescu, D.; Estess, P. The CD44-initiated pathway of T-cell extravasation uses VLA-4 but not LFA-1 for firm adhesion. J. Clin. Investig. 2000, 105, 683–691. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.S.; Hung, Y.; Su, C.H.; Peng, S.T.; Guo, Y.J.; Lai, M.C.; Liu, C.Y.; Hsu, J.W. CD44 cross-linking induces integrin-mediated adhesion and transendothelial migration in breast cancer cell line by up-regulation of LFA-1 (αLβ2) and VLA-4 (α4β1). Exp. Cell Res. 2005, 304, 116–126. [Google Scholar] [CrossRef] [PubMed]
- Olaku, V.; Matzke, A.; Mitchell, C.; Hasenauer, S.; Sakkaravarthi, A.; Pace, G.; Ponta, H.; Orian-Rousseau, V. c-Met recruits ICAM-1 as a coreceptor to compensate for the loss of CD44 in Cd44 null mice. Mol. Biol. Cell 2011, 22, 2777–2786. [Google Scholar] [CrossRef] [PubMed]
- Doetschman, T. Influence of genetic background on genetically engineered mouse phenotypes. Methods Mol. Biol. 2009, 530, 423–433. [Google Scholar] [CrossRef]
- Sanford, L.P.; Kallapur, S.; Ormsby, I.; Doetschman, T. Influence of genetic background on knockout mouse phenotypes. Methods Mol. Biol. 2001, 158, 217–225. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Rao, B.; Lou, J.; Li, J.; Liu, Z.; Li, A.; Cui, G.; Ren, Z.; Yu, Z. The Function of the HGF/c-Met Axis in Hepatocellular Carcinoma. Front. Cell Dev. Biol. 2020, 8, 55. [Google Scholar] [CrossRef] [PubMed]
- Yimlamai, D.; Christodoulou, C.; Galli, G.G.; Yanger, K.; Pepe-Mooney, B.; Gurung, B.; Shrestha, K.; Cahan, P.; Stanger, B.Z.; Camargo, F.D. Hippo pathway activity influences liver cell fate. Cell 2014, 157, 1324–1338. [Google Scholar] [CrossRef] [PubMed]
- Stamenkovic, I.; Yu, Q. Merlin, a “magic” linker between extracellular cues and intracellular signaling pathways that regulate cell motility, proliferation, and survival. Curr. Protein Pept. Sci. 2010, 11, 471–484. [Google Scholar] [CrossRef]
- Zeilstra, J.; Joosten, S.P.; Dokter, M.; Verwiel, E.; Spaargaren, M.; Pals, S.T. Deletion of the WNT target and cancer stem cell marker CD44 in Apc(Min/+) mice attenuates intestinal tumorigenesis. Cancer Res. 2008, 68, 3655–3661. [Google Scholar] [CrossRef]
- Li, H.; Shukla, S.; Frappart, L.; Herrlich, P.; Ploubidou, A. cd44 deletion suppresses atypia in the precancerous mouse testis. Mol. Carcinog. 2019, 58, 621–626. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Bollam, S.R.; White, S.M.; Laughlin, S.Z.; Graham, G.T.; Wadhwa, M.; Chen, H.; Nguyen, C.; Vitte, J.; Giovannini, M.; et al. Rac1-Mediated DNA Damage and Inflammation Promote Nf2 Tumorigenesis but Also Limit Cell-Cycle Progression. Dev. Cell 2016, 39, 452–465. [Google Scholar] [CrossRef] [PubMed]
- Yi, C.; Shen, Z.; Stemmer-Rachamimov, A.; Dawany, N.; Troutman, S.; Showe, L.C.; Liu, Q.; Shimono, A.; Sudol, M.; Holmgren, L.; et al. The p130 isoform of angiomotin is required for Yap-mediated hepatic epithelial cell proliferation and tumorigenesis. Sci. Signal. 2013, 6, ra77. [Google Scholar] [CrossRef]
- Perez, A.; Neskey, D.M.; Wen, J.; Pereira, L.; Reategui, E.P.; Goodwin, W.J.; Carraway, K.L.; Franzmann, E.J. CD44 interacts with EGFR and promotes head and neck squamous cell carcinoma initiation and progression. Oral Oncol. 2013, 49, 306–313. [Google Scholar] [CrossRef] [PubMed]
- Yin, J.; Zhang, H.; Wu, X.; Zhang, Y.; Li, J.; Shen, J.; Zhao, Y.; Xiao, Z.; Lu, L.; Huang, C.; et al. CD44 inhibition attenuates EGFR signaling and enhances cisplatin sensitivity in human EGFR wild-type non-small-cell lung cancer cells. Int. J. Mol. Med. 2020, 45, 1783–1792. [Google Scholar] [CrossRef] [PubMed]
- Morath, I.; Jung, C.; Leveque, R.; Linfeng, C.; Toillon, R.A.; Warth, A.; Orian-Rousseau, V. Differential recruitment of CD44 isoforms by ErbB ligands reveals an involvement of CD44 in breast cancer. Oncogene 2018, 37, 1472–1484. [Google Scholar] [CrossRef]
- Oliferenko, S.; Kaverina, I.; Small, J.V.; Huber, L.A. Hyaluronic acid (HA) binding to CD44 activates Rac1 and induces lamellipodia outgrowth. J. Cell Biol. 2000, 148, 1159–1164. [Google Scholar] [CrossRef]
- Murai, T.; Miyazaki, Y.; Nishinakamura, H.; Sugahara, K.N.; Miyauchi, T.; Sako, Y.; Yanagida, T.; Miyasaka, M. Engagement of CD44 promotes Rac activation and CD44 cleavage during tumor cell migration. J. Biol. Chem. 2004, 279, 4541–4550. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Stamenkovic, I.; Yu, Q. CD44 attenuates activation of the hippo signaling pathway and is a prime therapeutic target for glioblastoma. Cancer Res. 2010, 70, 2455–2464. [Google Scholar] [CrossRef]
- Yu, S.; Cai, X.; Wu, C.; Wu, L.; Wang, Y.; Liu, Y.; Yu, Z.; Qin, S.; Ma, F.; Thiery, J.P.; et al. Adhesion glycoprotein CD44 functions as an upstream regulator of a network connecting ERK, AKT and Hippo-YAP pathways in cancer progression. Oncotarget 2015, 6, 2951–2965. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; He, X.; Wan, Y.; Zhang, H.; Tang, T.; Zhang, M.; Yu, S.; Zhao, W.; Chen, L. CD44 promotes hepatocellular carcinoma progression via upregulation of YAP. Exp. Hematol. Oncol. 2021, 10, 54. [Google Scholar] [CrossRef]
- Fan, Z.; Xia, H.; Xu, H.; Ma, J.; Zhou, S.; Hou, W.; Tang, Q.; Gong, Q.; Nie, Y.; Bi, F. Standard CD44 modulates YAP1 through a positive feedback loop in hepatocellular carcinoma. Biomed. Pharmacother. 2018, 103, 147–156. [Google Scholar] [CrossRef] [PubMed]
- Ooki, T.; Murata-Kamiya, N.; Takahashi-Kanemitsu, A.; Wu, W.; Hatakeyama, M. High-Molecular-Weight Hyaluronan Is a Hippo Pathway Ligand Directing Cell Density-Dependent Growth Inhibition via PAR1b. Dev. Cell 2019, 49, 590–604.e9. [Google Scholar] [CrossRef] [PubMed]
- Sherman, L.; Wainwright, D.; Ponta, H.; Herrlich, P. A splice variant of CD44 expressed in the apical ectodermal ridge presents fibroblast growth factors to limb mesenchyme and is required for limb outgrowth. Genes Dev. 1998, 12, 1058–1071. [Google Scholar] [CrossRef] [PubMed]
- Kaya, G.; Rodriguez, I.; Jorcano, J.L.; Vassalli, P.; Stamenkovic, I. Selective suppression of CD44 in keratinocytes of mice bearing an antisense CD44 transgene driven by a tissue-specific promoter disrupts hyaluronate metabolism in the skin and impairs keratinocyte proliferation. Genes Dev. 1997, 11, 996–1007. [Google Scholar] [CrossRef]
- Matzke, A.; Sargsyan, V.; Holtmann, B.; Aramuni, G.; Asan, E.; Sendtner, M.; Pace, G.; Howells, N.; Zhang, W.; Ponta, H.; et al. Haploinsufficiency of c-Met in cd44−/− mice identifies a collaboration of CD44 and c-Met in vivo. Mol. Cell. Biol. 2007, 27, 8797–8806. [Google Scholar] [CrossRef]
- Fujisaki, T.; Tanaka, Y.; Fujii, K.; Mine, S.; Saito, K.; Yamada, S.; Yamashita, U.; Irimura, T.; Eto, S. CD44 stimulation induces integrin-mediated adhesion of colon cancer cell lines to endothelial cells by up-regulation of integrins and c-Met and activation of integrins. Cancer Res. 1999, 59, 4427–4434. [Google Scholar]
- Zen, K.; Liu, D.Q.; Guo, Y.L.; Wang, C.; Shan, J.; Fang, M.; Zhang, C.Y.; Liu, Y. CD44v4 is a major E-selectin ligand that mediates breast cancer cell transendothelial migration. PLoS ONE 2008, 3, e1826. [Google Scholar] [CrossRef]
- Gong, Y.; Zhang, Y.; Feng, S.; Liu, X.; Lu, S.; Long, M. Dynamic contributions of P- and E-selectins to β2-integrin-induced neutrophil transmigration. FASEB J. 2017, 31, 212–223. [Google Scholar] [CrossRef]
- Videira, P.A.; Silva, M.; Martin, K.C.; Sackstein, R. Ligation of the CD44 Glycoform HCELL on Culture-Expanded Human Monocyte-Derived Dendritic Cells Programs Transendothelial Migration. J. Immunol. 2018, 201, 1030–1043. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name: | Cat. No.: | Company: | Application: B (Blocking) IF (Immunofluorescence) IHC (Immunohistochemistry) WB (Western Blotting) |
|---|---|---|---|
| CD44, clone E7K2Y | 37259 | Cell Signaling Technology, Leiden, The Netherlands | IHC 1:100 WB 1:1000 |
| CD44, clone IM7 | 553131 | BD Biosciences, Heidelberg, Germany | B 1:50 |
| CD44, clone KM201 | MABT78 | Merck KGaA, Darmstadt, Germany | IF 1:50 |
| CK19, clone B-1 | sc-374192 | Santa Cruz Biotechnology, Inc, Heidelberg, Germany | IF 1:100 IHC 1:200 WB 1:500 |
| GAPDH, clone G-9 | 365062 | Santa Cruz Biotechnology, Inc, Heidelberg, Germany | WB 1:1000 |
| HNF4α, clone K9218 | MA1-199 | Thermo Fisher Scientific GmbH, Darmstadt, Germany | IF 1:200 IHC 1:200 WB 1:1000 |
| Ki67, clone D3B5 | 12202 | Cell Signaling Technology, Leiden, The Netherlands | IHC 1:200 |
| LATS1, clone C66B5 | 3477 | Cell Signaling Technology, Leiden, The Netherlands | WB 1:1000 |
| Phospho-LATS1(Thr1079), clone D57D3 | 8654 | Cell Signaling Technology, Leiden, The Netherlands | WB 1:1000 |
| MET, clone B2 | sc-8057 | Santa Cruz Biotechnology, Inc, Heidelberg, Germany | WB 1:500 |
| Phospho-MET | 3126 | Cell Signaling Technology, Leiden, The Netherlands | WB 1:1000 |
| NF2 / Merlin, clone EPR2573(2) | ab109244 | Abcam, Berlin, Germany | WB 1:1000 |
| p44/42 MAPK (ERK1/2), clone L34F12 | 4696 | Cell Signaling Technology, Leiden, The Netherlands | WB 1:1000 |
| Phospho-p44/42 MAPK (ERK1/2) (Thr202/Tyr204) | 9101 | Cell Signaling Technology, Leiden, The Netherlands | WB 1:1000 |
| SOX9 polyclonal | AB5535 | Merck KgaA, Darmstadt, Germany | IF 1:500 IHC 1:200 WB 1:2000 |
| YAP polyclonal | 4912 | Cell Signaling Technology, Leiden, The Netherlands | IF 1:100 IHC 1:200 WB 1:1000 |
| Phospho-YAP (Ser127) polyclonal | 4911 | Cell Signaling Technology, Leiden, The Netherlands | WB 1:1000 |
| Name: | Cat. No.: | Company: | Application: IF (Immunofluorescence) IHC (Immunohistochemistry) WB (Western Blotting) |
|---|---|---|---|
| Goat anti-Mouse IgG (H+L) Secondary Antibody, HRP | 31430 | Thermo Fisher Scientific GmbH, Darmstadt, Germany | WB 1:1000 |
| Goat anti-Rabbit IgG F(ab’)2 Secondary Antibody, HRP | 31461 | Thermo Fisher Scientific GmbH, Darmstadt, Germany | WB 1:1000 |
| Goat anti-mouse (H+L) Secondary Antibody, Alexa Fluor 488 | A-11001 | Thermo Fisher Scientific GmbH, Darmstadt, Germany | IF 1:1000 |
| Goat anti-Rabbit IgG (H+L) Secondary Antibody, Alexa Fluor 488 | A-11034 | Thermo Fisher Scientific GmbH, Darmstadt, Germany | IF: 1:500 |
| Goat anti-Rat IgG (H+L) Secondary Antibody, Alexa Fluor 555 | A-21434 | Thermo Fisher Scientific GmbH, Darmstadt, Germany | IF: 1:500 |
| Name | Primer Sequence 5′-…….-3′ |
|---|---|
| Actin Fwd4 | TGC CCT GAG GCT CTT TTC CA |
| Actin Rev4 | TTG GCA TAG AGG TCT TTA CGG AT |
| Actin Fwd5 | TGT TAC CAA CTG GGA CGA CA |
| Actin Rev5 | ACC AGA GGC ATA CAG GGA CA |
| Cd44 stable Ex5 Fwd1 | AGC ACC CCA GAA AGC TAC ATT |
| Cd44all Fwd1 | TCT GCC AGG CTT TCA ACA GT |
| Cd44all Rev1 | CTG CAC AGA TAG CGT TGG GA |
| Cd44v3 Rev1 | TGA TCC AGA AAA ACT GGG GTA |
| Cd44v6 Rev1 | GTT CTG AAA CCA CGT CTC CTG |
| Cd44v7 Rev1 | TGA TCC AGA AAA ACT GGG GTA |
| Ccn1 Fwd1 | AGA GGC TTC CTG TCT TTG GC |
| Ccn1 Rev1 | CCA AGA CGT GGT CTG AAC GA |
| Ccn2 Fwd3 | GTG CCA GAA CGC ACA CTG |
| Ccn2 Rev3 | CCC CGG TTA CAC TCC AAA |
| Hes1 Fwd1 | CCA GCC AGT GTC AAC ACG A |
| Hes1 Rev1 | AAT GCC GGG AGC TAT CTT TCT |
| Hprt Fwd5 | CCA TCA CAT TGT GGC CCT CT |
| Hprt Rev5 | AAT GTA ATC CAG CAG GTC AGC A |
| Icam1Fwd2 | TCC GTG GGG AGG AGA TAC TG |
| Icam1Rev2 | GGC ATG AGA AAT TGG CTC CG |
| Itgal Fwd1 | CCA CTT CCA CTT CCC GAT CT |
| Itgal Rev1 | AGG TCT CAG GAT AGG CTG CAT |
| Itga4 Fwd2 | TGT AGG ACA CAC CAG GCA TTC |
| Itga4 Rev2 | TGA TGC CCA AGG TGG TAT GTG |
| Itgb1 Fwd1 | TCG ATC CTG TGA CCC ATT GC |
| Itgb1 Rev1 | AGT CTC CAC AAC ATG CAC GA |
| Itgb2 Fwd1 | GCA GAA GGA CGG AAG GAA CA |
| Itgb2 Rev1 | CCA GAT GAC CAG GAG GAG GA |
| Sox9 Fwd1 | CAA GAC TCT GGG CAA GCT CTG |
| Sox9 Rev1 | TCC GCT TGT CCG TTC TTC AC |
| Tbp Fwd1 | GGC CTC TCA GAA GCA TCA CTA |
| Tbp Rev1 | GCC AAG CCC TGA GCA TAA |
| Vcam1 Fwd1 | AAG GGA CGA TTC CGG CAT TT |
| Vcam1 Rev1 | TCG GGC ACA TTT CCA CAA GT |
| Genotype | Age | Number of Mice |
|---|---|---|
| Control: Alb-Cre | 2-wk-old | 0 |
| 6-wk-old | 4 | |
| 20-wk-old | 11 | |
| 32-wk-old | 8 | |
| 48-wk-old | 5 | |
| Cd44+/+;Nf2flox/flox;Alb-Cre | 2-wk-old | 5 |
| 6-wk-old | 5 | |
| 20-wk-old | 12 | |
| 32-wk-old | 17 | |
| 48-wk-old | 9 | |
| Cd44−/−;Nf2flox/flox;Alb-Cre | 2-wk-old | 3 |
| 6-wk-old | 6 | |
| 20-wk-old | 12 | |
| 32-wk-old | 10 | |
| 48-wk-old | 9 | |
| Cd44flox/flox;Nf2flox/flox;Alb-Cre | 2-wk-old | 0 |
| 6-wk-old | 0 | |
| 20-wk-old | 0 | |
| 32-wk-old | 1 | |
| 48-wk-old | 5 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gerardo-Ramírez, M.; Giam, V.; Becker, D.; Groth, M.; Hartmann, N.; Morrison, H.; May-Simera, H.L.; Radsak, M.P.; Marquardt, J.U.; Galle, P.R.; et al. Deletion of Cd44 Inhibits Metastasis Formation of Liver Cancer in Nf2-Mutant Mice. Cells 2023, 12, 1257. https://doi.org/10.3390/cells12091257
Gerardo-Ramírez M, Giam V, Becker D, Groth M, Hartmann N, Morrison H, May-Simera HL, Radsak MP, Marquardt JU, Galle PR, et al. Deletion of Cd44 Inhibits Metastasis Formation of Liver Cancer in Nf2-Mutant Mice. Cells. 2023; 12(9):1257. https://doi.org/10.3390/cells12091257
Chicago/Turabian StyleGerardo-Ramírez, Monserrat, Vanessa Giam, Diana Becker, Marco Groth, Nils Hartmann, Helen Morrison, Helen L. May-Simera, Markus P. Radsak, Jens U. Marquardt, Peter R. Galle, and et al. 2023. "Deletion of Cd44 Inhibits Metastasis Formation of Liver Cancer in Nf2-Mutant Mice" Cells 12, no. 9: 1257. https://doi.org/10.3390/cells12091257