Molecular Phylogeny and Historical Biogeography of the Butterfly Tribe Aeromachini Tutt (Lepidoptera: Hesperiidae) from China


 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Taxon Sampling
2.2. DNA Extraction, PCR Amplification and Sequencing
2.3. Phylogenetic Analyses
2.4. Divergence Time Estimates
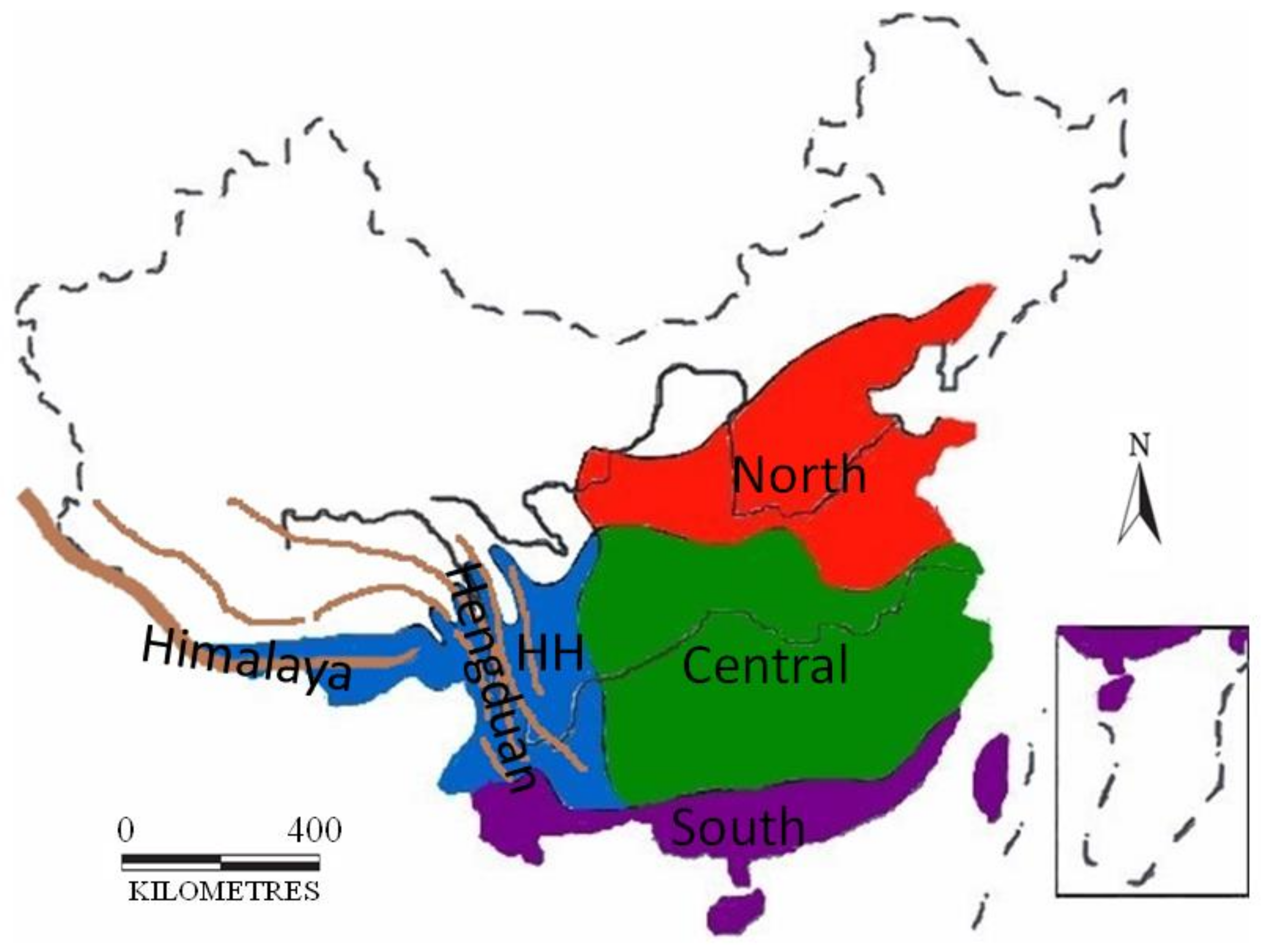
2.5. Ancestral Areas
3. Results
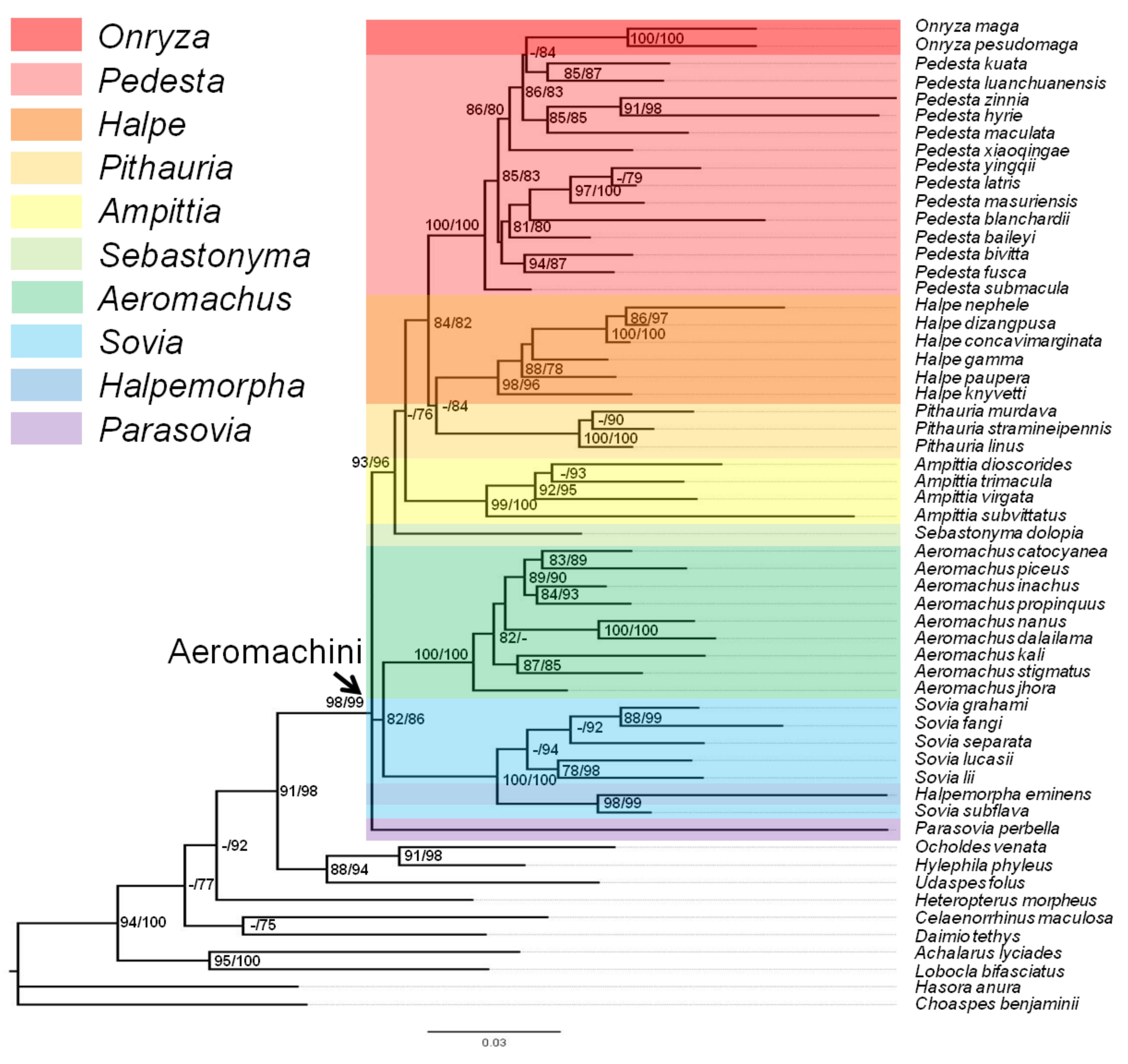
3.1. Analysis and Tree Topology
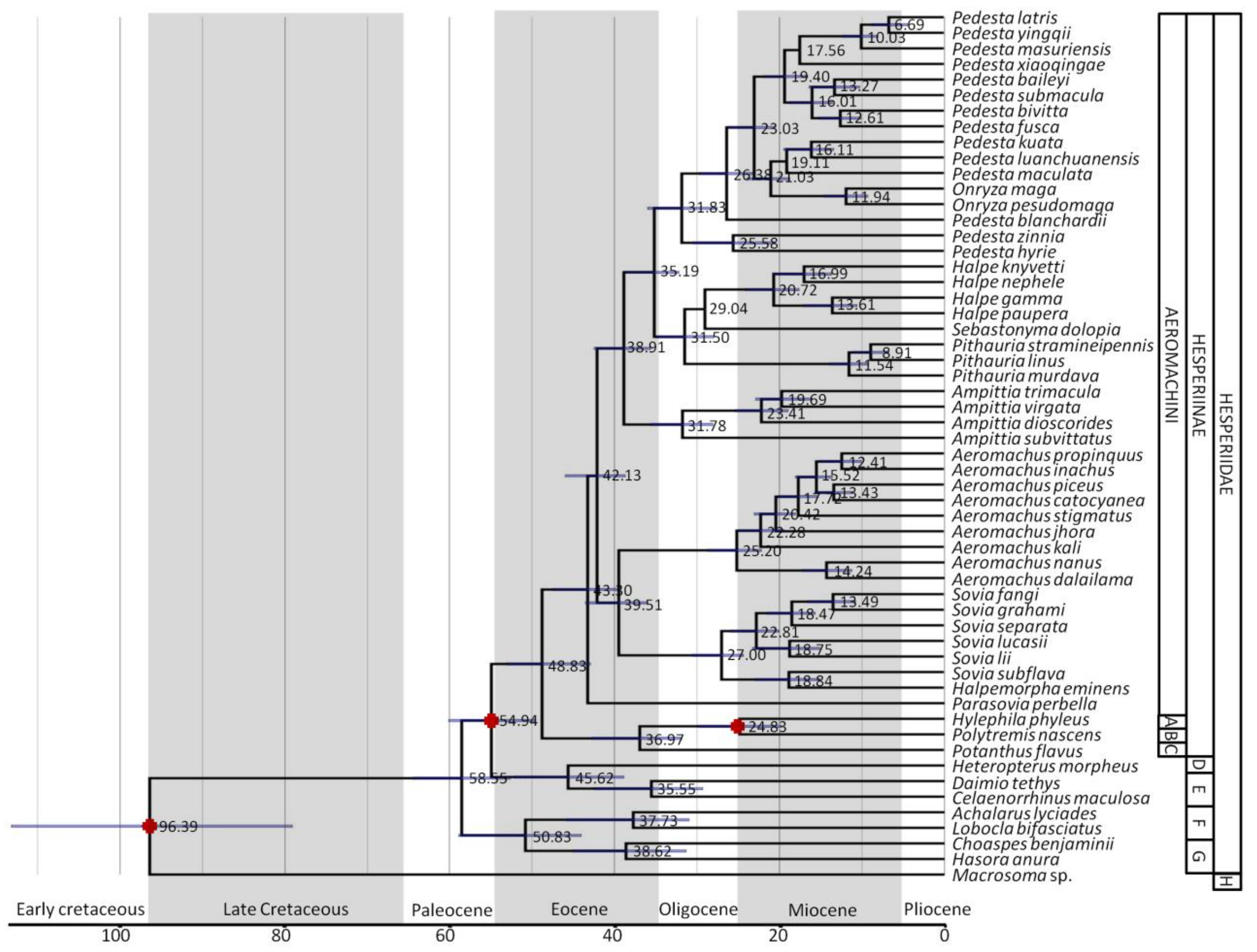
3.2. Divergence Time Estimates
3.3. Ancestral Areas
4. Discussion
4.1. Taxonomic Implications
4.2. Historical Biogeography
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Boggs, C.L.; Watt, W.B.; Ehrlich, P.R. Butterflies: Evolution and Ecology Taking Flight; University of Chicago Press: Chicago, IL, USA, 2003; p. 14. [Google Scholar]
- Brab, M.F.; Roger, V.; Pierce, N.E. Molecular phylogeny and systematics of the Pieridae (Lepidoptera: Papilionoidea): Higher classification and biogeography. Zool. J. Linn. Sci. 2006, 147, 239–275. [Google Scholar] [CrossRef]
- Caterino, M.S.; Reed, R.D.; Kuo, M.M.; Sperling, F.A.H. A partitioned likelihood analysis of swallowtail butterfly phylogeny (Lepidoptera: Papilionidae). Syst. Biol. 2001, 50, 106–127. [Google Scholar] [CrossRef]
- Freitas, A.V.L.; Brown, K.S. Phylogeny of the Nymphalidae (Lepidoptera). Syst. Biol. 2004, 53, 363–383. [Google Scholar] [CrossRef]
- Warren, A.D.; Ogawa, J.R.; Brower, A.V.Z. Phylogenetic relationships of subfamilies and circumscription of tribes in the family Hesperiidae (Lepidoptera: Hesperioidea). Cladistics 2008, 24, 1235. [Google Scholar] [CrossRef]
- Warren, A.D.; Ogawa, J.R.; Brower, A.V.Z. Revised classification of the family Hesperiidae (Lepidoptera: Hesperioidea) based on combined molecular and morphological data. Syst. Entomol. 2009, 34, 467–523. [Google Scholar] [CrossRef]
- Sahoo, R.K.; Warren, A.D.; Wahlberg, N.; Brower, A.V.Z.; Lukhtanov, V.A.; Kodandaramaiah, U. Ten genes and two topologies: An exploration of higher relationships in skipper butterflies (Hesperiidae). PeerJ 2016, 4, e2653. [Google Scholar] [CrossRef]
- Toussaint, E.F.A.; Breinholt, J.W.; Earl, C.; Warren, A.D.; Brower, A.V.Z.; Yago, M.; Dexter, K.M.; Espeland, M.; Pierce, N.E.; Lohman, D.J.; et al. Anchored phylogenomics illuminates the skipper butterfly tree of life. BMC Evol. Biol. 2018, 18, 101. [Google Scholar] [CrossRef] [PubMed]
- Eliot, J.N. An analysis of the Eurasian and Australian Neptini. Bull. Br. Mus. Nat. Hist. 1969, 15, l50–155. [Google Scholar]
- Huang, H. Systematic and Taxonomic Study on the Tribe Aeromachini Tutt (Lepidoptera: Hesperiidae) from China. Master’s Thesis, Shanghai Normal University, Shanghai, China, 2009. [Google Scholar]
- Evans, W.H. A revision of the Genus Aeromachus de N. (Lepidoptera: Hesperiidae). Proc. R. Soc. B 1943, 12, 97–101. [Google Scholar] [CrossRef]
- Devyatkin, A.L. New Hesperiidae from North Vietnam, with the description of a new genus (Lepidoptera: Rhopalocera). Atalanta 1996, 27, 595–604. [Google Scholar]
- Huang, H.; Zhan, C.H. Notes on the genera Thoressa and Pedesta, with description of a new species from South China. Neue Entomol. Nachr. 2004, 57, 179–186. [Google Scholar]
- Jiang, W.B.; Zhu, J.Q.; Song, C.; Li, X.Y.; Yang, Y.; Yu, W.D. Molecular phylogeny of the butterfly genus Polytremis (Hesperiidae, Hesperiinae, Baorini) in China. PLoS ONE 2013, 8, e84098. [Google Scholar] [CrossRef] [PubMed]
- Jiang, W.B.; He, H.Y.; Li, Y.D.; Ren, M.Y.; Ma, Y.Z.; Zheng, L.L.; Zhu, J.Q.; Yu, W.D. Taxonomic status and molecular phylogeography of two sibling species of Polytremis (Lepidoptera: Hesperiidae). Sci. Rep. 2016, 6, 20820. [Google Scholar] [CrossRef]
- Jiang, W.B.; He, H.Y.; Li, Y.Y.; Wang, Y.; Ge, C.; Zhu, J.Q.; Yu, W.D. Molecular phylogeny of the tutterfly tribe Baorini Doherty, 1886 (Hesperiidae, Hesperiinae) in China. Zootaxa 2019, 4565, 108–116. [Google Scholar] [CrossRef]
- Nguyen, L.T.; Schmidt, H.A.; Von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Huang, Z.F.; Chiba, H.; Jin, J.; Kizhakke, A.G.; Wang, M.; Kunte, K.; Fan, X.L. A multilocus phylogenetic framework of the tribe Aeromachini (Lepidoptera: Hesperiidae: Hesperiinae), with implications for taxonomy and biogeography. Syst. Entomol. 2019, 44, 163–178. [Google Scholar] [CrossRef]
- Rand, D.B.; Heath, A.; Suderman, T.; Pierce, N.E. Phylogeny and life history evolution of the genus Chrysoritis within the Aphnaeini (Lepidoptera: Lycaenidae), inferred from mitochondrial cytochrome oxidase I sequences. Mol. Phylogenet. Evol. 2000, 17, 85–96. [Google Scholar] [CrossRef]
- Jiang, W.B.; Zhu, J.Q.; Chen, M.H.; Yang, Q.C.; Du, X.; Chen, S.Y.; Zhang, L.N.; Yu, Y.M.; Yu, W.D. Wolbachia infection status and genetic structure in natural populations of Polytremis nascens (Lepidoptera: Hesperiidae). Infect. Genet. Evol. 2014, 27, 202–211. [Google Scholar] [CrossRef] [PubMed]
- Raupach, M.J.; Astrin, J.J.; Hannig, K.; Peters, M.K.; Stoeckle, M.Y.; Wägele, J.W. Molecular species identification of Central European ground beetles (Coleoptera: Carabidae) using nuclear rDNA expansion segments and DNA barcodes. Front. Zool. 2010, 7, 26. [Google Scholar] [CrossRef]
- Jiang, W.B.; Zhu, J.Q.; Wu, Y.J.; Li, L.Z.; Li, Y.Y.; Ge, C.; Wang, Y.; Endersby, N.M.; Hoffmann, A.A.; Yu, W.D. Influence of Wolbachia infection on mitochondrial DNA variation in the genus Polytremis (Lepidoptera: Hesperiidae). Mol. Phylogenet. Evol. 2018, 129, 158–170. [Google Scholar] [CrossRef] [PubMed]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor an analysis program for Windows 95/98/NT. Nucleic Acids Symp. 1999, 41, 95–98. [Google Scholar]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed]
- Lanfear, R.; Calcott, B.; Ho, S.Y.W.; Guindon, S. PartitionFinder: Combined selection of partitioning schemes and substitution models for phylogenetic analyses. Mol. Biol. Evol. 2012, 29, 1695–1701. [Google Scholar] [CrossRef]
- Hoang, D.T.; Chernomor, O.; von Haeseler, A.; Minh, B.Q.; Vinh, L.S. UFBoot2: Improving the ultrafast bootstrap approximation. Mol. Biol. Evol. 2018, 35, 518–522. [Google Scholar] [CrossRef] [PubMed]
- Minh, B.Q.; Nguyen, M.A.T.; Von Haeseler, A. Ultrafast approximation for phylogenetic bootstrap. Mol. Biol. Evol. 2013, 30, 1188–1195. [Google Scholar] [CrossRef] [PubMed]
- Excoffier, L.; Laval, G.; Schneider, S. Arlequin ver. 3.0: An integrated software package for population genetics data analysis. Evol. Bioinform. 2005, 1, 47–50. [Google Scholar] [CrossRef]
- Heikkilä, M.; Kaila, L.; Mutanen, M.; Peña, C.; Wahlberg, N. Cretaceous origin and repeated tertiary diversification of the redefined butterflies. Proc. R. Soc. Lond. B Biol. Sci. 2012, 279, 1093–1099. [Google Scholar] [CrossRef] [PubMed]
- Chazot, N.; Wahlberg, N.; Freitas, A.V.L.; Miller, C.; Labandeira, C.; Sohn, J.; Sahoo, R.K.; Seraphim, N.; De Jong, R.; Keikkila, M. The trials and tribulations of priors and posteriors in Bayesian timing of divergence analyses: The age of butterfles revisited. BioRxiv 2018. [Google Scholar] [CrossRef]
- De Jong, R. Reconstructing a 55-million-year-old butterfly (Lepidoptera: Hesperiidae). Eur. J. Entomol. 2016, 113, 423–428. [Google Scholar] [CrossRef]
- Fu, Y.X.; Li, W.H. Statistical tests of neutrality of mutations. Genetics 1993, 133, 693–709. [Google Scholar] [PubMed]
- Chou, I. Monographia Rhopalocerorum Sinensium; Henan Scientific and Technological Press: Zhengzhou, China, 1994; p. 854. [Google Scholar]
- Fan, X.L.; Wang, M. A New Species of the Genus Thoressa Swinhoe (Lepidoptera: Hesperiidae) from China. Entomol. News 2009, 120, 438–441. [Google Scholar] [CrossRef]
- Huang, H. Some new butterflies from China-2. Atalanta 2002, 33, 109–122. [Google Scholar]
- Huang, H. A list of butterflies collected from Nujiang and Dulongjiang, China with descriptions of new species, new subspecies and revisional notes. Neue Entomol. Nachr. 2003, 55, 3–114. [Google Scholar]
- Huang, H.; Wu, C.S. New and little known Chinese butterflies in the collection of the Institute of Zoology, Academia Sinica, Beijing-l. Neue Entomol. Nachr. 2003, 55, 115–144. [Google Scholar]
- Wang, Z.G.; Niu, Y. Description of new species of Chinese butterflies (in Chinese). Entomotaxonomia 2002, 24, 276–282. [Google Scholar]
- Xue, G.X.; Li, M.; Nan, W.H.; Jia, X.L.; Huang, S.Y.; Sun, H.; Li, X.J. A new species of the genus Sovia (Lepidoptera: Hesperiidae) from Qinling-Daba Mountains of China. Zootaxa 2015, 3985, 583–590. [Google Scholar] [CrossRef] [PubMed]
- Xue, G.X.; Li, Y.F.; Liu, Z.H.; Li, M.; Ren, Y.D. Distribution of Onryza maga (Leech, 1890) (Lepidoptera: Hesperiidae) with description of female genitalia and taxonomic notes. Entomol. Fenn. 2016, 27, 71–76. [Google Scholar]
- Yuan, F.; Yuan, X.Q.; Wang, Z.Q. Checklist of the genus Halpe (Lepidoptera: Hesperiidae) from China with description of a new species. Acta Zootaxonomica Sin. 2007, 32, 308–311. [Google Scholar]
- Zhu, J.Q.; Mao, W.W.; Chen, Z.B. A new species of Onryza Watson, 1893 (Lepidoptera: Hesperiidae) from China. Zootaxa 2017, 4216, 94–100. [Google Scholar] [CrossRef] [PubMed]
- Ali, S.S.; Yu, Y.; Pfosser, M.; Wetschnig, W. Inferences of biogeographical histories within subfamily Hyacinthoideae using S-DIVA and Bayesian binary MCMC analysis implemented in RASP (Reconstruct Ancestral State in Phylogenies). Ann. Bot. 2012, 109, 95–107. [Google Scholar] [CrossRef]
- Yu, Y.; Harris, A.J.; He, X.J. S-DIVA (statistical dispersal-vicariance analysis): A tool for inferring biogeographic histories. Mol. Phylogenet. Evol. 2010, 56, 848–850. [Google Scholar] [CrossRef] [PubMed]
- Ree, R.H.; Smith, S.A. Maximum likelihood inference of geographic range evolution by dispersal, local extinction, and cladogenesis. Syst. Biol. 2008, 57, 4–14. [Google Scholar] [CrossRef] [PubMed]
- Ree, R.H.; Moore, B.R.; Webb, C.O.; Donoghue, M.J.; Crandall, K. A likelihood framework for inferring the evolution of geographic range on phylogenetic trees. Evolution 2005, 59, 2299–2311. [Google Scholar] [CrossRef]
- Che, J.; Zhoua, W.; Hua, J.S.; Yana, F.; Papenfuss, T.J.; Wake, D.B.; Zhang, Y.P. Spiny frogs (Paini) illuminate the history of the Himalayan region and Southeast Asia. Proc. Natl. Acad. Sci. USA 2010, 107, 13765–13770. [Google Scholar] [CrossRef]
- Procheş, Ş.; Ramdhani, S. The world’s zoogeographical regions confimed by cross-taxon analyses. Bioscience 2012, 62, 260–270. [Google Scholar] [CrossRef]
- Zhang, R. Zoogeographical regions of China. Bull. Biol. 1987, 24, 196–202. [Google Scholar]
- Huang, H. Notes on the genus Thoressa Swinhoe, [1913] from China, with the description of a new species. Atlanta 2011, 42, 193–200. [Google Scholar]
- Zhai, Q.; Xue, G.X.; Li, M. DNA barcoding-based sexual association of Sovia lucasii and S. lii (Lepidoptera: Hesperiidae), with description of a new subspecies. PLoS ONE 2017, 12, e0183847. [Google Scholar] [CrossRef]
- Evans, W.H. A Catalogue of the Hesperiidae from Europe, Asia and Australia in the British Museum (Natural History); Trustees of the British Museum: London, UK, 1949; p. 12458. [Google Scholar]
- Cong, Q.; Shen, J.; Li, W.; Borek, D.; Otwinowski, Z.; Grishin, N.V. The first complete genomes of Metalmarks and the classification of butterfly families. Genomics 2017, 109, 485–493. [Google Scholar] [CrossRef] [PubMed]
- Espeland, M.; Breinholt, J.; Willmott, K.R.; Warren, A.D.; Vila, R.; Toussaint, E.F.A.; Maunsell, S.C.; Aduse-Poku, K.; Talavera, G.; Eastwood, R.; et al. A comprehensive and dated phylogenomic analysis of butterflies. Curr. Biol. 2018, 28, 770–778. [Google Scholar] [CrossRef] [PubMed]
- Sahoo, R.K.; Warren, A.D.; Collins, S.C.; Kodandaramaiah, U. Hostplant change and paleoclimatic events explain diversification shifts in skipper butterflies (Family: Hesperiidae). BMC Evol. Biol. 2017, 17, 174. [Google Scholar] [CrossRef]
- Wahlberg, N.; Leneveu, J.; Kodandaramaiah, U.; Pena, C.; Nylin, S.; Freitas, A.V.L.; Brower, A.V.Z. Nymphalid butterflies diversify following near demise at the Cretaceous/Tertiary boundary. Proc. Biol. Sci. 2009, 276, 4295–4302. [Google Scholar] [CrossRef] [PubMed]
- Rasnitsyn, A.P.; Quicke, D.L.J. History of Insects; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2002. [Google Scholar]
- Harrison, T.M.; Copeland, P.; Kidd, W.; Yin, A. Raising Tibet. Science 1992, 255, 1663–1670. [Google Scholar] [CrossRef] [PubMed]
- Coleman, M.; Hodges, K. Evidence for Tibetan plateau uplift before 14 Myr ago from a new minimum age for east-west extension. Nature 1995, 374, 49–52. [Google Scholar] [CrossRef]
- Shi, Y.F.; Li, J.J.; Li, B.Y.; Yao, T.D.; Wang, S.M.; Li, S.J.; Cui, Z.J.; Wang, F.B.; Pan, B.T.; Fang, X.M.; et al. Uplift of the Qinghai-Xizang (Tibetan) plateau and East Asia environmental change during late Cenozoic. Acta Geograph. Sin. 1999, 54, 10–21. [Google Scholar]
- Spicer, R.A.; Harris, N.B.W.; Widdowson, M.; Herman, A.B.; Guo, S.; Valdes, P.J.; Wolfe, J.A.; Kelley, S.P. Constant elevation of southern Tibet over the past 15 million years. Nature 2003, 421, 622–624. [Google Scholar] [CrossRef]
- Clift, P.D. Controls on the erosion of Cenozoic Asia and the flx of clastic sediment to the ocean. Earth Planet. Sci. Lett. 2006, 241, 571–580. [Google Scholar] [CrossRef]
- Li, S.; Hughes, A.C.; Su, T.; Anberree, J.L.; Oskolski, A.A.; Sun, M.; Ferguson, D.K.; Zhou, Z.K. Fire dynamics under monsoonal climate in Yunnan, SW China: Past, present and future. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2017, 465, 168–176. [Google Scholar] [CrossRef]
- Tang, C.Q.; Matsui, T.; Ohashi, H.; Dong, Y.F.; Momohara, A.; Herrando-Moraira, S.; Qian, S.H.; Yang, Y.C.; Ohsawa, M.; Luu, H.T. Identifying long-term stable refugia for relict plant species in East Asia. Nat. Commun. 2018, 9, 4488. [Google Scholar] [CrossRef] [PubMed]
- Tzedakis, P.C.; Lawson, I.T.; Frogley, M.R.; Hewitt, G.M.; Preece, R.C. Buffered tree population changes in a quaternary refugium: Evolutionary implications. Science 2002, 297, 2044–2047. [Google Scholar] [CrossRef] [PubMed]
- Hampe, A.; Petit, R.J. Conserving biodiversity under climate change: The rear edge matters. Ecol. Lett. 2005, 8, 461–467. [Google Scholar] [CrossRef] [PubMed]
- Harrison, S.; Noss, R. Viewpoint: Part of a special issue on endemics hotspots, endemism hotspots are linked to stable climatic refugia. Ann. Bot. 2017, 119, 207–214. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene/Region | Primers | Sequence (5′–3′) | Amplicon Length | Annealing Temperature | Reference |
|---|---|---|---|---|---|
| COI | HCO2198 LCO1490 | TAAACTTCAGGGTGACCAAAAAATCA GGTCAACAAATCATAAAGATATTGG | 487 bp | 42 °C | [19] |
| COII | PIERRE EVA | AGAGCCTCTCCTTTAATAGAACA GAGACCATTACTTGCTTTCAGTCATCT | 637 bp | 45 °C | [20] |
| D3 region of 28S rDNA | CD3F CD3R | GGACCCGTCTTGAAACAC GCATAGTTCACCATCTTTC | 240 bp | 52 °C | [21] |
| V4 region of 18S rDNA | CV4F CV4R | TGGTGCCAGCAGCCGCGGTAA CCTCTAACGTCGCAATACGAATGCCC | 381 bp | 56 °C | [21] |
| V7 region of 18S rDNA | CV7F CV7R | CTTAAAGGAATTGACGGAGGGCACCACC GATTCCTTCAGTGTAGCGCGCGTG | 400 bp | 58 °C | [21] |
| Gene Fragment | Nucleotide Sites | Variable Sites | Parsimony Informative Sites | A | T | C | G | Accession Numbers |
|---|---|---|---|---|---|---|---|---|
| COI | 478 | 259 | 219 | 28.9% | 39.5% | 17.6% | 13.9% | MK344780-MK344909 |
| COII | 625 | 293 | 260 | 35.6% | 40.8% | 13.5% | 10.1% | MK344911-MK345027 |
| D3 region of 28S rDNA | 235 | 100 | 52 | 25.2% | 19.3% | 25.1% | 30.4% | MK345289-MK345418 |
| V4 region of 18S rDNA | 392 | 69 | 37 | 24.3% | 28.1% | 20.2% | 27.4% | MK345029-MK345156 |
| V7 region of 18S rDNA | 363 | 154 | 88 | 21.6% | 24.4% | 24.1% | 29.8% | MK345158-MK345287 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Y.; Zhu, J.; Ge, C.; Wang, Y.; Zhao, Z.; Ma, S.; Hoffmann, A.A.; Endersby, N.M.; Liu, Q.; Yu, W.; et al. Molecular Phylogeny and Historical Biogeography of the Butterfly Tribe Aeromachini Tutt (Lepidoptera: Hesperiidae) from China. Cells 2019, 8, 294. https://doi.org/10.3390/cells8040294
Li Y, Zhu J, Ge C, Wang Y, Zhao Z, Ma S, Hoffmann AA, Endersby NM, Liu Q, Yu W, et al. Molecular Phylogeny and Historical Biogeography of the Butterfly Tribe Aeromachini Tutt (Lepidoptera: Hesperiidae) from China. Cells. 2019; 8(4):294. https://doi.org/10.3390/cells8040294
Chicago/Turabian StyleLi, Yuanyuan, Jianqing Zhu, Chen Ge, Ying Wang, Zimiao Zhao, Shuojia Ma, Ary A. Hoffmann, Nancy M. Endersby, Qunxiu Liu, Weidong Yu, and et al. 2019. "Molecular Phylogeny and Historical Biogeography of the Butterfly Tribe Aeromachini Tutt (Lepidoptera: Hesperiidae) from China" Cells 8, no. 4: 294. https://doi.org/10.3390/cells8040294