The Petunia CHANEL Gene is a ZEITLUPE Ortholog Coordinating Growth and Scent Profiles

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Gene Identification and Phylogenetic Analysis
2.2. Silencing of PhCHL
2.3. Plant Growth Conditions and Sampling
2.4. Housekeeping Genes and Gene Expression Analysis by qPCR
2.5. Image Acquisition
2.6. Scent Analysis
3. Results
3.1. The Petunia Genomes Have a Single ZTL Gene
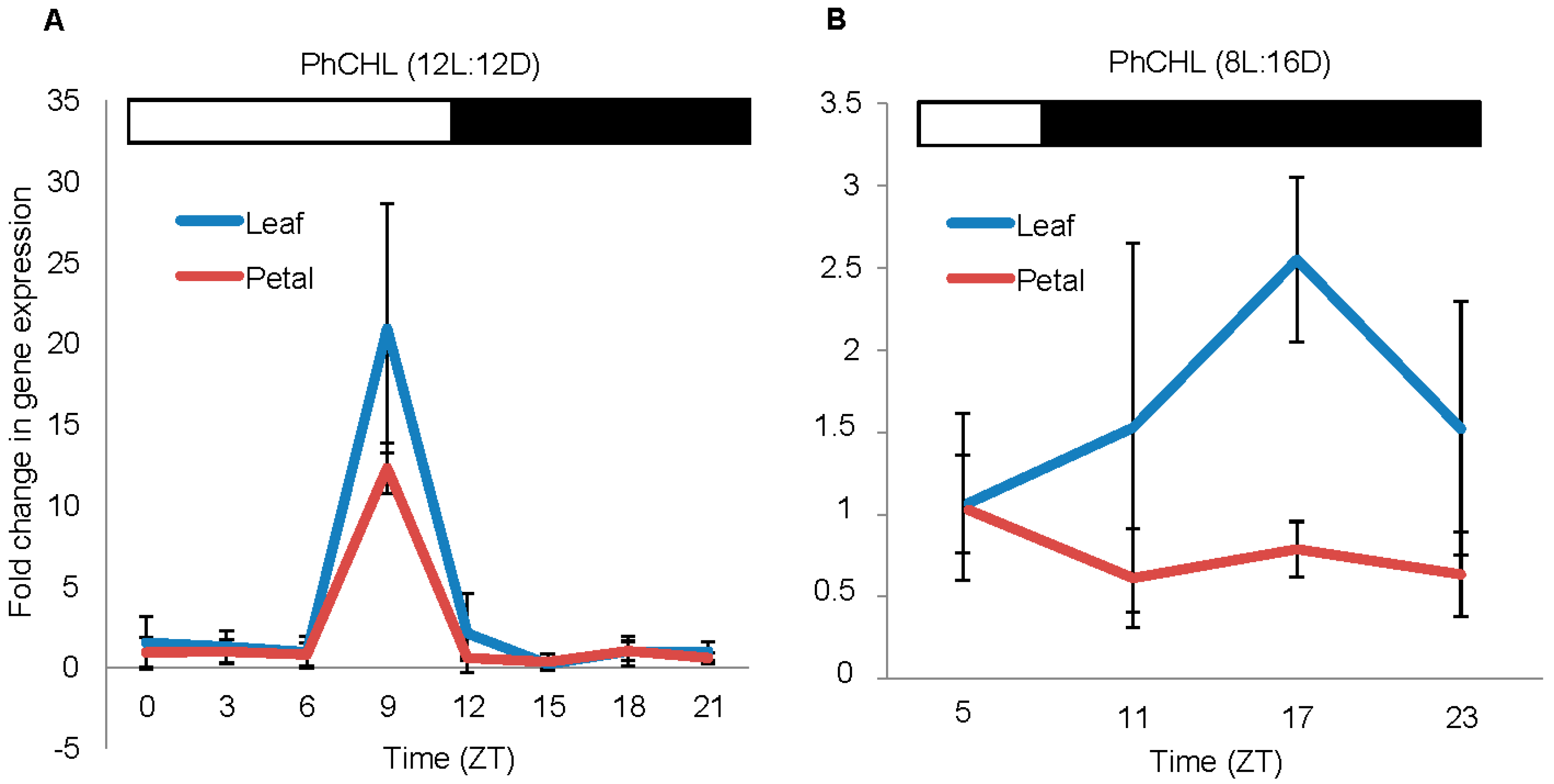
3.2. The Expression of PhCHL is Organ Specific and Affected by Day Length
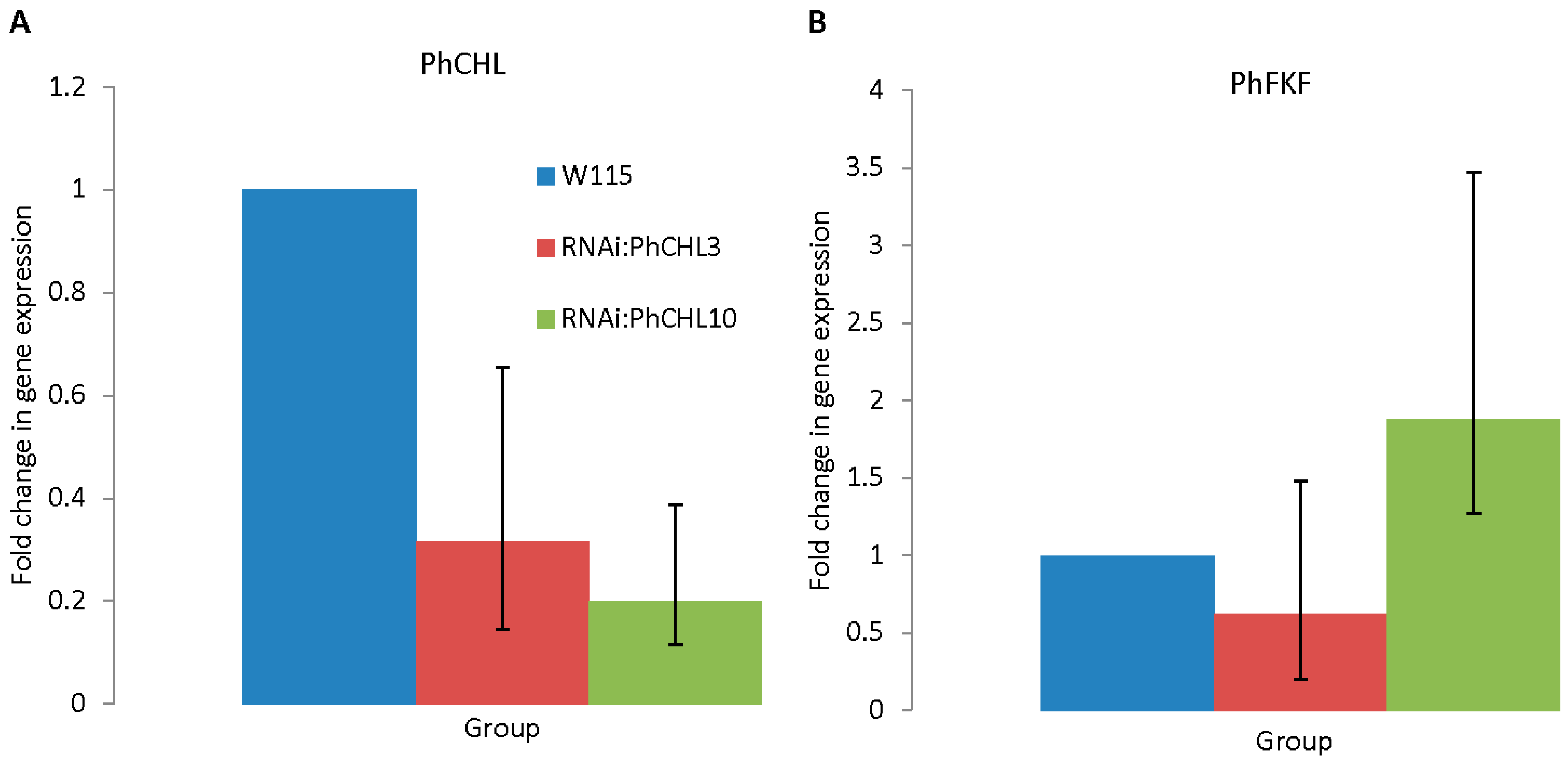
3.3. Silencing PhCHL Does Not Affect PhFKF
3.4. PhCHL Is a Positive Regulator of Lateral Organ Growth
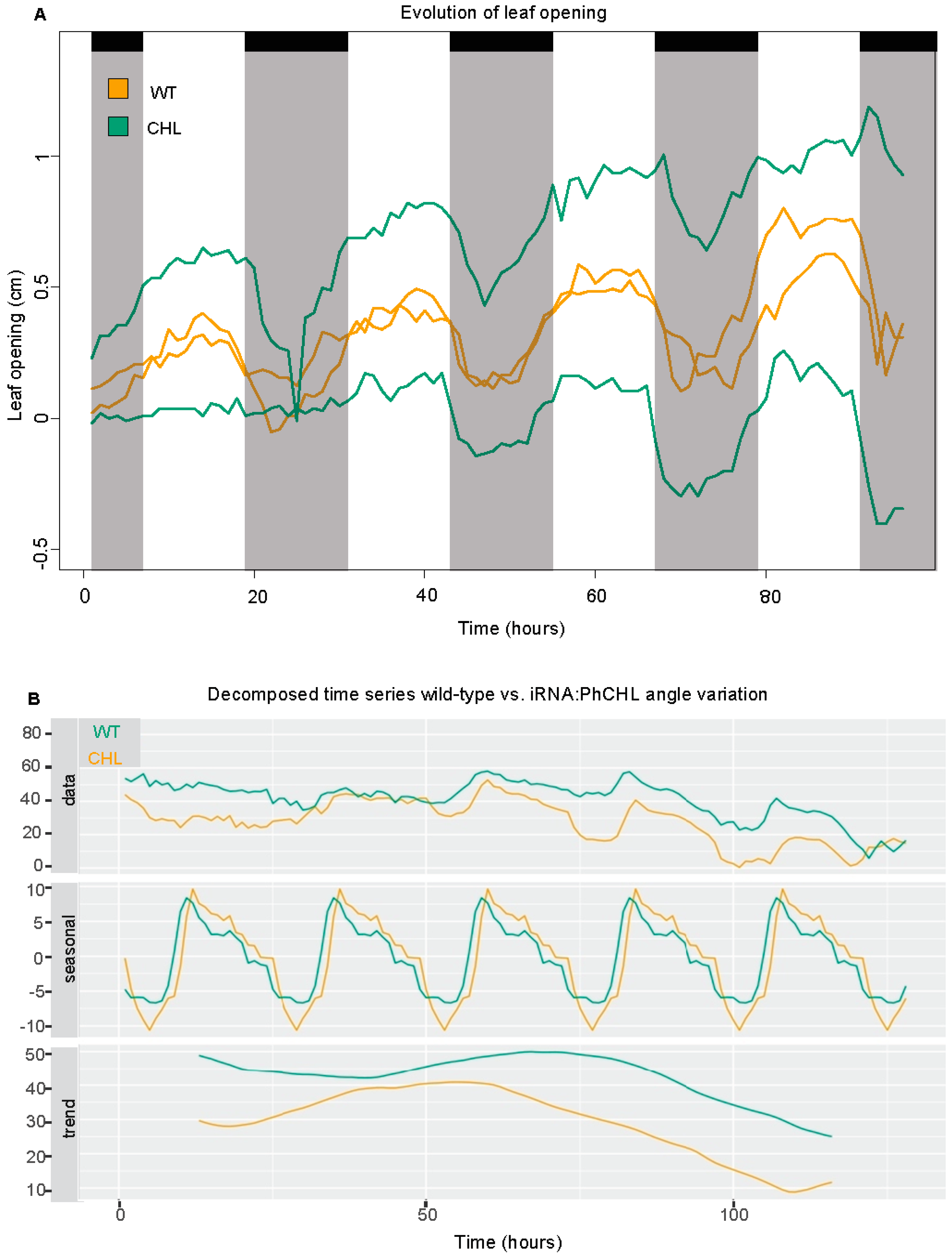
3.5. PhCHL Is Involved in Flower Angle Changes but Not Leaf Movement
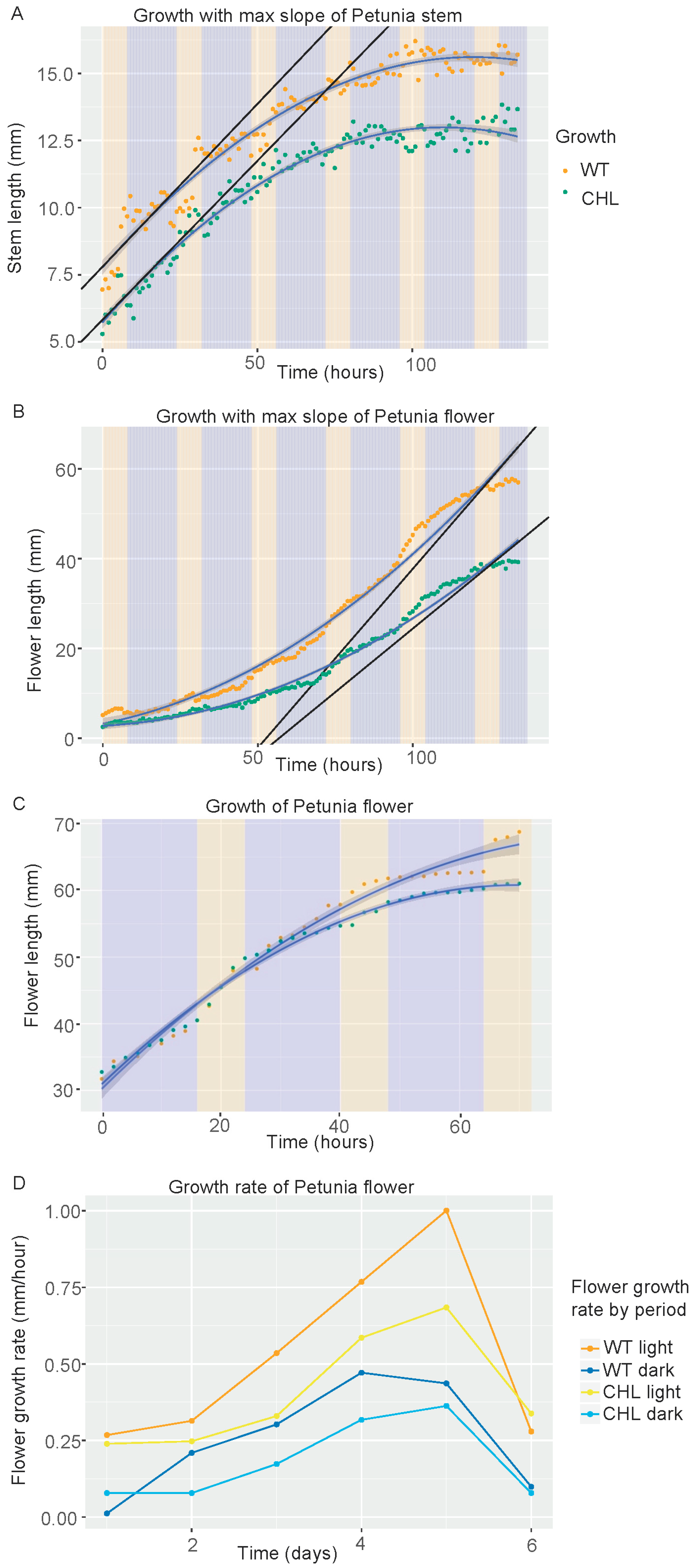
3.6. Differential Effect of PhCHL on Stem and Flower Growth Rate
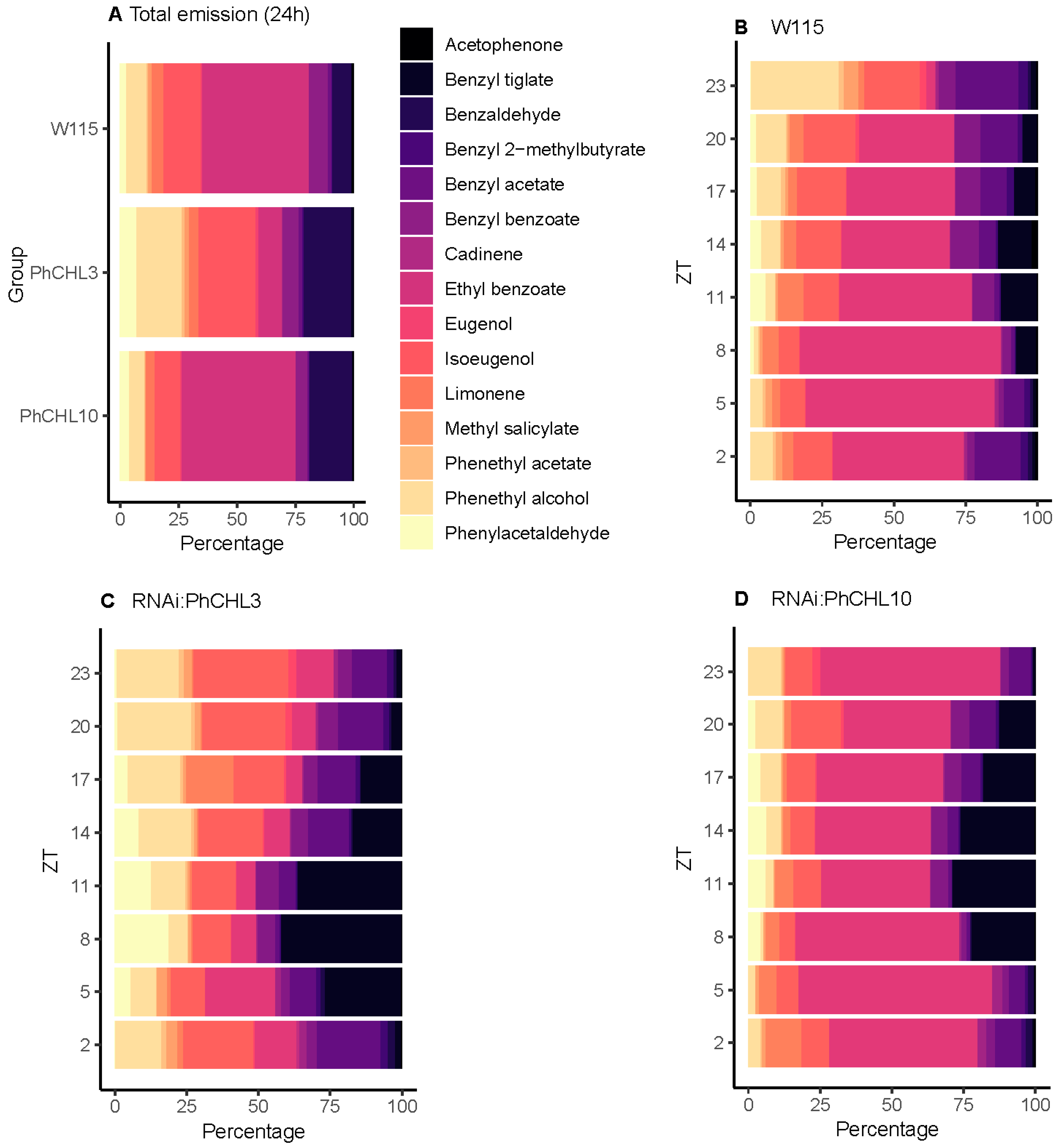
3.7. PhCHL Coordinates Daily Changes in Scent Profiles
3.8. PhCHL Is Required for Timing of Scent Profiles
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Barton, M.K. Twenty years on: The inner workings of the shoot apical meristem, a developmental dynamo. Dev. Biol. 2010, 341, 95–113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Angenent, G.C.; Immink, R.G.H.; Kaufmann, K. The “ABC” of MADS domain protein behaviour and interactions. Semin. Cell Dev. Biol. 2010, 21, 87–93. [Google Scholar]
- Egea-Cortines, M.; Saedler, H.; Sommer, H. Ternary complex formation between the MADS-box proteins SQUAMOSA, DEFICIENS and GLOBOSA is involved in the control of floral architecture in Antirrhinum majus. EMBO J. 1999, 18, 5370–5379. [Google Scholar] [CrossRef] [PubMed]
- Honma, T.; Goto, K. Complexes of MADS-box proteins are sufficient to convert leaves into floral organs. Nature 2001, 409, 525–529. [Google Scholar] [CrossRef] [PubMed]
- Bernardello, G.; Anderson, G.J.; Stuessy, T.F.; Crawford, D.J. A survey of floral traits, breeding systems, floral visitors, and pollination systems of the angiosperms of the Juan Fernandez Islands (Chile). Bot. Rev. 2001, 67, 255–308. [Google Scholar] [CrossRef]
- Hepworth, J.; Lenhard, M. Regulation of plant lateral-organ growth by modulating cell number and size. Curr. Opin. Plant Biol. 2014, 17, 36–42. [Google Scholar] [CrossRef] [PubMed]
- Manchado-Rojo, M.; Weiss, J.; Egea-Cortines, M. Validation of Aintegumenta as a gene to modify floral size in ornamental plants. Plant Biotechnol. J. 2014, 12, 1053–1065. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krizek, B.A. Ectopic expression AINTEGUMENTA in Arabidopsis plants results in increased growth of floral organs. Dev. Genet. 1999, 25, 224–236. [Google Scholar] [CrossRef]
- Kuluev, B.; Avalbaev, A.; Nurgaleeva, E.; Knyazev, A.; Nikonorov, Y.; Chemeris, A. Role of AINTEGUMENTA-like gene NtANTL in the regulation of tobacco organ growth. J. Plant Physiol. 2015, 189, 11–23. [Google Scholar] [CrossRef] [PubMed]
- Muhlemann, J.K.; Maeda, H.; Chang, C.Y.; Miguel, P.S.; Baxter, I.; Cooper, B.; Perera, M.A.; Nikolau, B.J.; Vitek, O.; Morgan, J.A.; et al. Developmental Changes in the Metabolic Network of Snapdragon Flowers. PLoS ONE 2012, 7, e40381. [Google Scholar] [CrossRef] [PubMed]
- Manchado-Rojo, M.; Delgado-Benarroch, L.; Roca, M.J.; Weiss, J.; Egea-Cortines, M. Quantitative levels of Deficiens and Globosa during late petal development show a complex transcriptional network topology of B function. Plant J. 2012, 72, 294–307. [Google Scholar] [CrossRef] [PubMed]
- Fenske, M.P.; Imaizumi, T. Circadian Rhythms in Floral Scent Emission. Front. Plant Sci. 2016, 7, 462. [Google Scholar] [CrossRef] [PubMed]
- Dudareva, N.; Murfitt, L.M.; Mann, C.J.; Gorenstein, N.; Kolosova, N.; Kish, C.M.; Bonham, C.; Wood, K. Developmental regulation of methyl benzoate biosynthesis and emission in snapdragon flowers. Plant Cell 2000, 12, 949–961. [Google Scholar] [CrossRef] [PubMed]
- Verdonk, J.C.; Ric de Vos, C.H.; Verhoeven, H.A.; Haring, M.A.; van Tunen, A.J.; Schuurink, R.C.; de Vos, C.H.R. Regulation of floral scent production in petunia revealed by targeted metabolomics. Phytochemistry 2003, 62, 997–1008. [Google Scholar] [CrossRef]
- Hendel-Rahmanim, K.; Masci, T.; Vainstein, A.; Weiss, D. Diurnal regulation of scent emission in rose flowers. Planta 2007, 226, 1491–1499. [Google Scholar] [CrossRef]
- Ruíz-Ramón, F.; Águila, D.J.; Egea-Cortines, M.; Weiss, J. Optimization of fragrance extraction: Daytime and flower age affect scent emission in simple and double narcissi. Ind. Crops Prod. 2014, 52, 671–678. [Google Scholar] [CrossRef]
- Yon, F.; Joo, Y.; Cort, L.; Rothe, E.; Baldwin, I.T.; Kim, S.; Kim, S. Silencing Nicotiana attenuata LHY and ZTL alters circadian rhythms in flowers. New Phytol. 2015, 203, 1058–1066. [Google Scholar] [CrossRef]
- Fenske, M.P.; Hewett Hazelton, K.D.; Hempton, A.K.; Shim, J.S.; Yamamoto, B.M.; Riffell, J.A.; Imaizumi, T. Circadian clock gene LATE ELONGATED HYPOCOTYL directly regulates the timing of floral scent emission in Petunia. Proc. Natl. Acad. Sci. USA 2015, 112, 9775–9780. [Google Scholar] [CrossRef] [Green Version]
- Bombarely, A.; Moser, M.; Amrad, A.; Bao, M.; Bapaume, L.; Barry, C.C.S.; Bliek, M.; Boersma, M.R.M.; Borghi, L.; Bruggmann, R.; et al. Insight into the evolution of the Solanaceae from the parental genomes of Petunia hybrida. Nat. Plants 2016, 2, 1–9. [Google Scholar] [CrossRef]
- Birney, E.; Clamp, M.; Durbin, R. GeneWise and Genomewise. Genome Res. 2004, 14, 988–995. [Google Scholar] [CrossRef] [Green Version]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [PubMed]
- Paradis, E.; Claude, J.; Strimmer, K. APE: Analyses of Phylogenetics and Evolution in R language. Bioinformatics 2004, 20, 289–290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schliep, K.P. Phangorn: Phylogenetic analysis in R. Bioinformatics 2011, 27, 592–593. [Google Scholar] [CrossRef]
- Yu, G.; Smith, D.K.; Zhu, H.; Guan, Y.; Lam, T.T.-Y. Ggtree: An r package for visualization and annotation of phylogenetic trees with their covariates and other associated data. Methods Ecol. Evol. 2017, 8, 28–36. [Google Scholar] [CrossRef]
- Helliwell, C.; Waterhouse, P. Constructs and methods for high-throughput gene silencing in plants. Methods 2003, 30, 289–295. [Google Scholar] [CrossRef]
- Hilson, P.; Allemeersch, J.; Altmann, T.; Aubourg, S.; Avon, A.; Beynon, J.; Bhalerao, R.P.; Bitton, F.; Caboche, M.; Cannoot, B.; et al. Versatile gene-specific sequence tags for Arabidopsis functional genomics: Trancript profiling and reverse genetics applications. Genome Res. 2004, 14, 26–2189. [Google Scholar] [CrossRef] [PubMed]
- Box, M.S.; Coustham, V.; Dean, C.; Mylne, J.S. Protocol: A simple phenol-based method for 96-well extraction of high quality RNA from Arabidopsis. Plant Methods 2011, 7, 7. [Google Scholar] [CrossRef]
- Mallona, I.; Lischewsky, S.; Weiss, J.; Hause, B.; Egea-Cortines, M. Validation of reference genes for quantitative real-time PCR during leaf and flower development in Petunia hybrida. BMC Plant Biol. 2010, 10, 4. [Google Scholar] [CrossRef] [PubMed]
- Pfaffl, M.W.; Tichopad, A.; Prgomet, C.; Neuvians, T.P. Determination of stable housekeeping genes, differentially regulated target genes and sample integrity: BestKeeper—Excel-based tool using pair-wise correlations. Biotechnol. Lett. 2004, 26, 509–515. [Google Scholar] [CrossRef] [PubMed]
- Andersen, C.L.; Jensen, J.L.; Orntoft, T.F. Normalization of real-time quantitative reverse transcription-PCR data: A model-based variance estimation approach to identify genes suited for normalization, applied to bladder and colon cancer data sets. Cancer Res. 2004, 64, 5245–5250. [Google Scholar] [CrossRef]
- Vandesompele, J.; De Preter, K.; Pattyn, F.; Poppe, B.; Van Roy, N.; De Paepe, A.; Speleman, F. Accurate normalization of realtime quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 2002, 3, RESEARCH0034. [Google Scholar] [CrossRef] [PubMed]
- Silver, N.; Best, S.; Jiang, J.; Thein, S.L. Selection of housekeeping genes for gene expression studies in human reticulocytes using real-time PCR. BMC Mol. Biol. 2006, 7, 33. [Google Scholar] [CrossRef] [PubMed]
- Xie, F.; Xiao, P.; Chen, D.; Xu, L.; Zhang, B. miRDeepFinder: A miRNA analysis tool for deep sequencing of plant small RNAs. Plant Mol. Biol. 2012, 80, 75–84. [Google Scholar] [CrossRef] [PubMed]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative CT method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef]
- Mallona, I.; Weiss, J.; Egea-Cortines, M. pcrEfficiency: A Web tool for PCR amplification efficiency prediction. BMC Bioinform. 2011, 12, 404. [Google Scholar] [CrossRef]
- Navarro, P.J.; Pérez, F.; Weiss, J.; Egea-Cortines, M. Machine learning and computer vision system for phenotype data acquisition and analysis in plants. Sensors 2016, 16, 641. [Google Scholar] [CrossRef]
- Kahm, M.; Hasenbrink, G.; Ludwig, J. Grofit: Fitting Biological Growth Curves with R. J. Stat. Softw. 2010, 33, 1–21. [Google Scholar] [CrossRef]
- Ruiz-Hernández, V.; Roca, M.J.; Egea-Cortines, M.; Weiss, J. A comparison of semi-quantitative methods suitable for establishing volatile profiles. Plant Methods 2018, 14, 67. [Google Scholar] [CrossRef] [PubMed]
- Hughes, M.E.; Hogenesch, J.B.; Kornacker, K. JTK_CYCLE: An efficient nonparametric algorithm for detecting rhythmic components in genome-scale data sets. J. Biol. Rhythm. 2010, 25, 372–380. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.; Anafi, R.C.; Hughes, M.E.; Kornacker, K.; Hogenesch, J.B. MetaCycle: An integrated R package to evaluate periodicity in large scale data. Bioinformatics 2016, 32, 3351–3353. [Google Scholar] [CrossRef]
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis; Springer: Berlin/Heidelberg, Germany, 2016; ISBN 978-3-319-24277-4. [Google Scholar]
- Garnier, S.; Ross, N.; Rudis, B.; Sciaini, M.; Scherer, C. Viridis: Default Color Maps from “Matplotlib”; R Project: Vienna, Austria, 2018; R Package Version 0.5; Available online: https://cran.r-project.org/web/packages/viridis/index.html (accessed on 11 April 2019).
- Kubota, A.; Kita, S.; Ishizaki, K.; Nishihama, R.; Yamato, K.T.; Kohchi, T. Co-option of a photoperiodic growth-phase transition system during land plant evolution. Nat. Commun. 2014, 5, 3668. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yon, F.; Seo, P.-J.; Ryu, J.Y.; Park, C.-M.; Baldwin, I.T.; Kim, S.-G. Identification and characterization of circadian clock genes in a native tobacco, Nicotiana attenuata. BMC Plant Biol. 2012, 12, 172. [Google Scholar] [CrossRef] [PubMed]
- Somers, D.E.; Schultz, T.F.; Milnamow, M.; Kay, S.A. ZEITLUPE encodes a novel clock-associated PAS protein from Arabidopsis. Cell 2000, 101, 319–329. [Google Scholar] [CrossRef]
- Navarro, P.J.; Fernández, C.; Weiss, J.; Egea-Cortines, M. Development of a configurable growth chamber with a vision system to study circadian rhythm in plants. Sensors 2012, 12, 15356–15375. [Google Scholar] [CrossRef] [PubMed]
- Nusinow, D.A.; Helfer, A.; Hamilton, E.E.; King, J.J.; Imaizumi, T.; Schultz, T.F.; Farré, E.M.; Kay, S.A.; Farre, E.M. The ELF4-ELF3-LUX complex links the circadian clock to diurnal control of hypocotyl growth. Nature 2011, 475, 398. [Google Scholar] [CrossRef] [PubMed]
- Amrad, A.; Moser, M.; Mandel, T.; de Vries, M.; Schuurink, R.C.; Freitas, L.; Kuhlemeier, C. Gain and Loss of Floral Scent Production through Changes in Structural Genes during Pollinator-Mediated Speciation. Curr. Biol. 2016, 26, 3303–3312. [Google Scholar] [CrossRef] [PubMed]
- Endo, M. Tissue-specific circadian clocks in plants. Curr. Opin. Plant Biol. 2016, 29, 44–49. [Google Scholar] [CrossRef] [Green Version]
- Weiss, J.; Terry, M.I.; Martos-Fuentes, M.; Letourneux, L.; Ruiz-hernández, V.; Fernández, J.A.; Egea-cortines, M. Diel pattern of circadian clock and storage protein gene expression in leaves and during seed filling in cowpea (Vigna unguiculata). BMC Plant Biol. 2018, 18, 33–53. [Google Scholar] [CrossRef]
- De Mairan, J.-J. Observation Botanique; Histoire de l’Académie Royale des Sciences Paris; Institut de France: Paris, France, 1729; p. 35. [Google Scholar]
- Ku, L.; Wei, X.; Zhang, S.; Zhang, J.; Guo, S.; Chen, Y. Cloning and Characterization of a Putative TAC1 Ortholog Associated with Leaf Angle in Maize (Zea mays L.). PLoS ONE 2011, 6, e20621. [Google Scholar] [CrossRef] [PubMed]
- Kao, W.-Y.; Tsai, T.-T. Tropic leaf movements, photosynthetic gas exchange, leaf δ13C and chlorophyll a fluorescence of three soybean species in response to water availability. Plant Cell Environ. 1998, 21, 1055–1062. [Google Scholar] [CrossRef] [Green Version]
- Park, D.H. Control of Circadian Rhythms and Photoperiodic Flowering by the Arabidopsis GIGANTEA Gene. Science 1999, 285, 1579–1582. [Google Scholar] [CrossRef] [PubMed]
- Doyle, M.R.; Davis, S.J.; Bastow, R.M.; McWatters, H.G.; Kozma-Bognar, L.; Nagy, F.; Millar, A.J.; Amasino, R.M. The ELF4 gene controls circadian rhythms and flowering time in Arabidopsis thaliana. Nature 2002, 419, 74–77. [Google Scholar] [CrossRef] [PubMed]
- Kennaway, R.; Coen, E.; Green, A.; Bangham, A. Generation of diverse biological forms through combinatorial interactions between tissue polarity and growth. PLoS Comput. Biol. 2011, 7, e1002071. [Google Scholar] [CrossRef]
- Beemster, G.T.S.; De Vusser, K.; De Tavernier, E.; De Bock, K.; Inze, D. Variation in growth rate between Arabidopsis ecotypes is correlated with cell division and A-type cyclin-dependent kinase activity. Plant Physiol. 2002, 129, 854–864. [Google Scholar] [CrossRef]
- Feys, K.; Demuynck, K.; De Block, J.; Bisht, A.; De Vliegher, A.; Inzé, D.; Nelissen, H. Growth rate rather than growth duration drives growth heterosis in maize B104 hybrids. Plant Cell Environ. 2017, 41, 374–382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thain, S.C.; Murtas, G.; Lynn, J.R.; McGrath, R.B.; Millar, A.J. The circadian clock that controls gene expression in Arabidopsis is tissue specific. Plant Physiol. 2002, 130, 102–110. [Google Scholar] [CrossRef] [PubMed]
- Bordage, S.; Sullivan, S.; Laird, J.; Millar, A.J.; Nimmo, H.G. Organ specificity in the plant circadian system is explained by different light inputs to the shoot and root clocks. New Phytol. 2016, 212, 136–149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Voß, U.; Wilson, M.H.; Kenobi, K.; Gould, P.D.; Robertson, F.C.; Peer, W.A.; Lucas, M.; Swarup, K.; Casimiro, I.; Holman, T.J.; et al. The circadian clock rephases during lateral root organ initiation in Arabidopsis thaliana. Nat. Commun. 2015, 6, 7641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farré, E.M. The regulation of plant growth by the circadian clock. Plant Biol. 2012, 14, 401–410. [Google Scholar] [CrossRef]
- Fung-Uceda, J.; Lee, K.; Seo, P.J.; Polyn, S.; De Veylder, L.; Mas, P. The Circadian Clock Sets the Time of DNA Replication Licensing to Regulate Growth in Arabidopsis. Dev. Cell 2018, 45, 101–113. [Google Scholar] [CrossRef] [PubMed]
- Hoballah, M.E.; Stuurman, J.; Turlings, T.C.J.; Guerin, P.M.; Connétable, S.; Kuhlemeier, C.; Connetable, S.; Kuhlemeier, C. The composition and timing of flower odour emission by wild Petunia axillaris coincide with the antennal perception and nocturnal activity of the pollinator Manduca sexta. Planta 2005, 222, 141–150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Unsicker, S.B.; Kunert, G.; Gershenzon, J. Protective perfumes: The role of vegetative volatiles in plant defense against herbivores. Curr. Opin. Plant Biol. 2009, 12, 479–485. [Google Scholar] [CrossRef]
- Pichersky, E.; Gershenzon, J. The formation and function of plant volatiles: Perfumes for pollinator attraction and defense. Curr. Opin. Plant Biol. 2002, 5, 237–243. [Google Scholar] [CrossRef]
- Junker, R.R.; Blüthgen, N. Floral scents repel potentially nectar-thieving ants. Evol. Ecol. Res. 2008, 10, 295–308. [Google Scholar]
- Aharoni, A. Terpenoid Metabolism in Wild-Type and Transgenic Arabidopsis Plants. Plant Cell Online 2003, 15, 2866–2884. [Google Scholar] [CrossRef] [PubMed]
- Yon, F.; Kessler, D.; Joo, Y.; Cortés Llorca, L.; Kim, S.G.; Baldwin, I.T. Fitness consequences of altering floral circadian oscillations for Nicotiana attenuata. J. Integr. Plant Biol. 2017, 59, 180–189. [Google Scholar] [CrossRef] [PubMed]
- Bhatia, V.; Maisnam, J.; Jain, A.; Sharma, K.K.; Bhattacharya, R. Aphid-repellent pheromone E-β-farnesene is generated in transgenic Arabidopsis thaliana over-expressing farnesyl diphosphate synthase2. Ann. Bot. 2015, 115, 581–591. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Measure | RNAi:PhCHL | W115 | p-Value | ||
|---|---|---|---|---|---|
| Line 3 | Line 10 | RNAi:PhCHL3 | RNAi:PhCHL10 | ||
| Plant height | 25.1 ± 1.89 | 21.7 ± 4.18 | 37.33 ± 3.0 | 0.07 | 0.022 * |
| Floral limb | 4.37 ± 0.44 | 4.76 ± 0.55 | 5.38 ± 0.29 | 0.00009 *** | 0.0038 ** |
| Floral tube | 4.07 ± 0.37 | 4.81 ± 0.51 | 4.82 ± 0.35 | 0.00055 *** | NS |
| Chlorophyll | 39.43 ± 6.32 | 36.65 ± 9.77 | 39.30 ± 9.80 | NS | NS |
| Group | Rate | Max.Growth/Std.Error | Max.Slope/Std.Error | Area under Model |
|---|---|---|---|---|
| WT | Stem growth | 16.084/0.142 | 0.121/0.005 | 1778.3 |
| RNAi:PhCHL | Stem growth | 12.984/0.096 | 0.118/0.006 | 1471.53 |
| WT | Flower growth | 57.698/0.403 | 0.798/0.013 | 3598.88 |
| RNAi:PhCHL | Flower growth | 39.677/0.344 | 0.566/0.011 | 2328.95 |
| RT | Name | CAS |
|---|---|---|
| 4.873 | Benzaldehyde | 100-52-7 |
| 6.435 | Limonene | 138-86-3 |
| 6.539 | Benzyl alcohol | 100-51-6 |
| 6.735 | Phenylacetaldehyde | 122-78-1 |
| 7.230 | Acetophenone | 98-86-2 |
| 7.825 | Methyl benzoate | 93-58-3 |
| 8.149 | Phenylethyl alcohol | 60-12-8 |
| 9.125 | Benzyl acetate | 140-11-4 |
| 9.244 | Ethyl benzoate | 93-89-0 |
| 9.668 | Methyl salicylate | 119-36-8 |
| 10.720 | Phenylethyl acetate | 103-45-7 |
| 12.287 | Eugenol | 97-53-0 |
| 12.711 | Benzyl 2-methylbutyrate | 56423-40-6 |
| 13.587 | Isoeugenol (isomers) | 97-45-1; 5932-68-3 |
| 14.254 | Benzyl tiglate | 37526-88-8 |
| 14.625 | Cadinene | 483-76-1 |
| 17.525 | Benzyl benzoate | 120-51-4 |
| Volatile | W115 p Value | Phase | PhCHL3 p Value | Phase | PhCHL10 p Value | Phase |
|---|---|---|---|---|---|---|
| Benzyl tiglate | 1.000 | 2 | 0.014 | 0.5 | 0.476 | 5 |
| Cadinene | 0.445 | 5 | 0.246 | 23 | 1.000 | 9.5 |
| Ethyl benzoate | 1.000 | 11 | 1.000 | 20 | 1.000 | 20 |
| Acetophenone | 1.000 | 12.5 | 1.000 | 8 | 0.210 | 11 |
| Benzaldehyde | 0.000 | 15.5 | 0.000 | 12.5 | 0.000 | 15.5 |
| Benzyl benzoate | 0.001 | 15.5 | 0.023 | 17 | 0.001 | 17 |
| Limonene | 0.165 | 15.5 | 0.280 | 17 | 1.000 | 11 |
| Phenylacetaldehyde | 0.001 | 15.5 | 0.000 | 12.5 | 0.000 | 20 |
| Methyl benzoate | 0.017 | 17 | 0.003 | 15.5 | 0.011 | 17 |
| Isoeugenol | 0.001 | 18.5 | 0.007 | 18.5 | 0.014 | 17 |
| Benzyl acetate | 0.003 | 20 | 0.001 | 20 | 0.005 | 20 |
| Eugenol | 0.001 | 20 | 0.000 | 20 | 0.019 | 20 |
| Methyl salicylate | 1.000 | 20 | 0.009 | 20 | 1.000 | 15.5 |
| Phenylethyl acetate | 0.048 | 20 | 0.002 | 20 | 0.000 | 20 |
| Phenylethyl alcohol | 0.000 | 20 | 0.000 | 18.5 | 0.000 | 15.5 |
| Benzyl alcohol | 0.000 | 21.5 | 0.007 | 20 | 0.000 | 20 |
| Benzyl 2-methylbutyrate | 1.000 | 23 | 0.011 | 23 | 0.194 | 2 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Terry, M.I.; Pérez-Sanz, F.; Díaz-Galián, M.V.; Pérez de los Cobos, F.; Navarro, P.J.; Egea-Cortines, M.; Weiss, J. The Petunia CHANEL Gene is a ZEITLUPE Ortholog Coordinating Growth and Scent Profiles. Cells 2019, 8, 343. https://doi.org/10.3390/cells8040343
Terry MI, Pérez-Sanz F, Díaz-Galián MV, Pérez de los Cobos F, Navarro PJ, Egea-Cortines M, Weiss J. The Petunia CHANEL Gene is a ZEITLUPE Ortholog Coordinating Growth and Scent Profiles. Cells. 2019; 8(4):343. https://doi.org/10.3390/cells8040343
Chicago/Turabian StyleTerry, Marta I., Fernando Pérez-Sanz, M. Victoria Díaz-Galián, Felipe Pérez de los Cobos, Pedro J. Navarro, Marcos Egea-Cortines, and Julia Weiss. 2019. "The Petunia CHANEL Gene is a ZEITLUPE Ortholog Coordinating Growth and Scent Profiles" Cells 8, no. 4: 343. https://doi.org/10.3390/cells8040343