
Forced Swimming-Induced Depressive-like Behavior and Anxiety Are Reduced by Chlorpheniramine via Suppression of Oxidative and Inflammatory Mediators and Activating the Nrf2-BDNF Signaling Pathway

 , , , , ,
, , , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
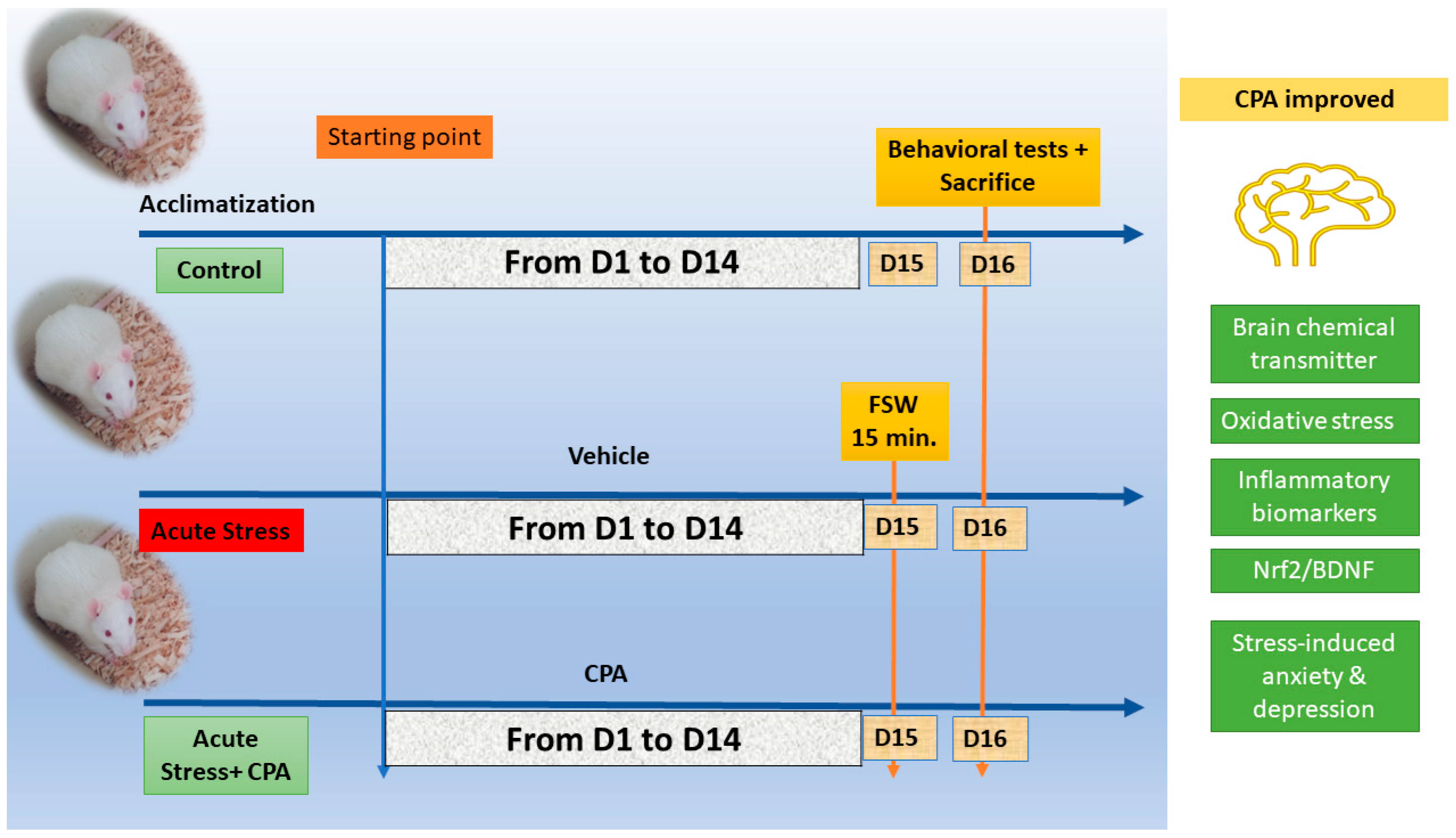
2.2. Experimental Design
2.3. Behavioral Tests
2.3.1. Forced Swim Test (FST) and Immobility Time
2.3.2. Open Field Test
2.3.3. Elevated Plus Maze (EPM) Test
2.4. Biochemical Analysis
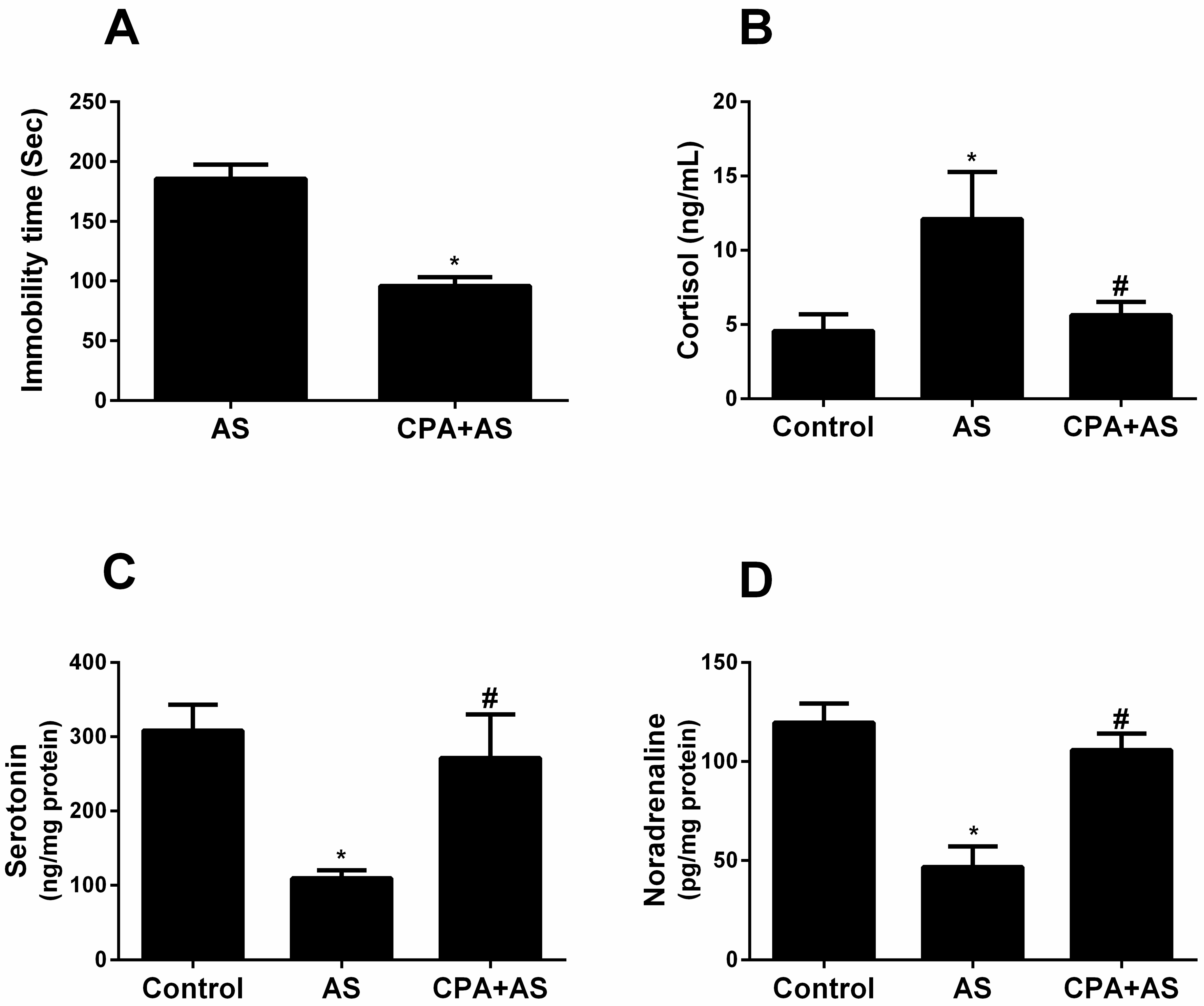
2.4.1. Estimation of Serum Cortisol Levels
2.4.2. Estimation of Serotonin and Noradrenaline Levels
2.4.3. Real-Time Polymerase Chain Reaction (RT-PCR) for Estimation of Gene Expression of BDNF and Nrf2
2.4.4. Estimation of Biomarkers the Oxidative Stress Malondialdehyde (MDA) Levels and Superoxide Dismutase (SOD) and the Pro-Inflammatory Biomarkers IL-1 β and IL-6 in Brain Tissues
2.5. Statistical Analysis
3. Results
3.1. CPA Protects against Stress-Induced Depressive-like Behavior
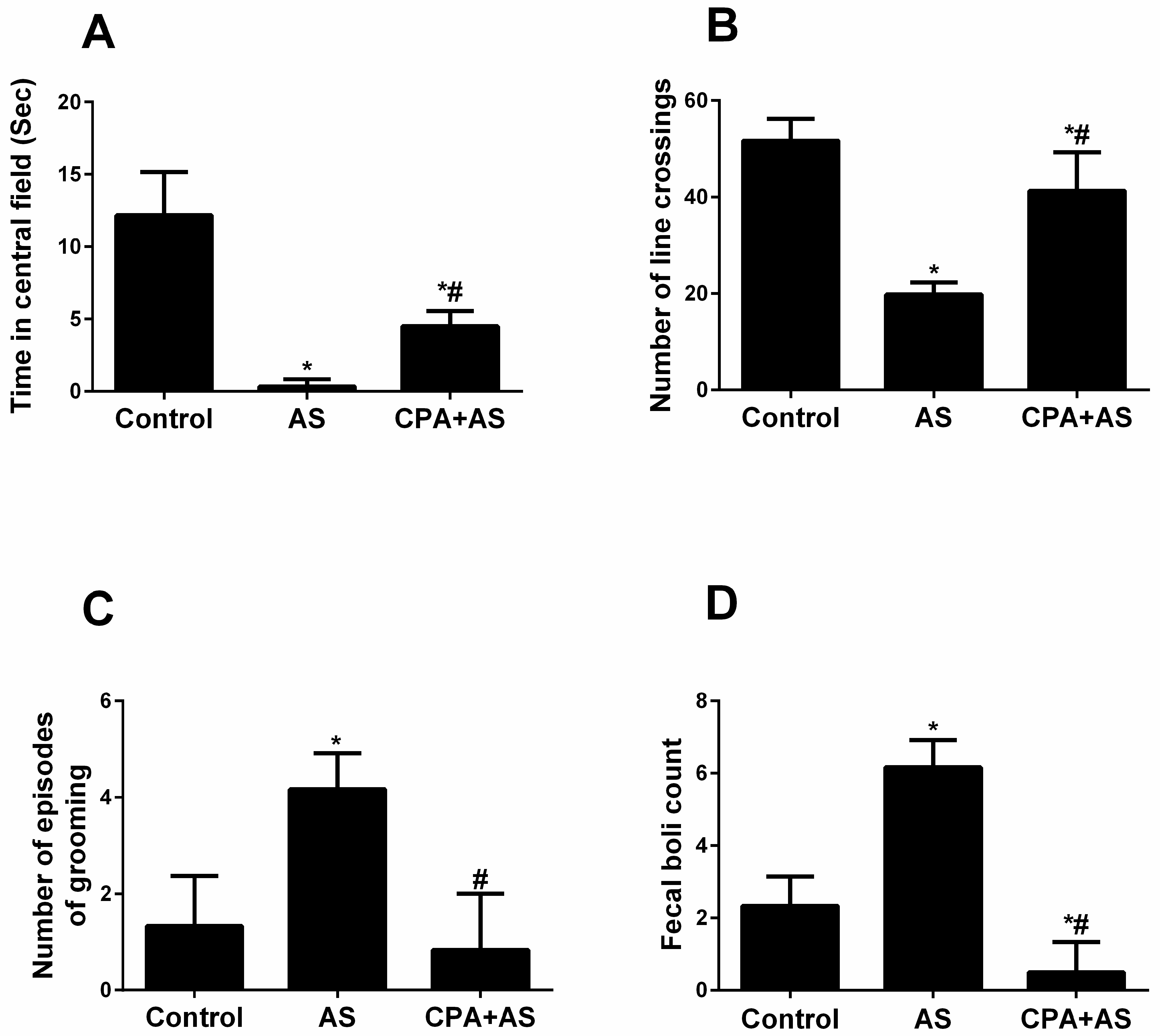
3.2. CPA Protects against Stress-Induced Anxiety Using an Open Field Test
3.2.1. Time Spent in the Field’s Centre
3.2.2. Number of Line Crossings
3.2.3. Number of Grooming Episodes
3.2.4. Fecal Boli Count
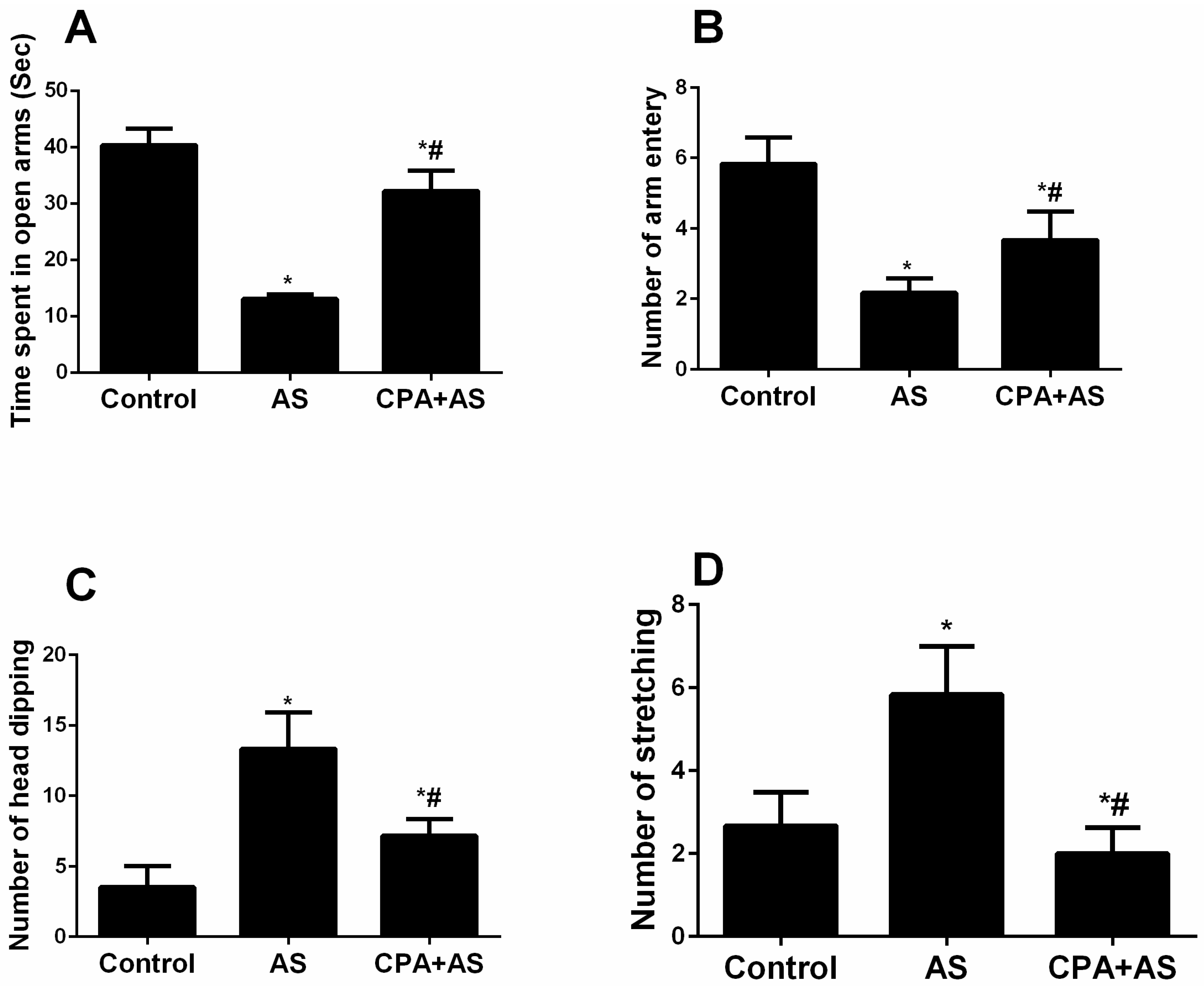
3.3. CPA Protects against Stress-Induced Anxiety Using an Elevated Plus Maize Test (EPMT)
3.3.1. Time Spent in the Open Arms
3.3.2. Number of Arm Entries
3.3.3. Number of Head Dipping
3.3.4. Stretched-Attend Postures
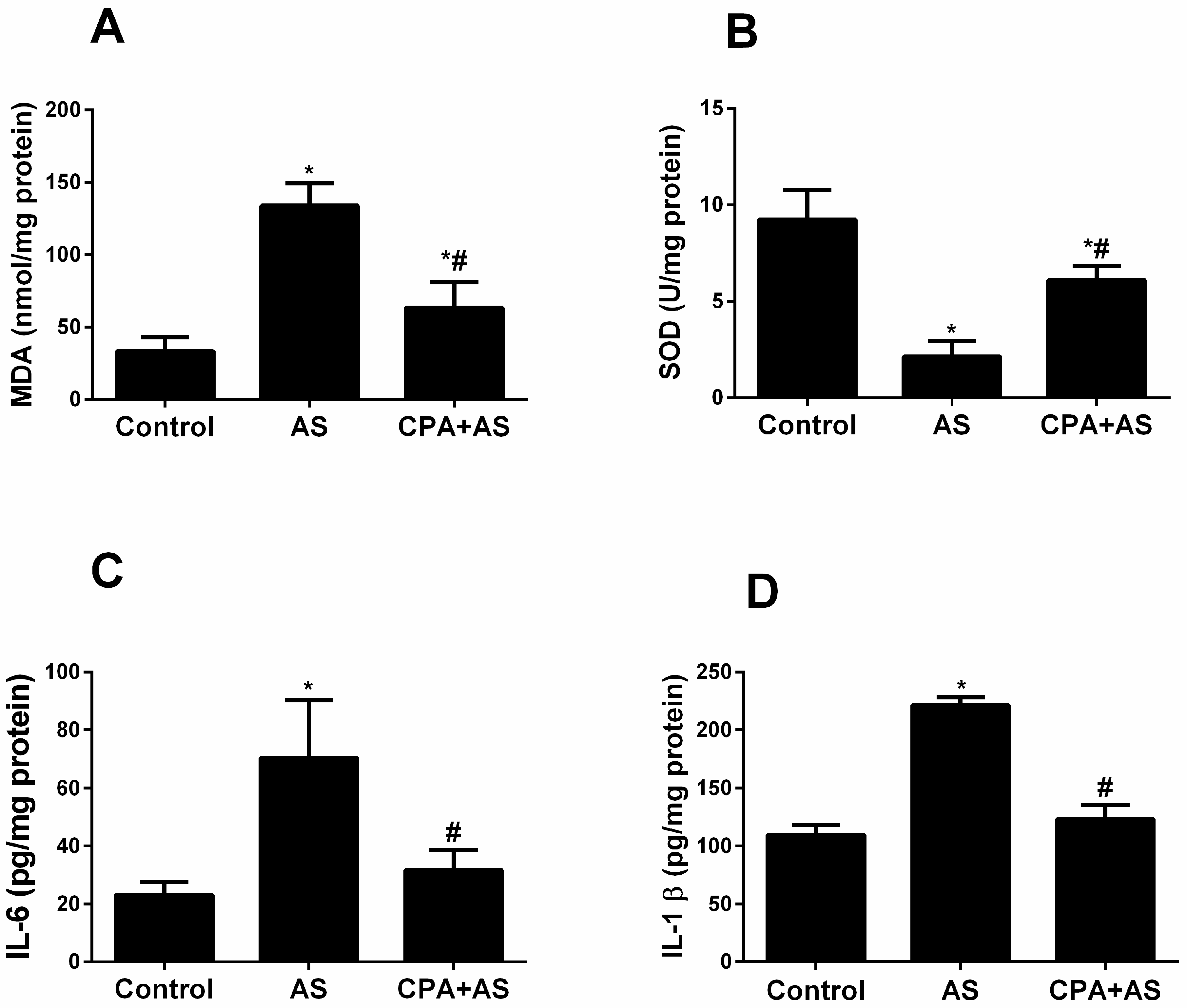
3.4. CPA Attenuated Stress-Induced Oxidative Stress and Inflammatory Biomarkers in Hippocampal Tissues
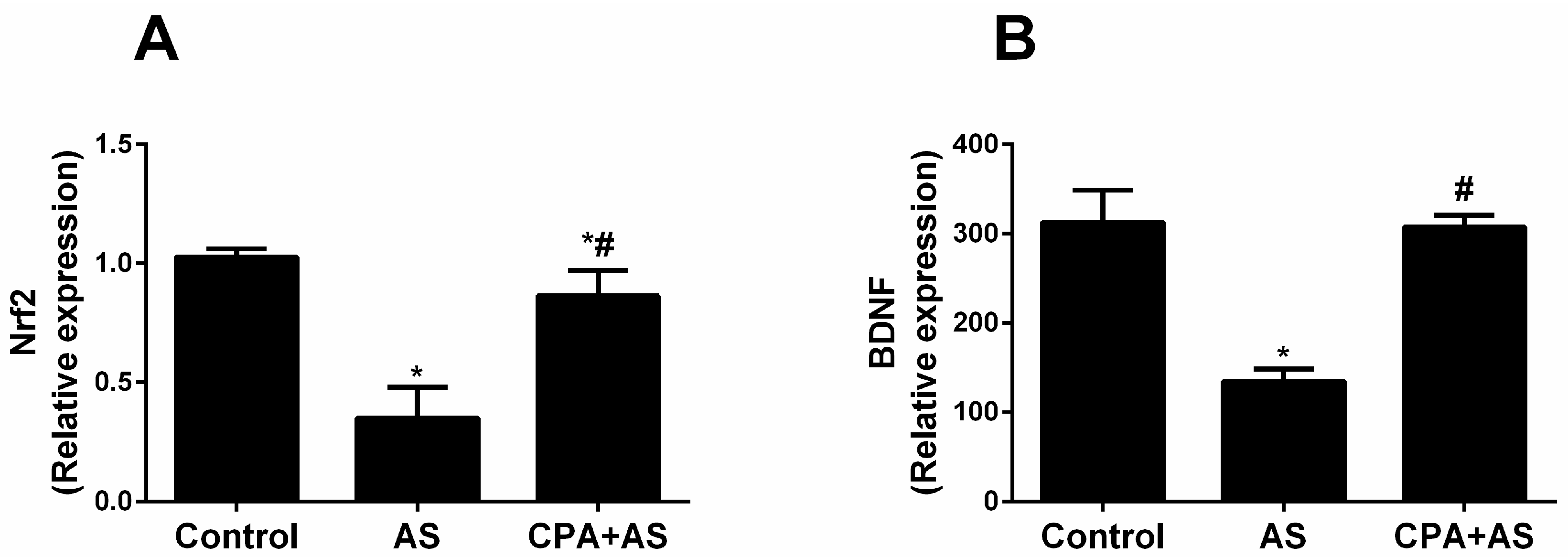
3.5. CPA Attenuated Stress-Induced Decrease in Nrf2 and BDNF in Cerebral Cortex Tissues
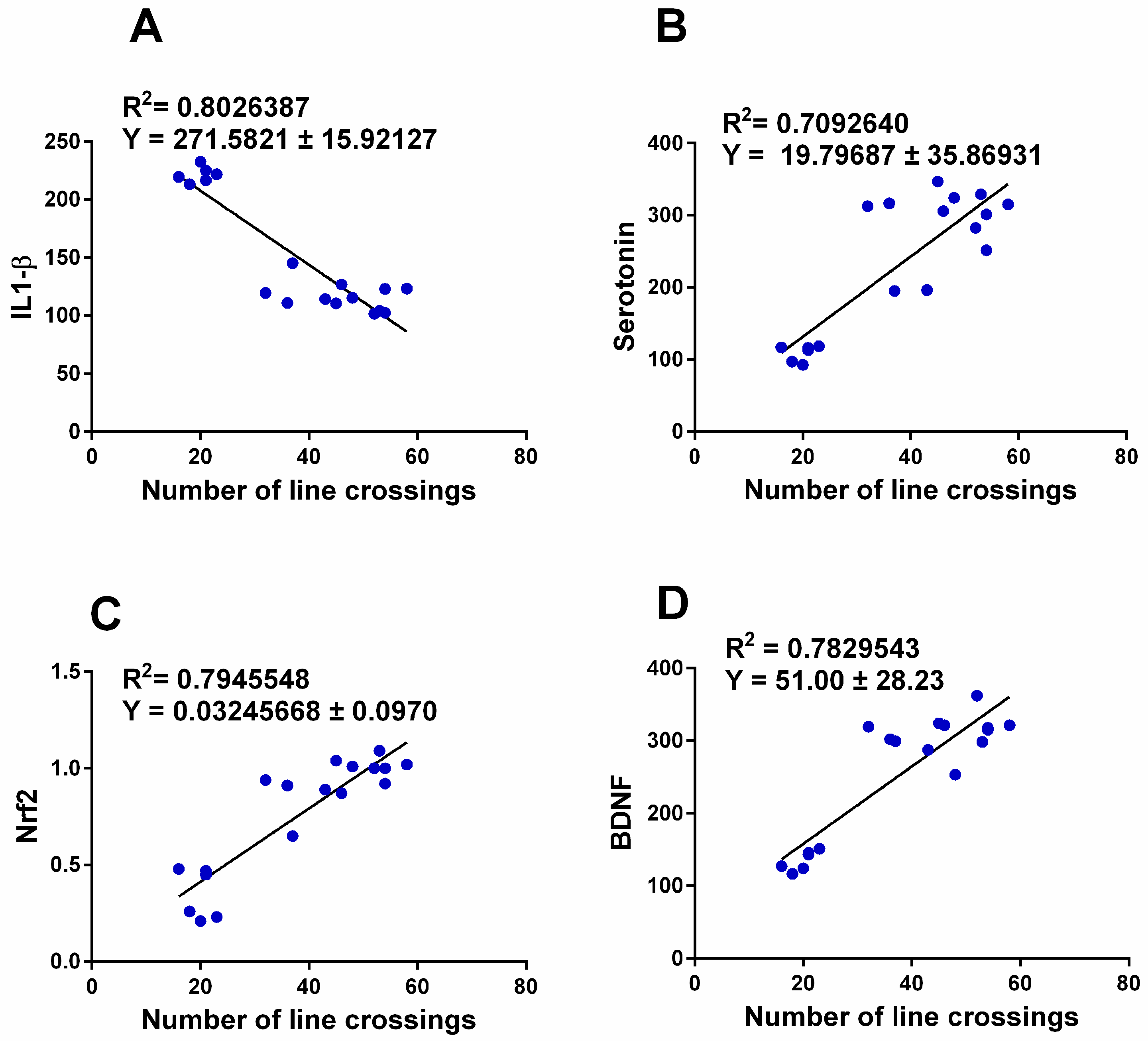
3.6. Correlation between Anxiety-Related Behavior and Changes in Brain Transmitters, Inflammatory Biomarkers, and Nrf2-BDNF Signaling Pathway
4. Discussion
5. Study Limitation
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Ghanem, I.; Castelo, B. Coping strategies and depressive symptoms in cancer patients. Clin. Transl. Oncol. 2020, 22, 330–336. [Google Scholar] [CrossRef] [PubMed]
- Bulos, E.M.; Pobbe, R.L.; Zangrossi, H., Jr. Behavioral consequences of predator stress in the rat elevated T-maze. Physiol. Behav. 2015, 146, 28–35. [Google Scholar] [CrossRef] [PubMed]
- Perez-Tejada, J.; Garmendia, L.; Labaka, A.; Vegas, O.; Gómez-Lazaro, E.; Arregi, A. Active and Passive Coping Strategies: Comparing Psychological Distress, Cortisol, and Proinflammatory Cytokine Levels in Breast Cancer Survivors. Clin. J. Oncol. Nurs. 2019, 23, 583–590. [Google Scholar] [CrossRef]
- Belovicova, K.; Bogi, E.; Csatlosova, K.; Dubovicky, M. Animal tests for anxiety-like and depression-like behavior in rats. Interdiscip. Toxicol. 2017, 10, 40–43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Porsolt, R.D.; Le Pichon, M.; Jalfre, M. Depression: A new animal model sensitive to antidepressant treatments. Nature 1977, 266, 730–732. [Google Scholar] [CrossRef] [PubMed]
- Commons, K.G.; Cholanians, A.B.; Babb, J.A.; Ehlinger, D.G. The Rodent Forced Swim Test Measures Stress-Coping Strategy, Not Depression-like Behavior. ACS Chem. Neurosci. 2017, 8, 955–960. [Google Scholar] [CrossRef] [Green Version]
- Rihmer, Z.; Dome, P.; Katona, C. Serotonin and depression—A riposte to Moncrieff et al. (2022). Neuropsychopharmacol. Hung. 2022, 24, 120–125. [Google Scholar]
- Chen, B.; Xu, J.; Chen, S.; Mou, T.; Wang, Y.; Wang, H.; Zhang, Z.; Ren, F.; Wang, Z.; Jin, K.; et al. Dysregulation of striatal dopamine D2/D3 receptor-mediated by hypocretin induces depressive behaviors in rats. J. Affect. Disord. 2023, 325, 256–263. [Google Scholar] [CrossRef]
- Strawn, J.R.; Mills, J.A.; Poweleit, E.A.; Ramsey, L.B.; Croarkin, P.E. Adverse Effects of Antidepressant Medications and their Management in Children and Adolescents. Pharmacotherapy 2023, 43, 675. [Google Scholar] [CrossRef]
- Cheng, Z.; Zhao, F.; Piao, J.; Cui, R.; Li, B. Rasd2 mediates acute fasting-induced antidepressant-like effects via dopamine D2 receptor activation in ovariectomized mice. Int. J. Neuropsychopharmacol. 2022, 26, 217–229. [Google Scholar] [CrossRef]
- Vedhara, K.; Hyde, J.; Gilchrist, I.D.; Tytherleigh, M.; Plummer, S. Acute stress, memory, attention and cortisol. Psychoneuroendocrinology 2000, 25, 535–549. [Google Scholar] [CrossRef] [PubMed]
- Koelsch, S.; Boehlig, A.; Hohenadel, M.; Nitsche, I.; Bauer, K.; Sack, U. The impact of acute stress on hormones and cytokines, and how their recovery is affected by music-evoked positive mood. Sci. Rep. 2016, 6, 23008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shati, A.A.; Zaki, M.S.A.; Alqahtani, Y.A.; Al-Qahtani, S.M.; Haidara, M.A.; Dawood, A.F.; AlMohanna, A.M.; El-Bidawy, M.H.; Alaa Eldeen, M.; Eid, R.A. Antioxidant Activity of Vitamin C against LPS-Induced Septic Cardiomyopathy by Down-Regulation of Oxidative Stress and Inflammation. Curr. Issues Mol. Biol. 2022, 44, 2387–2400. [Google Scholar] [CrossRef]
- Ma, Q. Transcriptional responses to oxidative stress: Pathological and toxicological implications. Pharmacol. Ther. 2010, 125, 376–393. [Google Scholar] [CrossRef]
- Agnihotri, A.; Aruoma, O.I. Alzheimer’s Disease and Parkinson’s Disease: A Nutritional Toxicology Perspective of the Impact of Oxidative Stress, Mitochondrial Dysfunction, Nutrigenomics and Environmental Chemicals. J. Am. Coll. Nutr. 2020, 39, 16–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abbah, J.; Vacher, C.M.; Goldstein, E.Z.; Li, Z.; Kundu, S.; Talbot, B.; Bhattacharya, S.; Hashimoto-Torii, K.; Wang, L.; Banerjee, P.; et al. Oxidative Stress-Induced Damage to the Developing Hippocampus Is Mediated by GSK3β. J. Neurosci. 2022, 42, 4812–4827. [Google Scholar] [CrossRef] [PubMed]
- Novochadlo, M.; Goldim, M.P.; Bonfante, S.; Joaquim, L.; Mathias, K.; Metzker, K.; Machado, R.S.; Lanzzarin, E.; Bernades, G.; Bagio, E.; et al. Folic acid alleviates the blood brain barrier permeability and oxidative stress and prevents cognitive decline in sepsis-surviving rats. Microvasc. Res. 2021, 137, 104193. [Google Scholar] [CrossRef] [PubMed]
- Savage, K.; Kingshott, D.; Gubko, A.; Thee, A.W.; Burjawi, T.; Croft, K.; Sarris, J.; Stough, C. The Relationship between Oxidative Stress and Anxiety in a Healthy Older Population. Exp. Aging Res. 2021, 47, 322–346. [Google Scholar] [CrossRef]
- Gałecki, P.; Szemraj, J.; Bieńkiewicz, M.; Zboralski, K.; Gałecka, E. Oxidative stress parameters after combined fluoxetine and acetylsalicylic acid therapy in depressive patients. Hum. Psychopharmacol. 2009, 24, 277–286. [Google Scholar] [CrossRef]
- Hussain, T.; Kandeel, M.; Metwally, E.; Murtaza, G.; Kalhoro, D.H.; Yin, Y.; Tan, B.; Chughtai, M.I.; Yaseen, A.; Afzal, A.; et al. Unraveling the harmful effect of oxidative stress on male fertility: A mechanistic insight. Front. Endocrinol. 2023, 14, 1070692. [Google Scholar] [CrossRef]
- Steptoe, A.; Hamer, M.; Chida, Y. The effects of acute psychological stress on circulating inflammatory factors in humans: A review and meta-analysis. Brain Behav. Immun. 2007, 21, 901–912. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, K.T.; Deak, T.; Owens, S.M.; Kohno, T.; Fleshner, M.; Watkins, L.R.; Maier, S.F. Exposure to acute stress induces brain interleukin-1beta protein in the rat. J. Neurosci. 1998, 18, 2239–2246. [Google Scholar] [CrossRef] [Green Version]
- Johnson, J.D.; Barnard, D.F.; Kulp, A.C.; Mehta, D.M. Neuroendocrine Regulation of Brain Cytokines After Psychological Stress. J. Endocr. Soc. 2019, 3, 1302–1320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, C.Y.; Ren, X.M.; Li, H.B.; Wei, W.; Wang, K.X.; Li, Y.M.; Hu, J.L.; Li, X. Simvastatin alleviates inflammation and oxidative stress in rats with cerebral hemorrhage through Nrf2-ARE signaling pathway. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 6321–6329. [Google Scholar] [CrossRef] [PubMed]
- Ma, Q. Xenobiotic-activated receptors: From transcription to drug metabolism to disease. Chem. Res. Toxicol. 2008, 21, 1651–1671. [Google Scholar] [CrossRef]
- Li, W.; Khor, T.O.; Xu, C.; Shen, G.; Jeong, W.S.; Yu, S.; Kong, A.N. Activation of Nrf2-antioxidant signaling attenuates NFkappaB-inflammatory response and elicits apoptosis. Biochem. Pharmacol. 2008, 76, 1485–1489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yao, W.; Lin, S.; Su, J.; Cao, Q.; Chen, Y.; Chen, J.; Zhang, Z.; Hashimoto, K.; Qi, Q.; Zhang, J.C. Activation of BDNF by transcription factor Nrf2 contributes to antidepressant-like actions in rodents. Transl. Psychiatry 2021, 11, 140. [Google Scholar] [CrossRef] [PubMed]
- Duman, R.S.; Malberg, J.; Thome, J. Neural plasticity to stress and antidepressant treatment. Biol. Psychiatry 1999, 46, 1181–1191. [Google Scholar] [CrossRef]
- Castrén, E.; Rantamäki, T. The role of BDNF and its receptors in depression and antidepressant drug action: Reactivation of developmental plasticity. Dev. Neurobiol. 2010, 70, 289–297. [Google Scholar] [CrossRef]
- Miyata, S.; Hirano, S.; Ohsawa, M.; Kamei, J. Chlorpheniramine exerts anxiolytic-like effects and activates prefrontal 5-HT systems in mice. Psychopharmacology 2011, 213, 441–452. [Google Scholar] [CrossRef]
- Gammoh, O.; Mayyas, F.; Darwish Elhajji, F. Chlorpheniramine and escitalopram: Similar antidepressant and nitric oxide lowering roles in a mouse model of anxiety. Biomed. Rep. 2017, 6, 675–680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bloomfield, M.A.; McCutcheon, R.A.; Kempton, M.; Freeman, T.P.; Howes, O. The effects of psychosocial stress on dopaminergic function and the acute stress response. eLife 2019, 8, e46797. [Google Scholar] [CrossRef]
- Karamanakos, P.N.; Pappas, P.; Marselos, M. Involvement of the brain serotonergic system in the locomotor stimulant effects of chlorpheniramine in Wistar rats: Implication of postsynaptic 5-HT1A receptors. Behav. Brain Res. 2004, 148, 199–208. [Google Scholar] [CrossRef] [PubMed]
- Lazarus, B.; Wu, A.; Shin, J.I.; Sang, Y.; Alexander, G.C.; Secora, A.; Inker, L.A.; Coresh, J.; Chang, A.R.; Grams, M.E. Association of Metformin Use With Risk of Lactic Acidosis Across the Range of Kidney Function: A Community-Based Cohort Study. JAMA Intern. Med. 2018, 178, 903–910. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yankelevitch-Yahav, R.; Franko, M.; Huly, A.; Doron, R. The forced swim test as a model of depressive-like behavior. J. Vis. Exp. 2015, 97, e52587. [Google Scholar] [CrossRef] [Green Version]
- Yavuz, Y.; Oyku Ozen, D.; Yagmur Erol, Z.; Goren, H.; Yilmaz, B. Effects of endocrine disruptors on the electrical activity of leptin receptor neurons in the dorsomedial hypothalamus and anxiety-like behavior in male mice. Environ. Pollut. 2023, 324, 121366. [Google Scholar] [CrossRef]
- Estaphan, S.; Curpăn, A.S.; Khalifa, D.; Rashed, L.; Ciobica, A.; Cantemir, A.; Ciobica, A.; Trus, C.; Ali, M.; ShamsEldeen, A. Combined Low Dose of Ketamine and Social Isolation: A Possible Model of Induced Chronic Schizophrenia-Like Symptoms in Male Albino Rats. Brain Sci. 2021, 11, 917. [Google Scholar] [CrossRef]
- Zargar, S.; Alonazi, M.; Rizwana, H.; Wani, T.A. Resveratrol Reverses Thioacetamide-Induced Renal Assault with respect to Oxidative Stress, Renal Function, DNA Damage, and Cytokine Release in Wistar Rats. Oxid. Med. Cell Longev. 2019, 2019, 1702959. [Google Scholar] [CrossRef] [Green Version]
- Thivisol, U.; Binder, M.D.; Hannan, A.J.; Pang, T.Y. Loss of Tyro3 causes anxiety-relevant behavioural changes in female mice. Brain Res. 2023, 1807, 148319. [Google Scholar] [CrossRef]
- Cryan, J.F.; Valentino, R.J.; Lucki, I. Assessing substrates underlying the behavioral effects of antidepressants using the modified rat forced swimming test. Neurosci. Biobehav. Rev. 2005, 29, 547–569. [Google Scholar] [CrossRef]
- Chang, M.; Zhang, L.; Dai, H.; Sun, L. Genistein acts as antidepressant agent against chronic mild stress-induced depression model of rats through augmentation of brain-derived neurotrophic factor. Brain Behav. 2021, 11, e2300. [Google Scholar] [CrossRef]
- Amin, F.; Ibrahim, M.A.A.; Rizwan-Ul-Hasan, S.; Khaliq, S.; Gabr, G.A.; Muhammad; Khan, A.; Sidhom, P.A.; Tikmani, P.; Shawky, A.M.; et al. Interactions of Apigenin and Safranal with the 5HT1A and 5HT2A Receptors and Behavioral Effects in Depression and Anxiety: A Molecular Docking, Lipid-Mediated Molecular Dynamics, and In Vivo Analysis. Molecules 2022, 27, 8658. [Google Scholar] [CrossRef] [PubMed]
- Serafim, K.R.; Kishi, M.S.; Canto-de-Souza, A.; Mattioli, R. H₁ but not H₂ histamine antagonist receptors mediate anxiety-related behaviors and emotional memory deficit in mice subjected to elevated plus-maze testing. Braz. J. Med. Biol. Res. 2013, 46, 440–446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palotai, M.; Telegdy, G.; Tanaka, M.; Bagosi, Z.; Jászberényi, M. Neuropeptide AF induces anxiety-like and antidepressant-like behavior in mice. Behav. Brain Res. 2014, 274, 264–269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brito, A.F.; Braga, P.; Moreira, L.K.S.; Silva, D.M.; Silva, D.P.B.; Sanz, G.; Vaz, B.G.; de Carvalho, F.S.; Lião, L.M.; Silva, R.R.; et al. A new piperazine derivative: 1-(4-(3,5-di-tert-butyl-4-hydroxybenzyl) piperazin-1-yl)-2-methoxyethan-1-one with antioxidant and central activity. Naunyn Schmiedebergs Arch. Pharmacol. 2018, 391, 255–269. [Google Scholar] [CrossRef]
- Almeida, L.S.; Santana, I.G.C.; da Silva Moreira, L.K.; Turones, L.C.; Sanz, G.; Vaz, B.G.; de Carvalho, F.S.; Lião, L.M.; Menegatti, R.; Costa, E.A.; et al. Neuropharmacological Activity of the New Piperazine Derivative 2-(4-((1-Phenyl-1H-Pyrazol-4-yl)Methyl)Piperazin-1-yl)Ethyl Acetate is Modulated by Serotonergic and GABAergic Pathways. CNS Neurol. Disord. Drug Targets 2022, 21, 520–532. [Google Scholar] [CrossRef]
- Gammoh, O.S.; Al-Smadi, A.; Al-Awaida, W.; Badr, M.M.; Qinna, N.A. Increased Salivary Nitric Oxide and G6PD Activity in Refugees with Anxiety and Stress. Stress Health 2016, 32, 435–440. [Google Scholar] [CrossRef]
- Machawal, L.; Kumar, A. Possible involvement of nitric oxide mechanism in the neuroprotective effect of rutin against immobilization stress induced anxiety like behaviour, oxidative damage in mice. Pharmacol. Rep. 2014, 66, 15–21. [Google Scholar] [CrossRef]
- Kala, M.; Nivsarkar, M. Role of cortisol and superoxide dismutase in psychological stress induced anovulation. Gen. Comp. Endocrinol. 2016, 225, 117–124. [Google Scholar] [CrossRef]
- Biala, G.; Pekala, K.; Boguszewska-Czubara, A.; Michalak, A.; Kruk-Slomka, M.; Budzynska, B. Behavioral and Biochemical Interaction Between Nicotine and Chronic Unpredictable Mild Stress in Mice. Mol. Neurobiol. 2017, 54, 904–921. [Google Scholar] [CrossRef] [Green Version]
- Misra, U.K.; Singh, S.K.; Kalita, J.; Kumar, A. Astrocyte activation following nitrous oxide exposure is related to oxidative stress and glutamate excitotoxicity. Brain Res. 2020, 1730, 146645. [Google Scholar] [CrossRef] [PubMed]
- Shaban, N.Z.; Mohammed, A.S.; Abu-Serie, M.M.; Maher, A.M.; Habashy, N.H. Inhibition of oxidative stress, IL-13, and WNT/β-catenin in ovalbumin-sensitized rats by a novel organogel of Punica granatum seed oil saponifiable fraction. Biomed. Pharmacother. 2022, 154, 113667. [Google Scholar] [CrossRef] [PubMed]
- Lal, R.; Dharavath, R.N.; Chopra, K. Alpha-Lipoic Acid Ameliorates Doxorubicin-Induced Cognitive Impairments by Modulating Neuroinflammation and Oxidative Stress via NRF-2/HO-1 Signaling Pathway in the Rat Hippocampus. Neurochem. Res. 2023, 48, 2476–2489. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; Zhou, Y.; Huang, Z.; Wang, M.; Jiang, J.; Yan, M.; Xiang, W.; Li, S.; Yu, Y.; Chen, L.; et al. Notopterol improves cognitive dysfunction and depression-like behavior via inhibiting STAT3/NF-ĸB pathway mediated inflammation in glioma-bearing mice. Int. Immunopharmacol. 2023, 118, 110041. [Google Scholar] [CrossRef] [PubMed]
- Parekh, S.V.; Adams, L.O.; Barkell, G.A.; Lysle, D.T. MDMA administration attenuates hippocampal IL-β immunoreactivity and subsequent stress-enhanced fear learning: An animal model of PTSD. Brain Behav. Immun. Health 2022, 26, 100542. [Google Scholar] [CrossRef]
- Murdaca, G.; Spanò, F.; Cagnati, P.; Puppo, F. Free radicals and endothelial dysfunction: Potential positive effects of TNF-α inhibitors. Redox. Rep. 2013, 18, 95–99. [Google Scholar] [CrossRef]
- Sherwood, A.; Hinderliter, A.L.; Watkins, L.L.; Waugh, R.A.; Blumenthal, J.A. Impaired endothelial function in coronary heart disease patients with depressive symptomatology. J. Am. Coll. Cardiol. 2005, 46, 656–659. [Google Scholar] [CrossRef] [Green Version]
- Rajendran, P.; Rengarajan, T.; Thangavel, J.; Nishigaki, Y.; Sakthisekaran, D.; Sethi, G.; Nishigaki, I. The vascular endothelium and human diseases. Int. J. Biol. Sci. 2013, 9, 1057–1069. [Google Scholar] [CrossRef] [Green Version]
- Cooper, D.C.; Tomfohr, L.M.; Milic, M.S.; Natarajan, L.; Bardwell, W.A.; Ziegler, M.G.; Dimsdale, J.E. Depressed mood and flow-mediated dilation: A systematic review and meta-analysis. Psychosom. Med. 2011, 73, 360–369. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sum of Squares | df | Mean Square | F | Overall p Value | ||
|---|---|---|---|---|---|---|
| MDA | Between Groups | 31,911.434 | 2 | 15,955.717 | 73.322 | <0.001 |
| Within Groups | 3264.182 | 15 | 217.612 | |||
| Total | 35,175.616 | 17 | ||||
| SOD | Between Groups | 151.919 | 2 | 75.959 | 65.939 | <0.001 |
| Within Groups | 17.280 | 15 | 1.152 | |||
| Total | 169.198 | 17 | ||||
| IL-1 B | Between Groups | 44,693.818 | 2 | 22,346.909 | 247.961 | <0.001 |
| Within Groups | 1351.840 | 15 | 90.123 | |||
| Total | 46,045.658 | 17 | ||||
| Noradrenaline | Between Groups | 17,965.671 | 2 | 8982.836 | 97.998 | <0.001 |
| Within Groups | 1374.947 | 15 | 91.663 | |||
| Total | 19,340.618 | 17 | ||||
| Cortisol | Between Groups | 199.564 | 2 | 99.782 | 24.530 | <0.001 |
| Within Groups | 61.016 | 15 | 4.068 | |||
| Total | 260.580 | 17 | ||||
| BDNF | Between Groups | 123,369.854 | 2 | 61,684.927 | 111.713 | <0.001 |
| Within Groups | 8282.623 | 15 | 552.175 | |||
| Total | 131,652.478 | 17 | ||||
| No of line crossings (open field test) | Between Groups | 3164.778 | 2 | 1582.389 | 52.109 | <0.001 |
| Within Groups | 455.500 | 15 | 30.367 | |||
| Total | 3620.278 | 17 | ||||
| Time spent in open arms (S) (elevated plus maze test) | Between Groups | 2362.333 | 2 | 1181.167 | 155.190 | <0.001 |
| Within Groups | 114.167 | 15 | 7.611 | |||
| Total | 2476.500 | 17 | ||||
| Head dipping (elevated plus maze test) | Between Groups | 296.333 | 2 | 148.167 | 43.016 | <0.001 |
| Within Groups | 51.667 | 15 | 3.444 | |||
| Total | 348.000 | 17 | ||||
| Stretching (elevated plus maze test) | Between Groups | 50.333 | 2 | 25.167 | 31.027 | <0.001 |
| Within Groups | 12.167 | 15 | 0.811 | |||
| Total | 62.500 | 17 | ||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alamri, H.S.; Mufti, R.; Sabir, D.K.; Abuderman, A.A.; Dawood, A.F.; ShamsEldeen, A.M.; Haidara, M.A.; Isenovic, E.R.; El-Bidawy, M.H. Forced Swimming-Induced Depressive-like Behavior and Anxiety Are Reduced by Chlorpheniramine via Suppression of Oxidative and Inflammatory Mediators and Activating the Nrf2-BDNF Signaling Pathway. Curr. Issues Mol. Biol. 2023, 45, 6449-6465. https://doi.org/10.3390/cimb45080407
Alamri HS, Mufti R, Sabir DK, Abuderman AA, Dawood AF, ShamsEldeen AM, Haidara MA, Isenovic ER, El-Bidawy MH. Forced Swimming-Induced Depressive-like Behavior and Anxiety Are Reduced by Chlorpheniramine via Suppression of Oxidative and Inflammatory Mediators and Activating the Nrf2-BDNF Signaling Pathway. Current Issues in Molecular Biology. 2023; 45(8):6449-6465. https://doi.org/10.3390/cimb45080407
Chicago/Turabian StyleAlamri, Hasan S., Rana Mufti, Deema Kamal Sabir, Abdulwahab A. Abuderman, Amal F. Dawood, Asmaa M. ShamsEldeen, Mohamed A. Haidara, Esma R. Isenovic, and Mahmoud H. El-Bidawy. 2023. "Forced Swimming-Induced Depressive-like Behavior and Anxiety Are Reduced by Chlorpheniramine via Suppression of Oxidative and Inflammatory Mediators and Activating the Nrf2-BDNF Signaling Pathway" Current Issues in Molecular Biology 45, no. 8: 6449-6465. https://doi.org/10.3390/cimb45080407